
Molecular Aspects of Regeneration Mechanisms in Holothurians


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Regeneration after Fission
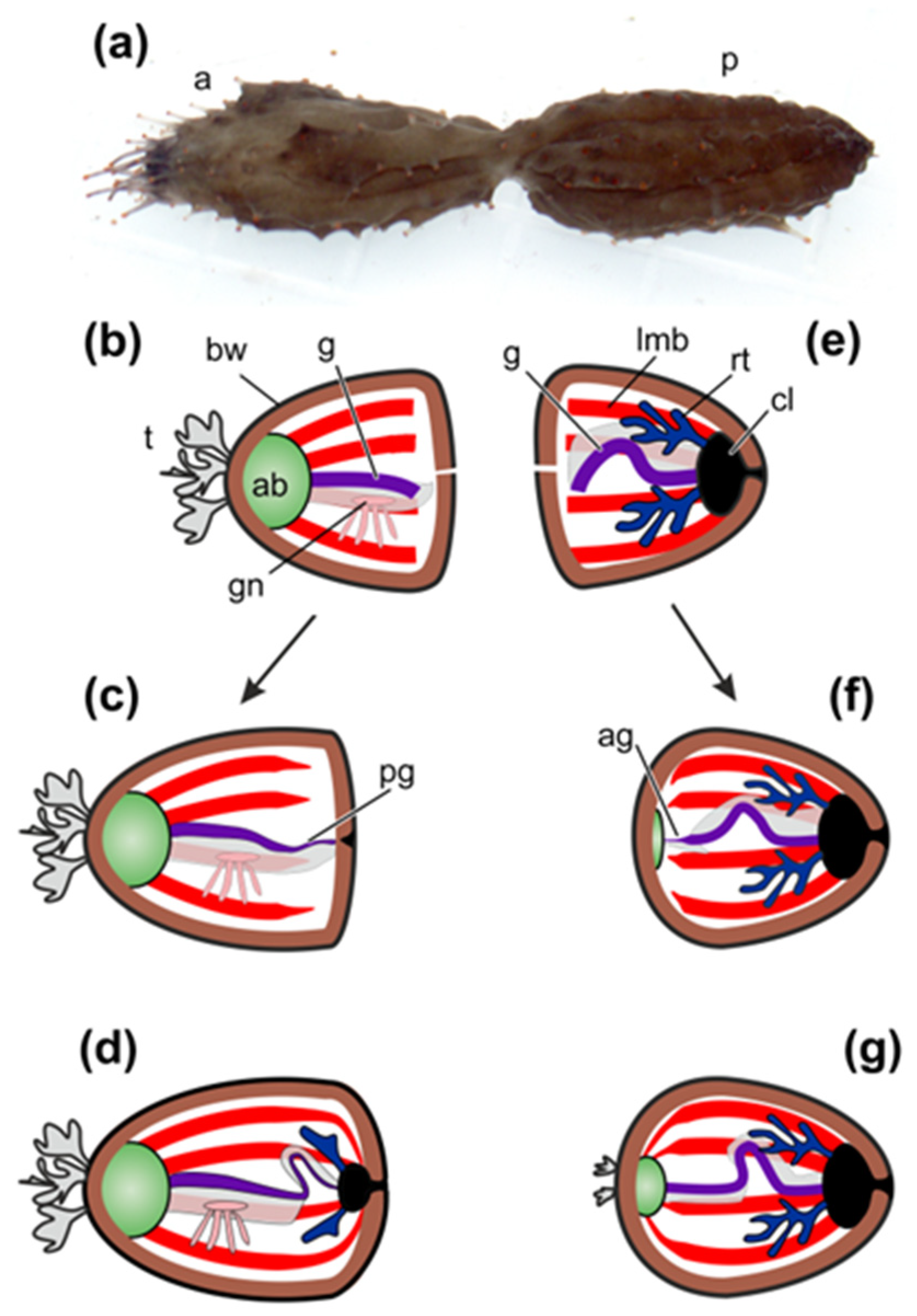
2.1. Morphological Aspects of Fission and Regeneration
2.2. Structural Components of Connective Tissue
2.3. Proteins Modifying ECM
2.4. Fission Mechanisms
2.5. Preparation for Whole Body Regeneration
3. Early Events after Evisceration
4. Regeneration after Posterior Evisceration
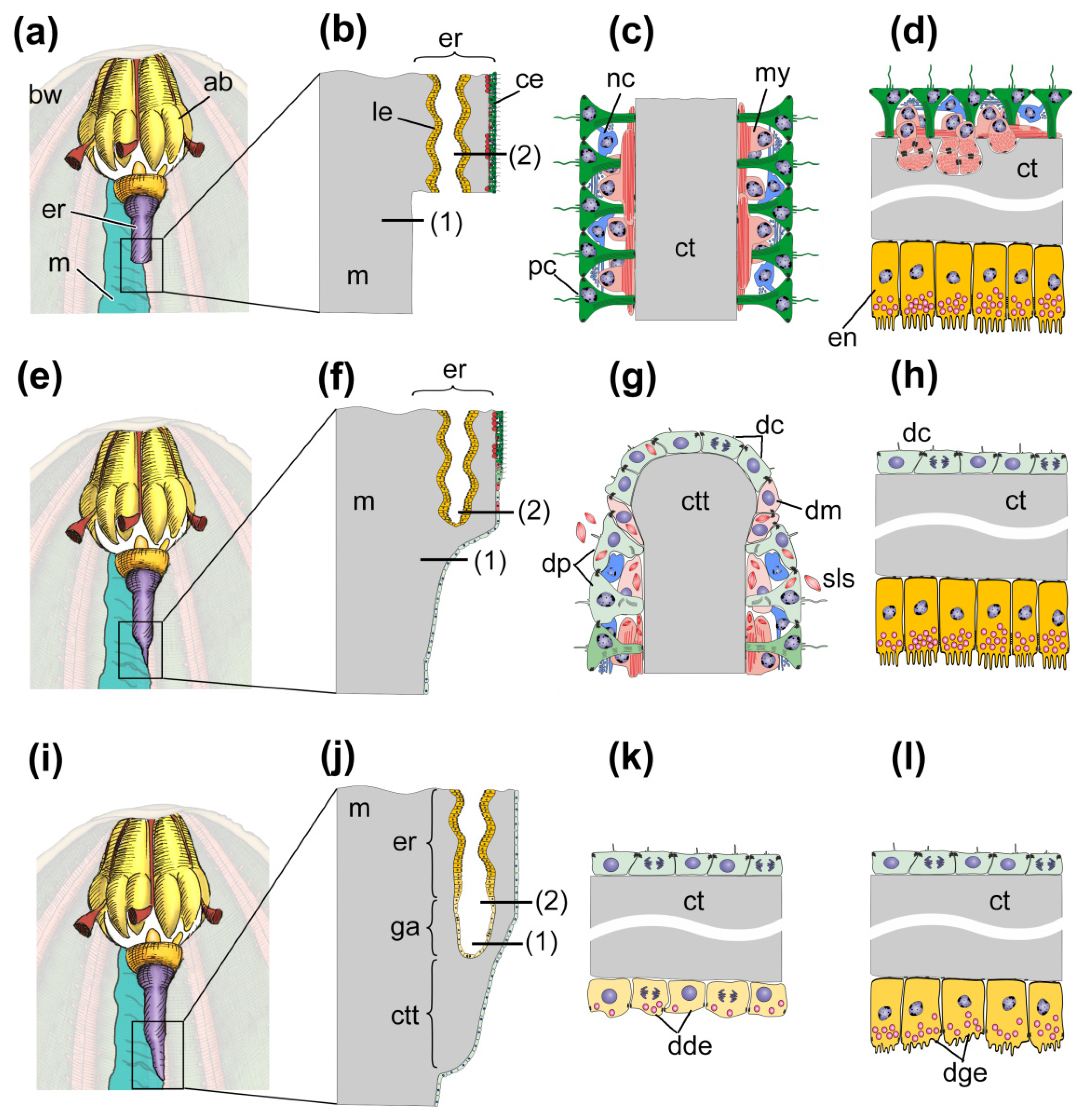
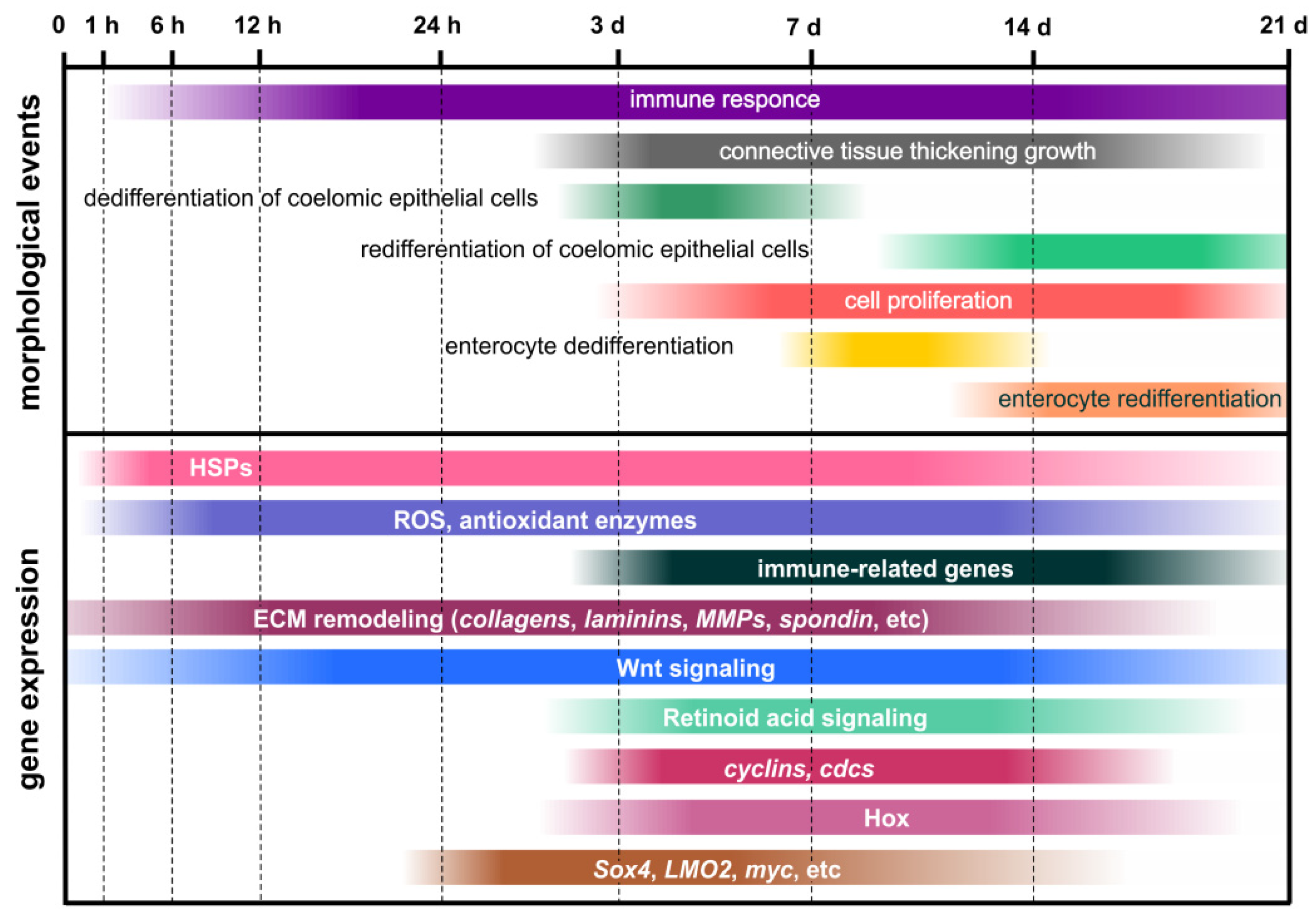
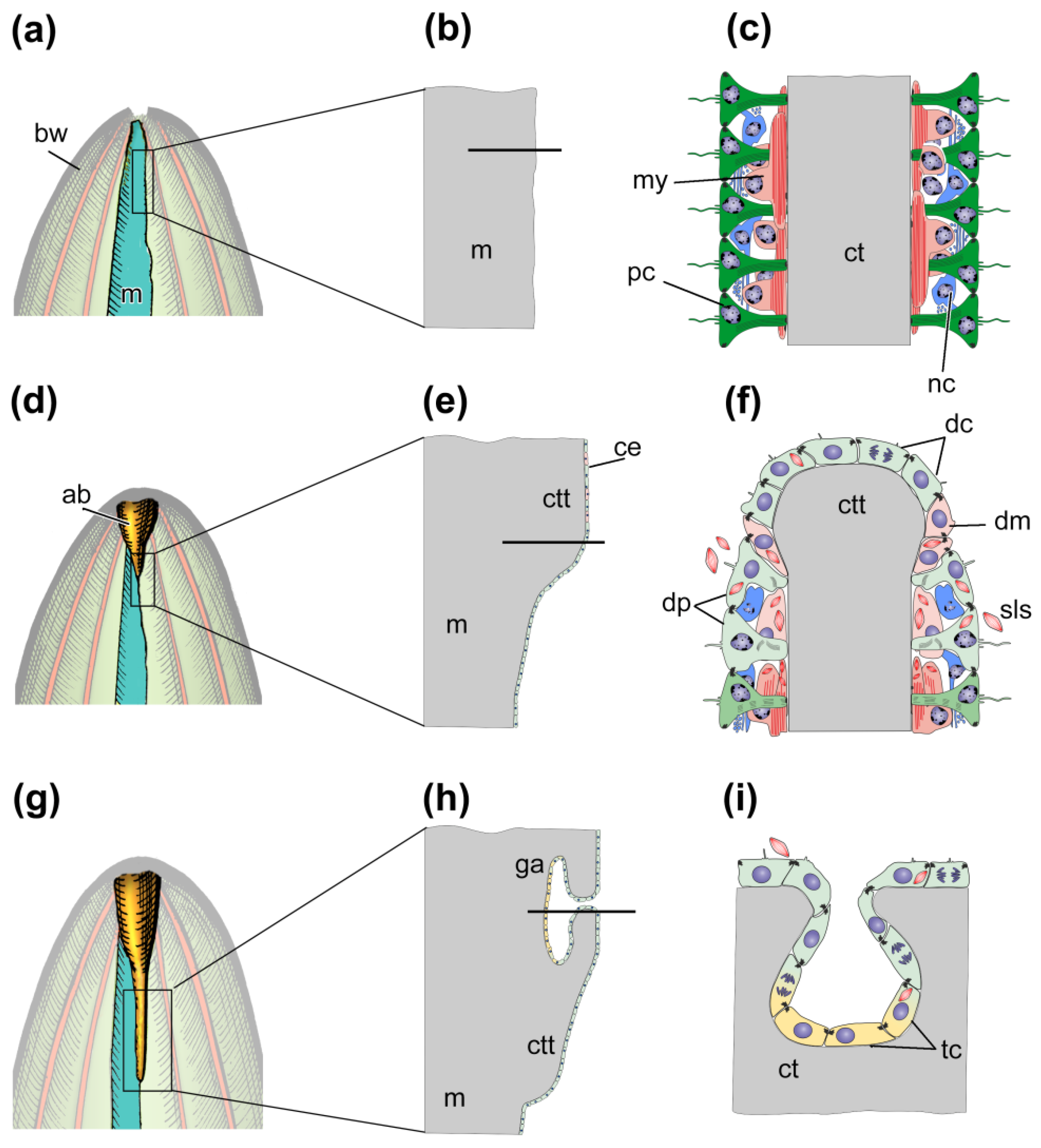
4.1. Morphological Aspects of Regeneration
4.1.1. Initial Stage (1–24 h Post-Evisceration)
4.1.2. Preparation to Regeneration (1–3 Days Post-Evisceration, dpe)
4.1.3. Onset of Regeneration (7–10 dpe)
4.1.4. Regeneration and Growth of Gut (10–21 dpe)
4.2. Transcriptomic, Proteomic and Metabolomic Data
4.2.1. 1–3 Days Post-Evisceration
4.2.2. 7 Days Post-Evisceration
4.2.3. 14–21 Days Post-Evisceration
4.3. Analysis of Expression of Single Genes
4.3.1. ECM Remodeling
4.3.2. Cell Proliferation and Apoptosis
4.3.3. Wnt Signaling Pathway
4.3.4. Retinoic Acid Signaling
5. Regeneration after Anterior Evisceration
5.1. Morphological Aspects of Regeneration
5.2. Transcriptomic Data
5.3. Wnt Signaling Pathway
5.4. Analysis of Expression of Single Genes
6. Nervous System Regeneration
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bluhm, H.; Gebruk, A. Holothuroidea (Echinodermata) of the Peru Basin—Ecological and taxonomic remarks based on underwater images. Mar. Ecol. 1999, 20, 167–195. [Google Scholar] [CrossRef]
- Solan, M.; Germano, J.D.; Rhoads, D.C.; Smith, C.; Michaud, E.; Parry, D.; Wenzhofer, F.; Kennedy, B.; Henriques, C.; Battle, E.; et al. Towards a greater understanding of pattern, scale and process in marine benthic systems: A picture is worth a thousand worms. J. Exp. Mar. Biol. Ecol. 2003, 285, 313–338. [Google Scholar] [CrossRef]
- Ohta, S. Photographic observations of the deep sea pelagothuriid holothurian Enypniastes (Elasipoda, Holothurioidea). J. Oceanogr. Soc. Jpn. 1985, 41, 121–133. [Google Scholar] [CrossRef]
- Conand, C. Population status, fisheries and trade of sea cucumbers in Africa and the Indian Ocean. In Sea Cucumbers. A Global Review of Fisheries and Trade. FAO Fisheries and Aquaculture Technical Paper. No. 516; Toral-Granda, V., Lovatelli, A., Vasconcellos, M., Eds.; FAO: Rome, Italy, 2008; pp. 143–193. [Google Scholar]
- Uthicke, S.; Byrne, M.; Conand, C. Genetic barcoding of commercial Bêche-de-mer species (Echinodermata: Holothuroidea). Mol. Ecol. Resour. 2010, 10, 634–646. [Google Scholar] [CrossRef]
- Candia Carnevali, M.D. Regeneration in echinoderms: Repair, regrowth, cloning. Invertebr. Surv. J. 2006, 3, 64–76. [Google Scholar]
- Dolmatov, I.Y. Regeneration in echinoderms. Russ. J. Mar. Biol. 1999, 25, 225–233. [Google Scholar]
- Dolmatov, I.Y. New data on asexual reproduction, autotomy, and regeneration in holothurians of the order Dendrochirotida. Russ. J. Mar. Biol. 2014, 40, 228–232. [Google Scholar] [CrossRef]
- Dolmatov, I.Y.; Ginanova, T.T. Muscle regeneration in holothurians. Microsc. Res. Tech. 2001, 55, 452–463. [Google Scholar] [CrossRef] [PubMed]
- Dolmatov, I.Y.; Ginanova, T.T. Post-autotomy regeneration of the respiratory trees in the holothurian Apostichopus japonicus (Holothurioidea, Aspidochirotida). Cell Tissue Res. 2009, 336, 41–58. [Google Scholar] [CrossRef]
- Kille, F.R. Regeneration of the reproductive system following binary fission in the sea cucumber Holothuria parvula. Biol. Bull. 1942, 83, 55–66. [Google Scholar] [CrossRef]
- Mashanov, V.S.; García-Arrarás, J.E. Gut regeneration in holothurians: A snapshot of recent developments. Biol. Bull. 2011, 221, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Hyman, L.H. The Invertebrates: Echinodermata. The Coelome Bilateria; McGraw-Hill Book Co.: New York, NY, USA, 1955; p. 763. [Google Scholar]
- Monticelli, F.S. Sull’ autotomia delle Cucumaria planci (Br.). Atti Accad. Naz. Lincei Cl. Sci. Fis. Mat. Nat. Rendi. 1896, 5, 231–239. [Google Scholar]
- Torelle, E. Regeneration in holothuria. Zool. Anz. 1910, 35, 15–22. [Google Scholar]
- Reichenbach, N.; Holloway, S. Potential for asexual propagation of several commercially important species of tropical sea cucumbers (Echinodermata). J. World Aquac. Soc. 1995, 26, 272–278. [Google Scholar] [CrossRef]
- Reichenbach, N.; Nishar, Y.; Saeed, A. Species and size-related trends in asexual propagation of commercially important species of tropical sea cucumbers (Holothuroidea). J. World Aquac. Soc. 1996, 27, 475–482. [Google Scholar] [CrossRef]
- Bertolini, F. Rigenerazione dell’apparato digerente nello Holothuria. Pubbl. Staz. Zool. Napoli 1932, 12, 432–443. [Google Scholar]
- Byrne, M. Evisceration behavior and the seasonal incidence of evisceration in the holothurian Eupentacta quinquesemita (Selenka). Ophelia 1985, 24, 75–90. [Google Scholar] [CrossRef]
- Swan, E.F. Seasonal Evisceration in the sea cucumber, Parastichopus californicus (Stimpson). Science 1961, 133, 1078–1079. [Google Scholar] [CrossRef]
- Emson, R.H.; Mladenov, P.V. Studies of the fissiparous holothurian Holothuria parvula (Selenka) (Echinodermata, Holothuroidea). J. Exp. Mar. Biol. Ecol. 1987, 111, 195–211. [Google Scholar] [CrossRef]
- Emson, R.H.; Wilkie, I.C. Fission and autotomy in echinoderms. Oceanogr. Mar. Biol. Annu. Rev. 1980, 18, 155–250. [Google Scholar]
- Dolmatov, I.Y.; Nguyen, A.K.; Kamenev, Y.O. Asexual reproduction, evisceration, and regeneration in holothurians (Holothuroidea) from Nha Trang Bay of the South China Sea. Russ. J. Mar. Biol. 2012, 38, 243–252. [Google Scholar] [CrossRef]
- Dolmatov, I.Y.; Mashanov, V.S. Regeneration in Holothurians; Dalnauka: Vladivostok, Russia, 2007; p. 212. [Google Scholar]
- Vogt, G. Hidden treasures in stem cells of indeterminately growing bilaterian invertebrates. Stem Cell Rev. Rep. 2012, 8, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Eliseikina, M.G.; Magarlamov, T.Y.; Dolmatov, I.Y. Stem cells of holothuroid coelomocytes. In Echinoderms: Durham; Harris, L.G., Bottger, S.A., Walker, C.W., Lesser, M.P., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 163–166. [Google Scholar]
