
High Genetic Diversity despite Conserved Karyotype Organization in the Giant Trahiras from Genus Hoplias (Characiformes, Erythrinidae)

 ,
,  , ,
, ,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Individuals, Collection Sites, and Classical Cytogenetic Methods
2.2. Fish-Based Experiments
2.3. DArTseq Genotyping and Genetic Diversity
2.4. Genetic Relationships and Phylogenetic Estimation
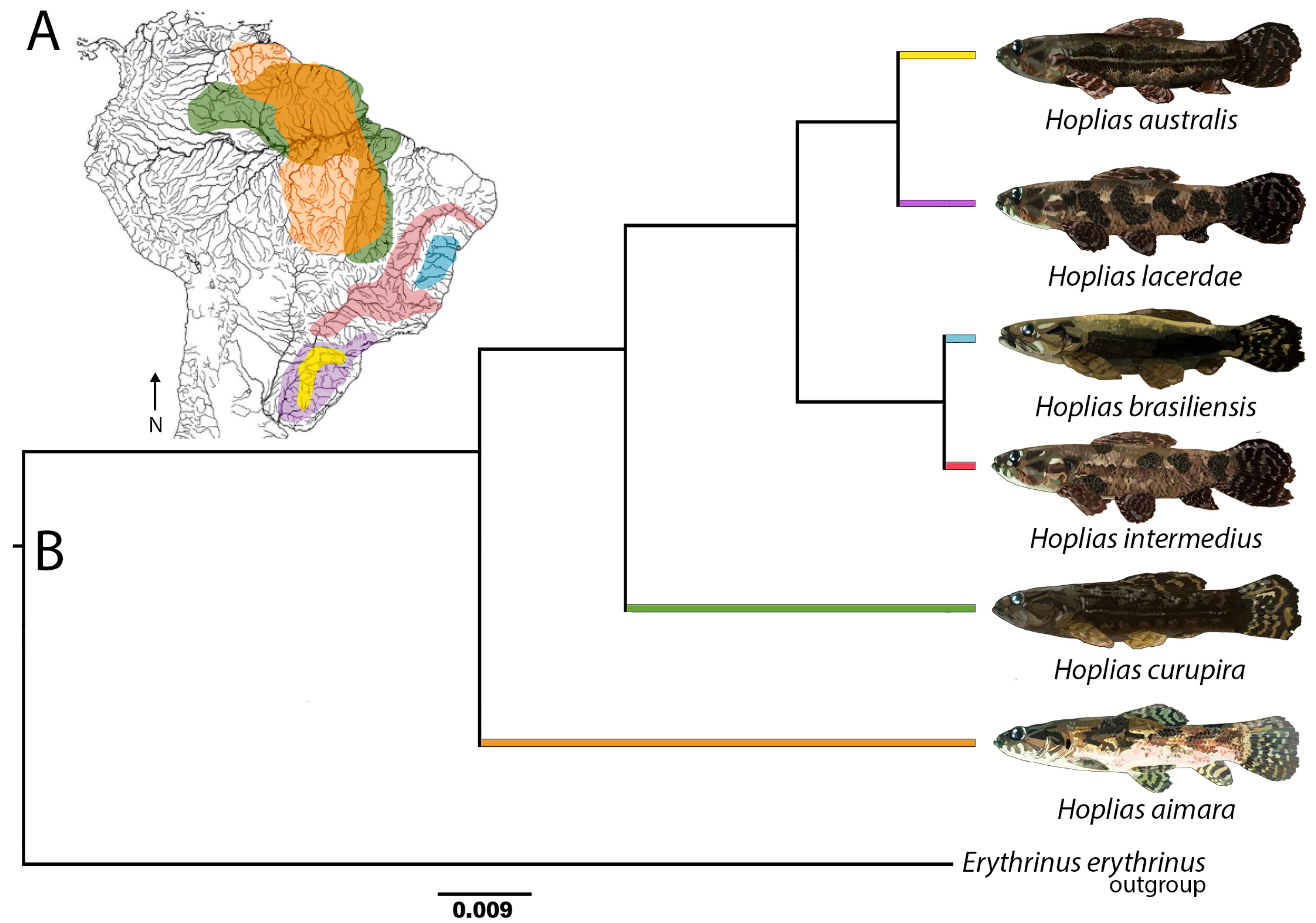
3. Results
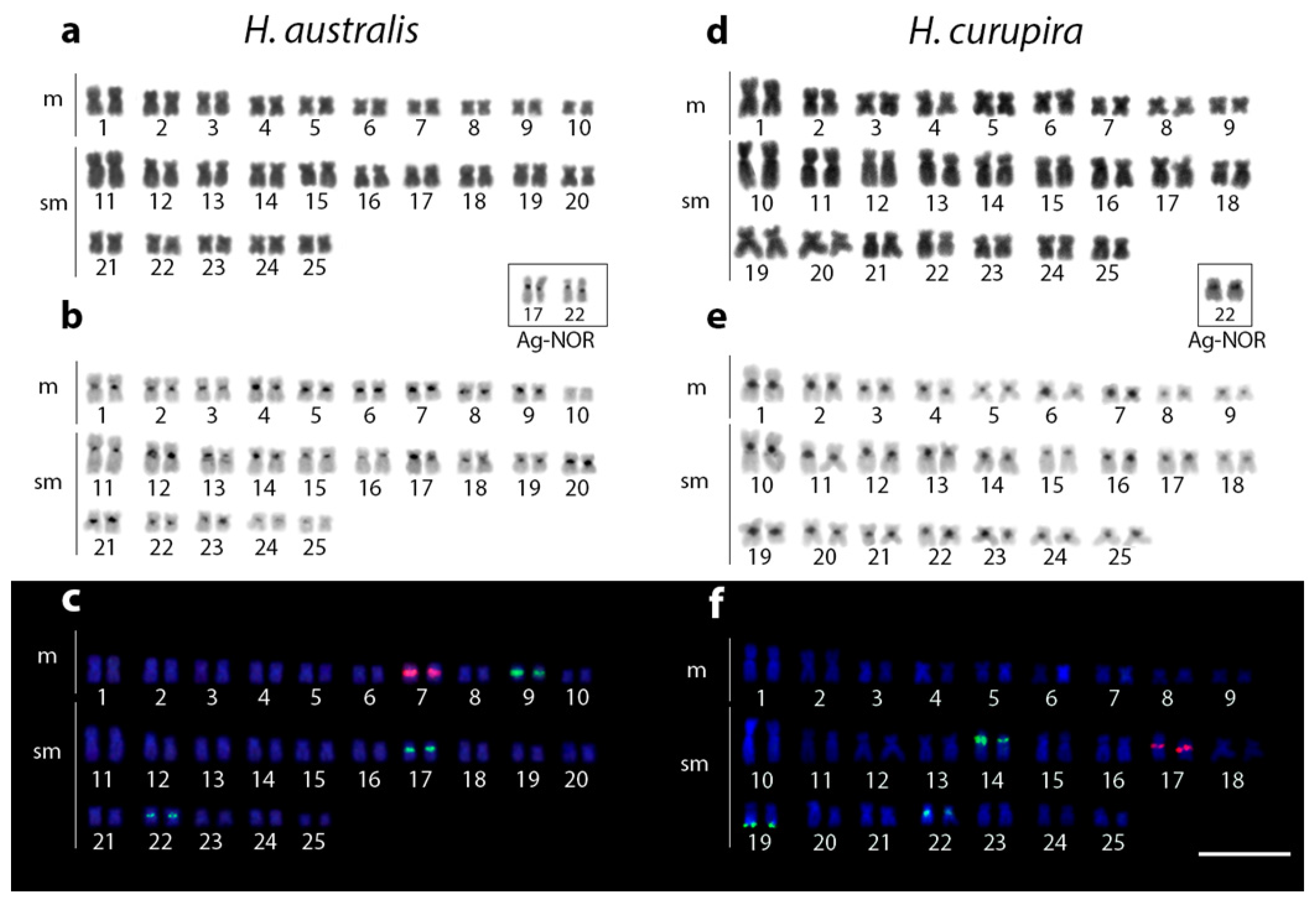
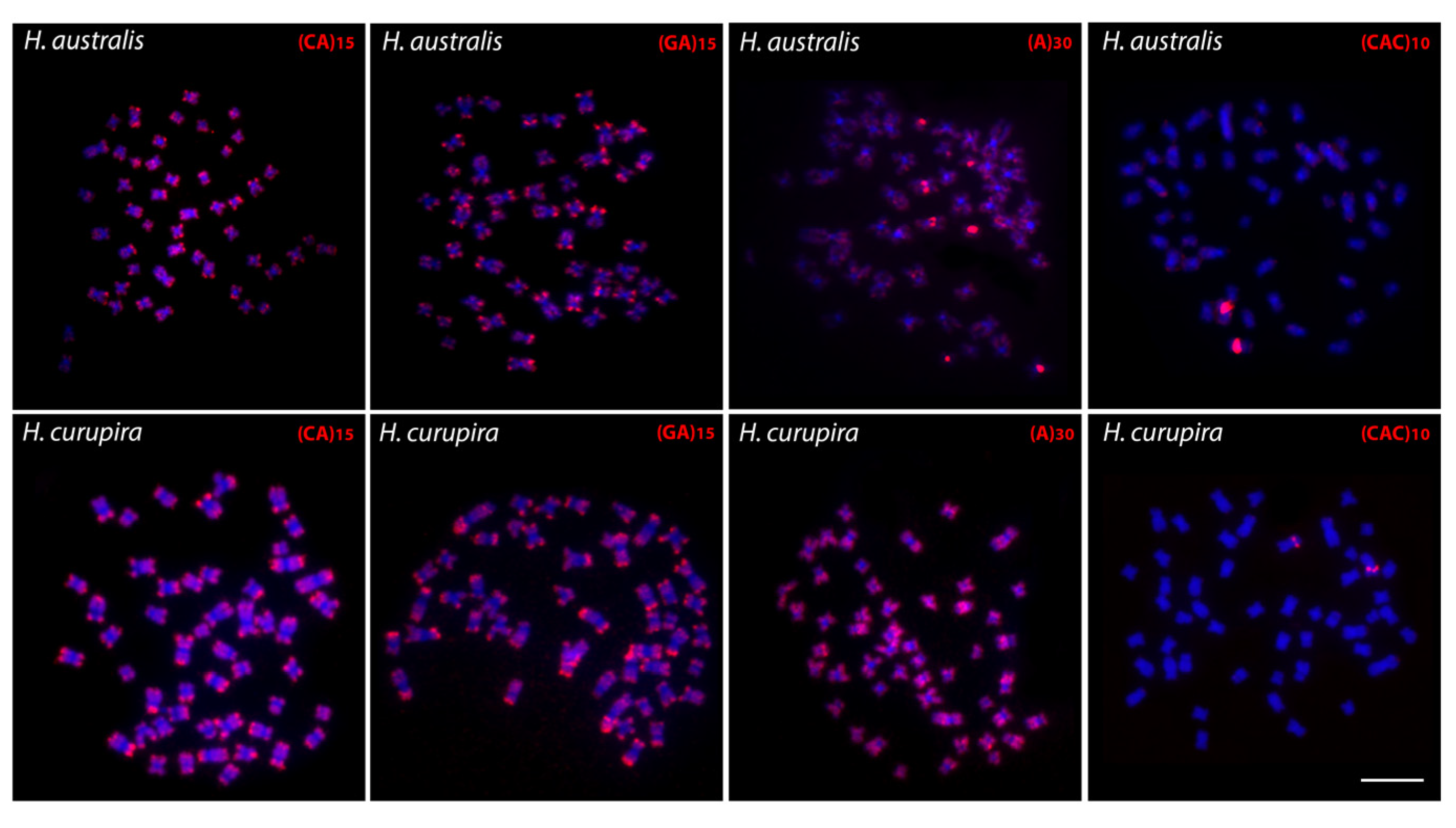
3.1. Karyotypes, C-Banding, and Chromosomal Mapping of Repetitive Sequences
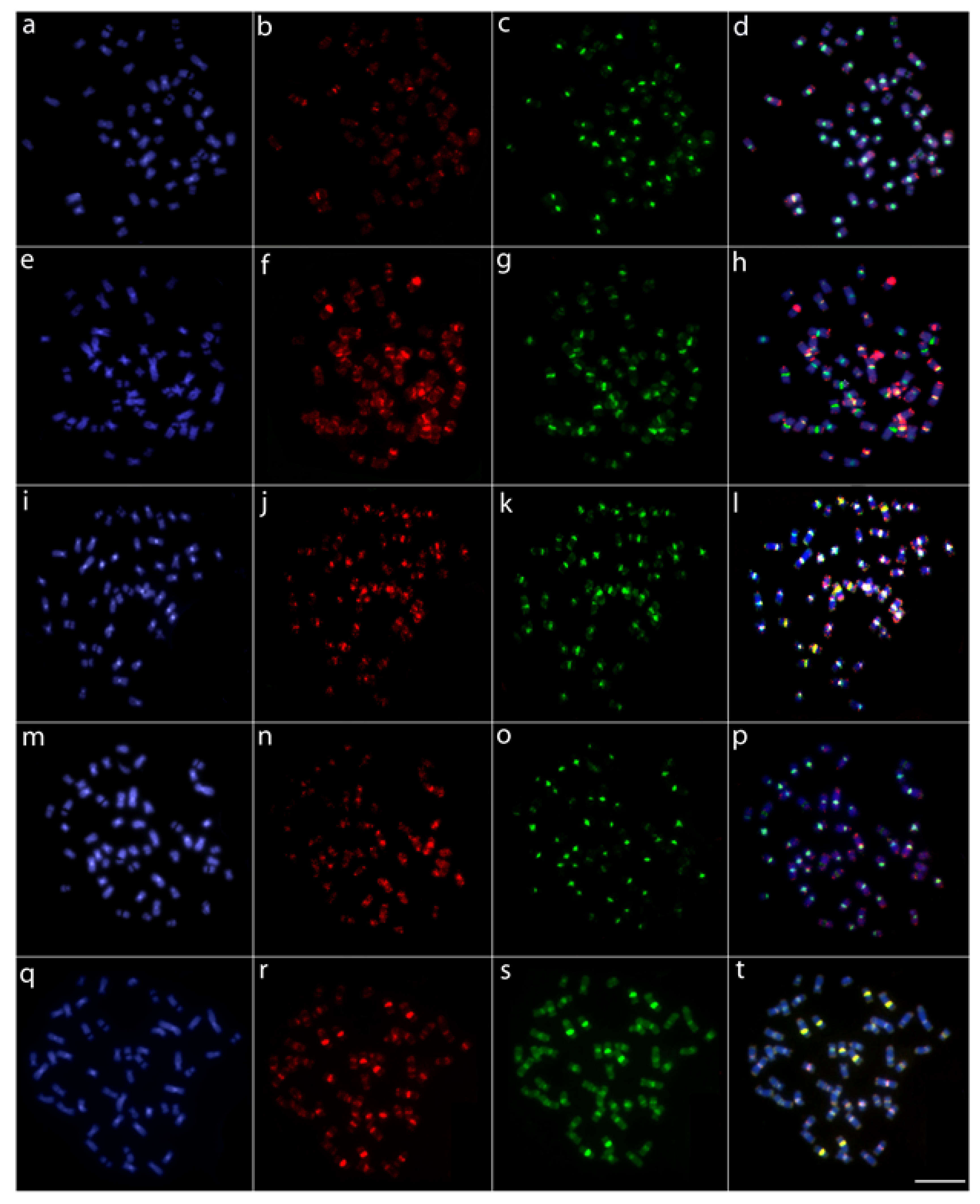
