
Structural Protein Analysis of Driver Gene Mutations in Conjunctival Melanoma

 ,
,  , and
, and
Abstract
:1. Introduction
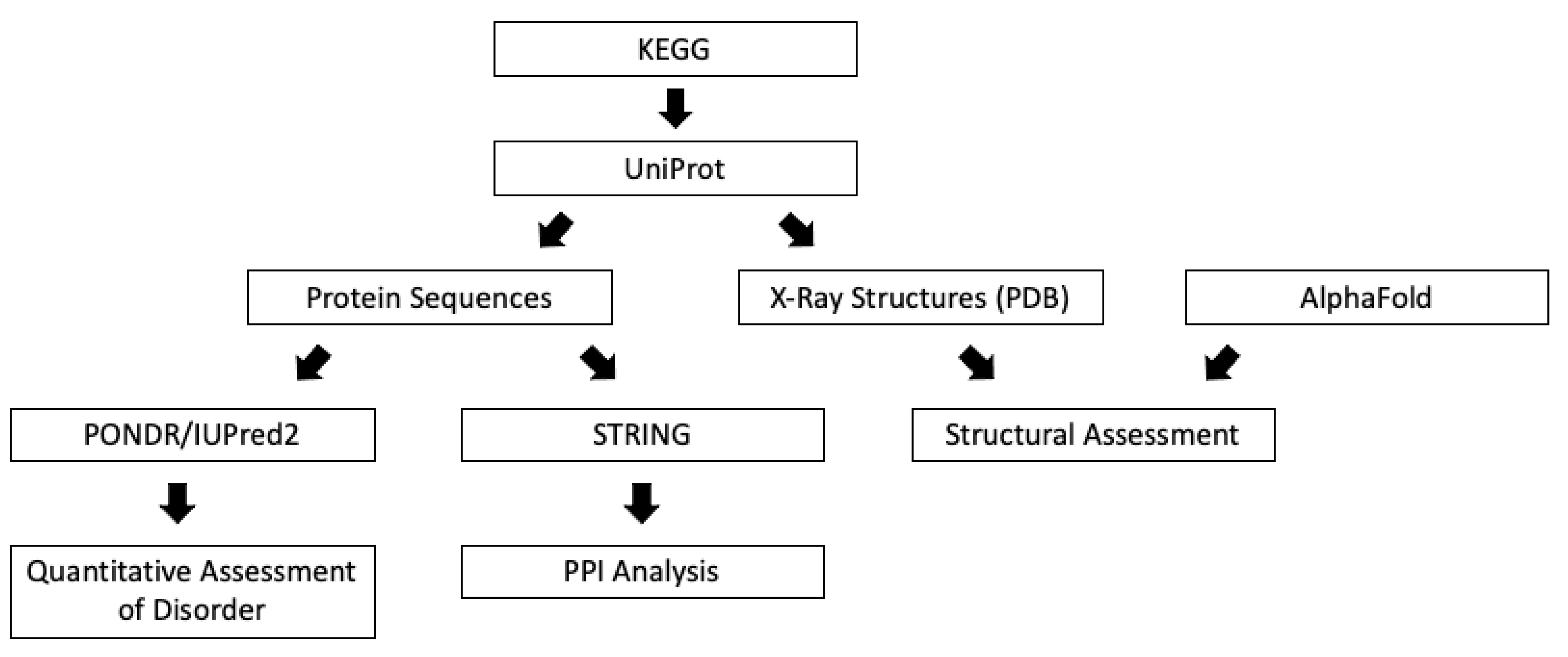
2. Materials and Methods
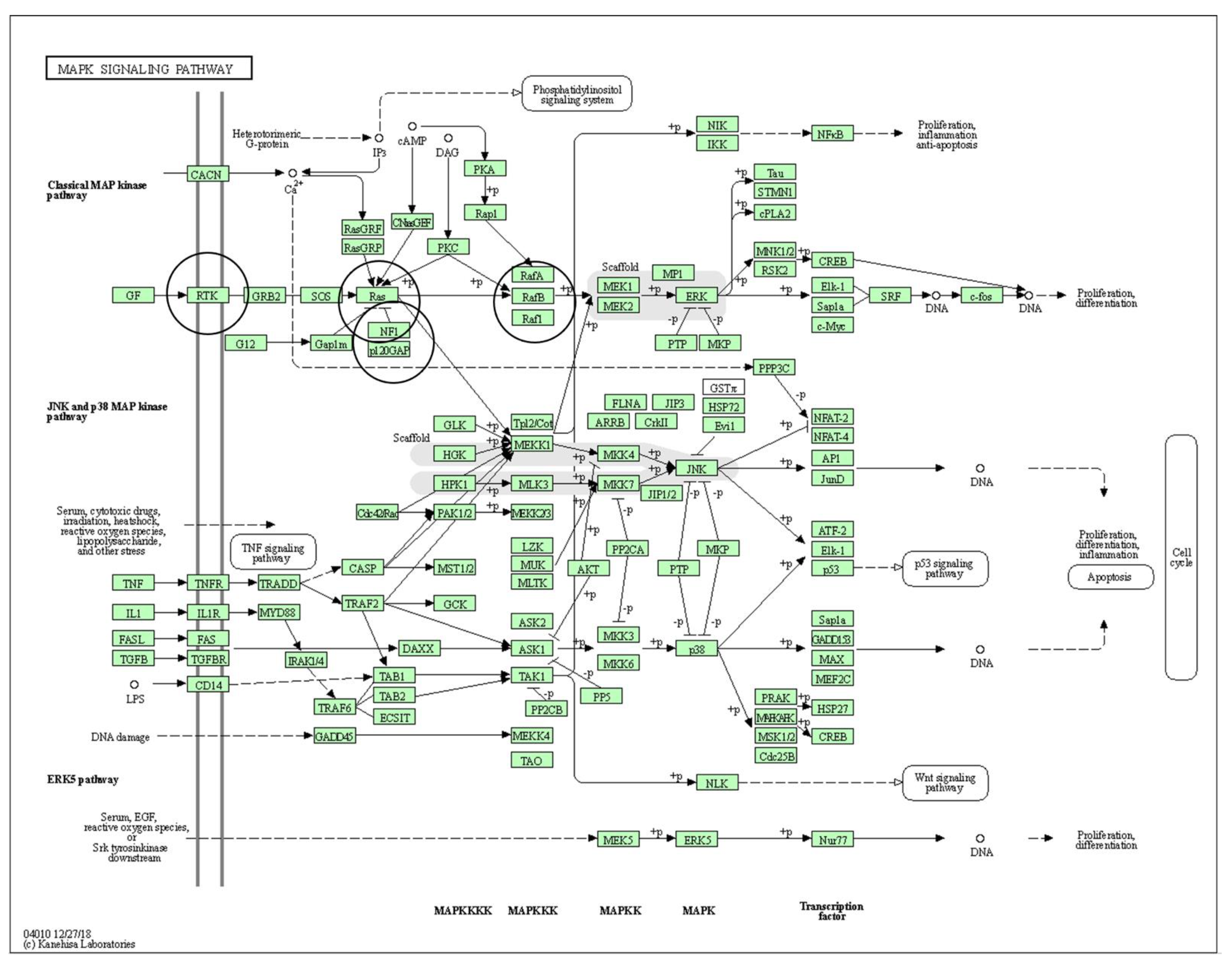
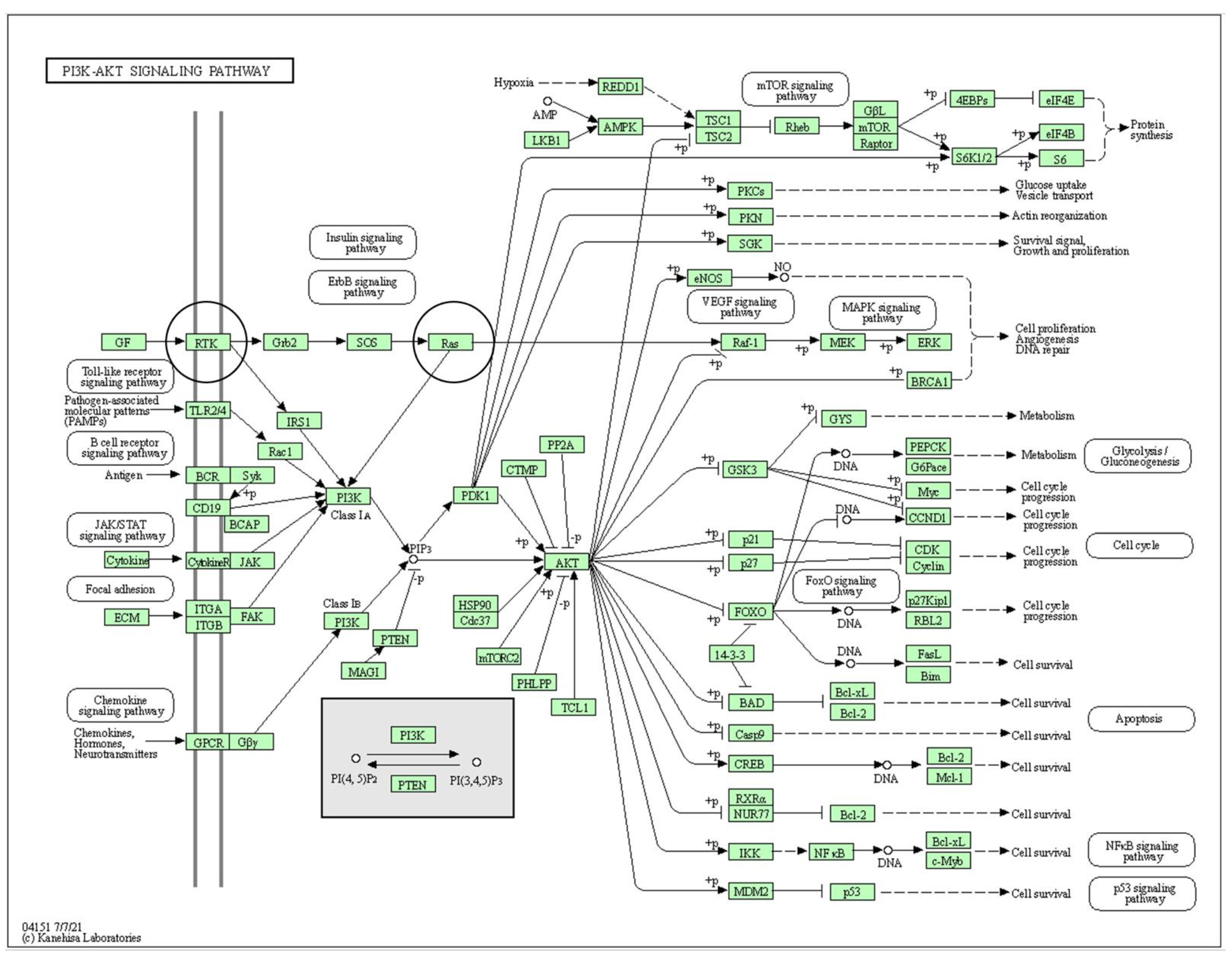
2.1. Pathways of Interest
2.2. Protein Sequences
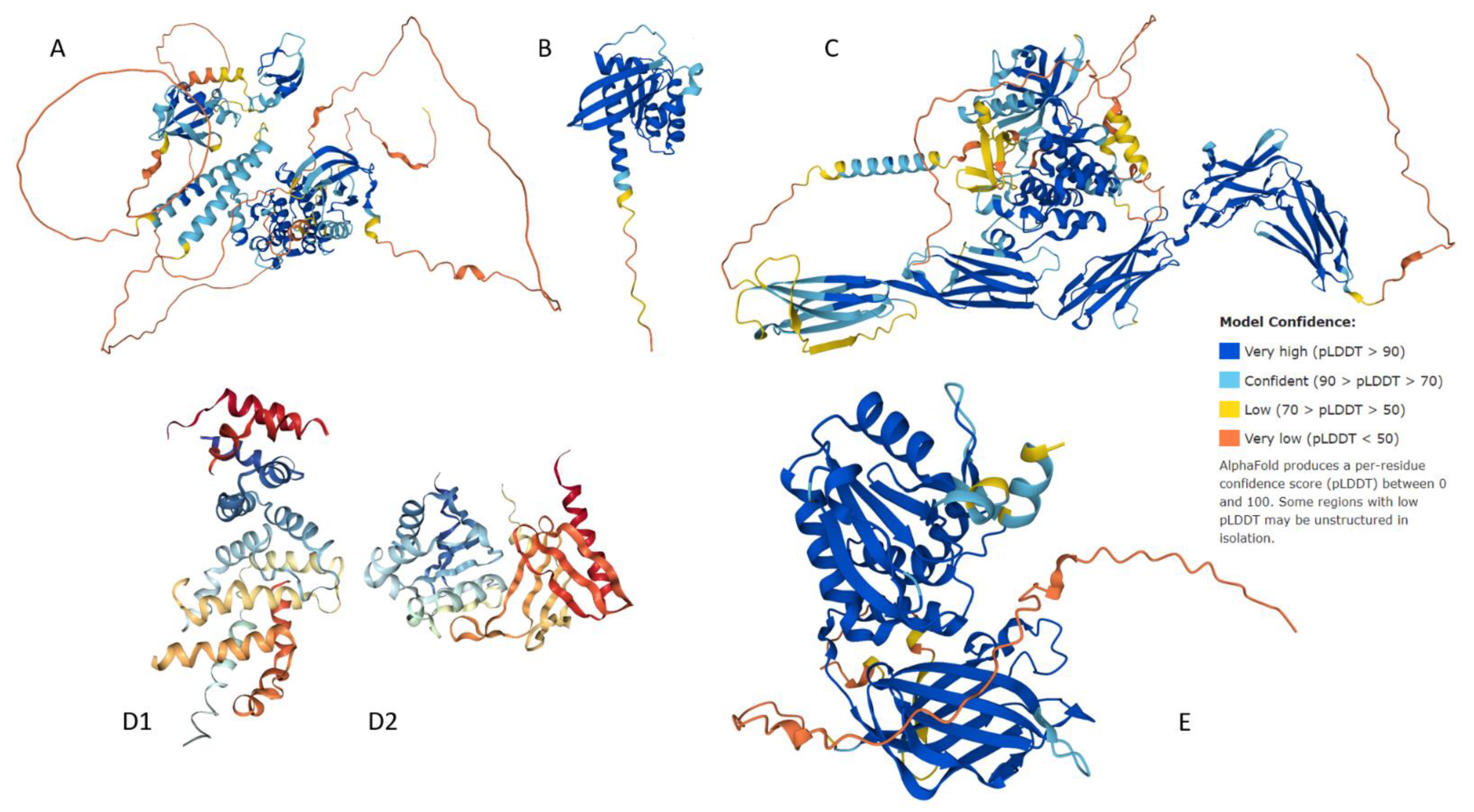
2.3. Structural Assessment
2.4. Quantitative Disorder-Based Predictions
2.5. Protein-Protein Interaction Network
3. Results
3.1. Pathways with Proteins of Interest
3.2. Protein Sequences
3.3. Structural Assessment
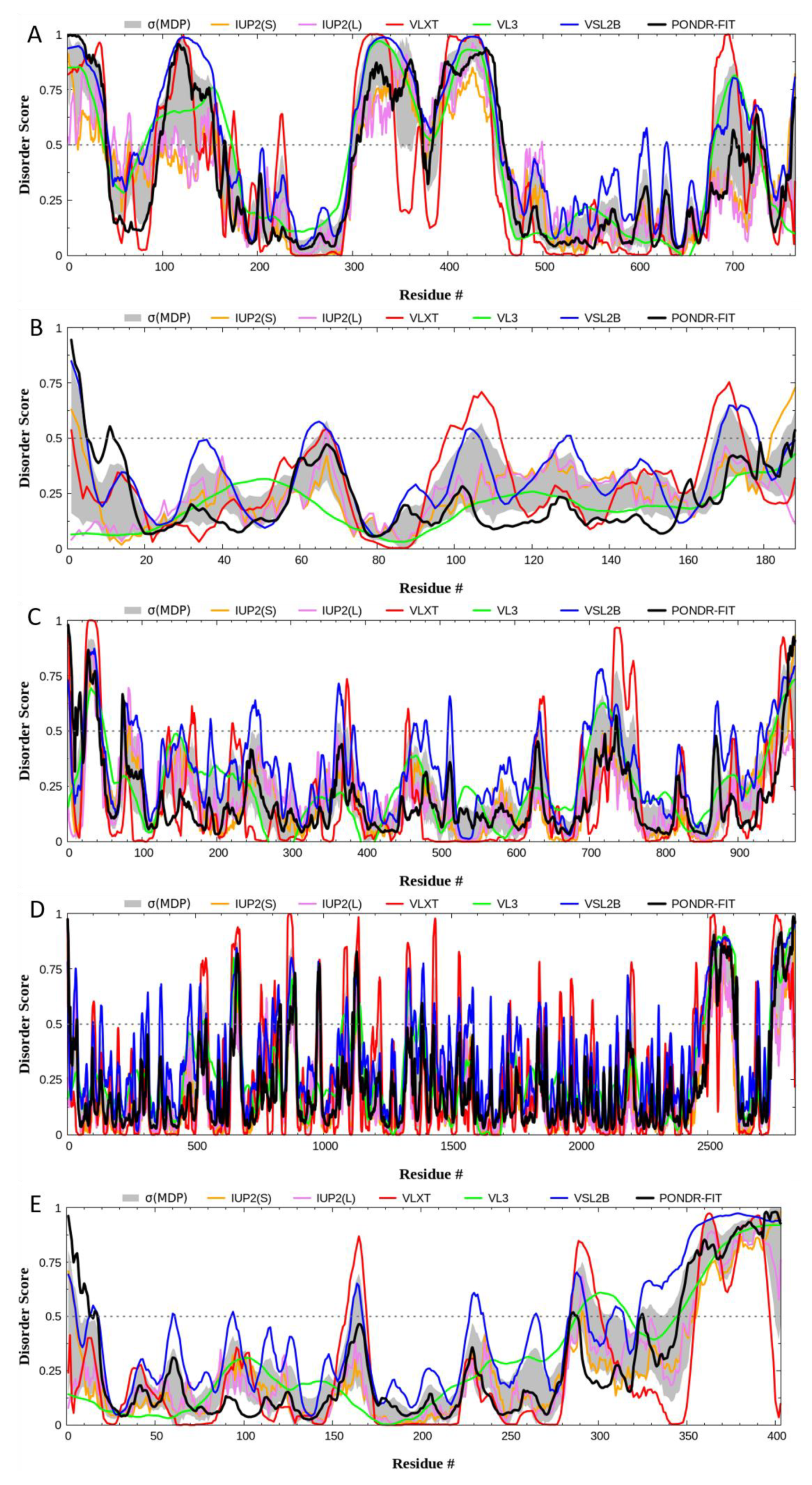
3.4. Quantitative Disorder Based Predictions
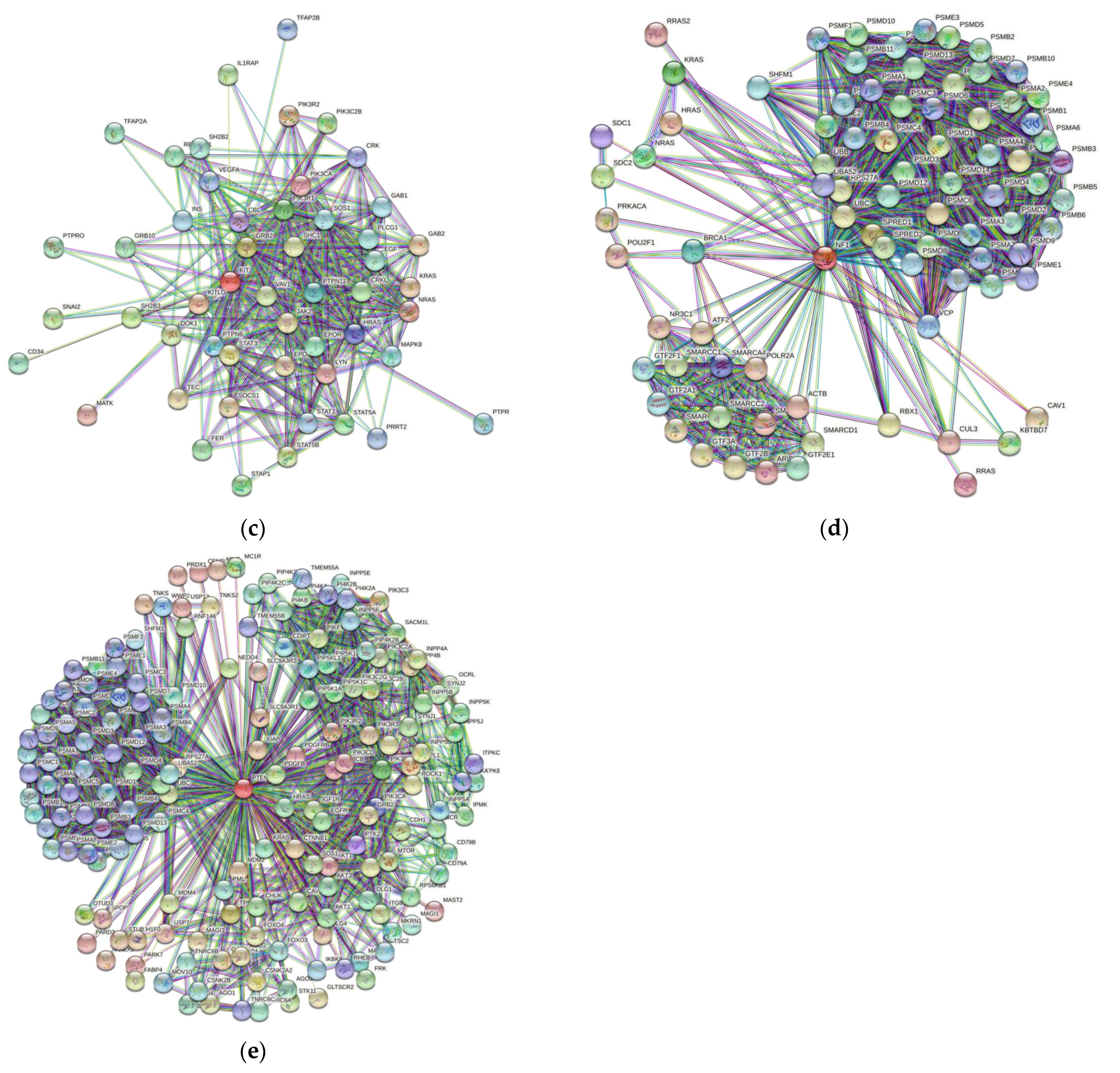
3.5. Protein-Protein Interaction Network
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Vora, G.K.; Demirci, H.; Marr, B.; Mruthyunjaya, P. Advances in the management of conjunctival melanoma. Surv. Ophthalmol. 2017, 62, 26–42. [Google Scholar] [CrossRef] [Green Version]
- Grimes, J.; Shah, N.V.; Samie, F.H.; Carvajal, R.D.; Marr, B.P. Conjunctival Melanoma: Current Treatments and Future Options. Am. J. Clin. Dermatol. 2020, 21, 371–381. [Google Scholar] [CrossRef]
- Mor, J.M.; Rokohl, A.C.; Koch, K.R.; Heindl, L.M. Sentinel lymph node biopsy in the management of conjunctival melanoma: Current insights. Clin. Ophthalmol. 2019, 13, 1297–1302. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.; Nanji, A.A.; Galor, A.; Karp, C.L. Management of conjunctival malignant melanoma: A review and update. Expert Rev. Ophthalmol. 2014, 9, 185–204. [Google Scholar] [CrossRef] [Green Version]