- Zavalnaya, E.G.; Shamshurina, E.V.; Eliseikina, M.G. The Immunocytochemical Identification of PIWI-positive cells during the recovery of a coelomocyte population after evisceration in the holothurian Eupentacta fraudatrix (Djakonov et Baranova, 1958) (Holothuroidea: Dendrochirota). Russ. J. Mar. Biol. 2020, 46, 97–104. [Google Scholar] [CrossRef]
- Mashanov, V.S.; Zueva, O.R.; Garcia-Arraras, J.E. Expression of pluripotency factors in echinoderm regeneration. Cell Tissue Res. 2015, 359, 521–536. [Google Scholar] [CrossRef] [Green Version]
- Mashanov, V.; Zueva, O.; Mashanova, D.; García-Arrarás, J.E. Expression of stem cell factors in the adult sea cucumber digestive tube. Cell Tissue Res. 2017, 370, 427–440. [Google Scholar] [CrossRef]
- Mashanov, V.S.; Zueva, O.R.; García-Arrarás, J.E. Transcriptomic changes during regeneration of the central nervous system in an echinoderm. BMC Genom. 2014, 15, 357. [Google Scholar] [CrossRef] [Green Version]
- Maki, N.; Suetsugu-Maki, R.; Sano, S.; Nakamura, K.; Nishimura, O.; Tarui, H.; Del Rio-Tsonis, K.; Ohsumi, K.; Agata, K.; Tsonis, P.A. Oocyte-type linker histone B4 is required for transdifferentiation of somatic cells in vivo. FASEB J. 2010, 24, 3462–3467. [Google Scholar] [CrossRef] [Green Version]
- Maki, N.; Suetsugu-Maki, R.; Tarui, H.; Agata, K.; Del Rio-Tsonis, K.; Tsonis, P.A. Expression of stem cell pluripotency factors during regeneration in newts. Dev. Dyn. 2009, 238, 1613–1616. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Pao, G.M.; Satoh, A.; Cummings, G.; Monaghan, J.R.; Harkins, T.T.; Bryant, S.V.; Randal Voss, S.; Gardiner, D.M.; Hunter, T. Activation of germline-specific genes is required for limb regeneration in the Mexican axolotl. Dev. Biol. 2012, 370, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Dolmatov, I.Y.; Eliseikina, M.G.; Bulgakov, A.A.; Ginanova, T.T.; Lamash, N.E.; Korchagin, V.P. Muscle regeneration in the holothurian Stichopus japonicus. Roux Arch. Dev. Biol. 1996, 205, 486–493. [Google Scholar] [CrossRef]
- Frolova, L.T.; Dolmatov, I.Y. Microscopic anatomy of the digestive system in normal and regenerating specimens of the brittlestar Amphipholis kochii. Biol. Bull. 2010, 218, 303–316. [Google Scholar] [CrossRef] [PubMed]
- García-Arrarás, J.E.; Dolmatov, I.Y. Echinoderms: Potential model systems for studies on muscle regeneration. Curr. Pharm. Des. 2010, 16, 942–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Arrarás, J.E.; Estrada-Rodgers, L.; Santiago, R.; Torres, I.I.; Díaz-Miranda, L.; Torres-Avillán, I. Cellular mechanisms in the regeneration of the intestine of the sea cucumber, Holothuria glaberrima Selenka (Holothuroidea: Echinodermata). J. Exp. Zool. 1998, 281, 288–304. [Google Scholar] [CrossRef]
- Mladenov, P.V.; Bisgrove, B.; Asotra, S.; Burke, R.D. Mechanisms of arm tip regeneration in the sea star, Leptasterias hexactis. Roux Arch. Dev. Biol. 1989, 198, 19–28. [Google Scholar] [CrossRef]
- Mashanov, V.S.; Zueva, O.R.; Heinzeller, T. Regeneration of the radial nerve cord in a holothurian: A promising new model system for studying post-traumatic recovery in the adult nervous system. Tissue Cell 2008, 40, 351–372. [Google Scholar] [CrossRef]
- Mashanov, V.S.; Frolova, L.T.; Dolmatov, I.Y. Structure of the digestive tube in the holothurian Eupentacta fraudatrix (Holothuroidea, Dendrochirota). Russ. J. Mar. Biol. 2004, 30, 314–322. [Google Scholar] [CrossRef]
- Shukalyuk, A.I.; Dolmatov, I.Y. Regeneration of the digestive tube in the holothurian Apostichopus japonicus after evisceration. Russ. J. Mar. Biol. 2001, 27, 168–173. [Google Scholar] [CrossRef]
- Dolmatov, I.Y. Regeneration of the aquapharyngeal complex in the holothurian Eupentacta fraudatrix (Holothuroidea, Dendrochirota). In Keys for Regeneration. Monographs in Developmental Biology; Taban, C.H., Boilly, B., Eds.; Karger: Basel, Switzerland, 1992; Volume 23, pp. 40–50. [Google Scholar]
- Dolmatov, I.Y. Proliferation of tissues of regenerating aquapharyngeal complex in holothurians. Russ. J. Dev. Biol. 1993, 24, 72–81. [Google Scholar]
- Kalacheva, N.V.; Eliseikina, M.G.; Frolova, L.T.; Dolmatov, I.Y. Regeneration of the digestive system in the crinoid Himerometra robustipinna occurs by transdifferentiation of neurosecretory-like cells. PLoS ONE 2017, 12, e0182001. [Google Scholar] [CrossRef] [Green Version]
- Kamenev, Y.O.; Dolmatov, I.Y. Posterior regeneration following fission in the holothurian Cladolabes schmeltzii (Dendrochirotida: Holothuroidea). Microsc. Res. Tech. 2015, 78, 540–552. [Google Scholar] [CrossRef]
- Mashanov, V.S.; Dolmatov, I.Y.; Heinzeller, T. Transdifferentiation in holothurian gut regeneration. Biol. Bull. 2005, 209, 184–193. [Google Scholar] [CrossRef]
- Kalacheva, N.V.; Dolmatov, I.Y. Cellular source of digestive system regeneration in Lamprometra palmata and Anneissia bennetti. In Proceedings of the Abstracts of 10th European Conference on Echinoderms, Borissiak Paleontological Institute RAS, Moscow, Russia, 16‒19 September 2019; p. 42. [Google Scholar]
- Mozzi, D.; Dolmatov, I.Y.; Bonasoro, F.; Candia Carnevali, M.D. Visceral regeneration in the crinoid Antedon mediterranea: Basic mechanisms, tissues and cells involved in gut regrowth. Centr. Eur. J. Biol. 2006, 1, 609–635. [Google Scholar] [CrossRef] [Green Version]
- Dolmatov, I.Y. Development and evolution of the muscle system in the Echinodermata. In Echinoderms: Durham; Harris, L.G., Bottger, S.A., Walker, C.W., Lesser, M.P., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 163–166. [Google Scholar]
- Dolmatov, I.Y.; Ivantey, V.A. Histogenesis of longitudinal muscle bands in holothurians. Russ. J. Dev. Biol. 1993, 24, 67–72. [Google Scholar]
- Dolmatov, I.Y.; Mashanov, V.S.; Zueva, O.R. Derivation of muscles of the Aristotle’s lantern from coelomic epithelia. Cell Tissue Res. 2007, 327, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Candelaria, A.G.; Murray, G.; File, S.K.; García-Arrarás, J.E. Contribution of mesenterial muscle dedifferentiation to intestine regeneration in the sea cucumber Holothuria glaberrima. Cell Tissue Res. 2006, 325, 55–65. [Google Scholar] [CrossRef] [PubMed]
- García-Arrarás, J.E.; Bello, S.A.; Malavez, S. The mesentery as the epicenter for intestinal regeneration. Semin. Cell Dev. Biol. 2019, 92, 45–54. [Google Scholar] [CrossRef]
