
The Role of Structural Variation in Adaptation and Evolution of Yeast and Other Fungi

Abstract
:1. Introduction
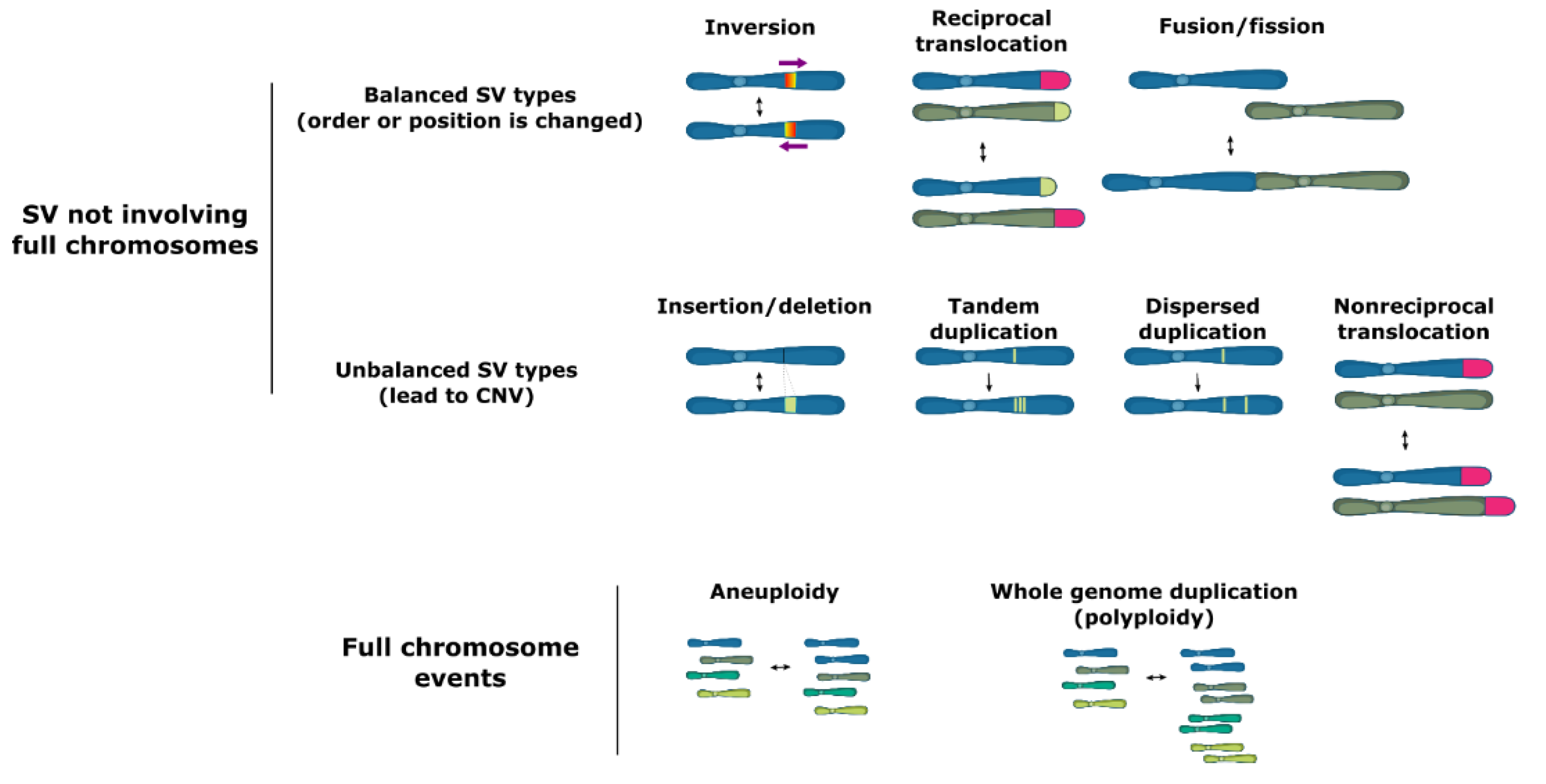
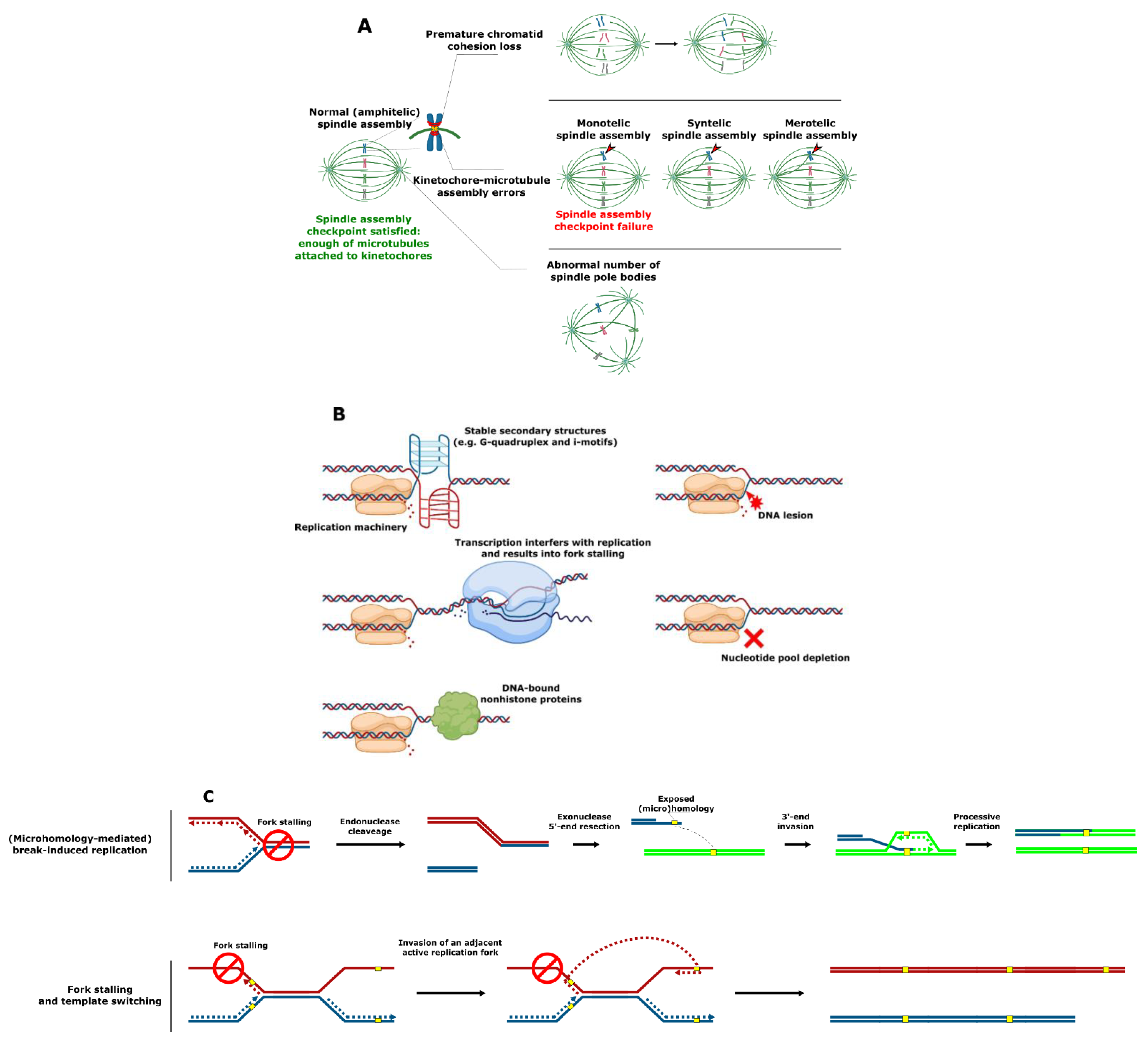
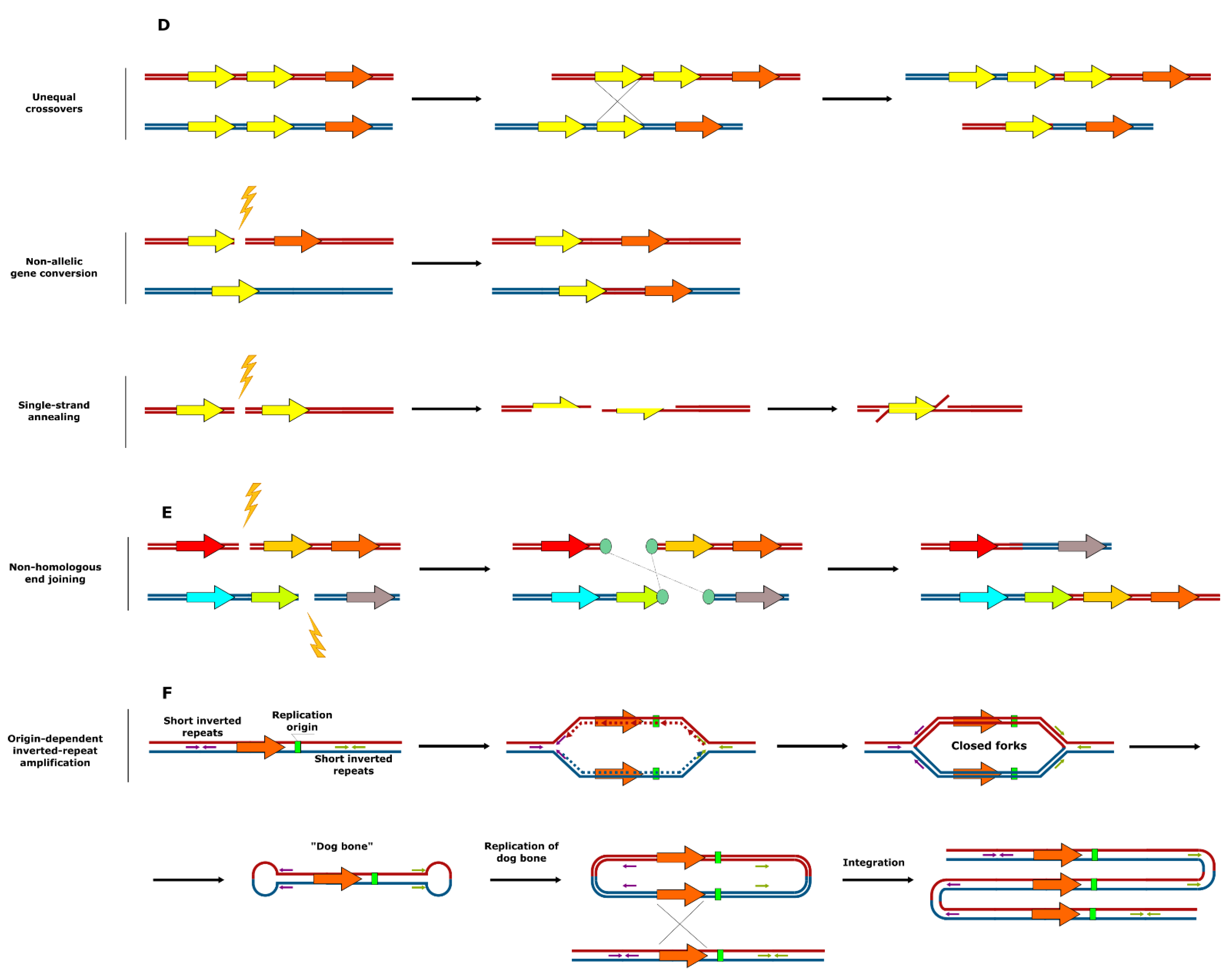
2. Mechanisms of SV Formation
3. Gene Duplications: Fruitful Soil for Evolution and Adaptation
4. Whole Genome Duplication
5. Aneuploidy and Segmental Duplications: A “Quick Fix” for Evolutionary Adaptation?
6. Balanced SV Events
7. Examples of Adaptive SV Events
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stewart, N.B.; Rogers, R.L. Chromosomal rearrangements as a source of new gene formation in Drosophila yakuba. PLoS Genet. 2019, 15, e1008314. [Google Scholar] [CrossRef]
- Huang, Y.C.; Dang, V.D.; Chang, N.C.; Wang, J. Multiple large inversions and breakpoint rewiring of gene expression in the evolution of the fire ant social supergene. Proc. Biol. Sci. 2018, 285. [Google Scholar] [CrossRef]
- Lavington, E.; Kern, A.D. The Effect of Common Inversion Polymorphisms on Patterns of Transcriptional Variation in Drosophila melanogaster. G3 Genes Genomes Genet. 2017, 7, 3659. [Google Scholar] [CrossRef] [Green Version]
- Gamazon, E.R.; Stranger, B.E. The impact of human copy number variation on gene expression. Brief. Funct. Genom. 2015, 14, 352–357. [Google Scholar] [CrossRef] [Green Version]
- Radke, D.W.; Lee, C. Adaptive potential of genomic structural variation in human and mammalian evolution. Brief. Funct. Genom. 2015, 14, 358–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, M.J.; Jiggins, C.D. Supergenes and their role in evolution. Heredity 2014, 113, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avril, A.; Purcell, J.; Brelsford, A.; Chapuisat, M. Asymmetric assortative mating and queen polyandry are linked to a supergene controlling ant social organization. Mol. Ecol. 2019, 28, 1428–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coughlan, J.M.; Willis, J.H. Dissecting the role of a large chromosomal inversion in life history divergence throughout the Mimulus guttatus species complex. Mol. Ecol. 2019, 28, 1343–1357. [Google Scholar] [CrossRef] [PubMed]
- Faria, R.; Chaube, P.; Morales, H.E.; Larsson, T.; Lemmon, A.R.; Lemmon, E.M.; Rafajlović, M.; Panova, M.; Ravinet, M.; Johannesson, K.; et al. Multiple chromosomal rearrangements in a hybrid zone between Littorina saxatilis ecotypes. Mol. Ecol. 2019, 28, 1375–1393. [Google Scholar] [CrossRef] [Green Version]
- Wellband, K.; Mérot, C.; Linnansaari, T.; Elliott, J.A.K.; Curry, R.A.; Bernatchez, L. Chromosomal fusion and life history-associated genomic variation contribute to within-river local adaptation of Atlantic salmon. Mol. Ecol. 2019, 28, 1439–1459. [Google Scholar] [CrossRef]
- Wellenreuther, M.; Mérot, C.; Berdan, E.; Bernatchez, L. Going beyond SNPs: The role of structural genomic variants in adaptive evolution and species diversification. Mol. Ecol. 2019, 28, 1203–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrad, D.F.; Pinto, D.; Redon, R.; Feuk, L.; Gokcumen, O.; Zhang, Y.; Aerts, J.; Andrews, T.D.; Barnes, C.; Campbell, P.; et al. Origins and functional impact of copy number variation in the human genome. Nature 2010, 464, 704–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudmant, P.H.; Rausch, T.; Gardner, E.J.; Handsaker, R.E.; Abyzov, A.; Huddleston, J.; Zhang, Y.; Ye, K.; Jun, G.; Fritz, M.H.; et al. An integrated map of structural variation in 2504 human genomes. Nature 2015, 526, 75–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffares, D.C.; Jolly, C.; Hoti, M.; Speed, D.; Shaw, L.; Rallis, C.; Balloux, F.; Dessimoz, C.; Bähler, J.; Sedlazeck, F.J. Transient structural variations have strong effects on quantitative traits and reproductive isolation in fission yeast. Nat. Commun. 2017, 8, 14061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audano, P.A.; Sulovari, A.; Graves-Lindsay, T.A.; Cantsilieris, S.; Sorensen, M.; Welch, A.E.; Dougherty, M.L.; Nelson, B.J.; Shah, A.; Dutcher, S.K.; et al. Characterizing the Major Structural Variant Alleles of the Human Genome. Cell 2019, 176, 663–675.e619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weischenfeldt, J.; Symmons, O.; Spitz, F.; Korbel, J.O. Phenotypic impact of genomic structural variation: Insights from and for human disease. Nat. Rev. Genet. 2013, 14, 125–138. [Google Scholar] [CrossRef]
- Hollox, E.J.; Huffmeier, U.; Zeeuwen, P.L.J.M.; Palla, R.; Lascorz, J.; Rodijk-Olthuis, D.; van de Kerkhof, P.C.M.; Traupe, H.; de Jongh, G.; Heijer, M.D.; et al. Psoriasis is associated with increased β-defensin genomic copy number. Nat. Genet. 2008, 40, 23–25. [Google Scholar] [CrossRef] [Green Version]
- Stefansson, H.; Rujescu, D.; Cichon, S.; Pietilainen, O.P.; Ingason, A.; Steinberg, S.; Fossdal, R.; Sigurdsson, E.; Sigmundsson, T.; Buizer-Voskamp, J.E.; et al. Large recurrent microdeletions associated with schizophrenia. Nature 2008, 455, 232–236. [Google Scholar] [CrossRef]
- Traherne, J.A.; Martin, M.; Ward, R.; Ohashi, M.; Pellett, F.; Gladman, D.; Middleton, D.; Carrington, M.; Trowsdale, J. Mechanisms of copy number variation and hybrid gene formation in the KIR immune gene complex. Hum. Mol. Genet. 2010, 19, 737–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polley, S.; Louzada, S.; Forni, D.; Sironi, M.; Balaskas, T.; Hains, D.S.; Yang, F.; Hollox, E.J. Evolution of the rapidly mutating human salivary agglutinin gene (DMBT1) and population subsistence strategy. Proc. Natl. Acad. Sci. USA 2015, 112, 5105–5110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durbin, R.M.; Altshuler, D.; Durbin, R.M.; Abecasis, G.R.; Bentley, D.R.; Chakravarti, A.; Clark, A.G.; Collins, F.S.; De La Vega, F.M.; Donnelly, P.; et al. A map of human genome variation from population-scale sequencing. Nature 2010, 467, 1061–1073. [Google Scholar] [CrossRef] [Green Version]
- Weissensteiner, M.H.; Bunikis, I.; Catalan, A.; Francoijs, K.J.; Knief, U.; Heim, W.; Peona, V.; Pophaly, S.D.; Sedlazeck, F.J.; Suh, A.; et al. Discovery and population genomics of structural variation in a songbird genus. Nat. Commun. 2020, 11, 3403. [Google Scholar] [CrossRef]
- Catanach, A.; Crowhurst, R.; Deng, C.; David, C.; Bernatchez, L.; Wellenreuther, M. The genomic pool of standing structural variation outnumbers single nucleotide polymorphism by threefold in the marine teleost Chrysophrys auratus. Mol. Ecol. 2019, 28, 1210–1223. [Google Scholar] [CrossRef] [PubMed]
- Lucek, K.; Gompert, Z.; Nosil, P. The role of structural genomic variants in population differentiation and ecotype formation in Timema cristinae walking sticks. Mol. Ecol. 2019, 28, 1224–1237. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, M.; Emerson, J.J.; Macdonald, S.J.; Long, A.D. Structural variants exhibit widespread allelic heterogeneity and shape variation in complex traits. Nat. Commun. 2019, 10, 4872. [Google Scholar] [CrossRef] [Green Version]
- Fisher, K.J.; Buskirk, S.W.; Vignogna, R.C.; Marad, D.A.; Lang, G.I. Adaptive genome duplication affects patterns of molecular evolution in Saccharomyces cerevisiae. PLoS Genet. 2018, 14, e1007396. [Google Scholar] [CrossRef] [PubMed]
- Steenwyk, J.; Rokas, A. Extensive Copy Number Variation in Fermentation-Related Genes among Saccharomyces cerevisiae Wine Strains. G3 Genes Genomes Genet. 2017, 7, 1475. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Zhang, L.J.; Fang, Y.H.; Jin, X.N.; Qi, L.; Wu, X.C.; Zheng, D.Q. Genomic structural variation contributes to phenotypic change of industrial bioethanol yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2016, 16, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treu, L.; Toniolo, C.; Nadai, C.; Sardu, A.; Giacomini, A.; Corich, V.; Campanaro, S. The impact of genomic variability on gene expression in environmental Saccharomyces cerevisiae strains. Environ. Microbiol. 2014, 16, 1378–1397. [Google Scholar] [CrossRef]
- Gresham, D.; Desai, M.M.; Tucker, C.M.; Jenq, H.T.; Pai, D.A.; Ward, A.; DeSevo, C.G.; Botstein, D.; Dunham, M.J. The Repertoire and Dynamics of Evolutionary Adaptations to Controlled Nutrient-Limited Environments in Yeast. PLoS Genet. 2008, 4, e1000303. [Google Scholar] [CrossRef] [Green Version]
- Dunham, M.J.; Badrane, H.; Ferea, T.; Adams, J.; Brown, P.O.; Rosenzweig, F.; Botstein, D. Characteristic genome rearrangements in experimental evolution of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2002, 99, 16144. [Google Scholar] [CrossRef] [Green Version]
- Thompson, S.L.; Bakhoum, S.F.; Compton, D.A. Mechanisms of chromosomal instability. Curr. Biol. 2010, 20, R285–R295. [Google Scholar] [CrossRef] [Green Version]
- Myung, K.; Datta, A.; Kolodner, R.D. Suppression of spontaneous chromosomal rearrangements by S phase checkpoint functions in Saccharomyces cerevisiae. Cell 2001, 104, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Labib, K.; Hodgson, B. Replication fork barriers: Pausing for a break or stalling for time? EMBO Rep. 2007, 8, 346–353. [Google Scholar] [CrossRef] [Green Version]
- Admire, A.; Shanks, L.; Danzl, N.; Wang, M.; Weier, U.; Stevens, W.; Hunt, E.; Weinert, T. Cycles of chromosome instability are associated with a fragile site and are increased by defects in DNA replication and checkpoint controls in yeast. Genes Dev. 2006, 20, 159–173. [Google Scholar] [CrossRef] [Green Version]