- Shields, C.L.; Chien, J.L.; Surakiatchanukul, T.; Sioufi, K.; Lally, S.E.; Shields, J.A. Conjunctival Tumors: Review of Clinical Features, Risks, Biomarkers, and Outcomes—The 2017 J. Donald M. Gass Lecture. Asia-Pac. J. Ophthalmol. 2017, 6, 109–120. [Google Scholar] [CrossRef]
- Jain, P.; Finger, P.T.; Fili, M.; Damato, B.; Coupland, S.E.; Heimann, H.; Kenawy, N.; Brouwer, N.J.; Marinkovic, M.; Van Duinen, S.G.; et al. Conjunctival melanoma treatment outcomes in 288 patients: A multicentre international data-sharing study. Br. J. Ophthalmol. 2020, 105, 1358–1364. [Google Scholar] [CrossRef]
- Koç, I.; Kıratlı, H. Current Management of Conjunctival Melanoma Part 1: Clinical Features, Diagnosis and Histopathology. Turk. J. Ophthalmol. 2020, 50, 293–303. [Google Scholar] [CrossRef]
- Benage, M.J.; Morrow, N.C.; Janson, B.J.; Greiner, M.A. Evaluation of interferon α 2b as adjunctive therapy for conjunctival melanoma. Am. J. Ophthalmol. Case Rep. 2019, 15, 100467. [Google Scholar] [CrossRef]
- Mazzini, C.; Pieretti, G.; Vicini, G.; Nicolosi, C.; Virgili, G.; Giansanti, F. Extensive conjunctival melanoma successfully treated with surgical resection and pre- and postoperative topical mitomycin C. Eur. J. Ophthalmol. 2020. [Google Scholar] [CrossRef]
- Yin, V.T.; Merritt, H.A.; Sniegowski, M.; Esmaeli, B. Eyelid and ocular surface carcinoma: Diagnosis and management. Clin. Dermatol. 2015, 33, 159–169. [Google Scholar] [CrossRef]