3.2. Comparative Genomic Hybridization (CGH)
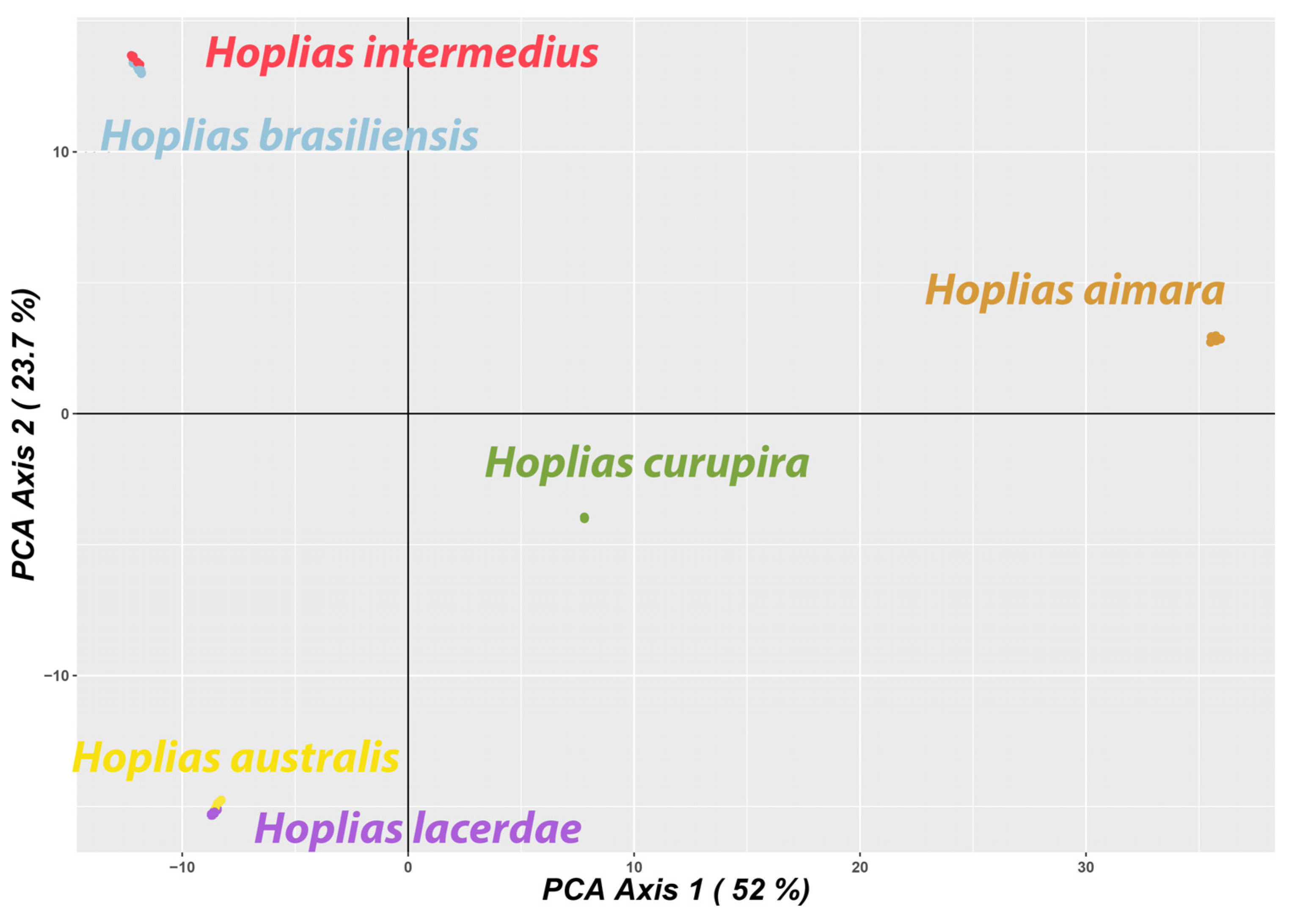
3.3. Comparative Analyses Using Diversity Arrays Technology Sequencing Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World; John Wiley & Sons: Hoboken, NJ, USA, 2016; ISBN 111834233X. [Google Scholar]
- Reis, R.E.; Albert, J.S.; Di Dario, F.; Mincarone, M.M.; Petry, P.; Rocha, L.A. Fish Biodiversity and Conservation in South America. J. Fish Biol. 2016, 89, 12–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fricke, R.; Eschmeyer, W.N.; van der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 12 February 2020).