- Brewer, B.J.; Lockshon, D.; Fangman, W.L. The arrest of replication forks in the rDNA of yeast occurs independently of transcription. Cell 1992, 71, 267–276. [Google Scholar] [CrossRef]
- Paeschke, K.; Bochman, M.L.; Garcia, P.D.; Cejka, P.; Friedman, K.L.; Kowalczykowski, S.C.; Zakian, V.A. Pif1 family helicases suppress genome instability at G-quadruplex motifs. Nature 2013, 497, 458–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallgren, M.; Mohammad, J.B.; Yan, K.-P.; Pourbozorgi-Langroudi, P.; Ebrahimi, M.; Sabouri, N. G-rich telomeric and ribosomal DNA sequences from the fission yeast genome form stable G-quadruplex DNA structures in vitro and are unwound by the Pfh1 DNA helicase. Nucleic Acids Res. 2016, 44, 6213–6231. [Google Scholar] [CrossRef] [Green Version]
- Capra, J.A.; Paeschke, K.; Singh, M.; Zakian, V.A. G-quadruplex DNA sequences are evolutionarily conserved and associated with distinct genomic features in Saccharomyces cerevisiae. PLoS Comput. Biol. 2010, 6, e1000861. [Google Scholar] [CrossRef] [PubMed]
- Hershman, S.G.; Chen, Q.; Lee, J.Y.; Kozak, M.L.; Yue, P.; Wang, L.S.; Johnson, F.B. Genomic distribution and functional analyses of potential G-quadruplex-forming sequences in Saccharomyces cerevisiae. Nucleic Acids Res. 2008, 36, 144–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huppert, J.L.; Balasubramanian, S. Prevalence of quadruplexes in the human genome. Nucleic Acids Res. 2005, 33, 2908–2916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabouri, N.; Capra, J.A.; Zakian, V.A. The essential Schizosaccharomyces pombe Pfh1 DNA helicase promotes fork movement past G-quadruplex motifs to prevent DNA damage. BMC Biol. 2014, 12, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Huppert, J.L.; Balasubramanian, S. G-quadruplexes in promoters throughout the human genome. Nucleic Acids Res. 2007, 35, 406–413. [Google Scholar] [CrossRef]
- Drygin, D.; Siddiqui-Jain, A.; O’Brien, S.; Schwaebe, M.; Lin, A.; Bliesath, J.; Ho, C.B.; Proffitt, C.; Trent, K.; Whitten, J.P. Anticancer activity of CX-3543: A direct inhibitor of rRNA biogenesis. Cancer Res. 2009, 69, 7653–7661. [Google Scholar] [CrossRef] [Green Version]
- Ivessa, A.S.; Lenzmeier, B.A.; Bessler, J.B.; Goudsouzian, L.K.; Schnakenberg, S.L.; Zakian, V.A. The Saccharomyces cerevisiae helicase Rrm3p facilitates replication past nonhistone protein-DNA complexes. Mol. Cell 2003, 12, 1525–1536. [Google Scholar] [CrossRef]
- Lin, Y.-L.; Pasero, P. Interference between DNA replication and transcription as a cause of genomic instability. Curr. Genom. 2012, 13, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, H.; Aguilera, A. Transcription as a threat to genome integrity. Annu. Rev. Biochem. 2016, 85, 291–317. [Google Scholar] [CrossRef]
- Cabral, M.; Cheng, X.; Singh, S.; Ivessa, A.S. Absence of Non-histone Protein Complexes at Natural Chromosomal Pause Sites Results in Reduced Replication Pausing in Aging Yeast Cells. Cell Rep. 2016, 17, 1747–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, S.; Watson, A.; Sheedy, D.M.; Martin, B.; Carr, A.M. Gross chromosomal rearrangements and elevated recombination at an inducible site-specific replication fork barrier. Cell 2005, 121, 689–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bester, A.C.; Roniger, M.; Oren, Y.S.; Im, M.M.; Sarni, D.; Chaoat, M.; Bensimon, A.; Zamir, G.; Shewach, D.S.; Kerem, B. Nucleotide deficiency promotes genomic instability in early stages of cancer development. Cell 2011, 145, 435–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldivar, J.C.; Miuma, S.; Bene, J.; Hosseini, S.A.; Shibata, H.; Sun, J.; Wheeler, L.J.; Mathews, C.K.; Huebner, K. Initiation of genome instability and preneoplastic processes through loss of Fhit expression. PLoS Genet. 2012, 8, e1003077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, S.; Mizuno, K.I.; Blaisonneau, J.; Martineau, S.; Chanet, R.; Fréon, K.; Murray, J.M.; Carr, A.M.; Baldacci, G. Homologous recombination restarts blocked replication forks at the expense of genome rearrangements by template exchange. Mol. Cell 2010, 39, 346–359. [Google Scholar] [CrossRef]
- Mizuno, K.I.; Lambert, S.; Baldacci, G.; Murray, J.M.; Carr, A.M. Nearby inverted repeats fuse to generate acentric and dicentric palindromic chromosomes by a replication template exchange mechanism. Genes Dev. 2009, 23, 2876–2886. [Google Scholar] [CrossRef] [Green Version]
- Payen, C.; Koszul, R.; Dujon, B.; Fischer, G. Segmental duplications arise from Pol32-dependent repair of broken forks through two alternative replication-based mechanisms. PLoS Genet. 2008, 4, e1000175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petes, T.D. Unequal meiotic recombination within tandem arrays of yeast ribosomal DNA genes. Cell 1980, 19, 765–774. [Google Scholar] [CrossRef]
- Szostak, J.W.; Wu, R. Unequal crossing over in the ribosomal DNA of Saccharomyces cerevisiae. Nature 1980, 284, 426–430. [Google Scholar] [CrossRef]
- Welch, J.W.; Maloney, D.H.; Fogel, S. Unequal crossing-over and gene conversion at the amplified CUP1 locus of yeast. Mol. Gen. Genet. MGG 1990, 222, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Gangloff, S.; Zou, H.; Rothstein, R. Gene conversion plays the major role in controlling the stability of large tandem repeats in yeast. EMBO J. 1996, 15, 1715–1725. [Google Scholar] [CrossRef] [PubMed]
- Ozenberger, B.A.; Roeder, G.S. A unique pathway of double-strand break repair operates in tandemly repeated genes. Mol. Cell. Biol. 1991, 11, 1222–1231. [Google Scholar] [CrossRef]
- Fischer, G.; James, S.; Roberts, I.; Oliver, S.; Louis, E. Chromosomal evolution in Saccharomyces. Nature 2000, 405, 451–454. [Google Scholar] [CrossRef]
- Stankiewicz, P.; Lupski, J.R. Genome architecture, rearrangements and genomic disorders. Trends Genet. 2002, 18, 74–82. [Google Scholar] [CrossRef]
- Kellis, M.; Patterson, N.; Endrizzi, M.; Birren, B.; Lander, E.S. Sequencing and comparison of yeast species to identify genes and regulatory elements. Nature 2003, 423, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.L.; Byrne, K.P.; Wolfe, K.H. Additions, losses, and rearrangements on the evolutionary route from a reconstructed ancestor to the modern Saccharomyces cerevisiae genome. PLoS Genet. 2009, 5, e1000485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selmecki, A.M.; Maruvka, Y.E.; Richmond, P.A.; Guillet, M.; Shoresh, N.; Sorenson, A.L.; De, S.; Kishony, R.; Michor, F.; Dowell, R. Polyploidy can drive rapid adaptation in yeast. Nature 2015, 519, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.-X.; Li, J.; Aigrain, L.; Hallin, J.; Persson, K.; Oliver, K.; Bergström, A.; Coupland, P.; Warringer, J.; Lagomarsino, M.C.; et al. Contrasting evolutionary genome dynamics between domesticated and wild yeasts. Nat. Genet. 2017, 49, 913–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, Y.; Qi, L.; Wu, J.-K.; Wen, X.-P.; Tang, X.-X.; Ma, Z.-J.; Wu, X.-C.; Zhang, K.; Kokoska, R.J.; Zheng, D.-Q.; et al. Genome-wide mapping of spontaneous genetic alterations in diploid yeast cells. Proc. Natl. Acad. Sci. USA 2020, 117, 28191. [Google Scholar] [CrossRef] [PubMed]
- Kaya, A.; Mariotti, M.; Tyshkovskiy, A.; Zhou, X.; Hulke, M.L.; Ma, S.; Gerashchenko, M.V.; Koren, A.; Gladyshev, V.N. Molecular signatures of aneuploidy-driven adaptive evolution. Nat. Commun. 2020, 11, 588. [Google Scholar] [CrossRef] [Green Version]
- Raffaele, S.; Farrer, R.A.; Cano, L.M.; Studholme, D.J.; MacLean, D.; Thines, M.; Jiang, R.H.Y.; Zody, M.C.; Kunjeti, S.G.; Donofrio, N.M.; et al. Genome Evolution Following Host Jumps in the Irish Potato Famine Pathogen Lineage. Science 2010, 330, 1540. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.-J.; van der Does, H.C.; Borkovich, K.A.; Coleman, J.J.; Daboussi, M.-J.; Di Pietro, A.; Dufresne, M.; Freitag, M.; Grabherr, M.; Henrissat, B. Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature 2010, 464, 367–373. [Google Scholar] [CrossRef]
- Klosterman, S.J.; Subbarao, K.V.; Kang, S.; Veronese, P.; Gold, S.E.; Thomma, B.P.; Chen, Z.; Henrissat, B.; Lee, Y.-H.; Park, J. Comparative genomics yields insights into niche adaptation of plant vascular wilt pathogens. PLoS Pathog. 2011, 7, e1002137. [Google Scholar] [CrossRef] [Green Version]
- Chuma, I.; Isobe, C.; Hotta, Y.; Ibaragi, K.; Futamata, N.; Kusaba, M.; Yoshida, K.; Terauchi, R.; Fujita, Y.; Nakayashiki, H. Multiple translocation of the AVR-Pita effector gene among chromosomes of the rice blast fungus Magnaporthe oryzae and related species. PLoS Pathog. 2011, 7, e1002147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jonge, R.; Bolton, M.D.; Kombrink, A.; van den Berg, G.C.M.; Yadeta, K.A.; Thomma, B.P.H.J. Extensive chromosomal reshuffling drives evolution of virulence in an asexual pathogen. Genome Res. 2013, 23, 1271–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faino, L.; Seidl, M.F.; Shi-Kunne, X.; Pauper, M.; van den Berg, G.C.; Wittenberg, A.H.; Thomma, B.P. Transposons passively and actively contribute to evolution of the two-speed genome of a fungal pathogen. Genome Res. 2016, 26, 1091–1100. [Google Scholar] [CrossRef] [Green Version]
- Plissonneau, C.; Stürchler, A.; Croll, D. The Evolution of Orphan Regions in Genomes of a Fungal Pathogen of Wheat. mBio 2016, 7, e01231-16. [Google Scholar] [CrossRef] [Green Version]