- Kikuchi, I.; Kase, S.; Ishijima, K.; Ishida, S. Long-term follow-up of conjunctival melanoma treated with topical interferon α-2b eye drops as adjunctive therapy following surgical resection. Graefe’s Arch. Clin. Exp. Ophthalmol. 2017, 255, 2271–2276. [Google Scholar] [CrossRef]
- Garip, A.; Schaumberger, M.M.; Wolf, A.; Herold, T.R.; Miller, C.V.; Klingenstein, A.; Schebitz-Walter, K.; Hintschich, C.R. Evaluation of a short-term topical interferon α-2b treatment for histologically proven melanoma and primary acquired melanosis with atypia. Orbit 2016, 35, 29–34. [Google Scholar] [CrossRef]
- Finger, P.T.; Sedeek, R.W.; Chin, K.J. Topical Interferon Alfa in the Treatment of Conjunctival Melanoma and Primary Acquired Melanosis Complex. Am. J. Ophthalmol. 2008, 145, 124–129.e1. [Google Scholar] [CrossRef]
- Finger, P.T.; Czechonska, G.; Liarikos, S. Topical mitomycin C chemotherapy for conjunctival melanoma and PAM with atypia. Br. J. Ophthalmol. 1998, 82, 476–479. [Google Scholar] [CrossRef] [Green Version]
- Demirci, H.; McCormick, S.A.; Finger, P.T. Topical mitomycin chemotherapy for conjunctival malignant melanoma and primary acquired melanosis with atypia: Clinical experience with histopathologic observations. Arch. Ophthalmol. 2000, 118, 885–891. [Google Scholar]
- Kim, J.W.; Abramson, D.H. Topical treatment options for conjunctival neoplasms. Clin. Ophthalmol. 2008, 2, 503–515. [Google Scholar] [CrossRef] [Green Version]
