Heat Stress Affects H3K9me3 Level at Human Alpha Satellite DNA Repeats

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatic Analysis
2.2. Human Cell Lines
2.3. DNA Extraction and PCR Analysis in Cell Cultures
2.4. RNA Isolation and Reverse Transcription
2.5. Quantitative Real-Time PCR (qPCR) Analysis
2.6. Chromatin Immunoprecipitation
3. Results
3.1. Dispersion Profile and Polymorphism of Dispersed Alpha Satellite Repeats
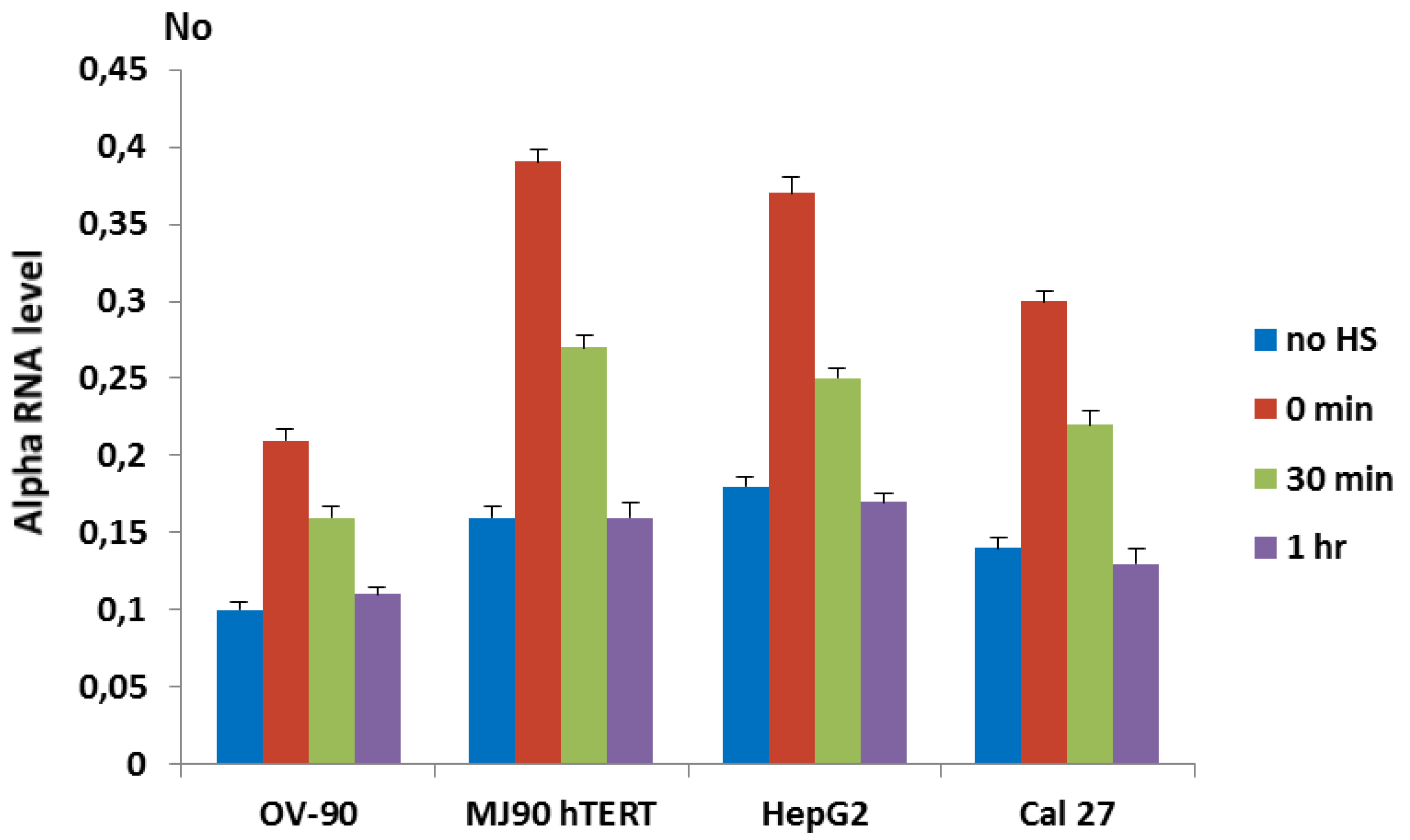
3.2. Alpha Satellite DNA Transcription after HS
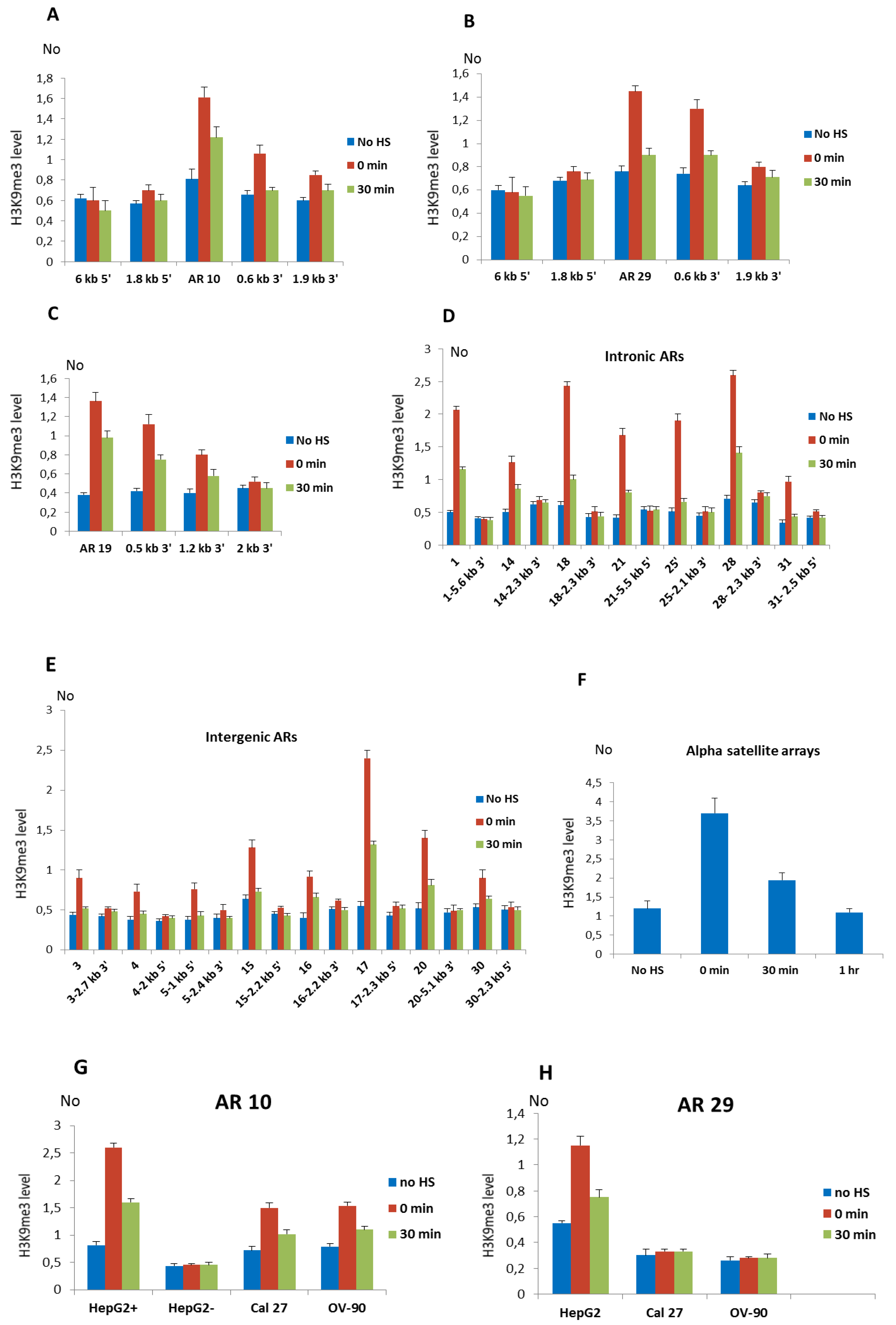
3.3. Dynamics of H3K9me3 at Alpha Satellite Repeats after HS
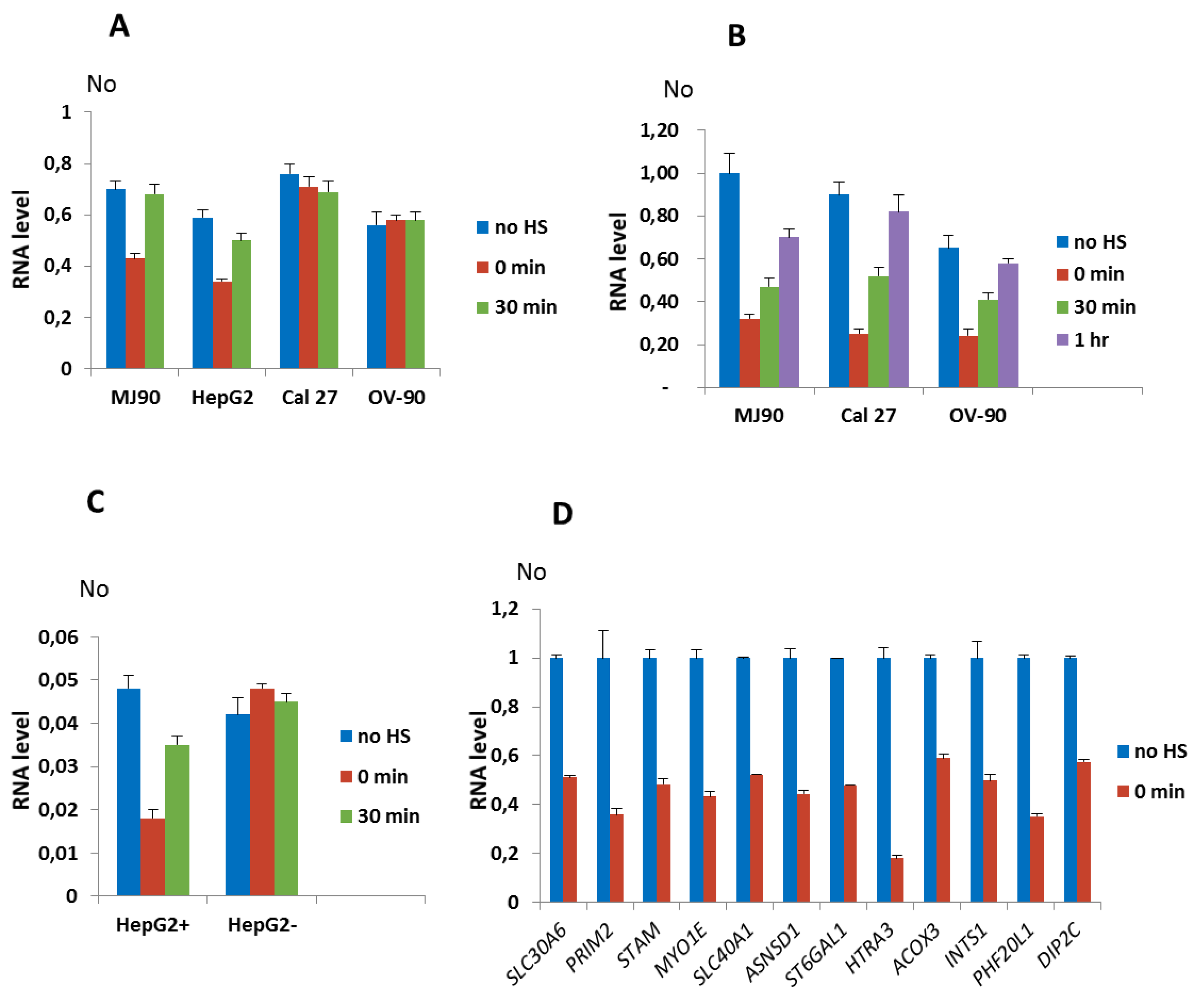
3.4. Expression of Alpha Repeat-Associated Genes after HS
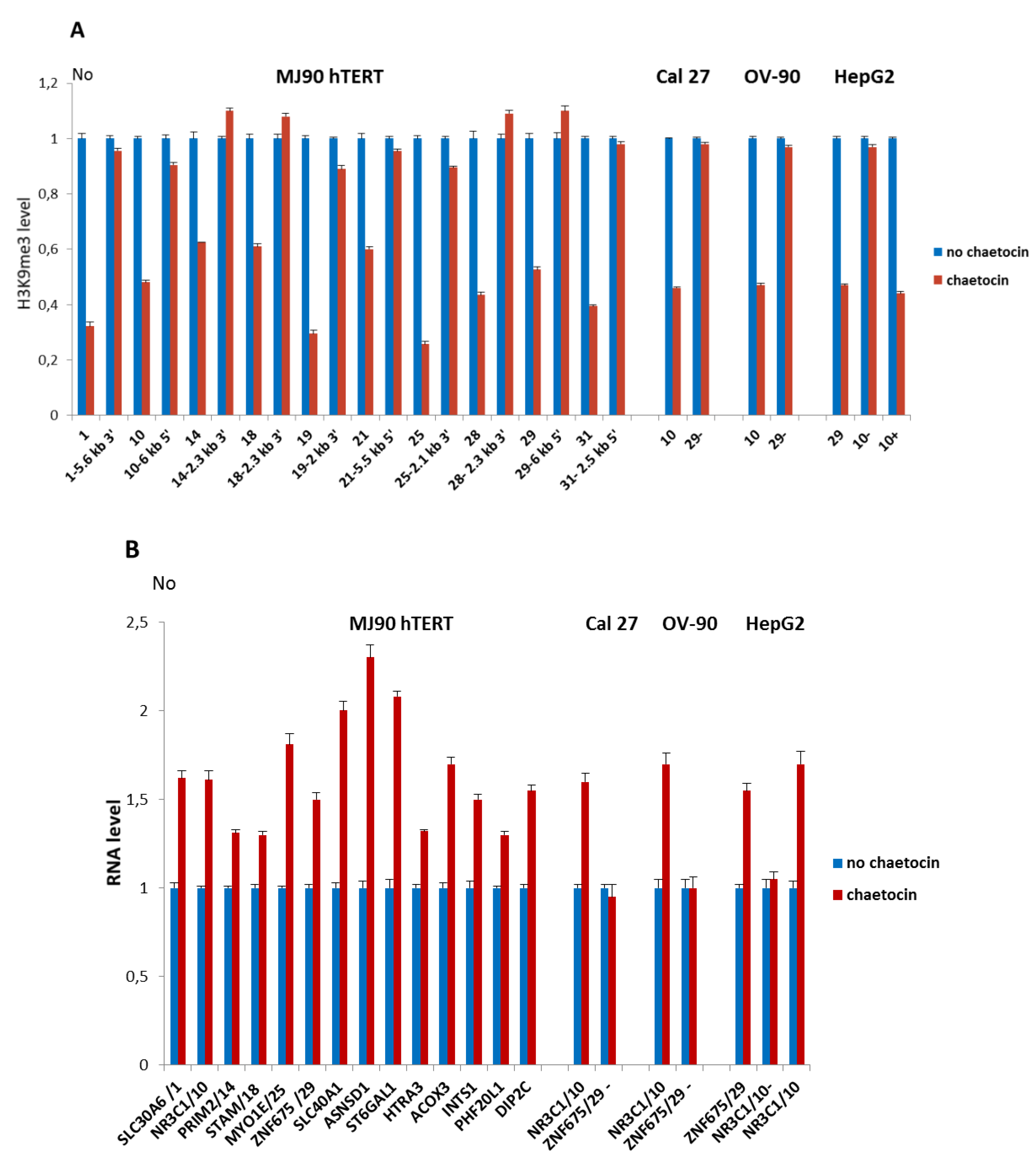
3.5. Influence of Suv39H1 Inhibition on Expression of Alpha Repeat-Associated Genes after HS
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Garrido-Ramos, M. Satellite DNA: An Evolving Topic. Genes 2017, 8, 230. [Google Scholar] [CrossRef] [PubMed]
- Brajković, J.; Feliciello, I.; Bruvo-Mađarić, B.; Ugarković, Đ. Satellite DNA-Like Elements Associated with Genes Within Euchromatin of the Beetle Tribolium castaneum. G3 Genes/Genomes/Genetics 2012, 2, 931–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feliciello, I.; Akrap, I.; Brajković, J.; Zlatar, I.; Ugarković, Đ. Satellite DNA as a driver of population divergence in the red flour beetle Tribolium castaneum. Genome Biol. Evol. 2014, 7, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, G.C.; Küttler, H.; Moreira-Filho, O.; Heslop-Harrison, J.S. The 1.688 Repetitive DNA of Drosophila: Concerted Evolution at Different Genomic Scales and Association with Genes. Mol. Biol. Evol. 2011, 29, 7–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larracuente, A.M. The organization and evolution of the Responder satellite in species of the Drosophila melanogaster group: Dynamic evolution of a target of meiotic drive. BMC Evol. Biol. 2014, 14, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Ruano, F.J.; López-León, M.D.; Cabrero, J.; Camacho, J.P.M. High-throughput analysis of the satellitome illuminates satellite DNA evolution. Sci. Rep. 2016, 6, 28333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ugarković, D. Functional elements residing within satellite DNAs. EMBO Rep. 2005, 6, 1035–1039. [Google Scholar] [CrossRef]