- Murray, G.; García-Arrarás, J.E. Myogenesis during holothurian intestinal regeneration. Cell Tissue Res. 2004, 318, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Ariza, L.; Carmona, R.; Cañete Sánchez, A.; Cano, E.; Muñoz-Chápuli, R. Coelomic epithelium-derived cells in visceral morphogenesis. Dev. Dyn. 2015, 245, 307–322. [Google Scholar] [CrossRef] [PubMed]
- Smiley, S. Holothuroidea. In Microscopic Anatomy of Invertebrates; Harrison, F.W., Chia, F.S., Eds.; Wiley-Liss Inc.: New York, NY, USA, 1994; Volume 14, pp. 401–471. [Google Scholar]
- Smiley, S.; McEuen, F.S.; Chaffee, C.; Krishnan, S. Echinodermata: Holothuroidea. In Reproduction of Marine Invertebrates; Giese, A.C., Pearse, J.S., Pearse, V.B., Eds.; Boxwood: Pacific Grove, CA, USA, 1991; Volume 6, pp. 633–750. [Google Scholar]
- Dolmatov, I.Y. Variability of regeneration mechanisms in echinoderms. Russ. J. Mar. Biol. 2020, 46, 391–404. [Google Scholar] [CrossRef]
- Dolmatov, I.Y. Asexual reproduction in holothurians. Sci. World J. 2014, 2014, 13. [Google Scholar] [CrossRef]
- Kamenev, Y.O.; Dolmatov, I.Y. Anterior regeneration after fission in the holothurian Cladolabes schmeltzii (Dendrochirotida: Holothuroidea). Microsc. Res. Tech. 2017, 80, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Leibson, N.L. Regeneration of digestive tube in holothurians Stichopus japonicus and Eupentacta fraudatrix. In Keys for Regeneration. Monographs in Developmental Biology; Taban, C.H., Boilly, B., Eds.; Karger: Basel, Switzerland, 1992; Volume 23, pp. 51–61. [Google Scholar]
- Wang, X.; Li, X. The morphological and histological observation of regeneration of alimentary tract in sea cucumber Apostichopus japonicus. J. Dalian Fish. Univ. 2007, 22, 340–346. [Google Scholar]
- Mladenov, P.V.; Burke, R.D. Echinodermata: Asexual propagation. In Reproductive Biology of Invertebrates. Asexual Propagation and Reproductive Strategies; Adiyodi, K.G., Adiyodi, R.G., Eds.; Oxford and IBH Publishing Co. PVT. LTD.: New Delhi, India; Bombay, India; Calcutta, India, 1994; Volume 6, pp. 339–383. [Google Scholar]
- Motokawa, T.; Sato, E.; Umeyama, K. Energy expenditure associated with softening and stiffening of echinoderm connective tissue. Biol. Bull. 2012, 222, 150–157. [Google Scholar] [CrossRef]
- Motokawa, T.; Tsuchi, A. Dynamic mechanical properties of body-wall dermis in various mechanical states and their implications for the behavior of sea cucumbers. Biol. Bull. 2003, 205, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, I.C. Autotomy as a prelude to regeneration in echinoderms. Microsc. Res. Tech. 2001, 55, 369–396. [Google Scholar] [CrossRef]
- Wilkie, I.C. Mutable collagenous tissue: Overview and biotechnological perspective. In Progress in Molecular and Subcellular Biology. Subseries Marine Molecular Biotechnology; Matranga, V., Ed.; Springer: Heidelberg, Germany, 2005; pp. 221–250. [Google Scholar]
- Wilkie, I.C. Variable tensility in echinoderm collagenous tissues: A review. Mar. Behav. Physiol. 1984, 11, 1–34. [Google Scholar] [CrossRef]
- Motokawa, T. Connective tissue catch in echinoderms. Biol. Rev. 1984, 59, 255–270. [Google Scholar] [CrossRef]
- Ribeiro, A.R.; Barbaglio, A.; Oliveira, M.J.; Ribeiro, C.C.; Wilkie, I.C.; Candia Carnevali, M.D.; Barbosa, M.A. Matrix metalloproteinases in a sea urchin ligament with adaptable mechanical properties. PLoS ONE 2012, 7, e49016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolmatov, I.Y.; Afanasyev, S.V.; Boyko, A.V. Molecular mechanisms of fission in echinoderms: Transcriptome analysis. PLoS ONE 2018, 13, e0195836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz-Pineda, P.A.; Ramírez-Gómez, F.; Pérez-Ortiz, J.; González-Díaz, S.; Jesús, F.S.; Hernández-Pasos, J.; Valle-Avila, C.D.; Rojas-Cartagena, C.; Suárez-Castillo, E.C.; Tossas, K.; et al. Gene expression profiling of intestinal regeneration in the sea cucumber. BMC Genom. 2009, 10, 262. [Google Scholar] [CrossRef] [Green Version]
- Rojas-Cartagena, C.; Ortiz-Pineda, P.; Ramirez-Gomez, F.; Suarez-Castillo, E.C.; Matos-Cruz, V.; Rodriguez, C.; Ortiz-Zuazaga, H.; Garcia-Arraras, J.E. Distinct profiles of expressed sequence tags during intestinal regeneration in the sea cucumber Holothuria glaberrima. Physiol. Genom. 2007, 31, 203–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Chen, M.; Yang, H.; Wang, T.; Liu, B.; Shu, C.; Gardiner, D.M. Large scale gene expression profiling during intestine and body wall regeneration in the sea cucumber Apostichopus japonicus. Comp. Biochem. Physiol. Part D Genom. Proteom. 2011, 6, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yang, H.; Chen, M.; Ma, D.; Lin, C. RNA-Seq reveals dynamic changes of gene expression in key stages of intestine regeneration in the sea cucumber Apostichopus japonicus. PLoS ONE 2013, 8, e69441. [Google Scholar]
- Hynes, R.O.; Naba, A. Overview of the matrisome—An inventory of extracellular matrix constituents and functions. Cold Spring Harb. Perspect. Biol. 2012, 4, a004903. [Google Scholar] [CrossRef] [Green Version]
- Tucker, R.P.; Chiquet-Ehrismann, R. Evidence for the evolution of tenascin and fibronectin early in the chordate lineage. Int. J. Biochem. Cell Biol. 2009, 41, 424–434. [Google Scholar] [CrossRef] [PubMed]
- Chiquet-Ehrismann, R.; Tucker, R.P. Tenascins and the importance of adhesion modulation. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarzbauer, J.E.; DeSimone, D.W. Fibronectins, their fibrillogenesis, and in vivo functions. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ba, H.; Yao, F.; Yang, L.; Qin, T.; Luan, H.; Li, Z.; Zou, X.; Hou, L. Identification and expression patterns of extracellular matrix-associated genes fibropellin-ia and tenascin involved in regeneration of sea cucumber Apostichopus japonicus. Gene 2015, 565, 96–105. [Google Scholar] [CrossRef]
- Hynes, R.O. The evolution of metazoan extracellular matrix. J. Cell Biol. 2012, 196, 671–679. [Google Scholar] [CrossRef] [Green Version]
- Thurmond, F.A.; Koob, T.J.; Bowness, J.M.; Trotter, J.A. Partial biochemical and immunologic characterization of fibrillin microfibrils from sea cucumber dermis. Connect. Tissue Res. 1997, 36, 211–222. [Google Scholar] [CrossRef]
- Ribeiro, A.R.; Barbaglio, A.; Benedetto, C.D.; Ribeiro, C.C.; Wilkie, I.C.; Carnevali, M.D.C.; Barbosa, M.A. New insights into mutable collagenous tissue: Correlations between the microstructure and mechanical state of a sea-urchin ligament. PLoS ONE 2011, 6, e24822. [Google Scholar] [CrossRef] [Green Version]