- Kase, S.; Ishijima, K.; Noda, M.; Ishida, S. [Two cases of conjunctival malignant melanoma treated with topical interferon α-2b drop as an adjuvant therapy]. Nippon. Ganka Gakkai Zasshi 2011, 115, 1043–1047. [Google Scholar]
- Shields, C.L.; Markowitz, J.S.; Belinsky, I.; Schwartzstein, H.; George, N.S.; Lally, S.E.; Mashayekhi, A.; Shields, J.A. Conjunctival Melanoma: Outcomes Based on Tumor Origin in 382 Consecutive Cases. Ophthalmology 2011, 118, 389–395.e2. [Google Scholar] [CrossRef]
- Wierenga, A.P.; Cao, J.; Luyten, G.P.; Jager, M.J. Immune Checkpoint Inhibitors in Uveal and Conjunctival Melanoma. Int. Ophthalmol. Clin. 2019, 59, 53–63. [Google Scholar] [CrossRef]
- Rossi, E.; Maiorano, B.A.; Pagliara, M.M.; Sammarco, M.G.; Dosa, T.; Martini, M.; Rindi, G.; Bria, E.; Blasi, M.A.; Tortora, G.; et al. Dabrafenib and Trametinib in BRAF Mutant Metastatic Conjunctival Melanoma. Front. Oncol. 2019, 9, 232. [Google Scholar] [CrossRef] [Green Version]
- Demirci, H.; Demirci, F.Y.; Ciftci, S.; Elner, V.; Wu, Y.-M.; Ning, Y.; Chinnaiyan, A.; Robinson, D.R. Integrative Exome and Transcriptome Analysis of Conjunctival Melanoma and Its Potential Application for Personalized Therapy. JAMA Ophthalmol. 2019, 137, 1444–1448. [Google Scholar] [CrossRef]
- Sun, J.; Carr, M.J.; Khushalani, N.I. Principles of Targeted Therapy for Melanoma. Surg. Clin. N. Am. 2020, 100, 175–188. [Google Scholar] [CrossRef]
- Gkiala, A.; Palioura, S. Conjunctival Melanoma: Update on Genetics, Epigenetics and Targeted Molecular and Immune-Based Therapies. Clin. Ophthalmol. 2020, 14, 3137–3152. [Google Scholar] [CrossRef]