- Hastings, P.J.; Lupski, J.R.; Rosenberg, S.M.; Ira, G. Mechanisms of change in gene copy number. Nat. Rev. Genet. 2009, 10, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, C.M.; Lupski, J.R. Mechanisms underlying structural variant formation in genomic disorders. Nat. Rev. Genet. 2016, 17, 224–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brewer, B.J.; Payen, C.; Raghuraman, M.K.; Dunham, M.J. Origin-dependent inverted-repeat amplification: A replication-based model for generating palindromic amplicons. PLoS Genet. 2011, 7, e1002016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brewer, B.J.; Payen, C.; Di Rienzi, S.C.; Higgins, M.M.; Ong, G.; Dunham, M.J.; Raghuraman, M.K. Origin-Dependent Inverted-Repeat Amplification: Tests of a Model for Inverted DNA Amplification. PLoS Genet. 2015, 11, e1005699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gresham, D.; Usaite, R.; Germann, S.M.; Lisby, M.; Botstein, D.; Regenberg, B. Adaptation to diverse nitrogen-limited environments by deletion or extrachromosomal element formation of the GAP1 locus. Proc. Natl. Acad. Sci. USA 2010, 107, 18551–18556. [Google Scholar] [CrossRef] [Green Version]
- Møller, H.D.; Andersen, K.S.; Regenberg, B. A model for generating several adaptive phenotypes from a single genetic event: Saccharomyces cerevisiae GAP1 as a potential bet-hedging switch. Commun. Integr. Biol. 2013, 6, e23933. [Google Scholar] [CrossRef]
- Cohen, S.; Segal, D. Extrachromosomal circular DNA in eukaryotes: Possible involvement in the plasticity of tandem repeats. Cytogenet. Genome Res. 2009, 124, 327–338. [Google Scholar] [CrossRef]
- Pontes, O.; Neves, N.; Silva, M.; Lewis, M.S.; Madlung, A.; Comai, L.; Viegas, W.; Pikaard, C.S. Chromosomal locus rearrangements are a rapid response to formation of the allotetraploid Arabidopsis suecica genome. Proc. Natl. Acad. Sci. USA 2004, 101, 18240–18245. [Google Scholar] [CrossRef] [Green Version]
- Madlung, A.; Tyagi, A.P.; Watson, B.; Jiang, H.; Kagochi, T.; Doerge, R.W.; Martienssen, R.; Comai, L. Genomic changes in synthetic Arabidopsis polyploids. Plant J. 2005, 41, 221–230. [Google Scholar] [CrossRef]
- Ohno, S.; Wolf, U.; Atkin, N.B. Evolution from fish to mammals by gene duplication. Hereditas 1968, 59, 169–187. [Google Scholar] [CrossRef]
- Ohno, S. Evolution by Gene Duplication; Springer Science & Business Media: Berlin, Germany, 2013. [Google Scholar]
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and Divergence of Saccharomyces cerevisiae Beer Yeasts. Cell 2016, 166, 1397–1410.e1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergström, A.; Simpson, J.T.; Salinas, F.; Barré, B.; Parts, L.; Zia, A.; Nguyen Ba, A.N.; Moses, A.M.; Louis, E.J.; Mustonen, V.; et al. A High-Definition View of Functional Genetic Variation from Natural Yeast Genomes. Mol. Biol. Evol. 2014, 31, 872–888. [Google Scholar] [CrossRef] [PubMed]
- Borneman, A.R.; Desany, B.A.; Riches, D.; Affourtit, J.P.; Forgan, A.H.; Pretorius, I.S.; Egholm, M.; Chambers, P.J. Whole-Genome Comparison Reveals Novel Genetic Elements That Characterize the Genome of Industrial Strains of Saccharomyces cerevisiae. PLoS Genet. 2011, 7, e1001287. [Google Scholar] [CrossRef] [Green Version]
- Dunn, B.; Richter, C.; Kvitek, D.J.; Pugh, T.; Sherlock, G. Analysis of the Saccharomyces cerevisiae pan-genome reveals a pool of copy number variants distributed in diverse yeast strains from differing industrial environments. Genome Res. 2012, 22, 908–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavelka, N.; Rancati, G.; Zhu, J.; Bradford, W.D.; Saraf, A.; Florens, L.; Sanderson, B.W.; Hattem, G.L.; Li, R. Aneuploidy confers quantitative proteome changes and phenotypic variation in budding yeast. Nature 2010, 468, 321–325. [Google Scholar] [CrossRef]
- Rancati, G.; Pavelka, N.; Fleharty, B.; Noll, A.; Trimble, R.; Walton, K.; Perera, A.; Staehling-Hampton, K.; Seidel, C.W.; Li, R. Aneuploidy Underlies Rapid Adaptive Evolution of Yeast Cells Deprived of a Conserved Cytokinesis Motor. Cell 2008, 135, 879–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selmecki, A.M.; Dulmage, K.; Cowen, L.E.; Anderson, J.B.; Berman, J. Acquisition of aneuploidy provides increased fitness during the evolution of antifungal drug resistance. PLoS Genet. 2009, 5, e1000705. [Google Scholar] [CrossRef]
- Voordeckers, K.; Kominek, J.; Das, A.; Espinosa-Cantú, A.; de Maeyer, D.; Arslan, A.; van Pee, M.; van der Zande, E.; Meert, W.; Yang, Y.; et al. Adaptation to High Ethanol Reveals Complex Evolutionary Pathways. PLoS Genet. 2015, 11, e1005635. [Google Scholar] [CrossRef]
- Purugganan, M.D.; Fuller, D.Q. The nature of selection during plant domestication. Nature 2009, 457, 843–848. [Google Scholar] [CrossRef]
- Low, W.Y.; Tearle, R.; Liu, R.; Koren, S.; Rhie, A.; Bickhart, D.M.; Rosen, B.D.; Kronenberg, Z.N.; Kingan, S.B.; Tseng, E.; et al. Haplotype-resolved genomes provide insights into structural variation and gene content in Angus and Brahman cattle. Nat. Commun. 2020, 11, 2071. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.H.; Shields, D.C. Molecular evidence for an ancient duplication of the entire yeast genome. Nature 1997, 387, 708–713. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.H. Origin of the Yeast Whole-Genome Duplication. PLoS Biol. 2015, 13, e1002221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcet-Houben, M.; Gabaldón, T. Beyond the Whole-Genome Duplication: Phylogenetic Evidence for an Ancient Interspecies Hybridization in the Baker’s Yeast Lineage. PLoS Biol. 2015, 13, e1002220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charron, G.; Marsit, S.; Hénault, M.; Martin, H.; Landry, C.R. Spontaneous whole-genome duplication restores fertility in interspecific hybrids. Nat. Commun. 2019, 10, 4126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, K.P.; Wolfe, K.H. The Yeast Gene Order Browser: Combining curated homology and syntenic context reveals gene fate in polyploid species. Genome Res. 2005, 15, 1456–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fares, M.A.; Keane, O.M.; Toft, C.; Carretero-Paulet, L.; Jones, G.W. The roles of whole-genome and small-scale duplications in the functional specialization of Saccharomyces cerevisiae genes. PLoS Genet. 2013, 9, e1003176. [Google Scholar] [CrossRef] [Green Version]
- Ohno, S. Gene duplication and the uniqueness of vertebrate genomes circa 1970–1999. Semin Cell Dev. Biol. 1999, 10, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Conery, J.S. The evolutionary demography of duplicate genes. Genome Evol. 2003, 3, 35–44. [Google Scholar]
- Kondrashov, F.A.; Rogozin, I.B.; Wolf, Y.I.; Koonin, E.V. Selection in the evolution of gene duplications. Genome Biol. 2002, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, T.J.; Spring, J. Genetic redundancy in vertebrates: Polyploidy and persistence of genes encoding multidomain proteins. Trends Genet. 1998, 14, 46–49. [Google Scholar] [CrossRef]
- Veitia, R.A. Nonlinear effects in macromolecular assembly and dosage sensitivity. J. Theor. Biol. 2003, 220, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Veitia, R.A. A sigmoidal transcriptional response: Cooperativity, synergy and dosage effects. Biol. Rev. 2003, 78, 149–170. [Google Scholar] [CrossRef]
- Force, A.; Lynch, M.; Pickett, F.B.; Amores, A.; Yan, Y.L.; Postlethwait, J. Preservation of duplicate genes by complementary, degenerative mutations. Genetics 1999, 151, 1531–1545. [Google Scholar] [CrossRef]
- Hughes, A.L. The evolution of functionally novel proteins after gene duplication. Proc. R. Soc. London. Ser. B Biol. Sci. 1994, 256, 119–124. [Google Scholar] [CrossRef]
- Bhat, P.J.; Murthy, T. Transcriptional control of the GAL/MEL regulon of yeast Saccharomyces cerevisiae: Mechanism of galactose-mediated signal transduction. Mol. Microbiol. 2001, 40, 1059–1066. [Google Scholar] [CrossRef]
- Taylor, J.S.; Raes, J. Duplication and divergence: The evolution of new genes and old ideas. Annu. Rev. Genet. 2004, 38, 615–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pougach, K.; Voet, A.; Kondrashov, F.A.; Voordeckers, K.; Christiaens, J.F.; Baying, B.; Benes, V.; Sakai, R.; Aerts, J.; Zhu, B. Duplication of a promiscuous transcription factor drives the emergence of a new regulatory network. Nat. Commun. 2014, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Teichmann, S.A.; Babu, M.M. Gene regulatory network growth by duplication. Nat. Genet. 2004, 36, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Voordeckers, K.; Pougach, K.; Verstrepen, K.J. How do regulatory networks evolve and expand throughout evolution? Curr. Opin. Biotechnol. 2015, 34, 180–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattenberger, F.; Sabater-Muñoz, B.; Toft, C.; Fares, M.A. The Phenotypic Plasticity of Duplicated Genes in Saccharomyces cerevisiae and the Origin of Adaptations. G3 2017, 7, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Ha, M.; Kim, E.-D.; Chen, Z.J. Duplicate genes increase expression diversity in closely related species and allopolyploids. Proc. Natl. Acad. Sci. USA 2009, 106, 2295–2300. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, S.; Liberles, D.A. Subfunctionalization of duplicated genes as a transition state to neofunctionalization. BMC Evol. Biol. 2005, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Peer, Y.; Mizrachi, E.; Marchal, K. The evolutionary significance of polyploidy. Nat. Rev. Genet. 2017, 18, 411–424. [Google Scholar] [CrossRef]