- Ayoub, N.; Goldshmidt, I.; Cohen, A. Position effect variegation at the mating-type locus of fission yeast: A cis-acting element inhibits covariegated expression of genes in the silent and expressed domains. Genetics 1999, 152, 495–508. [Google Scholar]
- Wang, J.; Jia, S.T.; Jia, S. New Insights into the Regulation of Heterochromatin. Trends Genet. 2016, 32, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Col, E.; Hoghoughi, N.; Dufour, S.; Penin, J.; Koskas, S.; Faure, V.; Ouzounova, M.; Hernandez-Vargash, H.; Reynoird, N.; Daujat, S.; et al. Bromodomain factors of BET family are new essential actors of pericentric heterochromatin transcriptional activation in response to heat shock. Sci. Rep. 2017, 7, 5418. [Google Scholar] [CrossRef]
- Jolly, C.; Metz, A.; Govin, J.; Vigneron, M.; Turner, B.M.; Khochbin, S.; Vourc, H.C. Stress-induced transcription of satellite III repeats. J. Cell Biol. 2003, 164, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzi, N.; Denegri, M.; Chiodi, I.; Corioni, M.; Valgardsdottir, R.; Cobianchi, F.; Riva, S.; Biamonti, G. Transcriptional Activation of a Constitutive Heterochromatic Domain of the Human Genome in Response to Heat Shock. Mol. Biol. Cell 2004, 15, 543–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seong, K.-H.; Li, D.; Shimizu, H.; Nakamura, R.; Ishii, S. Inheritance of Stress-Induced, ATF-2-Dependent Epigenetic Change. Cell 2011, 145, 1049–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezer, Z.; Ugarkovic, D. Satellite DNA-associated siRNAs as mediators of heat shock response in insects. RNA Biol. 2012, 9, 587–595. [Google Scholar] [CrossRef] [Green Version]
- Volpe, T.A.; Kidner, C.; Hall, I.M.; Teng, G.; Grewal, S.I.S.; Martienssen, R.A. Regulation of Heterochromatic Silencing and Histone H3 Lysine-9 Methylation by RNAi. Science 2002, 297, 1833–1837. [Google Scholar] [CrossRef] [Green Version]
- Grewal, S.I.S.; Elgin, S.C.R. Transcription and RNA interference in the formation of heterochromatin. Nature 2007, 447, 399–406. [Google Scholar] [CrossRef]