- Uversky, V.N. A decade and a half of protein intrinsic disorder: Biology still waits for physics. Protein Sci. 2013, 22, 693–724. [Google Scholar] [CrossRef] [Green Version]
- Daughdrill, G.W.; Pielak, G.J.; Uversky, V.N.; Cortese, M.S.; Dunker, A.K. Natively disordered proteins. In Protein Folding Handbook; Wiley-VCH: Weinheim, Germany, 2005; pp. 275–357. [Google Scholar]
- Uversky, V.N. Unusual biophysics of intrinsically disordered proteins. Biochim. Biophys. Acta—Proteins Proteom. 2013, 1834, 932–951. [Google Scholar] [CrossRef]
- Santofimia-Castaño, P.; Rizzuti, B.; Xia, Y.; Abian, O.; Peng, L.; Velazquez-Campoy, A.; Neira, J.L.; Iovanna, J. Targeting intrinsically disordered proteins involved in cancer. Cell. Mol. Life Sci. 2020, 77, 1695–1707. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 27, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nat. Cell Biol. 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Romero, P.; Obradovic, Z.; Li, X.; Garner, E.C.; Brown, C.J.; Dunker, A.K. Sequence complexity of disordered protein. Proteins Struct. Funct. Bioinform. 2001, 42, 38–48. [Google Scholar] [CrossRef]
- Peng, K.; Vucetic, S.; Radivojac, P.; Brown, C.J.; Dunker, A.K.; Obradovic, Z. Optimizing long intrinsic disorder predictors with protein evolutionary information. J. Bioinform. Comput. Biol. 2005, 3, 35–60. [Google Scholar] [CrossRef]