- Soltis, P.S.; Marchant, D.B.; van de Peer, Y.; Soltis, D.E. Polyploidy and genome evolution in plants. Curr. Opin. Genet. Dev. 2015, 35, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Arrigo, N.; Barker, M.S. Rarely successful polyploids and their legacy in plant genomes. Curr. Opin. Plant Biol. 2012, 15, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Mable, B.; Alexandrou, M.; Taylor, M. Genome duplication in amphibians and fish: An extended synthesis. J. Zool. 2011, 284, 151–182. [Google Scholar] [CrossRef]
- Otto, S.P.; Whitton, J. Polyploid incidence and evolution. Annu. Rev. Genet. 2000, 34, 401–437. [Google Scholar] [CrossRef] [Green Version]
- Meyers, L.A.; Levin, D.A. On the abundance of polyploids in flowering plants. Evolution 2006, 60, 1198–1206. [Google Scholar] [CrossRef] [PubMed]
- Albertin, W.; Marullo, P. Polyploidy in fungi: Evolution after whole-genome duplication. Proc. Biol. Sci. 2012, 279, 2497–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Peer, Y.; Maere, S.; Meyer, A. The evolutionary significance of ancient genome duplications. Nat. Rev. Genet. 2009, 10, 725–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaillon, O.; Aury, J.-M.; Brunet, F.; Petit, J.-L.; Stange-Thomann, N.; Mauceli, E.; Bouneau, L.; Fischer, C.; Ozouf-Costaz, C.; Bernot, A. Genome duplication in the teleost fish Tetraodon nigroviridis reveals the early vertebrate proto-karyotype. Nature 2004, 431, 946–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, A.; van de Peer, Y. From 2R to 3R: Evidence for a fish-specific genome duplication (FSGD). Bioessays 2005, 27, 937–945. [Google Scholar] [CrossRef] [Green Version]
- Adams, K.L.; Wendel, J.F. Polyploidy and genome evolution in plants. Curr. Opin. Plant Biol. 2005, 8, 135–141. [Google Scholar] [CrossRef]
- Tang, H.; Bowers, J.E.; Wang, X.; Ming, R.; Alam, M.; Paterson, A.H. Synteny and collinearity in plant genomes. Science 2008, 320, 486–488. [Google Scholar] [CrossRef] [Green Version]
- Boore, J.L. Two rounds of whole genome duplication in the ancestral vertebrate genome. PLoS Biol. 2006, 10, 314. [Google Scholar]
- Kellis, M.; Birren, B.W.; Lander, E.S. Proof and evolutionary analysis of ancient genome duplication in the yeast Saccharomyces cerevisiae. Nature 2004, 428, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Edgar, B.A.; Orr-Weaver, T.L. Endoreplication cell cycles: More for less. Cell 2001, 105, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Harari, Y.; Ram, Y.; Kupiec, M. Frequent ploidy changes in growing yeast cultures. Curr. Genet. 2018, 64, 1001–1004. [Google Scholar] [CrossRef]
- Ortiz-Merino, R.A.; Kuanyshev, N.; Braun-Galleani, S.; Byrne, K.P.; Porro, D.; Branduardi, P.; Wolfe, K.H. Evolutionary restoration of fertility in an interspecies hybrid yeast, by whole-genome duplication after a failed mating-type switch. PLoS Biol. 2017, 15, e2002128. [Google Scholar] [CrossRef]
- Steensels, J.; Gallone, B.; Verstrepen, K.J. Interspecific hybridization as a driver of fungal evolution and adaptation. Nat. Rev. Microbiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Kosheleva, K.; Desai, M.M. Recombination Alters the Dynamics of Adaptation on Standing Variation in Laboratory Yeast Populations. Mol. Biol. Evol. 2018, 35, 180–201. [Google Scholar] [CrossRef]
- Gorter, F.A.; Derks, M.F.L.; van den Heuvel, J.; Aarts, M.G.M.; Zwaan, B.J.; de Ridder, D.; de Visser, J.A.G.M. Genomics of Adaptation Depends on the Rate of Environmental Change in Experimental Yeast Populations. Mol. Biol. Evol. 2017, 34, 2613–2626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkataram, S.; Dunn, B.; Li, Y.; Agarwala, A.; Chang, J.; Ebel, E.R.; Geiler-Samerotte, K.; Herissant, L.; Blundell, J.R.; Levy, S.F.; et al. Development of a Comprehensive Genotype-to-Fitness Map of Adaptation-Driving Mutations in Yeast. Cell 2016, 166, 1585–1596. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Gresham, D. Molecular Specificity, Convergence and Constraint Shape Adaptive Evolution in Nutrient-Poor Environments. PLoS Genet. 2014, 10, e1004041. [Google Scholar] [CrossRef] [Green Version]
- Gerstein, A.C.; Chun, H.-J.E.; Grant, A.; Otto, S.P. Genomic Convergence toward Diploidy in Saccharomyces cerevisiae. PLoS Genet. 2006, 2, e145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harari, Y.; Ram, Y.; Rappoport, N.; Hadany, L.; Kupiec, M. Spontaneous Changes in Ploidy Are Common in Yeast. Curr. Biol. 2018, 28, 825–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsey, J.; Schemske, D.W. Pathways, Mechanisms, and Rates of Polyploid Formation in Flowering Plants. Annu. Rev. Ecol. Syst. 1998, 29, 467–501. [Google Scholar] [CrossRef] [Green Version]
- Sharp, N.P.; Sandell, L.; James, C.G.; Otto, S.P. The genome-wide rate and spectrum of spontaneous mutations differ between haploid and diploid yeast. Proc. Natl. Acad. Sci. USA 2018, 115, E5046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, M.; Sung, W.; Morris, K.; Coffey, N.; Landry, C.R.; Dopman, E.B.; Dickinson, W.J.; Okamoto, K.; Kulkarni, S.; Hartl, D.L.; et al. A genome-wide view of the spectrum of spontaneous mutations in yeast. Proc. Natl. Acad. Sci. USA 2008, 105, 9272–9277. [Google Scholar] [CrossRef] [Green Version]
- Nishant, K.T.; Wei, W.; Mancera, E.; Argueso, J.L.; Schlattl, A.; Delhomme, N.; Ma, X.; Bustamante, C.D.; Korbel, J.O.; Gu, Z.; et al. The Baker’s Yeast Diploid Genome Is Remarkably Stable in Vegetative Growth and Meiosis. PLoS Genet. 2010, 6, e1001109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.O.; Siegal, M.L.; Hall, D.W.; Petrov, D.A. Precise estimates of mutation rate and spectrum in yeast. Proc. Natl. Acad. Sci. USA 2014, 111, E2310–E2318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, A.; Lin, G.; Pankajam, A.V.; Chakraborty, P.; Bhat, N.; Steinmetz, L.M.; Nishant, K.T. Genome Dynamics of Hybrid Saccharomyces cerevisiae During Vegetative and Meiotic Divisions. G3 2017, 7, 3669–3679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, J.; de Chiara, M.; Friedrich, A.; Yue, J.X.; Pflieger, D.; Bergström, A.; Sigwalt, A.; Barre, B.; Freel, K.; Llored, A.; et al. Genome evolution across 1011 Saccharomyces cerevisiae isolates. Nature 2018, 556, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Epstein, C.J. Cell size, nuclear content, and the development of polyploidy in the mammalian liver. Proc. Natl. Acad. Sci. USA 1967, 57, 327. [Google Scholar] [CrossRef] [Green Version]
- Gregory, T.R. Coincidence, coevolution, or causation? DNA content, cellsize, and the C-value enigma. Biol. Rev. 2001, 76, 65–101. [Google Scholar] [CrossRef]
- Beaulieu, J.M.; Leitch, I.J.; Patel, S.; Pendharkar, A.; Knight, C.A. Genome size is a strong predictor of cell size and stomatal density in angiosperms. New Phytol. 2008, 179, 975–986. [Google Scholar] [CrossRef] [Green Version]
- Marad, D.A.; Buskirk, S.W.; Lang, G.I. Altered access to beneficial mutations slows adaptation and biases fixed mutations in diploids. Nat. Ecol. Evol. 2018, 2, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zeidler, A.F.B.; Song, W.; Puccia, C.M.; Malc, E.; Greenwell, P.W.; Mieczkowski, P.A.; Petes, T.D.; Argueso, J.L. Gene Copy-Number Variation in Haploid and Diploid Strains of the Yeast Saccharomyces cerevisiae. Genetics 2013, 193, 785. [Google Scholar] [CrossRef] [Green Version]
- Deutschbauer, A.M.; Jaramillo, D.F.; Proctor, M.; Kumm, J.; Hillenmeyer, M.E.; Davis, R.W.; Nislow, C.; Giaever, G. Mechanisms of haploinsufficiency revealed by genome-wide profiling in yeast. Genetics 2005, 169, 1915–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selmecki, A.; Forche, A.; Berman, J. Aneuploidy and isochromosome formation in drug-resistant Candida albicans. Science 2006, 313, 367–370. [Google Scholar] [CrossRef] [Green Version]
- Ni, M.; Feretzaki, M.; Li, W.; Floyd-Averette, A.; Mieczkowski, P.; Dietrich, F.S.; Heitman, J. Unisexual and Heterosexual Meiotic Reproduction Generate Aneuploidy and Phenotypic Diversity De Novo in the Yeast Cryptococcus neoformans. PLoS Biol. 2013, 11, e1001653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, R.J.; Forche, A.; Berman, J. Rapid mechanisms for generating genome diversity: Whole ploidy shifts, aneuploidy, and loss of heterozygosity. Cold Spring Harb. Perspect. Med. 2014, 4, a019604. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.-J.; Swamy, K.B.S.; Leu, J.-Y. Experimental Evolution Reveals Interplay between Sch9 and Polyploid Stability in Yeast. PLoS Genet. 2016, 12, e1006409. [Google Scholar] [CrossRef]
- Storchová, Z.; Breneman, A.; Cande, J.; Dunn, J.; Burbank, K.; O’Toole, E.; Pellman, D. Genome-wide genetic analysis of polyploidy in yeast. Nature 2006, 443, 541–547. [Google Scholar] [CrossRef]
- Storchova, Z. Ploidy changes and genome stability in yeast. Yeast 2014, 31, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Andalis, A.A.; Storchova, Z.; Styles, C.; Galitski, T.; Pellman, D.; Fink, G.R. Defects arising from whole-genome duplications in Saccharomyces cerevisiae. Genetics 2004, 167, 1109–1121. [Google Scholar] [CrossRef] [Green Version]