- Fagegaltier, D.; Bougé, A.-L.; Berry, B.; Poisot, É.; Sismeiro, O.; Coppée, J.-Y.; Théodore, L.; Voinnet, O.; Antoniewski, C. The endogenous siRNA pathway is involved in heterochromatin formation in Drosophila. Proc. Natl. Acad. Sci. USA 2009, 106, 21258–21263. [Google Scholar] [CrossRef] [Green Version]
- Feliciello, I.; Akrap, I.; Ugarković, Đ. Satellite DNA Modulates Gene Expression in the Beetle Tribolium castaneum after Heat Stress. PLoS Genet. 2015, 11, e1005466. [Google Scholar]
- McNulty, S.M.; Sullivan, B.A. Alpha satellite DNA biology: Finding function in the recesses of the genome. Chromosom. Res. 2018, 26, 115–138. [Google Scholar] [CrossRef]
- Lee, C.; Wevrick, R.; Fisher, R.B.; Ferguson-Smith, M.A.; Lin, C.C. Human centromeric DNAs. Hum. Genet. 1997, 100, 291–304. [Google Scholar] [CrossRef]
- Rudd, M.K.; Willard, H.F. Analysis of the centromeric regions of the human genome assembly. Trends Genet. 2004, 20, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.L.; Yewdell, W.T.; Bell, J.; McNulty, S.M.; Duda, Z.; O′Neill, R.J.; Sullivan, B.A.; Straight, A. RNA-dependent stabilization of SUV39H1 at constitutive heterochromatin. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Aerts, J.L.; Gonzales, M.I.; Topalian, S.L. Selection of appropriate control genes to assess expression of tumor antigens using real-time RT-PCR. BioTechniques 2004, 36, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.H.; Karlen, Y.; Bakker, O.; Hoff, M.J.B.V.D.; Moorman, A.F.M. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruijter, J.M.; Pfaffl, M.W.; Zhao, S.; Spiess, A.N.; Boggy, G.; Blom, J.; Rutledge, R.G.; Sisti, D.; Lievens, A.; de Preter, K.; et al. Evaluation of qPCR curve analysis methods for reliable biomarker discovery: Bias, resolution, precision, and implications. Methods 2013, 59, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Choo, K.; Vissel, B.; Nagy, A.; Earle, E.; Kalitsis, P. A survey of the genomic distribution of alpha satellite DNA on all the human chromosomes, and derivation of a new consensus sequence. Nucleic Acids Res. 1991, 19, 1179–1182. [Google Scholar] [CrossRef] [Green Version]
- Greiner, D.; Bonaldi, T.; Eskeland, R.; Roemer, E.; Imhof, A. Identification of a specific inhibitor of the histone methyltransferase SU(VAR)3-9. Nat. Methods 2005, 1, 143–145. [Google Scholar] [CrossRef]