- Oyakawa, O.T. Checklist of the Freshwater Fishes of South and Central America; EDIPUCRS: Porto Alegre, Brazil, 2003; pp. 238–240. [Google Scholar]
- Manel, S.; Guerin, P.-E.; Mouillot, D.; Blanchet, S.; Velez, L.; Albouy, C.; Pellissier, L. Global Determinants of Freshwater and Marine Fish Genetic Diversity. Nat. Commun. 2020, 11, 692. [Google Scholar] [CrossRef] [Green Version]
- Bertollo, L.A.C.; Born, G.G.; Dergam, J.A.; Fenocchio, A.S.; Moreira-Filho, O. A Biodiversity Approach in the Neotropical Erythrinidae Fish, Hoplias malabaricus. Karyotypic Survey, Geographic Distribution of Cytotypes and Cytotaxonomic Considerations. Chromosome Res. 2000, 8, 603–613. [Google Scholar] [CrossRef]
- De Bello Cioffi, M.; Franco, W.; Ferreira, R.; Bertollo, L.A.C. Chromosomes as tools for discovering Biodiversity—The case of Erythrinidae fish family. In Recent Trends Cytogenet Studies Methodol Appl; Tirunilai, P., Ed.; InTech: Rijeka, Croatia, 2012; pp. 125–146. ISBN 978-953-51-0178-9. [Google Scholar]
- Bertollo, L.A.C. Chromosome Evolution in the Neotropical Erythrinidae Fish Family: An Overview. In Fish Cytogenetics; Pisano, E., Ozouf-Costaz, C., Foresti, F., Kapoor, B.G., Eds.; CRC Press: Enfield, NH, USA, 2007; pp. 195–211. ISBN 9781578083305. [Google Scholar]
- De Bello Cioffi, M.; Bertollo, L.A.C. Initial Steps in XY Chromosome Differentiation in Hoplias malabaricus and the Origin of an X1X2Y Sex Chromosome System in This Fish Group. Heredity (Edinburgh) 2010. [Google Scholar] [CrossRef] [Green Version]
- De Bello Cioffi, M.; Yano, C.F.; Sember, A.; Bertollo, L.A.C. Chromosomal Evolution in Lower Vertebrates: Sex Chromosomes in Neotropical Fishes. Genes (Basel) 2017, 8, 258. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.F.; Lui, R.L.; Traldi, J.B.; Blanco, D.R.; Moreira-Filho, O. Comparative Cytogenetics of Hoplerythrinus unitaeniatus (Agassiz, 1829) (Characiformes, Erythrinidae) Species Complex from Different Brazilian Hydrographic Basins. Cytogenet. Genome Res. 2016, 149, 191–200. [Google Scholar] [CrossRef]
- Sember, A.; Bertollo, L.A.C.; Ráb, P.; Yano, C.F.; Hatanaka, T.; de Oliveira, E.A.; de Bello Cioffi, M. Sex Chromosome Evolution and Genomic Divergence in the Fish Hoplias malabaricus (Characiformes, Erythrinidae). Front. Genet. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Oyakawa, O.T.; Mattox, M.T. Revision of the Neotropical Trahiras of the Hoplias lacerdae Species-Group (Ostariophysi: Characiformes: Erythrinidae) with Descriptions of Two New Species. Neotrop. Ichthyol. 2009, 7. [Google Scholar] [CrossRef]
- Cardoso, Y.P.; Rosso, J.J.; Mabragaña, E.; González-Castro, M.; Delpiani, M.; Avigliano, E.; Bogan, S.; Covain, R.; Schenone, N.F.; Díaz de Astarloa, J.M. A Continental-Wide Molecular Approach Unraveling MtDNA Diversity and Geographic Distribution of the Neotropical Genus Hoplias. PLoS ONE 2018, 13, e0202024. [Google Scholar] [CrossRef] [Green Version]
- De Bello Cioffi, M.; Kejnovsky, E.; Bertollo, L.A.C. The Chromosomal Distribution of Microsatellite Repeats in the Genome of the Wolf Fish Hoplias malabaricus, Focusing on the Sex Chromosomes. Cytogenet. Genome Res. 2011, 132, 289–296. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, E.A.; Sember, A.; Bertollo, L.A.C.; Yano, C.F.; Ezaz, T.; Moreira-Filho, O.; Hatanaka, T.; Trifonov, V.; Liehr, T.; Al-Rikabi, A.B.H.; et al. Tracking the Evolutionary Pathway of Sex Chromosomes among Fishes: Characterizing the Unique XX/XY1Y2 System in Hoplias malabaricus (Teleostei, Characiformes). Chromosoma 2018, 127, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Bertollo, L.A.C.; Takahashi, C.; Moreira-Filho, O. Cytotaxonomic Considerations on Hoplias lacerdae (Pisces, Erythrinidae). Braz. J. Genet. 1978, 1, 103–120. [Google Scholar]