- Mayer, V.W.; Goin, C.J.; Arras, C.A.; Taylor-Mayer, R.E. Comparison of chemically induced chromosome loss in a diploid, triploid, and tetraploid strain of Saccharomyces cerevisiae. Mutat. Res. Genet. Toxicol. 1992, 279, 41–48. [Google Scholar] [CrossRef]
- Mayer, V.W.; Aguilera, A. High levels of chromosome instability in polyploids of Saccharomyces cerevisiae. Mutat. Res. Fundam. Mol. Mech. Mutagenes. 1990, 231, 177–186. [Google Scholar] [CrossRef]
- Mortimer, R.K. Radiobiological and genetic studies on a polyploid series (haploid to hexaploid) of Saccharomyces cerevisiae. Radiat. Res. 1958, 9, 312–326. [Google Scholar] [CrossRef]
- Scott, A.L.; Richmond, P.A.; Dowell, R.D.; Selmecki, A.M. The Influence of Polyploidy on the Evolution of Yeast Grown in a Sub-Optimal Carbon Source. Mol. Biol. Evol. 2017, 34, 2690–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorpe, P.H.; González-Barrera, S.; Rothstein, R. More is not always better: The genetic constraints of polyploidy. Trends Genet. 2007, 23, 263–266. [Google Scholar] [CrossRef]
- Torres, E.M.; Sokolsky, T.; Tucker, C.M.; Chan, L.Y.; Boselli, M.; Dunham, M.J.; Amon, A. Effects of aneuploidy on cellular physiology and cell division in haploid yeast. Science 2007, 317, 916–924. [Google Scholar] [CrossRef]
- Sheltzer, J.M.; Blank, H.M.; Pfau, S.J.; Tange, Y.; George, B.M.; Humpton, T.J.; Brito, I.L.; Hiraoka, Y.; Niwa, O.; Amon, A. Aneuploidy drives genomic instability in yeast. Science 2011, 333, 1026–1030. [Google Scholar] [CrossRef] [Green Version]
- Oromendia, A.B.; Dodgson, S.E.; Amon, A. Aneuploidy causes proteotoxic stress in yeast. Genes Dev. 2012, 26, 2696–2708. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Pavelka, N.; Bradford, W.D.; Rancati, G.; Li, R. Karyotypic determinants of chromosome instability in aneuploid budding yeast. PLoS Genet. 2012, 8, e1002719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oromendia, A.B.; Amon, A. Aneuploidy: Implications for protein homeostasis and disease. Dis. Model. Mech. 2014, 7, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnelly, N.; Storchova, Z. Causes and consequences of protein folding stress in aneuploid cells. Cell Cycle 2015, 14, 495–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passerini, V.; Ozeri-Galai, E.; de Pagter, M.S.; Donnelly, N.; Schmalbrock, S.; Kloosterman, W.P.; Kerem, B.; Storchova, Z. The presence of extra chromosomes leads to genomic instability. Nat. Commun. 2016, 7, 10754. [Google Scholar] [CrossRef] [Green Version]
- Ravichandran, M.C.; Fink, S.; Clarke, M.N.; Hofer, F.C.; Campbell, C.S. Genetic interactions between specific chromosome copy number alterations dictate complex aneuploidy patterns. Genes Dev. 2018, 32, 1485–1498. [Google Scholar] [CrossRef] [Green Version]
- Tsai, H.-J.; Nelliat, A.R.; Choudhury, M.I.; Kucharavy, A.; Bradford, W.D.; Cook, M.E.; Kim, J.; Mair, D.B.; Sun, S.X.; Schatz, M.C.; et al. Hypo-osmotic-like stress underlies general cellular defects of aneuploidy. Nature 2019, 570, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Torres, E.M.; Dephoure, N.; Panneerselvam, A.; Tucker, C.M.; Whittaker, C.A.; Gygi, S.P.; Dunham, M.J.; Amon, A. Identification of aneuploidy-tolerating mutations. Cell 2010, 143, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Sheltzer, J.M.; Torres, E.M.; Dunham, M.J.; Amon, A. Transcriptional consequences of aneuploidy. Proc. Natl. Acad. Sci. USA 2012, 109, 12644–12649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorburn, R.R.; Gonzalez, C.; Brar, G.A.; Christen, S.; Carlile, T.M.; Ingolia, N.T.; Sauer, U.; Weissman, J.S.; Amon, A. Aneuploid yeast strains exhibit defects in cell growth and passage through START. Mol. Biol. Cell 2013, 24, 1274–1289. [Google Scholar] [CrossRef]
- Dephoure, N.; Hwang, S.; O’Sullivan, C.; Dodgson, S.E.; Gygi, S.P.; Amon, A.; Torres, E.M. Quantitative proteomic analysis reveals posttranslational responses to aneuploidy in yeast. eLife 2014, 3, e03023. [Google Scholar] [CrossRef]
- Dodgson, S.E.; Santaguida, S.; Kim, S.; Sheltzer, J.; Amon, A. The pleiotropic deubiquitinase Ubp3 confers aneuploidy tolerance. Genes Dev. 2016, 30, 2259–2271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brennan, C.M.; Vaites, L.P.; Wells, J.N.; Santaguida, S.; Paulo, J.A.; Storchova, Z.; Harper, J.W.; Marsh, J.A.; Amon, A. Protein aggregation mediates stoichiometry of protein complexes in aneuploid cells. Genes Dev. 2019, 33, 1031–1047. [Google Scholar] [CrossRef] [Green Version]
- Hose, J.; Yong, C.M.; Sardi, M.; Wang, Z.; Newton, M.A.; Gasch, A.P. Dosage compensation can buffer copy-number variation in wild yeast. eLife 2015, 4, e05462. [Google Scholar] [CrossRef] [PubMed]
- Strope, P.K.; Skelly, D.A.; Kozmin, S.G.; Mahadevan, G.; Stone, E.A.; Magwene, P.M.; Dietrich, F.S.; McCusker, J.H. The 100-genomes strains, an S. cerevisiae resource that illuminates its natural phenotypic and genotypic variation and emergence as an opportunistic pathogen. Genome Res. 2015, 25, 762–774. [Google Scholar] [CrossRef] [Green Version]
- Van den Broek, M.; Bolat, I.; Nijkamp, J.; Ramos, E.; Luttik, M.A.; Koopman, F.; Geertman, J.; de Ridder, D.; Pronk, J.T.; Daran, J.-M. Chromosomal copy number variation in Saccharomyces pastorianus is evidence for extensive genome dynamics in industrial lager brewing strains. Appl. Environ. Microbiol. 2015, 81, 6253–6267. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.O.; Sherlock, G.; Petrov, D.A. Whole Genome Analysis of 132 Clinical Saccharomyces cerevisiae Strains Reveals Extensive Ploidy Variation. G3 2016, 6, 2421–2434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorter de Vries, A.R.; Pronk, J.T.; Daran, J.-M.G. Industrial Relevance of Chromosomal Copy Number Variation in Saccharomyces Yeasts. Appl. Environ. Microbiol. 2017, 83, e03206-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, S.F.; Han, P.J.; Wang, Q.M.; Liu, W.Q.; Shi, J.Y.; Li, K.; Zhang, X.L.; Bai, F.Y. The origin and adaptive evolution of domesticated populations of yeast from Far East Asia. Nat. Commun. 2018, 9, 2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fay, J.C.; Liu, P.; Ong, G.T.; Dunham, M.J.; Cromie, G.A.; Jeffery, E.W.; Ludlow, C.L.; Dudley, A.M. A polyploid admixed origin of beer yeasts derived from European and Asian wine populations. PLoS Biol. 2019, 17, e3000147. [Google Scholar] [CrossRef] [Green Version]
- Gallone, B.; Steensels, J.; Mertens, S.; Dzialo, M.C.; Gordon, J.L.; Wauters, R.; Thesseling, F.A.; Bellinazzo, F.; Saels, V.; Herrera-Malaver, B.; et al. Interspecific hybridization facilitates niche adaptation in beer yeast. Nat. Ecol. Evol. 2019, 3, 1562–1575. [Google Scholar] [CrossRef] [PubMed]
- Gasch, A.P.; Hose, J.; Newton, M.A.; Sardi, M.; Yong, M.; Wang, Z. Further support for aneuploidy tolerance in wild yeast and effects of dosage compensation on gene copy-number evolution. eLife 2016, 5, e14409. [Google Scholar] [CrossRef] [PubMed]
- Hose, J.; Escalante, L.E.; Clowers, K.J.; Dutcher, H.A.; Robinson, D.; Bouriakov, V.; Coon, J.J.; Shishkova, E.; Gasch, A.P. The genetic basis of aneuploidy tolerance in wild yeast. eLife 2020, 9. [Google Scholar] [CrossRef]
- Larrimore, K.E.; Barattin-Voynova, N.S.; Reid, D.W.; Ng, D.T.W. Aneuploidy-induced proteotoxic stress can be effectively tolerated without dosage compensation, genetic mutations, or stress responses. BMC Biol. 2020, 18, 117. [Google Scholar] [CrossRef] [PubMed]
- Scopel, E.F.C.; Hose, J.; Bensasson, D.; Gasch, A.P. Genetic variation in aneuploidy prevalence and tolerance across the Saccharomyces cerevisiae phylogeny. bioRxiv 2020. [Google Scholar] [CrossRef]
- Chen, G.; Bradford, W.D.; Seidel, C.W.; Li, R. Hsp90 stress potentiates rapid cellular adaptation through induction of aneuploidy. Nature 2012, 482, 246–250. [Google Scholar] [CrossRef] [Green Version]
- Yona, A.H.; Manor, Y.S.; Herbst, R.H.; Romano, G.H.; Mitchell, A.; Kupiec, M.; Pilpel, Y.; Dahan, O. Chromosomal duplication is a transient evolutionary solution to stress. Proc. Natl. Acad. Sci. USA 2012, 109, 21010–21015. [Google Scholar] [CrossRef] [Green Version]
- Sunshine, A.B.; Payen, C.; Ong, G.T.; Liachko, I.; Tan, K.M.; Dunham, M.J. The fitness consequences of aneuploidy are driven by condition-dependent gene effects. PLoS Biol. 2015, 13, e1002155. [Google Scholar] [CrossRef]
- Kaya, A.; Gerashchenko, M.V.; Seim, I.; Labarre, J.; Toledano, M.B.; Gladyshev, V.N. Adaptive aneuploidy protects against thiol peroxidase deficiency by increasing respiration via key mitochondrial proteins. Proc. Natl. Acad. Sci. USA 2015, 112, 10685–10690. [Google Scholar] [CrossRef] [Green Version]
- Linder, R.A.; Greco, J.P.; Seidl, F.; Matsui, T.; Ehrenreich, I.M. The stress-inducible peroxidase TSA2 underlies a conditionally beneficial chromosomal duplication in Saccharomyces cerevisiae. G3 Genes Genomes Genet. 2017, 7, 3177–3184. [Google Scholar] [CrossRef] [Green Version]
- Beaupere, C.; Dinatto, L.; Wasko, B.M.; Chen, R.B.; VanValkenburg, L.; Kiflezghi, M.G.; Lee, M.B.; Promislow, D.E.L.; Dang, W.; Kaeberlein, M.; et al. Genetic screen identifies adaptive aneuploidy as a key mediator of ER stress resistance in yeast. Proc. Natl. Acad. Sci. USA 2018, 115, 9586–9591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Teoh, F.; Tan, A.S.M.; Cao, Y.; Pavelka, N.; Berman, J. Aneuploidy Enables Cross-Adaptation to Unrelated Drugs. Mol. Biol. Evol. 2019, 36, 1768–1782. [Google Scholar] [CrossRef]