- Imai, K.; Kamio, N.; Cueno, M.E.; Saito, Y.; Inoue, H.; Saito, I.; Ochiai, K. Role of the histone H3 lysine 9 methyltransferase Suv39 h1 in maintaining Epsteinn-Barr virus latency in B95-8 cells. FEBS J. 2014, 281, 2148–2158. [Google Scholar] [CrossRef]
- Pecinka, A.; Dinh, H.Q.; Baubec, T.; Rosa, M.; Lettner, N.; Scheid, O.M. Epigenetic regulation of repetitive elements is attenuated by prolonged heat stress in Arabidopsis. Plant Cell 2010, 22, 3118–3129. [Google Scholar] [CrossRef] [Green Version]
- Tittel-Elmer, M.; Bucher, E.; Broger, L.; Mathieu, O.; Paszkowski, J.; Vaillant, I. Stress-Induced Activation of Heterochromatic Transcription. PLoS Genet. 2010, 6, e1001175. [Google Scholar] [CrossRef]
- Brajković, J.; Pezer, Ž.; Bruvo-Mađarić, B.; Sermek, A.; Feliciello, I.; Ugarković, Đ. Dispersion Profiles and Gene Associations of Repetitive DNAs in the Euchromatin of the Beetle Tribolium castaneum. G3 Genes/Genomes/Genetics 2018, 8, 875–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eymery, A.; Horard, B.; el Atifi-Borel, M.; Fourel, G.; Berger, F.; Vitte, A.-L.; Broeck, A.V.D.; Brambilla, E.; Fournier, A.; Callanan, M.; et al. A transcriptomic analysis of human centromeric and pericentric sequences in normal and tumor cells. Nucleic Acids Res. 2009, 37, 6340–6354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ninomiya, K.; Adachi, S.; Natsume, T.; Iwakiri, J.; Terai, G.; Asai, K.; Hirose, T. LncRNA-dependent nuclear stress bodies promote intron retention through SR protein phosphorylation. EMBO J. 2019, 39, e102729. [Google Scholar] [CrossRef] [PubMed]
- Saksouk, N.; Simboeck, E.; Dejardin, J. Constitutive heterochromatin formation and transcription in mammals. Epigenetics Chromatin 2015, 8, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, L.H.; Brettingham-Moore, K.; Chan, L.; Quach, J.M.; Anderson, M.A.; Northrop, E.L.; Hannan, R.D.; Saffery, R.; Shaw, M.L.; Williams, E.; et al. Centromere RNA is a key component for the assembly of nucleoproteins at the nucleolus and centromere. Genome Res. 2007, 17, 1146–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezer, Ž.; Ugarković, Đ. RNA Pol II Promotes Transcription of Centromeric Satellite DNA in Beetles. PLoS ONE 2008, 3, e1594. [Google Scholar] [CrossRef] [Green Version]