- Giusti, B.; Pepe, G. Fibrillins in tendon. Front. Aging Neurosci. 2016, 8, 237. [Google Scholar] [CrossRef]
- Olivieri, J.; Smaldone, S.; Ramirez, F. Fibrillin assemblies: Extracellular determinants of tissue formation and fibrosis. Fibrogenesis Tissue Repair. 2010, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Sengle, G.; Sakai, L.Y. The fibrillin microfibril scaffold: A niche for growth factors and mechanosensation? Matrix Biol. 2015, 47, 3–12. [Google Scholar] [CrossRef]
- Whittaker, C.A.; Bergeron, K.-F.; Whittle, J.; Brandhorst, B.P.; Burke, R.D.; Hynes, R.O. The echinoderm adhesome. Dev. Biol. 2006, 300, 252–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vega, S.; Iwamoto, T.; Yamada, Y. Fibulins: Multiple roles in matrix structures and tissue functions. Cell. Mol. Life Sci. 2009, 66, 1890–1902. [Google Scholar] [CrossRef]
- Lee, N.V.; Rodriguez-Manzaneque, J.C.; Thai, S.N.-M.; Twal, W.O.; Luque, A.; Lyons, K.M.; Argraves, W.S.; Iruela-Arispe, M.L. Fibulin-1 acts as a cofactor for the matrix metalloprotease ADAMTS-1. J. Biol. Chem. 2005, 280, 34796–34804. [Google Scholar] [CrossRef] [Green Version]
- Adams, J.C.; Lawler, J. The thrombospondins. Cold Spring Harb. Perspect. Biol. 2011, 3, a009712. [Google Scholar] [CrossRef] [PubMed]
- Bein, K.; Simons, M. Thrombospondin type 1 repeats interact with matrix metalloproteinase 2: Regulation of metalloproteinase activity. J. Biol. Chem. 2000, 275, 32167–32173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Kyriakides, T.R.; Bornstein, P. Matricellular proteins as modulators of cell-matrix interactions: Adhesive defect in thrombospondin 2-null fibroblasts is a consequence of increased levels of matrix metalloproteinase-2. Mol. Biol. Cell. 2000, 11, 3353–3364. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Strickland, D.K.; Bornstein, P. Extracellular matrix metalloproteinase 2 levels are regulated by the low density lipoprotein-related scavenger receptor and Thrombospondin 2. J. Biol. Chem. 2001, 276, 8403–8408. [Google Scholar] [CrossRef] [Green Version]
- Pasten, C.; Rosa, R.; Ortiz, S.; Gonzalez, S.; García-Arrarás, J.E. Characterization of proteolytic activities during intestinal regeneration of the sea cucumber, Holothuria glaberrima. Int. J. Dev. Biol. 2012, 56, 681–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.; Dahms, H.U.; Han, L.L.; Li, Q.Y.; Zhang, Q.Z.; Wu, R.J.; Tan, J.; Zou, X.Y.; Hou, L. Cloning and characterization of a trypsin-like serine protease gene, a novel regeneration-related gene from Apostichopus japonicus. Gene 2012, 502, 46–52. [Google Scholar] [CrossRef]
- Miao, T.; Wan, Z.; Sun, L.; Li, X.; Xing, L.; Bai, Y.; Wang, F.; Yang, H. Extracellular matrix remodeling and matrix metalloproteinases (ajMMP-2 like and ajMMP-16 like) characterization during intestine regeneration of sea cucumber Apostichopus japonicus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2017, 212, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Lamash, N.E.; Dolmatov, I.Y. Proteases from the regenerating gut of the holothurian Eupentacta fraudatrix. PLoS ONE 2013, 8, e58433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shulga, A.P.; Lamash, N.E. Proteinases with gelatinase activity and their role in ambulacrum regeneration in holothurians Eupentacta fraudatrix (D’yakonov and Baranova, 1958) and Cucumaria japonica (Semper, 1868) (Echinodermata: Holothuroidea). Russ. J. Mar. Biol. 2020, 46, 461–471. [Google Scholar] [CrossRef]
- Dolmatov, I.Y.; Shulga, A.P.; Ginanova, T.T.; Eliseikina, M.G.; Lamash, N.E. Metalloproteinase inhibitor GM6001 delays regeneration in holothurians. Tissue Cell 2019, 59, 1–9. [Google Scholar] [CrossRef]
- Quiñones, J.L.; Rosa, R.; Ruiz, D.L.; García-Arrarás, J.E. Extracellular matrix remodeling and metalloproteinase involvement during intestine regeneration in the sea cucumber Holothuria glaberrima. Dev. Biol. 2002, 250, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Dolmatov, I.Y.; Kalacheva, N.V.; Shulga, A.P.; Tkacheva, E.S.; Boyko, A.V.; Girich, A.S. Expression of MMP, TIMP, and Sox genes during regeneration in holothurian Eupentacta fraudatrix. Genes 2021, in press. [Google Scholar]
- Boyko, A.V.; Girich, A.S.; Tkacheva, E.S.; Dolmatov, I.Y. The Eupentacta fraudatrix transcriptome provides insights into regulation of cell transdifferentiation. Sci. Rep. 2020, 10, 1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clouse, R.M.; Linchangco, G.V.; Kerr, A.M.; Reid, R.W.; Janies, D.A. Phylotranscriptomic analysis uncovers a wealth of tissue inhibitor of metalloproteinases variants in echinoderms. R. Soc. Open Sci. 2015, 2, 150377. [Google Scholar] [CrossRef] [Green Version]
- Tipper, J.P.; Lyons-Levy, G.; Atkinson, M.A.; Trotter, J.A. Purification, characterization and cloning of tensilin, the collagen-fibril binding and tissue-stiffening factor from Cucumaria frondosa dermis. Matrix Biol. 2002, 21, 625–635. [Google Scholar] [CrossRef]
- Miller, A.K.; Kerr, A.M.; Paulay, G.; Reich, M.; Wilson, N.G.; Carvajal, J.I.; Rouse, G.W. Molecular phylogeny of extant Holothuroidea (Echinodermata). Mol. Phylogenet. Evol. 2017, 111, 110–131. [Google Scholar] [CrossRef] [Green Version]
- Shivdasani, R.A. Molecular regulation of vertebrate early endoderm development. Dev. Biol. 2002, 249, 191–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorn, A.M.; Wells, J.M. Molecular basis of vertebrate endoderm development. Int. Rev. Cytol. 2007, 259, 49–111. [Google Scholar]
- Laverriere, A.C.; MacNeill, C.; Mueller, C.; Poelmann, R.E.; Burch, J.B.; Evans, T. GATA-4/5/6, a subfamily of three transcription factors transcribed in developing heart and gut. J. Biol. Chem. 1994, 269, 23177–23184. [Google Scholar] [CrossRef]
- Quispe-Parra, D.J.; Medina-Feliciano, J.G.; Cruz-González, S.; Ortiz-Zuazaga, H.; García-Arrarás, J.E. Transcriptomic analysis of early stages of intestinal regeneration in Holothuria glaberrima. Sci. Rep. 2021, 11, 346. [Google Scholar] [CrossRef] [PubMed]
- Vazzana, M.; Siragusa, T.; Arizza, V.; Buscaino, G.; Celi, M. Cellular responses and HSP70 expression during wound healing in Holothuria tubulosa (Gmelin, 1788). Fish Shellfish Immunol. 2015, 42, 306–315. [Google Scholar] [CrossRef]
- Li, W.; Sahu, D.; Tsen, F. Secreted heat shock protein-90 (Hsp90) in wound healing and cancer. Biochim. Biophys. Acta 2012, 1823, 730–741. [Google Scholar] [CrossRef] [Green Version]