- Morelli, S.; Vicari, M.R.; Bertollo, L.A.C. Evolutionary Cytogenetics of the Hoplias lacerdae, Miranda Ribeiro, 1908 Group. A Particular Pathway Concerning the Other Erythrinidae Fish. Braz. J. Biol. 2007, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, E.A.; Bertollo, L.A.C.; Yano, C.F.; Liehr, T.; De Bello Cioffi, M. Comparative Cytogenetics in the Genus Hoplias (Characiformes, Erythrinidae) Highlights Contrasting Karyotype Evolution among Congeneric Species. Mol. Cytogenet. 2015, 8. [Google Scholar] [CrossRef] [Green Version]
- Mattox, G.T.; Toledo-Piza, M.; Oyakawa, O.T. Taxonomic Study of Hoplias aimara (Valenciennes, 1846) and Hoplias macrophthalmus (Pellegrin, 1907) (Ostariophysi, Characiformes, Erythrinidae). Copeia 2006. [Google Scholar] [CrossRef]
- Blanco, D.R.; Lui, R.L.; Vicari, M.R.; Bertollo, L.A.C.; Moreira-Filho, O. Comparative Cytogenetics of Giant Trahiras Hoplias aimara and H. intermedius (Characiformes, Erythrinidae): Chromosomal Characteristics of Minor and Major Ribosomal DNA and Cross-Species Repetitive Centromeric Sequences Mapping Differ among Morphologicall. Cytogenet. Genome Res. 2011, 900, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Leaché, A.D.; Bouckaert, R.R. Species Trees and Species Delimitation with SNAPP: A Tutorial and Worked Example. In Proceedings of the Workshop on Population and Speciation Genomics, Český Krumlov, Czech Republic, 21 January–3 February 2018. [Google Scholar]
- Bertollo, L.A.C.; De Bello Cioffi, M. Moreira-Filho, O. Direct chromosome preparation from Freshwater Teleost Fishes. In Fish Cytogenetic Techniques (Chondrichthyans and Teleosts); Ozouf-Costaz, C., Pisano, E., Foresti, F., Almeida Toledo, L.F., Eds.; CRC Press: Enfield, CT, USA, 2015; pp. 21–26. [Google Scholar]
- Sumner, A.T. A Simple Technique for Demonstrating Centromeric Heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Howell, W.M.; Black, D.A. Controlled Silver-Staining of Nucleolus Organizer Regions with a Protective Colloidal Developer: A 1-Step Method. Experientia 1980, 36, 1014–1015. [Google Scholar] [CrossRef]
- Pendás, A.M.; Móran, P.; Freije, J.P.; Garcia-Vásquez, E. Chromosomal Location and Nucleotide Sequence of Two Tandem Repeats of the Atlantic Salmon 5S RDNA. Cytogenet. Cell Genet. 1994, 67. [Google Scholar] [CrossRef]
- De Bello Cioffi, M.; Martins, C.; Centofante, L.; Jacobina, U.P.; Bertollo, L.A.C. Chromosomal Variability among Allopatric Populations of Erythrinidae Fish Hoplias malabaricus: Mapping of Three Classes of Repetitive DNAs. Cytogenet. Genome Res. 2009, 132–141. [Google Scholar] [CrossRef]
- Kubat, Z.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Microsatellite Accumulation in the Y Chromosome of Silene Latifolia. Genome 2008, 51, 350–356. [Google Scholar] [CrossRef] [Green Version]
- Yano, C.F.; Bertollo, L.A.C.; De Bello Cioffi, M. Fish-FISH: Molecular cytogenetics in fish species. In Fluorescence In Situ Hybridization (FISH)—Application Guide; Liehr, T., Ed.; Springer: Berlin, Germany, 2017; pp. 429–444. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 854; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Zwick, M.S.; Hanson, R.E.; Mcknight, T.D.; Islam-Faridi, M.H.; Stelly, D.M.; Wing, R.A.; Price, H.J. A Rapid Procedure for the Isolation of C 0 T-1 DNA from Plants. Genome 1997, 40, 138–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symonová, R.; Sember, A.; Majtánová, Z.; Ráb, P. Characterization of fish genome by GISH and CGH. In Fish Cytogenetic Techniques. Ray-Fin Fishes and Chondrichthyans; Ozouf-Costaz, C., Pisano, E., Foresti, F., de Almeida-Toledo, L.F., Eds.; CCR Press: Boca Raton, FL, USA, 2015; pp. 118–131. [Google Scholar]