- Hedouin, S.; Grillo, G.; Ivkovic, I.; Velasco, G.; Francastel, C. CENP-A chromatin disassembly in stressed and senescent murine cells. Sci. Rep. 2017, 7, 42520. [Google Scholar] [CrossRef] [Green Version]
- Eichten, S.R.; Ellis, N.; Makarevitch, I.; Yeh, C.-T.; Gent, J.I.; Guo, L.; McGinnis, K.M.; Zhang, X.; Schnable, P.S.; Vaughn, M.W.; et al. Spreading of Heterochromatin Is Limited to Specific Families of Maize Retrotransposons. PLoS Genet. 2012, 8, e1003127. [Google Scholar] [CrossRef]
- Sienski, G.; Dönertas, D.; Brennecke, J. Transcriptional Silencing of Transposons by Piwi and Maelstrom and Its Impact on Chromatin State and Gene Expression. Cell 2012, 151, 964–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.C.G.; Karpen, G.H. Pervasive epigenetic effects of Drosophila euchromatic transposable elements impact their evolution. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebollo, R.; Karimi, M.M.; Bilenky, M.; Gagnier, L.; Miceli-Royer, K.; Zhang, Y.; Goyal, P.; Keane, T.; Jones, S.J.M.; Hirst, M.; et al. Retrotransposon-Induced Heterochromatin Spreading in the Mouse Revealed by Insertional Polymorphisms. PLoS Genet. 2011, 7, e1002301. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Lee, C.H.; Swigut, T.; Grow, E.; Gu, B.; Bassik, M.C.; Wysocka, J. Selective silencing of euchromatic L1s revealed by genome-wide screens for L1 regulators. Nature 2018, 553, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Mahat, D.B.; Salamanca, H.H.; Duarte, F.M.; Danko, C.G.; Lis, J.T. Mammalian Heat Shock Response and Mechanisms Underlying Its Genome-wide Transcriptional Regulation. Mol. Cell 2016, 62, 63–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadmiel, M.; Cidlowski, J.A. Glucocorticoid receptor signaling in health and disease. Trends Pharmacol. Sci. 2013, 34, 518–530. [Google Scholar] [CrossRef] [Green Version]
- Narkiewicz, J.; Lapinska-Szumczyk, S.; Zurawa-Janicka, D.; Skorko-Glonek, J.; Emerich, J.; Lipinska, B. Expression of human HtrA1, HtrA2, HtrA3 and TGF-beta1 genes in primary endometrial cancer. Oncol. Rep. 2009, 21, 1529–1537. [Google Scholar] [PubMed]
- Zhao, J.; Zhang, J.; Zhang, X.; Feng, M.; Qu, J. High temperature requirement A3 (HTRA3) expression predicts postoperative recurrence and survival in patients with non-small-cell lung cancer. Oncotarget 2016, 7, 40725–40734. [Google Scholar] [CrossRef] [Green Version]