- Wallin, R.P.A.; Lundqvist, A.; Moré, S.H.; von Bonin, A.; Kiessling, R.; Ljunggren, H.-G. Heat-shock proteins as activators of the innate immune system. Trends Immunol. 2002, 23, 130–135. [Google Scholar] [CrossRef]
- Browne, C.L.; Swan, J.B.; Rankin, E.E.; Calvert, H.; Griffiths, S.; Tytell, M. Extracellular heat shock protein 70 has novel functional effects on sea urchin eggs and coelomocytes. J. Exp. Biol. 2007, 210, 1275–1287. [Google Scholar] [CrossRef] [Green Version]
- Gosslau, A.; Ruoff, P.; Mohsenzadeh, S.; Hobohm, U.; Rensing, L. Heat shock and oxidative stress-induced exposure of hydrophobic protein domains as common signal in the induction of hsp68. J. Biol. Chem. 2001, 276, 1814–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Pasos, J.; Valentín-Tirado, G.; García-Arrarás, J.E. Melanotransferrin: New homolog genes and their differential expression during intestinal regeneration in the sea cucumber Holothuria glaberrima. J. Exp. Zool. B Mol. Dev. Evol. 2017, 328, 259–274. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Gómez, F.; Ortíz-Pineda, P.A.; Rojas-Cartagena, C.; Suárez-Castillo, E.C.; García-Arrarás, J.E. Immune-related genes associated with intestinal tissue in the sea cucumber Holothuria glaberrima. Immunogenetics 2008, 60, 57–71. [Google Scholar] [CrossRef]
- Ramírez-Gómez, F.; Aponte-Rivera, F.; Méndez-Castaner, L.; García-Arrarás, J.E. Changes in holothurian coelomocyte populations following immune stimulation with different molecular patterns. Fish Shellfish Immunol. 2010, 29, 175–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zang, Y.; Tian, X.; Dong, S.; Dong, Y. Growth, metabolism and immune responses to evisceration and the regeneration of viscera in sea cucumber, Apostichopus japonicus. Aquaculture 2012, 358–359, 50–60. [Google Scholar] [CrossRef]
- Jiang, J.; Zhou, Z.; Dong, Y.; Zhao, Z.; Sun, H.; Wang, B.; Jiang, B.; Chen, Z.; Gao, S. Comparative expression analysis of immune-related factors in the sea cucumber Apostichopus japonicus. Fish Shellfish Immunol. 2018, 72, 342–347. [Google Scholar] [CrossRef]
- Zhou, Z.C.; Dong, Y.; Sun, H.J.; Yang, A.F.; Chen, Z.; Gao, S.; Jiang, J.W.; Guan, X.Y.; Jiang, B.; Wang, B. Transcriptome sequencing of sea cucumber (Apostichopus japonicus) and the identification of gene-associated markers. Mol. Ecol. Resour. 2014, 14, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Dolmatova, L.S.; Eliseykina, M.G.; Timchenko, N.F.; Kovaleva, A.L.; Shitkova, O.A. Generation of reactive oxygen species in different fractions of the coelomocytes of holothurian Eupentacta fraudatrix in response to the thermostable toxin of Yersinia pseudotuberculosis in vitro. Chin. J. Oceanol. Limn. 2003, 21, 293–304. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Ren, Y.; Li, Y.; Xia, B. Regulation of growth, intestinal microbiota, non-specific immune response and disease resistance of sea cucumber Apostichopus japonicus (Selenka) in biofloc systems. Fish Shellfish Immunol. 2018, 77, 175–186. [Google Scholar] [CrossRef]
- Thiel, M.; Zourelidis, C.; Peter, K. Die Rolle der polymorphkernigen neutrophilen Leukozyten in der Pathogenese des akuten Lungenversagens (ARDS). Der Anaesthesist 1996, 45, 113–130. [Google Scholar] [CrossRef] [PubMed]
- Dolmatova, L.S.; Dolmatov, I.Y. Markers for immunity deficiency in lead-treated holothurians. In Echinoderms: Durham; Harris, L.G., Bottger, S.A., Walker, C.W., Lesser, M.P., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 359–362. [Google Scholar]
- Dolmatova, L.S.; Eliseikina, M.G.; Romashina, V.V. Antioxidant enzymatic activity of coelomocytes of the Far East sea cucumber Eupentacta fraudatrix. J. Evol. Biochem. Phys. 2004, 40, 126–135. [Google Scholar] [CrossRef]
- Dolmatova, L.S.; Dolmatov, I.Y. Lead induces different responses of two subpopulations of phagocytes in the holothurian Eupentacta fraudatrix. J. Ocean Univ. China. 2018, 17, 1391–1403. [Google Scholar] [CrossRef]
- Wei, X.; Liu, X.; Yang, J.; Wang, S.; Sun, G.; Yang, J. Critical roles of sea cucumber C-type lectin in non-self recognition and bacterial clearance. Fish Shellfish Immunol. 2015, 45, 791–799. [Google Scholar] [CrossRef]
- Eliseikina, M.G.; Petrova, I.Y.; Magarlamov, T.Y.; Dolmatov, I.Y.; Bulgakov, A.A. Mannan-binding lectins of echinoderms are components of humoral defense. In Echinoderms; Heinzeller, T., Nebelsick, J.H., Eds.; Taylor and Francis Group: London, UK, 2004; pp. 119–125. [Google Scholar]
- Bulgakov, A.A.; Eliseikina, M.G.; Petrova, I.Y.; Nazarenko, E.L.; Kovalchuk, S.N.; Kozhemyako, V.B.; Rasskazov, V.A. Molecular and biological characterization of a mannan-binding lectin from the holothurian Apostichopus japonicus. Glycobiology 2007, 17, 1284–1298. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Cao, X.; Guo, M.; Li, C. Identification and characterization of a novel short-type peptidoglycan recognition protein in Apostichopus japonicus. Fish Shellfish Immunol. 2020, 99, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Santiago, P.; Roig-López, J.L.; Santiago, C.; García-Arrarás, J.E. Serum amyloid A protein in an echinoderm: Its primary structure and expression during intestinal regeneration in the sea cucumber Holothuria glaberrima. J. Exp. Zool. 2000, 288, 335–344. [Google Scholar] [CrossRef]
- Tossas, K.; Qi-Huang, S.; Cuyar, E.; García-Arrarás, J.E. Temporal and spatial analysis of enteric nervous system regeneration in the sea cucumber Holothuria glaberrima. Regeneration 2014, 1, 10–26. [Google Scholar] [CrossRef] [PubMed]
- García-Arrarás, J.E.; Valentín-Tirado, G.; Flores, J.E.; Rosa, R.J.; Rivera-Cruz, A.; San Miguel-Ruiz, J.E.; Tossas, K. Cell dedifferentiation and epithelial to mesenchymal transitions during intestinal regeneration in H. glaberrima. BMC Dev. Biol. 2011, 11, 61. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.N.; Sun, J.C.; Xu, Q.Z.; Li, X.N.; Zhang, L.B.; Yang, H.S. Metabolic responses to intestine regeneration in sea cucumbers Apostichopus japonicus. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 22, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Lin, C.; Li, X.; Xing, L.; Huo, D.; Sun, J.; Zhang, L.; Yang, H. Comparative phospho- and acetyl proteomics analysis of posttranslational modifications regulating intestine regeneration in sea cucumbers. Front. Physiol. 2018, 9, 836. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xu, D.; Xu, Q.; Sun, J.; Xing, L.; Zhang, L.; Yang, H. iTRAQ reveals proteomic changes during intestine regeneration in the sea cucumber Apostichopus japonicus. Comp. Biochem. Physiol. Part D Genom. Proteomics 2017, 22, 39–49. [Google Scholar] [CrossRef]