- Xue, B.; Dunbrack, R.; Williams, R.W.; Dunker, A.K.; Uversky, V.N. PONDR-FIT: A meta-predictor of intrinsically disordered amino acids. Biochim. Biophys. Acta—Proteins Proteom. 2010, 1804, 996–1010. [Google Scholar] [CrossRef] [Green Version]
- Mészáros, B.; Erdős, G.; Dosztányi, Z. IUPred2A: Context-dependent prediction of protein disorder as a function of redox state and protein binding. Nucleic Acids Res. 2018, 46, W329–W337. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y.; Furumichi, M.; Morishima, K.; Tanabe, M. New approach for understanding genome variations in KEGG. Nucleic Acids Res. 2019, 47, D590–D595. [Google Scholar] [CrossRef] [Green Version]
- Rajagopalan, K.; Mooney, S.M.; Parekh, N.; Getzenberg, R.H.; Kulkarni, P. A majority of the cancer/testis antigens are intrinsically disordered proteins. J. Cell. Biochem. 2011, 112, 3256–3267. [Google Scholar] [CrossRef] [Green Version]
- Nussinov, R.; Jang, H.; Tsai, C.-J.; Liao, T.-J.; Li, S.; Fushman, D.; Zhang, J. Intrinsic protein disorder in oncogenic KRAS signaling. Cell. Mol. Life Sci. 2017, 74, 3245–3261. [Google Scholar] [CrossRef]
- Biesaga, M.; Frigolé-Vivas, M.; Salvatella, X. Intrinsically disordered proteins and biomolecular condensates as drug targets. Curr. Opin. Chem. Biol. 2021, 62, 90–100. [Google Scholar] [CrossRef]
- Spriano, F.; Chung, E.Y.L.; Gaudio, E.; Tarantelli, C.; Cascione, L.; Napoli, S.; Jessen, K.; Carrassa, L.; Priebe, V.; Sartori, G.; et al. The ETS Inhibitors YK-4-279 and TK-216 Are Novel Antilymphoma Agents. Clin. Cancer Res. 2019, 25, 5167–5176. [Google Scholar] [CrossRef]