- Razanadrakoto, L.; Cormier, F.; Laurienté, V.; Dondi, E.; Gardano, L.; Katzav, S.; Guittat, L.; Varin-Blank, N. Mutation of Vav1 adaptor region reveals a new oncogenic activation. Oncotarget 2014, 6, 2524–2538. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Liu, L.; Shan, W.; Yang, Z. An integrated genomic analysis of Tudor domain–containing proteins identifies PHD finger protein 20-like 1 (PHF20L1) as a candidate oncogene in breast cancer. Mol. Oncol. 2015, 10, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Imbeault, M.; Helleboid, P.-Y.; Trono, D. KRAB zinc-finger proteins contribute to the evolution of gene regulatory networks. Nature 2017, 543, 550–554. [Google Scholar] [CrossRef]
- Feliciello, I.; Picariello, O.; Chinali, G. Intra-specific variability and unusual organization of the repetitive units in a satellite DNA from Rana dalmatina: Molecular evidence of a new mechanism of DNA repair acting on satellite DNA. Gene 2006, 383, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Sproul, J.S.; Khost, D.E.; Eickbush, D.G.; Negm, S.; Wei, X.; Wong, I.; Larracuente, A.M. Dynamic Evolution of Euchromatic Satellites on the X Chromosome in Drosophila melanogaster and the simulans Clade. Mol. Biol. Evol. 2020, 078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, H.S.; Kudoh, J.; Wattenhofer, M.; Shibuya, K.; Berry, A.; Chrast, R.; Guipponi, M.; Wang, J.; Kawasaki, K.; Asakawa, S.; et al. Insertion of beta-satellite repeats identifies a transmembrane protease causing both congenital and childhood onset autosomal recessive deafness. Nat. Genet. 2001, 27, 59–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedges, D.J.; Deininger, P.L. Inviting instability: Transposable elements, double-strand breaks, and the maintenance of genome integrity. Mutat. Res. Mol. Mech. Mutagen. 2007, 616, 46–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynn, E.K.; Kamat, A.; Lach, F.P.; Donovan, F.X.; Kimble, D.C.; Narisu, N.; Sanborn, E.; Boulad, F.; Davies, S.M.; Gillio, A.P.; et al. Comprehensive Analysis of Pathogenic Deletion Variants in Fanconi Anemia Genes. Hum. Mutat. 2014, 35, 1342–1353. [Google Scholar] [CrossRef] [Green Version]