- Lever, M.; Slow, S. The clinical significance of betaine, an osmolyte with a key role in methyl group metabolism. Clin. Biochem. 2010, 43, 732–744. [Google Scholar] [CrossRef]
- Eckelbager, K.J.; Young, C.M. Ovarian ultrastructure and vitellogenesis in ten species of shallow-water and bathyal sea cucumbers (Echinodermata: Holothuroidea). J. Mar. Biol. Ass. UK 1992, 72, 759–781. [Google Scholar] [CrossRef]
- Krishnan, S.; Dale, T. Ultrastructural studies on the testis of Cucumaria frondosa (Holothuroidea: Echinodermata). Norw. J. Zool. 1975, 23, 1–15. [Google Scholar]
- Lawrence, J.M. A Functional Biology of Echinoderms; Johns Hopkins University Press: Baltimore, MD, USA, 1987. [Google Scholar]
- Lawrence, J.M. Energetic costs of loss and regeneration of arms in stellate echinoderms. Integr. Comp. Biol. 2010, 50, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Mashanov, V.S.; Zueva, O.R.; García-Arrarás, J.E. Myc regulates programmed cell death and radial glia dedifferentiation after neural injury in an echinoderm. BMC Dev. Biol. 2015, 15, 24. [Google Scholar] [CrossRef] [Green Version]
- Miao, Q.; Hill, M.C.; Chen, F.; Mo, Q.; Ku, A.T.; Ramos, C.; Sock, E.; Lefebvre, V.; Nguyen, H. SOX11 and SOX4 drive the reactivation of an embryonic gene program during murine wound repair. Nat. Comm. 2019, 10, 4042. [Google Scholar] [CrossRef] [Green Version]
- Meng, S.; Matrone, G.; Lv, J.; Chen, K.; Wong, W.T.; Cooke, J.P. LIM domain only 2 regulates endothelial proliferation, angiogenesis, and tissue regeneration. J. Am. Heart Assoc. 2016, 5, e004117. [Google Scholar] [CrossRef] [Green Version]
- Nam, C.-H.; Rabbitts, T.H. The role of LMO2 in development and in T cell leukemia after chromosomal translocation or retroviral insertion. Mol. Ther. 2006, 13, 15–25. [Google Scholar] [CrossRef]
- Sun, L.; Sun, J.; Li, X.; Zhang, L. Understanding regulation of microRNAs on intestine regeneration in the sea cucumber Apostichopus japonicus using high-throughput sequencing. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 22, 1–9. [Google Scholar] [CrossRef] [PubMed]
- McGinnis, W.; Krumlauf, R. Homeobox genes and axial patterning. Cell 1992, 68, 283–302. [Google Scholar] [CrossRef]
- Novikova, E.L.; Bakalenko, N.I.; Nesterenko, A.Y.; Kulakova, M.A. HOX genes and animal regeneration. Russ. J. Dev. Biol. 2016, 47, 173–180. [Google Scholar] [CrossRef]
- Wang, K.C.; Helms, J.A.; Chang, H.Y. Regeneration, repair and remembering identity: The three Rs of HOX gene expression. Trends Cell Biol. 2009, 19, 268–275. [Google Scholar] [CrossRef] [Green Version]
- Mashanov, V.S.; Zueva, O.R.; García-Arrarás, J.E. Expression of Wnt9, TCTP, and Bmp1/Tll in sea cucumber visceral regeneration. Gene Expr. Patterns 2012, 12, 24–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, P.C.; Gungi, A.; Unni, M. Cellular and molecular mechanisms of Hydra regeneration. Results Probl. Cell Differ. 2019, 68, 259–290. [Google Scholar]
- Scimone, M.L.; Cote, L.E.; Rogers, T.; Reddien, P.W. Two FGFRL-Wnt circuits organize the planarian anteroposterior axis. eLife 2016, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Sprangers, J.; Zaalberg, I.C.; Maurice, M.M. Organoid-based modeling of intestinal development, regeneration, and repair. Cell Death Differ. 2021, 28, 95–107. [Google Scholar] [CrossRef]
- Sikes, J.M.; Newmark, P.A. Restoration of anterior regeneration in a planarian with limited regenerative ability. Nature 2013, 500, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Sun, L.; Yuan, J.; Sun, Y.; Gao, Y.; Zhang, L.; Li, S.; Dai, H.; Hamel, J.F.; Liu, C.; et al. The sea cucumber genome provides insights into morphological evolution and visceral regeneration. PLoS Biol. 2017, 15, e2003790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwiatkowski, D.J. Functions of gelsolin: Motility, signaling, apoptosis, cancer. Curr. Opin. Cell Biol. 1999, 11, 103–108. [Google Scholar] [CrossRef]
- Vázquez-Vélez, G.E.; Rodríguez-Molina, J.F.; Quiñones-Frías, M.C.; Pagán, M. A proteoglycan-like molecule offers insights into ground substance changes during holothurian intestinal regeneration. J. Histochem. Cytochem. 2016, 64, 381–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bello, S.A.; Torres-Gutiérrez, V.; Rodríguez-Flores, E.J.; Toledo-Román, E.J.; Rodríguez, N.; Díaz-Díaz, L.M.; Vázquez-Figueroa, L.D.; Cuesta, J.M.; Grillo-Alvarado, V.; Amador, A.; et al. Insights into intestinal regeneration signaling mechanisms. Dev. Biol. 2020, 458, 12–31. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, Y.; Nakajima, Y.I.; Kuranaga, E. Apoptosis in cellular society: Communication between apoptotic cells and their neighbors. Int. J. Mol. Sci. 2016, 17, 2144. [Google Scholar] [CrossRef] [PubMed]
- Mashanov, V.S.; Zueva, O.R.; Rojas-Catagena, C.; García-Arrarás, J.E. Visceral regeneration in a sea cucumber involves extensive expression of survivin and mortalin homologs in the mesothelium. BMC Dev. Biol. 2010, 10, 117. [Google Scholar] [CrossRef] [Green Version]
- Gauron, C.; Rampon, C.; Bouzaffour, M.; Ipendey, E.; Teillon, J.; Volovitch, M.; Vriz, S. Sustained production of ROS triggers compensatory proliferation and is required for regeneration to proceed. Sci. Rep. 2013, 3, 2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogarty, C.E.; Diwanji, N.; Lindblad, J.L.; Tare, M.; Amcheslavsky, A.; Makhijani, K.; Brückner, K.; Fan, Y.; Bergmann, A. Extracellular reactive oxygen species drive apoptosis-induced proliferation via Drosophila macrophages. Curr. Biol. 2016, 26, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Chera, S.; Ghila, L.; Dobretz, K.; Wenger, Y.; Bauer, C.; Buzgariu, W.; Martinou, J.C.; Galliot, B. Apoptotic cells provide an unexpected source of Wnt3 signaling to drive hydra head regeneration. Dev. Cell 2009, 17, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, S.P.; Altieri, D.C. Survivin at a glance. J. Cell Sci. 2019, 132, jcs223826. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Sun, L.; Yang, H.; Zhang, L.; Miao, T.; Xing, L.; Huo, D. Identification and expression characterization of WntA during intestinal regeneration in the sea cucumber Apostichopus japonicus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2017, 210, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Gao, Y.; Sun, L.; Jin, S.; Zhang, X.; Liu, C.; Li, F.; Xiang, J. Wnt signaling pathway linked to intestinal regeneration via evolutionary patterns and gene expression in the sea cucumber Apostichopus japonicus. Front. Genet. 2019, 10, 10. [Google Scholar] [CrossRef]