- De Moraes, R.L.R.; Sember, A.; Bertollo, L.A.; De Oliveira, E.A.; Rab, P.; Hatanaka, T.; Marinho, M.M.; Liehr, T.; Al-Rikabi, A.B.H.; Feldberg, E.; et al. Comparative Cytogenetics and Neo-Y Formation in Small-Sized Fish Species of the Genus Pyrrhulina (Characiformes, Lebiasinidae). Front. Genet. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- De Menezes Cavalcante Sassi, F.; De Oliveira, E.A.; Bertollo, L.A.C.; Nirchio, M.; Hatanaka, T.; Ferreira Marinho, M.M.; Moreira-Filho, O.; Aroutiounian, R.; Liehr, T.; Al-Rikabi, A.B.H.; et al. Chromosomal Evolution and Evolutionary Relationships of Lebiasina Species (Characiformes, Lebiasinidae). Int. J. Mol. Sci. 2019, 20, 2944. [Google Scholar] [CrossRef] [Green Version]
- De Menezes Cavalcante Sassi, F.; Hatanaka, T.; De Moraes, R.L.R.; Toma, G.A.; De Oliveira, E.A.; Liehr, T.; Rab, P.; Bertollo, L.A.C.; Viana, P.F.; Feldberg, E.; et al. An Insight into the Chromosomal Evolution of Lebiasinidae (Teleostei, Characiformes). Genes (Basel) 2020, 11, 365. [Google Scholar]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for Centromeric Position on Chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Gruber, B.; Unmack, P.J.; Berry, O.F.; Georges, A. Dartr: An r Package to Facilitate Analysis of SNP Data Generated from Reduced Representation Genome Sequencing. Mol. Ecol. Resour. 2018, 18, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Sherwin, W.B.; Chao, A.; Jost, L.; Smouse, P.E. Information Theory Broadens the Spectrum of Molecular Ecology and Evolution. Trends Ecol. Evol. 2017, 32, 948–963. [Google Scholar] [CrossRef] [PubMed]
- Bryant, D.; Bouckaert, R.; Felsenstein, J.; Rosenberg, N.A.; RoyChoudhury, A. Inferring Species Trees Directly from Biallelic Genetic Markers: Bypassing Gene Trees in a Full Coalescent Analysis. Mol. Biol. Evol. 2012, 29, 1917–1932. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An Advanced Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Kirkpatrick, M.; Barton, N. Chromosome Inversions, Local Adaptation and Speciation. Genetics 2006, 173, 419–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertollo, L.A.C.; Moreira-Filho, O.; Galetti, P.M., Jr. Cytogenetics and Taxonomy: Consideration Based on Chromosome Studies of Freshwater Fish. J. Fish Biol. 1986, 28. [Google Scholar] [CrossRef]
- Oliveira, C.; Almeida-Toledo, L.F.; Foresti, F.; Britski, H.; Toledo-Filho, S.A. Chromosome Formulae of Neotropical Freshwater Fishes. Braz. J. Genet. 1988, 11, 577–624. [Google Scholar]
- Borges, A.T.; De Bello Cioffi, M.; Bertollo, L.A.C.; Soares, R.X.; Costa, G.W.W.F.; Molina, W.F. Paracentric Inversions Differentiate the Conservative Karyotypes in Two Centropomus Species (Teleostei: Centropomidae). Cytogenet. Genome Res. 2019, 157, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Biémont, C.; Vieira, C. Junk DNA as an Evolutionary Force. Nature 2006, 443, 521–524. [Google Scholar] [CrossRef]
- Raskina, O.; Barber, J.C.; Nevo, E.; Belyayev, A. Repetitive DNA and Chromosomal Rearrangements: Speciation Related Events in Plant Genomes. Cytogenet. Genome Res. 2008, 120. [Google Scholar] [CrossRef] [PubMed]
- De Bello Cioffi, M.; Bertollo, L.A.C. Chromosomal distribution and evolution of repetitive DNAs in fish. In Repetitive DNA; Garrido-Ramos, M.A., Ed.; Karger: Basel, Switzerland, 2012; pp. 197–221. ISBN 9783318021509. [Google Scholar]
- Sochorová, J.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. Evolutionary Trends in Animal Ribosomal DNA Loci: Introduction to a New Online Database. Chromosoma 2018, 127, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symonová, R. Integrative RDNAomics—Importance of the Oldest Repetitive Fraction of the Eukaryote Genome. Genes (Basel) 2019, 10, 345. [Google Scholar] [CrossRef] [Green Version]