- Todd, R.T.; Selmecki, A. Expandable and reversible copy number amplification drives rapid adaptation to antifungal drugs. eLife 2020, 9. [Google Scholar] [CrossRef]
- Millet, C.; Ausiannikava, D.; Le Bihan, T.; Granneman, S.; Makovets, S. Cell populations can use aneuploidy to survive telomerase insufficiency. Nat. Commun. 2015, 6, 8664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millet, C.; Makovets, S. Aneuploidy as a mechanism of adaptation to telomerase insufficiency. Curr. Genet. 2016, 62, 557–564. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Yong, M.Y.J.; Yurieva, M.; Srinivasan, K.G.; Liu, J.; Lim, J.S.Y.; Poidinger, M.; Wright, G.D.; Zolezzi, F.; Choi, H. Gene essentiality is a quantitative property linked to cellular evolvability. Cell 2015, 163, 1388–1399. [Google Scholar] [CrossRef] [Green Version]
- Ryu, H.Y.; Wilson, N.R.; Mehta, S.; Hwang, S.S.; Hochstrasser, M. Loss of the SUMO protease Ulp2 triggers a specific multichromosome aneuploidy. Genes Dev. 2016, 30, 1881–1894. [Google Scholar] [CrossRef] [Green Version]
- Hughes, T.R.; Roberts, C.J.; Dai, H.; Jones, A.R.; Meyer, M.R.; Slade, D.; Burchard, J.; Dow, S.; Ward, T.R.; Kidd, M.J.; et al. Widespread aneuploidy revealed by DNA microarray expression profiling. Nat. Genet. 2000, 25, 333–337. [Google Scholar] [CrossRef]
- Pankajam, A.V.; Dash, S.; Saifudeen, A.; Dutta, A.; Nishant, K.T. Loss of Heterozygosity and Base Mutation Rates Vary among Saccharomyces cerevisiae Hybrid Strains. G3 Genes Genomes Genet. 2020, 10, 3309. [Google Scholar] [CrossRef]
- Tsai, H.J.; Nelliat, A. A Double-Edged Sword: Aneuploidy is a Prevalent Strategy in Fungal Adaptation. Genes 2019, 10, 787. [Google Scholar] [CrossRef] [Green Version]
- Berman, J. Ploidy plasticity: A rapid and reversible strategy for adaptation to stress. FEMS Yeast Res. 2016, 16, fow020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Rubinstein, B.; Li, R. Whole chromosome aneuploidy: Big mutations drive adaptation by phenotypic leap. Bioessays 2012, 34, 893–900. [Google Scholar] [CrossRef] [Green Version]
- Liebman, S.; Shalit, P.; Picologlou, S. Ty elements are involved in the formation of deletions in DEL1 strains of Saccharomyces cerevisiae. Cell 1981, 26, 401–409. [Google Scholar] [CrossRef]
- Rothstein, R.; Helms, C.; Rosenberg, N. Concerted deletions and inversions are caused by mitotic recombination between delta sequences in Saccharomyces cerevisiae. Mol. Cell. Biol. 1987, 7, 1198–1207. [Google Scholar] [CrossRef] [Green Version]
- Schiestl, R.H. Nonmutagenic carcinogens induce intrachromosomal recombination in yeast. Nature 1989, 337, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Dorsey, M.; Peterson, C.; Bray, K.; Paquin, C.E. Spontaneous amplification of the ADH4 gene in Saccharomyces cerevisiae. Genetics 1992, 132, 943–950. [Google Scholar] [CrossRef]
- Tourrette, Y.; Schacherer, J.; Fritsch, E.; Potier, S.; Souciet, J.L.; de Montigny, J. Spontaneous deletions and reciprocal translocations in Saccharomyces cerevisiae: Influence of ploidy. Mol. Microbiol. 2007, 64, 382–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pryde, F.E.; Gorham, H.C.; Louis, E.J. Chromosome ends: All the same under their caps. Curr. Opin. Genet. Dev. 1997, 7, 822–828. [Google Scholar] [CrossRef]
- Mefford, H.C.; Trask, B.J. The complex structure and dynamic evolution of human subtelomeres. Nat. Rev. Genet. 2002, 3, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Eichler, E.E.; Sankoff, D. Structural dynamics of eukaryotic chromosome evolution. Science 2003, 301, 793–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dujon, B. Yeast evolutionary genomics. Nat. Rev. Genet. 2010, 11, 512–524. [Google Scholar] [CrossRef]
- Winzeler, E.A.; Castillo-Davis, C.I.; Oshiro, G.; Liang, D.; Richards, D.R.; Zhou, Y.; Hartl, D.L. Genetic diversity in yeast assessed with whole-genome oligonucleotide arrays. Genetics 2003, 163, 79–89. [Google Scholar] [CrossRef]
- Brown, C.A.; Murray, A.W.; Verstrepen, K.J. Rapid expansion and functional divergence of subtelomeric gene families in yeasts. Curr. Biol. 2010, 20, 895–903. [Google Scholar] [CrossRef] [Green Version]
- Ames, R.M.; Rash, B.M.; Hentges, K.E.; Robertson, D.L.; Delneri, D.; Lovell, S.C. Gene duplication and environmental adaptation within yeast populations. Genome Biol. Evol. 2010, 2, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirschner, M.; Gerhart, J. Evolvability. Proc. Natl. Acad. Sci. USA 1998, 95, 8420–8427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bendixsen, D.P.; Gettle, N.; Gilchrist, C.; Zhang, Z.; Stelkens, R. Genomic evidence of an ancient East Asian divergence event in wild Saccharomyces cerevisiae. Genome Biol. Evol. 2021, 13. [Google Scholar] [CrossRef]
- Avelar, A.T.; Perfeito, L.; Gordo, I.; Ferreira, M.G. Genome architecture is a selectable trait that can be maintained by antagonistic pleiotropy. Nat. Commun. 2013, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Gordon, J.L.; Byrne, K.P.; Wolfe, K.H. Mechanisms of chromosome number evolution in yeast. PLoS Genet. 2011, 7, e1002190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, J.A.; Huang, J.C.; Pukkila-Worley, R.; Alspaugh, J.A.; Mitchell, T.G.; Heitman, J. Chromosomal Translocation and Segmental Duplication in Cryptococcus neoformans. Eukaryotic Cell 2005, 4, 401. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, R.E.; Sim, A.D.; Chettri, P.; Dupont, P.Y.; Guo, Y.; Hunziker, L.; McDougal, R.L.; van der Nest, A.; Fourie, A.; Wheeler, D. Global population genomics of the forest pathogen Dothistroma septosporum reveal chromosome duplications in high dothistromin-producing strains. Mol. Plant Pathol. 2019, 20, 784–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Sun, M.; Zhang, Y.; Song, Z.; Zhang, S.; Zhang, Q.; Xu, J.R.; Liu, H. Extensive chromosomal rearrangements and rapid evolution of novel effector superfamilies contribute to host adaptation and speciation in the basal ascomycetous fungi. Mol. Plant Pathol. 2020, 21, 330–348. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Friedrich, A.; de Montigny, J.; Schacherer, J. Chromosomal rearrangements as a major mechanism in the onset of reproductive isolation in Saccharomyces cerevisiae. Curr. Biol. 2014, 24, 1153–1159. [Google Scholar] [CrossRef] [Green Version]
- Noor, M.A.; Grams, K.L.; Bertucci, L.A.; Reiland, J. Chromosomal inversions and the reproductive isolation of species. Proc. Natl. Acad. Sci. USA 2001, 98, 12084–12088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieseberg, L.H. Chromosomal rearrangements and speciation. Trends Ecol. Evol. 2001, 16, 351–358. [Google Scholar] [CrossRef]
- Kirkpatrick, M.; Barton, N. Chromosome inversions, local adaptation and speciation. Genetics 2006, 173, 419–434. [Google Scholar] [CrossRef] [Green Version]
- Navarro, A.; Barton, N.H. Accumulating postzygotic isolation genes in parapatry: A new twist on chromosomal speciation. Evolution 2003, 57, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Seoighe, C.; Federspiel, N.; Jones, T.; Hansen, N.; Bivolarovic, V.; Surzycki, R.; Tamse, R.; Komp, C.; Huizar, L.; Davis, R.W.; et al. Prevalence of small inversions in yeast gene order evolution. Proc. Natl. Acad. Sci. USA 2000, 97, 14433–14437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faria, R.; Navarro, A. Chromosomal speciation revisited: Rearranging theory with pieces of evidence. Trends Ecol. Evol. 2010, 25, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Puig Giribets, M.; García Guerreiro, M.P.; Santos, M.; Ayala, F.J.; Tarrío, R.; Rodríguez-Trelles, F. Chromosomal inversions promote genomic islands of concerted evolution of Hsp70 genes in the Drosophila subobscura species subgroup. Mol. Ecol. 2019, 28, 1316–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayala, D.; Zhang, S.; Chateau, M.; Fouet, C.; Morlais, I.; Costantini, C.; Hahn, M.W.; Besansky, N.J. Association mapping desiccation resistance within chromosomal inversions in the African malaria vector Anopheles gambiae. Mol. Ecol. 2019, 28, 1333–1342. [Google Scholar] [CrossRef] [Green Version]
- Colson, I.; Delneri, D.; Oliver, S.G. Effects of reciprocal chromosomal translocations on the fitness of Saccharomyces cerevisiae. EMBO Rep. 2004, 5, 392–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, W.R.; Liti, G.; Rosa, C.; James, S.; Roberts, I.; Robert, V.; Jolly, N.; Tang, W.; Baumann, P.; Green, C.; et al. A Geographically Diverse Collection of Schizosaccharomyces pombe Isolates Shows Limited Phenotypic Variation but Extensive Karyotypic Diversity. G3 2011, 1, 615–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naseeb, S.; Carter, Z.; Minnis, D.; Donaldson, I.; Zeef, L.; Delneri, D. Widespread Impact of Chromosomal Inversions on Gene Expression Uncovers Robustness via Phenotypic Buffering. Mol. Biol. Evol. 2016, 33, 1679–1696. [Google Scholar] [CrossRef] [Green Version]
- Thomson, J.M.; Gaucher, E.A.; Burgan, M.F.; de Kee, D.W.; Li, T.; Aris, J.P.; Benner, S.A. Resurrecting ancestral alcohol dehydrogenases from yeast. Nat. Genet. 2005, 37, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Piškur, J.; Rozpędowska, E.; Polakova, S.; Merico, A.; Compagno, C. How did Saccharomyces evolve to become a good brewer? Trends Genet. 2006, 22, 183–186. [Google Scholar] [CrossRef]