- Sadar, M.D. Discovery of drugs that directly target the intrinsically disordered region of the androgen receptor. Expert Opin. Drug Discov. 2020, 15, 551–560. [Google Scholar] [CrossRef]
- Neira, J.L.; Bintz, J.; Arruebo, M.; Rizzuti, B.; Bonacci, T.; Vega, S.; Lanas, A.; Velázquez-Campoy, A.; Iovanna, J.L.; Abián, O. Identification of a Drug Targeting an Intrinsically Disordered Protein Involved in Pancreatic Adenocarcinoma. Sci. Rep. 2017, 7, 39732. [Google Scholar] [CrossRef]
- Quaglino, P.; Fava, P.; Tonella, L.; Rubatto, M.; Ribero, S.; Ferro, M.T. Treatment of Advanced Metastatic Melanoma. Dermatol. Pr. Concept. 2021, 11 (Suppl. 1), 2021164S. [Google Scholar] [CrossRef]
- Wu, B.; Shi, L. Frontline BRAF Testing-Guided Treatment for Advanced Melanoma in the Era of Immunotherapies: A Cost-Utility Analysis Based on Long-term Survival Data. JAMA Derm. 2020, 156, 1177–1184. [Google Scholar] [CrossRef]
- Sabbah, M.; Najem, A.; Krayem, M.; Awada, A.; Journe, F.; Ghanem, G. RTK Inhibitors in Melanoma: From Bench to Bedside. Cancers 2021, 13, 1685. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Known Gene Mutations | UniProt ID | Structure Source |
|---|---|---|---|
| BRAF | G469A, D594G, V600E/K/R | P15056 | AlphaFold2 |
| NRAS | Q12C, Q13D, Q61R/H/L/K | P01111 | AlphaFold2 |
| c-KIT | * | P10721 | AlphaFold2 |
| NF1 | More than 25 distinct mutations | P21359 | 1NF1and 3PEG (PDB IDs) |
| PTEN | Chromosome 10q deletion | P60484 | AlphaFold2 |
| Gene Name | |||||
|---|---|---|---|---|---|
| Predictor | BRAF | NRAS | c-KIT | NF1 | PTEN |