- Rosa, R.S. Diversidade e Conservação Dos Peixes Da Caatinga. Biodiversidade Da Caatinga Áreas E Ações Prioritárias Para A Conservaҫão 2004, 1, 149–161. [Google Scholar]
- Ribeiro, A.C. Tectonic History and the Biogeography of the Freshwater Fishes from the Coastal Drainages of Eastern Brazil: An Example of Faunal Evolution Associated with a Divergent Continental Margin. Neotrop. Ichthyol. 2006, 4, 225–246. [Google Scholar] [CrossRef]
- Tagliacollo, V.A.; Roxo, F.F.; Duke-Sylvester, S.M.; Oliveira, C.; Albert, J.S. Biogeographical Signature of River Capture Events in Amazonian Lowlands. J. Biogeogr. 2015, 42, 2349–2362. [Google Scholar] [CrossRef]
- Abreu, J.M.S.; Saraiva, A.C.S.; Albert, J.S.; Piorski, N.M. Paleogeographic Influences on Freshwater Fish Distributions in Northeastern Brazil. J. S. Am. Earth Sci. 2020, 102, 102692. [Google Scholar] [CrossRef]
- Oyakawa, O.T. Revisao Sistematica Das Especies Do Genero Hoplias (Grupo Lacerdae) Da Amazonia Brasileira e Regiao Leste Do Brasil (Teleostei, Erythrinidae). Master’s Thesis, University De São Paulo, São Paolo, Brazil, 1990. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Locality | N |
|---|---|---|
| Hoplias australis | Jacutinga river, Arabutã—SC (27°10′11.1″ S 52°05′08.1″ W) | 07♀, 04♂ |
| H. curupira | Parauapebas river, Canaã dos Carajás—PA (6°30′06.5″ S 50°02′35.3″ W) | 05♀, 03♂ |
| H. aimara | Xingu river, Querência—MT (12°35’10.1″ S 52°56’35.6″ W) | 01♀, 03♂ |
| H. intermedius | Cipó river, Santana de Pirapama—MG (18°53’52.5″ S 43°52’58.7″ W) | 04♀, 04♂ |
| H. brasiliensis | Paraguaçu river, Itaberaba—BA (12°44’56.8″ S 40°12’16.0″ W) | 02♀, 08♂ |
| H. lacerdae | Juquiá river, Registro—SP (24°24’01.6″ S 47°49’46.8″ W) | 06♀, 02♂ |
| Species | N | AR | PA | HO | HE | S |
|---|---|---|---|---|---|---|
| H. curupira | 3 | 1.006 | 652 | 0.003 | 0.002 | 1.004 |
| H. australis | 6 | 1.048 | 270 | 0.016 | 0.014 | 1.030 |
| H. aimara | 3 | 1.005 | 1512 | 0.003 | 0.002 | 1.004 |
| H. intermedius | 6 | 1.024 | 64 | 0.009 | 0.009 | 1.018 |
| H. brasiliensis | 6 | 1.049 | 130 | 0.017 | 0.015 | 1.031 |
| H. lacerdae | 5 | 1.011 | 184 | 0.005 | 0.004 | 1.008 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de M. C. Sassi, F.; Perez, M.F.; Oliveira, V.C.S.; Deon, G.A.; de Souza, F.H.S.; Ferreira, P.H.N.; de Oliveira, E.A.; Hatanaka, T.; Liehr, T.; Bertollo, L.A.C.; et al. High Genetic Diversity despite Conserved Karyotype Organization in the Giant Trahiras from Genus Hoplias (Characiformes, Erythrinidae). Genes 2021, 12, 252. https://doi.org/10.3390/genes12020252
de M. C. Sassi F, Perez MF, Oliveira VCS, Deon GA, de Souza FHS, Ferreira PHN, de Oliveira EA, Hatanaka T, Liehr T, Bertollo LAC, et al. High Genetic Diversity despite Conserved Karyotype Organization in the Giant Trahiras from Genus Hoplias (Characiformes, Erythrinidae). Genes. 2021; 12(2):252. https://doi.org/10.3390/genes12020252
Chicago/Turabian Stylede M. C. Sassi, Francisco, Manolo F. Perez, Vanessa Cristina S. Oliveira, Geize A. Deon, Fernando H. S. de Souza, Pedro H. N. Ferreira, Ezequiel A. de Oliveira, Terumi Hatanaka, Thomas Liehr, Luiz A. C. Bertollo, and et al. 2021. "High Genetic Diversity despite Conserved Karyotype Organization in the Giant Trahiras from Genus Hoplias (Characiformes, Erythrinidae)" Genes 12, no. 2: 252. https://doi.org/10.3390/genes12020252
APA Stylede M. C. Sassi, F., Perez, M. F., Oliveira, V. C. S., Deon, G. A., de Souza, F. H. S., Ferreira, P. H. N., de Oliveira, E. A., Hatanaka, T., Liehr, T., Bertollo, L. A. C., & de B. Cioffi, M. (2021). High Genetic Diversity despite Conserved Karyotype Organization in the Giant Trahiras from Genus Hoplias (Characiformes, Erythrinidae). Genes, 12(2), 252. https://doi.org/10.3390/genes12020252