- Conant, G.C.; Wolfe, K.H. Increased glycolytic flux as an outcome of whole-genome duplication in yeast. Mol. Syst. Biol. 2007, 3, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hoek, M.J.; Hogeweg, P. Metabolic adaptation after whole genome duplication. Mol. Biol. Evol. 2009, 26, 2441–2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.J.; Todd, K.M.; Rosenzweig, R.F. Multiple duplications of yeast hexose transport genes in response to selection in a glucose-limited environment. Mol. Biol. Evol. 1998, 15, 931–942. [Google Scholar] [CrossRef] [Green Version]
- Lauer, S.; Avecilla, G.; Spealman, P.; Sethia, G.; Brandt, N.; Levy, S.F.; Gresham, D. Single-cell copy number variant detection reveals the dynamics and diversity of adaptation. PLoS Biol. 2018, 16, e3000069. [Google Scholar] [CrossRef] [Green Version]
- Nikitin, D.; Tosato, V.; Zavec, A.B.; Bruschi, C.V. Cellular and molecular effects of nonreciprocal chromosome translocations in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2008, 105, 9703–9708. [Google Scholar] [CrossRef] [Green Version]
- Janbon, G.; Sherman, F.; Rustchenko, E. Monosomy of a specific chromosome determines sorbose utilization: A novel regulatory mechanism in Candida albicans. Proc. Natl. Acad. Sci. USA 1998, 95, 5150. [Google Scholar] [CrossRef] [Green Version]
- Ezov, T.K.; Boger-Nadjar, E.; Frenkel, Z.E.; Katsperovski, I.; Kemeny, S.; Nevo, E.; Korol, A.; Kashi, Y. Molecular-genetic biodiversity in a natural population of the yeast Saccharomyces cerevisiae from “Evolution Canyon”: Microsatellite polymorphism, ploidy and controversial sexual status. Genetics 2006, 174, 1455–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.-L.; Lai, H.-Y.; Tung, S.-Y.; Leu, J.-Y. Dynamic large-scale chromosomal rearrangements fuel rapid adaptation in yeast populations. PLoS Genet. 2013, 9, e1003232. [Google Scholar] [CrossRef] [Green Version]
- Gerstein, A.C.; Ono, J.; Lo, D.S.; Campbell, M.L.; Kuzmin, A.; Otto, S.P. Too much of a good thing: The unique and repeated paths toward copper adaptation. Genetics 2015, 199, 555–571. [Google Scholar] [CrossRef] [Green Version]
- Will, J.L.; Kim, H.S.; Clarke, J.; Painter, J.C.; Fay, J.C.; Gasch, A.P. Incipient balancing selection through adaptive loss of aquaporins in natural Saccharomyces cerevisiae populations. PLoS Genet. 2010, 6, e1000893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.-J.; Lu, M.-Y.; Chang, Y.-W.; Li, W.-H. Experimental Evolution of Yeast for High-Temperature Tolerance. Mol. Biol. Evol. 2018, 35, 1823–1839. [Google Scholar] [CrossRef]
- Caspeta, L.; Chen, Y.; Ghiaci, P.; Feizi, A.; Buskov, S.; Hallström, B.M.; Petranovic, D.; Nielsen, J. Altered sterol composition renders yeast thermotolerant. Science 2014, 346, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Morard, M.; Macías, L.G.; Adam, A.C.; Lairón-Peris, M.; Pérez-Torrado, R.; Toft, C.; Barrio, E. Aneuploidy and Ethanol Tolerance in Saccharomyces cerevisiae. Front. Genet. 2019, 10, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto-Yamamoto, N.; Kitano, K.; Shiki, K.; Yoshida, Y.; Suzuki, T.; Iwata, T.; Yamane, Y.; Hara, S. SSU1-R, a sulfite resistance gene of wine yeast, is an allele of SSU1 with a different upstream sequence. J. Ferment. Bioeng. 1998, 86, 427–433. [Google Scholar] [CrossRef]
- Pérez-Ortın, J.E.; Querol, A.; Puig, S.; Barrio, E. Molecular characterization of a chromosomal rearrangement involved in the adaptive evolution of yeast strains. Genome Res. 2002, 12, 1533–1539. [Google Scholar] [CrossRef] [Green Version]
- Yuasa, N.; Nakagawa, Y.; Hayakawa, M.; Iimura, Y. Distribution of the sulfite resistance gene SSU1-R and the variation in its promoter region in wine yeasts. J. Biosc. Bioeng. 2004, 98, 394–397. [Google Scholar] [CrossRef]
- Zimmer, A.; Durand, C.; Loira, N.; Durrens, P.; Sherman, D.J.; Marullo, P. QTL dissection of lag phase in wine fermentation reveals a new translocation responsible for Saccharomyces cerevisiae adaptation to sulfite. PLoS ONE 2014, 9, e86298. [Google Scholar] [CrossRef] [Green Version]
- García-Ríos, E.; Nuévalos, M.; Barrio, E.; Puig, S.; Guillamón, J.M. A new chromosomal rearrangement improves the adaptation of wine yeasts to sulfite. Environ. Microbiol. 2019, 21, 1771–1781. [Google Scholar] [CrossRef] [Green Version]
- Selmecki, A.; Gerami-Nejad, M.; Paulson, C.; Forche, A.; Berman, J. An isochromosome confers drug resistance in vivo by amplification of two genes, ERG11 and TAC1. Mol. Microbiol. 2008, 68, 624–641. [Google Scholar] [CrossRef] [PubMed]
- Sionov, E.; Lee, H.; Chang, Y.C.; Kwon-Chung, K.J. Cryptococcus neoformans overcomes stress of azole drugs by formation of disomy in specific multiple chromosomes. PLoS Pathog. 2010, 6, e1000848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sionov, E.; Chang, Y.C.; Kwon-Chung, K.J. Azole heteroresistance in Cryptococcus neoformans: Emergence of resistant clones with chromosomal disomy in the mouse brain during fluconazole treatment. Antimicrob. Agents Chemother. 2013, 57, 5127–5130. [Google Scholar] [CrossRef] [Green Version]
- Ngamskulrungroj, P.; Chang, Y.; Hansen, B.; Bugge, C.; Fischer, E.; Kwon-Chung, K.J. Characterization of the chromosome 4 genes that affect fluconazole-induced disomy formation in Cryptococcus neoformans. PLoS ONE 2012, 7, e33022. [Google Scholar] [CrossRef] [Green Version]
- Ford, C.B.; Funt, J.M.; Abbey, D.; Issi, L.; Guiducci, C.; Martinez, D.A.; Delorey, T.; Li, B.y.; White, T.C.; Cuomo, C.; et al. The evolution of drug resistance in clinical isolates of Candida albicans. eLife 2015, 4, e00662. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Kravets, A.; Bethlendy, G.; Welle, S.; Rustchenko, E. Chromosome 5 monosomy of Candida albicans controls susceptibility to various toxic agents, including major antifungals. Antimicrob. Agents Chemother. 2013, 57, 5026–5036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osherov, N.; Kontoyiannis, D.P.; Romans, A.; May, G.S. Resistance to itraconazole in Aspergillus nidulans and Aspergillus fumigatus is conferred by extra copies of the A. nidulans P-450 14α-demethylase gene, pdmA. J. Antimicrob. Chemother. 2001, 48, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Jones, L.; Riaz, S.; Morales-Cruz, A.; Amrine, K.C.; McGuire, B.; Gubler, W.D.; Walker, M.A.; Cantu, D. Adaptive genomic structural variation in the grape powdery mildew pathogen, Erysiphe necator. BMC Genom. 2014, 15, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forche, A.; Solis, N.V.; Swidergall, M.; Thomas, R.; Guyer, A.; Beach, A.; Cromie, G.A.; Le, G.T.; Lowell, E.; Pavelka, N. Selection of Candida albicans trisomy during oropharyngeal infection results in a commensal-like phenotype. PLoS Genet. 2019, 15, e1008137. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, M.; Pontes, A.; Almeida, P.; Barbosa, R.; Serra, M.; Libkind, D.; Hutzler, M.; Gonçalves, P.; Sampaio, J.P. Distinct Domestication Trajectories in Top-Fermenting Beer Yeasts and Wine Yeasts. Curr. Biol. 2016, 26, 2750–2761. [Google Scholar] [CrossRef] [Green Version]
- Hope, E.A.; Amorosi, C.J.; Miller, A.W.; Dang, K.; Heil, C.S.; Dunham, M.J. Experimental evolution reveals favored adaptive routes to cell aggregation in yeast. Genetics 2017, 206, 1153–1167. [Google Scholar] [CrossRef] [Green Version]
- Hunter, A.J.; Jin, B.; Kelly, J.M. Independent duplications of α-amylase in different strains of Aspergillus oryzae. Fungal Genet. Biol. 2011, 48, 438–444. [Google Scholar] [CrossRef]
- Gibbons, J.G.; Salichos, L.; Slot, J.C.; Rinker, D.C.; McGary, K.L.; King, J.G.; Klich, M.A.; Tabb, D.L.; McDonald, W.H.; Rokas, A. The evolutionary imprint of domestication on genome variation and function of the filamentous fungus Aspergillus oryzae. Curr. Biol. 2012, 22, 1403–1409. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.K.; Liu, T.; Parreiras, L.S.; Williams, D.L.; Wohlbach, D.J.; Bice, B.D.; Ong, I.M.; Breuer, R.J.; Qin, L.; Busalacchi, D.; et al. Harnessing genetic diversity in Saccharomyces cerevisiae for fermentation of xylose in hydrolysates of alkaline hydrogen peroxide-pretreated biomass. Appl. Environ. Microbiol. 2014, 80, 540–554. [Google Scholar] [CrossRef] [Green Version]
- Gasch, A.P.; Huang, M.; Metzner, S.; Botstein, D.; Elledge, S.J.; Brown, P.O. Genomic expression responses to DNA-damaging agents and the regulatory role of the yeast ATR homolog Mec1p. Mol. Biol. Cell 2001, 12, 2987–3003. [Google Scholar] [CrossRef] [Green Version]
- De Kok, S.; Nijkamp, J.F.; Oud, B.; Roque, F.C.; de Ridder, D.; Daran, J.-M.; Pronk, J.T.; van Maris, A.J. Laboratory evolution of new lactate transporter genes in a jen1 Δ mutant of Saccharomyces cerevisiae and their identification as ADY2 alleles by whole-genome resequencing and transcriptome analysis. FEMS Yeast Res. 2012, 12, 359–374. [Google Scholar] [CrossRef] [PubMed]
- Sirr, A.; Cromie, G.A.; Jeffery, E.W.; Gilbert, T.L.; Ludlow, C.L.; Scott, A.C.; Dudley, A.M. Allelic variation, aneuploidy, and nongenetic mechanisms suppress a monogenic trait in yeast. Genetics 2015, 199, 247–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mount, H.O.C.; Revie, N.M.; Todd, R.T.; Anstett, K.; Collins, C.; Costanzo, M.; Boone, C.; Robbins, N.; Selmecki, A.; Cowen, L.E. Global analysis of genetic circuitry and adaptive mechanisms enabling resistance to the azole antifungal drugs. PLoS Genet. 2018, 14, e1007319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haugen, A.C.; Kelley, R.; Collins, J.B.; Tucker, C.J.; Deng, C.; Afshari, C.A.; Brown, J.M.; Ideker, T.; van Houten, B. Integrating phenotypic and expression profiles to map arsenic-response networks. Genome Biol. 2004, 5, R95. [Google Scholar] [CrossRef] [Green Version]