- Sun, L.N.; Yang, H.S.; Chen, M.Y.; Xu, D.X. Cloning and expression analysis of Wnt6 and Hox6 during intestinal regeneration in the sea cucumber Apostichopus japonicus. Genet. Mol. Res. 2013, 12, 5321–5334. [Google Scholar] [CrossRef]
- Quispe-Parra, D.; Valentín, G.; García-Arrarás, J.E. A roadmap for intestinal regeneration. Int. J. Dev. Biol. 2020. [Google Scholar] [CrossRef]
- Huelsken, J.; Behrens, J. The Wnt signalling pathway. J. Cell Sci. 2002, 115, 3977–3978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Orte, E.; Sáenz-Narciso, B.; Moreno, S.; Cabello, J. Multiple functions of the noncanonical Wnt pathway. Trends Genet. 2013, 29, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Green, J.; Nusse, R.; van Amerongen, R. The role of Ryk and Ror receptor tyrosine kinases in Wnt signal transduction. Cold Spring Harb. Perspect. Biol. 2014, 6, a009175. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Mazariegos, J.; Schubert, M.; Laudet, V. Evolution of retinoic acid receptors and retinoic acid signaling. In The Biochemistry of Retinoic Acid Receptors I: Structure, Activation, and Function at the Molecular Level; Asson-Batres, M.A., Rochette-Egly, C., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 55–73. [Google Scholar]
- Mendoza-Parra, M.-A.; Gronemeyer, H. Integrative genomic to dissect retinoid functions. In The Biochemistry of Retinoic Acid Receptors I: Structure, Activation, and Function at the Molecular Level; Asson-Batres, M.A., Rochette-Egly, C., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 181–202. [Google Scholar]
- Yamakawa, S.; Morino, Y.; Kohtsuka, H.; Wada, H. Retinoic acid signaling regulates the metamorphosis of feather stars (Crinoidea, Echinodermata): Insight into the evolution of the animal life cycle. Biomolecules 2019, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- Maden, M. RA signaling in limb development and regeneration in different species. Sub-Cell. Biochem. 2020, 95, 87–117. [Google Scholar]
- Viera-Vera, J.; García-Arrarás, J.E. Molecular characterization and gene expression patterns of retinoid receptors, in normal and regenerating tissues of the sea cucumber, Holothuria glaberrima. Gene 2018, 654, 23–35. [Google Scholar] [CrossRef]
- Viera-Vera, J.; García-Arrarás, J.E. Retinoic acid signaling is associated with cell proliferation, muscle cell dedifferentiation, and overall rudiment size during intestinal regeneration in the sea cucumber, Holothuria glaberrima. Biomolecules 2019, 9, 873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, T.; Trojanowska, M.; Watson, D.K. Ets proteins in biological control and cancer. J. Cell. Biochem. 2004, 91, 896–903. [Google Scholar] [CrossRef] [Green Version]
- Oikawa, T.; Yamada, T. Molecular biology of the Ets family of transcription factors. Gene 2003, 303, 11–34. [Google Scholar] [CrossRef]
- Vervoort, M.; Meulemeester, D.; Béhague, J.; Kerner, P. Evolution of Prdm genes in animals: Insights from comparative. Genom. Mol. Biol. Evol. 2016, 33, 679–696. [Google Scholar] [CrossRef] [Green Version]
- Ilsley, M.D.; Gillinder, K.R.; Magor, G.W.; Huang, S.; Bailey, T.L.; Crossley, M.; Perkins, A.C. Krüppel-like factors compete for promoters and enhancers to fine-tune transcription. Nucleic Acids Res. 2017, 45, 6572–6588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, S.; Yuan, J.; Zhao, D.; Shen, N.; Chen, W.; Ding, Y.; Yu, D.; Li, J.; Pan, F.; Zhu, M.; et al. Regulation of mice liver regeneration by early growth response-1 through the GGPPS/RAS/MAPK pathway. Int. J. Biochem. Cell Biol. 2015, 64, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Wang, Y.; Liang, J.; Liu, Z.; Sun, X.; Cai, K. MiR-301b promotes the proliferation, mobility, and epithelial-to-mesenchymal transition of bladder cancer cells by targeting EGR1. Biochem. Cell Biol. 2017, 95, 571–577. [Google Scholar] [CrossRef]
- Riddle, M.R.; Weintraub, A.; Nguyen, K.C.; Hall, D.H.; Rothman, J.H. Transdifferentiation and remodeling of post-embryonic C. elegans cells by a single transcription factor. Development 2013, 140, 4844–4849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Materna, S.C.; Ransick, A.; Li, E.; Davidson, E.H. Diversification of oral and aboral mesodermal regulatory states in pregastrular sea urchin embryos. Dev. Biol. 2013, 375, 92–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lentjes, M.H.; Niessen, H.E.; Akiyama, Y.; de Bruïne, A.P.; Melotte, V.; van Engeland, M. The emerging role of GATA transcription factors in development and disease. Expert Rev. Mol. Med. 2016, 18, e3. [Google Scholar] [CrossRef] [PubMed]
- Gehrke, A.R.; Neverett, E.; Luo, Y.-J.; Brandt, A.; Ricci, L.; Hulett, R.E.; Gompers, A.; Ruby, J.G.; Rokhsar, D.S.; Reddien, P.W.; et al. Acoel genome reveals the regulatory landscape of whole-body regeneration. Science 2019, 363, eaau6173. [Google Scholar] [CrossRef]
- Yamane, M.; Ohtsuka, S.; Matsuura, K.; Nakamura, A.; Niwa, H. Overlapping functions of Krüppel-like factor family members: Targeting multiple transcription factors to maintain the naïve pluripotency of mouse embryonic stem cells. Development 2018, 145, dev162404. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, F.; Fernandez-Serra, M.; Squarzoni, P.; Archimandritis, A.; Arnone, M.I. Identification and developmental expression of the ets gene family in the sea urchin (Strongylocentrotus purpuratus). Dev. Biol. 2006, 300, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Nigmatullina, L.; Norkin, M.; Dzama, M.M.; Messner, B.; Sayols, S.; Soshnikova, N. Id2 controls specification of Lgr5+ intestinal stem cell progenitors during gut development. EMBO J. 2017, 36, 869–885. [Google Scholar] [CrossRef] [Green Version]
- Hishikawa, K.; Marumo, T.; Miura, S.; Nakanishi, A.; Matsuzaki, Y.; Shibata, K.; Ichiyanagi, T.; Kohike, H.; Komori, T.; Takahashi, I.; et al. Musculin/MyoR is expressed in kidney side population cells and can regulate their function. J. Cell Biol. 2005, 169, 921–928. [Google Scholar] [CrossRef] [Green Version]