- Halbach, R.; Miesen, P.; Joosten, J.; Taşköprü, E.; Rondeel, I.; Pennings, B.; Vogels, C.B.F.; Merkling, S.H.; Koenraadt, C.J.; Lambrechts, L.; et al. A satellite repeat-derived piRNA controls embryonic development of Aedes. Nature 2020, 580, 274–277. [Google Scholar] [CrossRef]
- Ugarković, Ð.; Plohl, M. Variation in satellite DNA profiles—Causes and effects. EMBO J. 2002, 21, 5955–5959. [Google Scholar] [CrossRef] [Green Version]
- Feliciello, I.; Picariello, O.; Chinali, G. The first characterisation of the overall variability of repetitive units in a species reveals unexpected features of satellite DNA. Gene 2005, 349, 153–164. [Google Scholar] [CrossRef]
- Satyaki, P.R.; Cuykendall, T.N.; Wei, K.H.; Brideau, N.J.; Kwak, H.; Aruna, S.; Ferree, P.M.; Ji, S.; Barbash, D.A. The Hmr and Lhr hybrid incompatibility genes suppress a broad range of heterochroma. PLoS Genet. 2014, 10, e1004240. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alpha Repeat No /Number of Monomers / Similarity to Consensus % | The Increase of H3K9me3 (×) after 2 h HS and p Values | Alpha Repeat-Associated Gene | Cell Line | Position of Alpha Repeat Relative to Gene and Distance (bp) | Distance of Alpha Repeat from TSS (bp) | The Gene Expression downregulation (×) after1 h/2 h HS and p Values |
|---|---|---|---|---|---|---|
| 29/0 | 1.0 (0.211) | ZNF675 | Cal 27 | / | / | -/1.0 (0.059) |
| 29/0 | 1.0 (0.162) | ZNF675 | OV-90 | / | / | -/1.0 (0.087) |
| 29/0.6 / 88% | 1.9 (0.012) | ZNF675 | MJ90 hTERT | intron | 25,978 | -/1.7 (0.002) |
| 29/0.6 / 88% | 2.1 (0.008) | ZNF675 | HepG2 | intron | 25,978 | -/1.7 (0.003) |
| 10/1.1 / 86% | 2.0 (0.011) | NR3C1 | MJ90 hTERT | intron | 125,971 | -/3.5 (0.002) |
| 10/1.1 / 86% | 3.2 (0.014) | NR3C1+allele | HepG2 | intron | 125,971 | -/2.7 (0.004) |
| 10/0 | 1.0 (0.158) | NR3C1-allele | HepG2 | / | / | -/1.0 (0.061) |
| 10/1.1 / 86% | 2.1 (0.012) | NR3C1 | Cal 27 | intron | 125,971 | -/3.7 (0.007) |
| 10/1.1 / 86% | 1.7 (0.017) | NR3C1 | OV-90 | intron | 125,971 | -/2.7 (0.012) |
| 28/1.2 / 70% | 3.7 (0.006) | VAV1 | MJ90 hTERT | intron | 25,978 | / |
| 28/1.2 / 70% | - | VAV1 | 697 | intron | 46,492 | 2.6/- (0.001) |
| 1/0.7 / 82% | 4.1 (0.009) | SLC30A6 | MJ90 hTERT | intron | 48,094 | -/2.0 (0.003) |