- Dunn, B.; Levine, R.P.; Sherlock, G. Microarray karyotyping of commercial wine yeast strains reveals shared, as well as unique, genomic signatures. BMC Genom. 2005, 6, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carreto, L.; Eiriz, M.F.; Gomes, A.C.; Pereira, P.M.; Schuller, D.; Santos, M.A. Comparative genomics of wild type yeast strains unveils important genome diversity. BMC Genom. 2008, 9, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naseeb, S.; Delneri, D. Impact of Chromosomal Inversions on the Yeast DAL Cluster. PLoS ONE 2012, 7, e42022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naumov, G.I.; James, S.A.; Naumova, E.S.; Louis, E.J.; Roberts, I.N. Three new species in the Saccharomyces sensu stricto complex: Saccharomyces cariocanus, Saccharomyces kudriavzevii and Saccharomyces mikatae. Int. J. Syst. Evol. Microbiol. 2000, 50, 1931–1942. [Google Scholar] [CrossRef] [PubMed]
- Dymond, J.S.; Richardson, S.M.; Coombes, C.E.; Babatz, T.; Muller, H.; Annaluru, N.; Blake, W.J.; Schwerzmann, J.W.; Dai, J.; Lindstrom, D.L. Synthetic chromosome arms function in yeast and generate phenotypic diversity by design. Nature 2011, 477, 471–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Li, Y.; Chen, X.; Ding, M.; Wu, Y.; Yuan, Y.-J. SCRaMbLE generates evolved yeasts with increased alkali tolerance. Microb. Cell Factories 2019, 18, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Luo, Z.; Wang, Y.; Pham, N.T.; Tuck, L.; Pérez-Pi, I.; Liu, L.; Shen, Y.; French, C.; Auer, M. Rapid pathway prototyping and engineering using in vitro and in vivo synthetic genome SCRaMbLE-in methods. Nat. Commun. 2018, 9, 1936. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Wang, L.; Wang, Y.; Zhang, W.; Guo, Y.; Shen, Y.; Jiang, L.; Wu, Q.; Zhang, C.; Cai, Y. Identifying and characterizing SCRaMbLEd synthetic yeast using ReSCuES. Nat. Commun. 2018, 9, 1930. [Google Scholar] [CrossRef]
- Blount, B.; Gowers, G.F.; Ho, J.; Ledesma-Amaro, R.; Jovicevic, D.; McKiernan, R.; Xie, Z.; Li, B.; Yuan, Y.; Ellis, T. Rapid host strain improvement by in vivo rearrangement of a synthetic yeast chromosome. Nat. Commun. 2018, 9, 1932. [Google Scholar] [CrossRef]
- Wang, J.; Xie, Z.-X.; Ma, Y.; Chen, X.-R.; Huang, Y.-Q.; He, B.; Jia, B.; Li, B.-Z.; Yuan, Y.-J. Ring synthetic chromosome V SCRaMbLE. Nat. Commun. 2018, 9, 3783. [Google Scholar] [CrossRef]
- Jia, B.; Wu, Y.; Li, B.-Z.; Mitchell, L.A.; Liu, H.; Pan, S.; Wang, J.; Zhang, H.-R.; Jia, N.; Li, B. Precise control of SCRaMbLE in synthetic haploid and diploid yeast. Nat. Commun. 2018, 9, 1933. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhu, R.-Y.; Mitchell, L.A.; Ma, L.; Liu, R.; Zhao, M.; Jia, B.; Xu, H.; Li, Y.-X.; Yang, Z.-M. In vitro DNA SCRaMbLE. Nat. Commun. 2018, 9, 1935. [Google Scholar] [CrossRef] [PubMed]
- Gowers, G.O.F.; Chee, S.M.; Bell, D.; Suckling, L.; Kern, M.; Tew, D.; McClymont, D.W.; Ellis, T. Improved betulinic acid biosynthesis using synthetic yeast chromosome recombination and semi-automated rapid LC-MS screening. Nat. Commun. 2020, 11, 868. [Google Scholar] [CrossRef] [PubMed]
- Bansal, V.; Boucher, C. Sequencing Technologies and Analyses: Where Have We Been and Where Are We Going? iScience 2019, 18, 37–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Phenotype | Gene(s) | Type of SV | Organism | References |
|---|---|---|---|---|
| Adaptation to: | ||||
| Glucose limitation | HXT6 and HXT7 | Increased copy number | S. cerevisiae | [31,246] |
| Sulfate limitation | SUL1 | Increased copy number | S. cerevisiae | [30,196] |
| Poor nitrogen sources (glutamine or glutamate) | GAP1 | Aneuploidies, nonreciprocal translocations, tandem duplication, complex CNVs | S. cerevisiae | [79,247] |
| Poor nitrogen sources (allantoin) | DAL4 | Increased copy number | S. cerevisiae | [139] |
| Poor nitrogen sources (urea) | DUR3 | Increased copy number | S. cerevisiae | [139] |
| Poor carbon sources (raffinose) 1 | Chr XIII duplication | S. cerevisiae | [64] | |
| Poor carbon sources (L-sorbose) | SOU1 | Chr V monosomy | C. albicans | [249] |
| High temperature (40 °C) | PAU5 and several genes introgressed from S. paradoxus | Increased copy number | S. cerevisiae | [148] |
| Elevated concentration of copper sulfate | CUP1, CUP2, SCO1, and SCO2 | Increased copy number | S. cerevisiae | [65,148,250,251,252] |
| Elevated concentration of sodium chloride | ENA5, MDM36, and VAR1589 introgressed from S. paradoxus | Increased copy number | S. cerevisiae | [148] |
| Elevated concentration of lithium chloride | ENA5 and VAR1589 | Increased copy number | S. cerevisiae | [148] |
| Sodium meta-arsenite | Chr XVI subtelomeric region containing ARR1, ARR2, and ARR3 genes | Increased copy number | S. cerevisiae | [65,148] |
| Nonpreferred carbon sources (galactose) | Two genes introgressed from S. paradoxus | Increased copy number | S. cerevisiae | [148] |
| Freeze–thaw cycles | AQY2 | Increased copy number | S. cerevisiae S. paradoxus | [182,253] |
| High temperature (39 °C) | HCM1, YCR016W, RRT12, YCR102C, and IMG2 | Increased copy number | S. cerevisiae | [195] |
| High temperature (42 °C) | Chr III segmental duplication and aneuploidy | S. cerevisiae | [254,255] | |
| High ethanol concentration | Chr III and Chr XII duplication | S. cerevisiae | [93,256] | |
| Sulfite | SSU1 | Translocations VIII-t-XVI XV-t-XVI | S. cerevisiae | [29,257,258,259,260] |
| Sulfite | SSU1 | Inversion in Chr XVI | S. cerevisiae | [261] |
| Nystatin | ADH4 and HXK2 | Increased copy number | S. cerevisiae | [148] |
| Fluconazole | ERG11 | Increased copy number | S. cerevisiae C. albicans C. neoformans | [194,262,263,264,265,266] |
| Fluconazole | TAC1, MRR1, CDR1 and CDR2 | Increased copy number | C. albicans | [262,266] |
| Fluconazole | AFR1, SEY1 and GLO3 | Increased copy number | C. neoformans | [263,264,265] |
| Fluconazole, amphotericin B, caspofungin | Chr V monosomy | C. albicans | [200,267] | |
| Itraconazole | pdmA | Increased copy number | Aspergillus fumigatus | [268] |
| Radicicol | STI1, PDR5 | Increased copy number | S. cerevisiae | [194] |
| Tunicamycin | ALG7, PRE7, YBR085C-A | Increased copy number | S. cerevisiae | [199] |
| 4-Nitroquinoline-N-oxide | ATR1 | Increased copy number | S. cerevisiae | [90] |
| Sterol demethylase inhibitors | Mutated CYP51 | Increased copy number | Erysiphe necator | [269] |
| Hydroxyurea, caspofungin | Chr II trisomy | C. albicans | [200] | |
| Inflammatory host response | Chr V or VI triplication | C. albicans | [270] | |
| Modulation of pathogenicity | Ave1, XLOC_009059, XLOC_008951, etc. | Translocations and inversions | Verticillium dahliae | [72] |
| Industrially relevant phenotype: | ||||
| Improved growth in maltose-containing medium | MAL1, MAL3, MPH2, MPH3, and YPR196W | Increased copy number | S. cerevisiae | [86,271] |
| Improved growth in galactose-containing medium | GAL7, GAL10, GAL1 | Increased copy number | S. cerevisiae | [187] |
| Desirable cell aggregation | FLO1, Lg-FLO1, FLO5 and FLO10 | Increased copy number | S. cerevisiae | [184,272] |
| Low diacetyl production | ILV5 and ILV3 | Increased copy number | S. pastorianus | [184] |
| Improved starch consumption | α-amylase gene | Increased copy number | Aspergillus oryzae | [273,274] |
| Increased ethanol yield | Chr XI duplication | S. cerevisiae | [28] | |
| Improved xylose fermentation in the presence of ferulic and p-coumaric acids | Chr XIV duplication | S. cerevisiae | [275] | |
| Compensation of mutation: | ||||
| Deletion of MEC1 | RNR1 | Increased copy number | S. cerevisiae | [276] |
| Deficiency of EST2 | PRP8, UTP9, KOG1, and SCH9 | Chr VIII monosomy | S. cerevisiae | [202] |
| Deletion of RNR1 | RNR3 | Increased copy number | S. cerevisiae | [206] |
| Deletion of RPS24A | RPS24B | Increased copy number | S. cerevisiae | [206] |
| Deletion of MYO2 | RLM1 | Increased copy number | S. cerevisiae | [91] |
| Deletion of JEN1 | ADY2 | Increased copy number + SNV | S. cerevisiae | [277] |
| Deletion of GAL7 | GAL80 | Increased copy number | S. cerevisiae | [278] |
| Deletion of RGD1 | NPR2 | Increased copy number | C. albicans | [279] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorkovskiy, A.; Verstrepen, K.J. The Role of Structural Variation in Adaptation and Evolution of Yeast and Other Fungi. Genes 2021, 12, 699. https://doi.org/10.3390/genes12050699
Gorkovskiy A, Verstrepen KJ. The Role of Structural Variation in Adaptation and Evolution of Yeast and Other Fungi. Genes. 2021; 12(5):699. https://doi.org/10.3390/genes12050699
Chicago/Turabian StyleGorkovskiy, Anton, and Kevin J. Verstrepen. 2021. "The Role of Structural Variation in Adaptation and Evolution of Yeast and Other Fungi" Genes 12, no. 5: 699. https://doi.org/10.3390/genes12050699
APA StyleGorkovskiy, A., & Verstrepen, K. J. (2021). The Role of Structural Variation in Adaptation and Evolution of Yeast and Other Fungi. Genes, 12(5), 699. https://doi.org/10.3390/genes12050699