- DeBenedittis, P.; Jiao, K. Alternative splicing of T-box transcription factor genes. Biochem. Biophys. Res. Commun. 2011, 412, 513–517. [Google Scholar] [CrossRef] [Green Version]
- Takashima, Y.; Suzuki, A. Regulation of organogenesis and stem cell properties by T-box transcription factors. Cell. Mol. Life Sci. 2013, 70, 3929–3945. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Park, M.K.; Park, J.H.; Lee, H.J.; Shin, D.H.; Kang, Y.; Lee, C.H.; Kong, G. Loss of the polycomb protein Mel-18 enhances the epithelial–mesenchymal transition by ZEB1 and ZEB2 expression through the downregulation of miR-205 in breast cancer. Oncogene 2014, 33, 1325–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.F.; Zhang, X.W.; Hua, R.X.; Du, Y.Q.; Huang, M.Z.; Liu, Y.; Cheng, Y.F.; Guo, W.J. Mel-18 negatively regulates stem cell-like properties through downregulation of miR-21 in gastric cancer. Oncotarget 2016, 7, 63352–63361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalinkova, L.; Zmetakova, I.; Smolkova, B.; Minarik, G.; Sedlackova, T.; Horvathova Kajabova, V.; Cierna, Z.; Mego, M.; Fridrichova, I. Decreased methylation in the SNAI2 and ADAM23 genes associated with de-differentiation and haematogenous dissemination in breast cancers. BMC Cancer 2018, 18, 875. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, Q.; Dai, X.; Yan, Y.; Yang, Y.; Li, H.; Zhou, X.; Gao, W.; Li, X.; Xi, Z. Transdifferentiation of type II alveolar epithelial cells induces reactivation of dormant tumor cells by enhancing TGF-β1/SNAI2 signaling. Onco. Rep. 2018, 39, 1874–1882. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, K.; Qian, C.N.; Leach, R. DNA methylation is associated with transcription of Snail and Slug genes. Biochem. Biophys. Res. Commun. 2013, 430, 1083–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamata, Y.U.; Sumida, T.; Kobayashi, Y.; Ishikawa, A.; Kumamaru, W.; Mori, Y. Introduction of ID2 enhances invasiveness in ID2-null oral squamous cell carcinoma cells via the SNAIL axis. Cancer Genom. Proteom. 2016, 13, 493–497. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.; Yang, X.; Pursell, B.; Mercurio, A. Id2 complexes with the SNAG domain of Snai1 inhibiting Snai1-mediated repression of integrin 4. Mol. Cell. Biol. 2013, 33. [Google Scholar] [CrossRef] [Green Version]
- Radice, G.P. The spreading of epithelial cells during wound closure in Xenopus larvae. Dev. Biol. 1980, 76, 26–46. [Google Scholar] [CrossRef]
- Schöck, F.; Perrimon, N. Molecular mechanisms of epithelial morphogenesis. Annu. Rev. Cell Dev. Biol. 2002, 18, 463–493. [Google Scholar] [CrossRef] [Green Version]
- Schöck, F.; Perrimon, N. Cellular processes associated with germ band retraction in Drosophila. Dev. Biol. 2002, 248, 29–39. [Google Scholar] [CrossRef] [Green Version]
- McLellan, A.S.; Langlands, K.; Kealey, T. Exhaustive identification of human class II basic helix–loop–helix proteins by virtual library screening. Mech. Dev. 2002, 119, S285–S291. [Google Scholar] [CrossRef]
- Girich, A.S.; Isaeva, M.P.; Dolmatov, I.Y. Wnt and frizzled expression during regeneration of internal organs in the holothurian Eupentacta fraudatrix. Wound Repair Regen. 2017, 25, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Girich, A.S. (NSCMB FEB RAS, Vladivostok, Russia). Personal communication, 2021.
- Witte, F.; Dokas, J.; Neuendorf, F.; Mundlos, S.; Stricker, S. Comprehensive expression analysis of all Wnt genes and their major secreted antagonists during mouse limb development and cartilage differentiation. Gene Expr. Patterns 2009, 9, 215–223. [Google Scholar] [CrossRef]
- Rai, M.F.; Schmidt, E.J.; McAlinden, A.; Cheverud, J.M.; Sandell, L.J. Molecular insight into the association between cartilage regeneration and ear wound healing in genetic mouse models: Targeting new genes in regeneration. G3 2013, 3, 1881–1891. [Google Scholar] [CrossRef] [Green Version]
- Ozeki, N.; Mogi, M.; Hase, N.; Hiyama, T.; Yamaguchi, H.; Kawai, R.; Kondo, A.; Nakata, K. Wnt16 signaling is required for IL-1beta-induced matrix metalloproteinase-13-regulated proliferation of human stem cell-derived osteoblastic cells. Int. J. Mol. Sci. 2016, 17, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girich, A.S.; Dolmatov, I.Y.; Lamash, N.E. The distribution of the Wnt5 protein in the tissues of the holothurian Eupentacta fraudatrix (Djakonov et Baranova, 1958) (Holothuroidea: Dendrochirotida) in the norm and during regeneration. Russ. J. Mar. Biol. 2014, 40, 66–70. [Google Scholar] [CrossRef]
- Adell, T.; Salò, E.; Boutros, M.; Bartscherer, K. Smed-Evi/Wntless is required for β-catenin-dependent and -independent processes during planarian regeneration. Development 2009, 136, 905–910. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.; Ma, X.; Meng, J.; Zhang, C.; Ma, K. Role of Wnt-5A in interleukin-1–induced matrix metalloproteinase expression in rabbit temporomandibular joint condylar chondrocytes. Arthritis Rheum. 2009, 60, 2714–2722. [Google Scholar] [CrossRef] [PubMed]
- Bankaitis, E.D.; Ha, A.; Kuo, C.J.; Magness, S.T. Reserve stem cells in intestinal homeostasis and injury. Gastroenterology 2018, 155, 1348–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garipova, V.A.; Girich, A.S.; Dolmatov, I.Y. HOX genes in the holothurian Eupentacta fraudatrix and their participation in the regeneration. In Proceedings of the Systems Biology and Biomedicine (SBioMed-2018), Novosibirsk, Russia, 22–23 August 2018; p. 44. [Google Scholar]
- San Miguel-Ruiz, J.E.; Maldonado-Soto, A.R.; García-Arrarás, J.E. Regeneration of the radial nerve cord in the sea cucumber Holothuria glaberrima. BMC Dev. Biol. 2009, 9, 3. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dolmatov, I.Y. Molecular Aspects of Regeneration Mechanisms in Holothurians. Genes 2021, 12, 250. https://doi.org/10.3390/genes12020250
Dolmatov IY. Molecular Aspects of Regeneration Mechanisms in Holothurians. Genes. 2021; 12(2):250. https://doi.org/10.3390/genes12020250
Chicago/Turabian StyleDolmatov, Igor Yu. 2021. "Molecular Aspects of Regeneration Mechanisms in Holothurians" Genes 12, no. 2: 250. https://doi.org/10.3390/genes12020250
APA StyleDolmatov, I. Y. (2021). Molecular Aspects of Regeneration Mechanisms in Holothurians. Genes, 12(2), 250. https://doi.org/10.3390/genes12020250