| PONDR® VLXT | 33.55% | 14.29% | 12.30% | 19.90% | 17.62% |
| PONDR® VL3 | 43.08% | 0.00% | 9.53% | 13.46% | 22.08% |
| PONDR® VSL2 | 45.95% | 14.81% | 21.82% | 22.19% | 31.76% |
| Average | 40.86% | 9.70% | 14.55% | 18.52% | 23.82% |
| Gene Name | Proteins in Network | Expected Number of Interactions | Predicted Number of Interactions | p-Value |
|---|---|---|---|---|
| BRAF | 109 | 253 | 2213 | <10−16 |
| NRAS | 327 | 1619 | 7795 | <10−16 |
| c-KIT | 60 | 177 | 495 | <10−16 |
| NF1 | 90 | 388 | 1790 | <10−16 |
| PTEN | 157 | 622 | 2297 | <10−16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djulbegovic, M.B.; Uversky, V.N.; Harbour, J.W.; Galor, A.; Karp, C.L. Structural Protein Analysis of Driver Gene Mutations in Conjunctival Melanoma. Genes 2021, 12, 1625. https://doi.org/10.3390/genes12101625
Djulbegovic MB, Uversky VN, Harbour JW, Galor A, Karp CL. Structural Protein Analysis of Driver Gene Mutations in Conjunctival Melanoma. Genes. 2021; 12(10):1625. https://doi.org/10.3390/genes12101625
Chicago/Turabian StyleDjulbegovic, Mak B., Vladimir N. Uversky, J. William Harbour, Anat Galor, and Carol L. Karp. 2021. "Structural Protein Analysis of Driver Gene Mutations in Conjunctival Melanoma" Genes 12, no. 10: 1625. https://doi.org/10.3390/genes12101625
APA StyleDjulbegovic, M. B., Uversky, V. N., Harbour, J. W., Galor, A., & Karp, C. L. (2021). Structural Protein Analysis of Driver Gene Mutations in Conjunctival Melanoma. Genes, 12(10), 1625. https://doi.org/10.3390/genes12101625