| 14/1.7 / 70% | 2.5 (0.011) | PRIM2 | MJ90 hTERT | intron | 59,161 | -/2.8 (0.015) |
| 18/0.5 / 76% | 4.3 (0.012) | STAM | MJ90 hTERT | intron | 9163 | -/2.0 (0.004) |
| 19/2.4 / 74% | 3.6 (0.006) | PLA2G12B | MJ90 hTERT | intron | 15,104 | / |
| 25/1.2 / 88% | 3.9 (0.018) | MYO1E | MJ90 hTERT | intron | 152,899 | -/2.3 (0.005) |
| 21/0.7 / 85% | 4.0 (0.005) | DLG2 | MJ90 hTERT | intron | 197,351 | / |
| 31/1.3 / 71% | 2.8 (0.011) | MAP7 | MJ90 hTERT | intron | 15,483 | / |
| 3/1.4 / 77% | 2.1 (0.012) | SLC40A1 | MJ90 hTERT | 5′ / 38,888 | 38,888 | -/2.0 (0.003) |
| 3/1.4 / 77% | - | ASNSD1 | MJ90 hTERT | 5′ / 46,190 | 46,190 | -/2.3 (0.014) |
| 4/1.4 / 70% | 1.9 (0.014) | ST6GAL1 | MJ90 hTERT | 3′ / 29,520 | 174,981 | -/2.1 (0.001) |
| 5/3.4 / 75% | 1.8 (0.012) | HTRA3 | MJ90 hTERT | 3′ / 34,942 | 72,296 | -/5.6 (0.011) |
| 5/3.4 / 75% | - | ACOX3 | MJ90 hTERT | 3′ / 24,328 | 98,092 | -/1.7 (0.009) |
| 15/2.0 / 86% | 2.0 (0.011) | INTS1 | MJ90 hTERT | 5′ / 14,580 | 14,850 | -/2.0 (0.013) |
| 16/0.5 / 72% | 2.3 (0.018) | PHF20L1 | MJ90 hTERT | 3′ / 15,772 | 89,221 | -/2.9 (0.010) |
| 17/0.5 / 94% | 4.1 (0.011) | DIP2C | MJ90 hTERT | 5′ / 18,158 | 18,158 | -/1.8 (0.010) |
| 20/1.0 / 86% | 2.4 (0.014) | OR6A2 | MJ90 hTERT | 5′/ 14,531 | 14,531 | / |
| 30/1.0 / 70% | 1.7 (0.012) | PPP2R3B | MJ90 hTERT | 5′ / 16,060 | 16,060 | / |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feliciello, I.; Sermek, A.; Pezer, Ž.; Matulić, M.; Ugarković, Đ. Heat Stress Affects H3K9me3 Level at Human Alpha Satellite DNA Repeats. Genes 2020, 11, 663. https://doi.org/10.3390/genes11060663
Feliciello I, Sermek A, Pezer Ž, Matulić M, Ugarković Đ. Heat Stress Affects H3K9me3 Level at Human Alpha Satellite DNA Repeats. Genes. 2020; 11(6):663. https://doi.org/10.3390/genes11060663
Chicago/Turabian StyleFeliciello, Isidoro, Antonio Sermek, Željka Pezer, Maja Matulić, and Đurđica Ugarković. 2020. "Heat Stress Affects H3K9me3 Level at Human Alpha Satellite DNA Repeats" Genes 11, no. 6: 663. https://doi.org/10.3390/genes11060663
APA StyleFeliciello, I., Sermek, A., Pezer, Ž., Matulić, M., & Ugarković, Đ. (2020). Heat Stress Affects H3K9me3 Level at Human Alpha Satellite DNA Repeats. Genes, 11(6), 663. https://doi.org/10.3390/genes11060663