
Intestinal Transcriptome Analysis Reveals Enrichment of Genes Associated with Immune and Lipid Mechanisms, Favoring Soybean Meal Tolerance in High-Growth Zebrafish (Danio Rerio)

 , , , , and
, , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Generation of Zebrafish Population
2.2. Fish Rearing and Experimental Diet
2.3. Growth Measurement and Intestinal Samples
2.4. Selection of Fish for RNA Sequencing
2.5. RNA Extraction and Library Preparation
2.6. Sequencing Data Processing and Differential Gene Expression
2.7. Gene Ontology and KEGG Eenrichment Analyses
2.8. Data Deposition
2.9. Histological Analysis
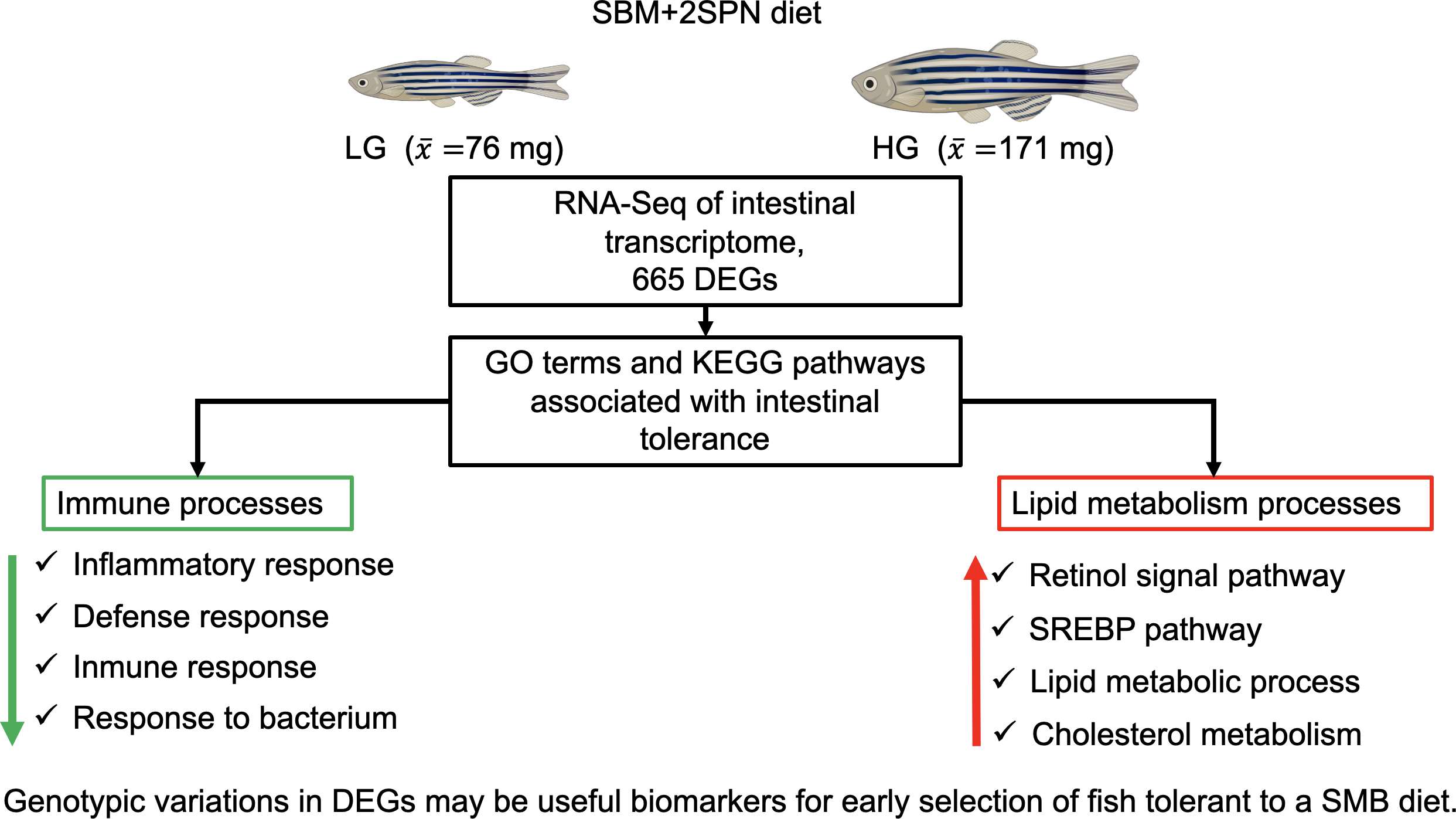
3. Results
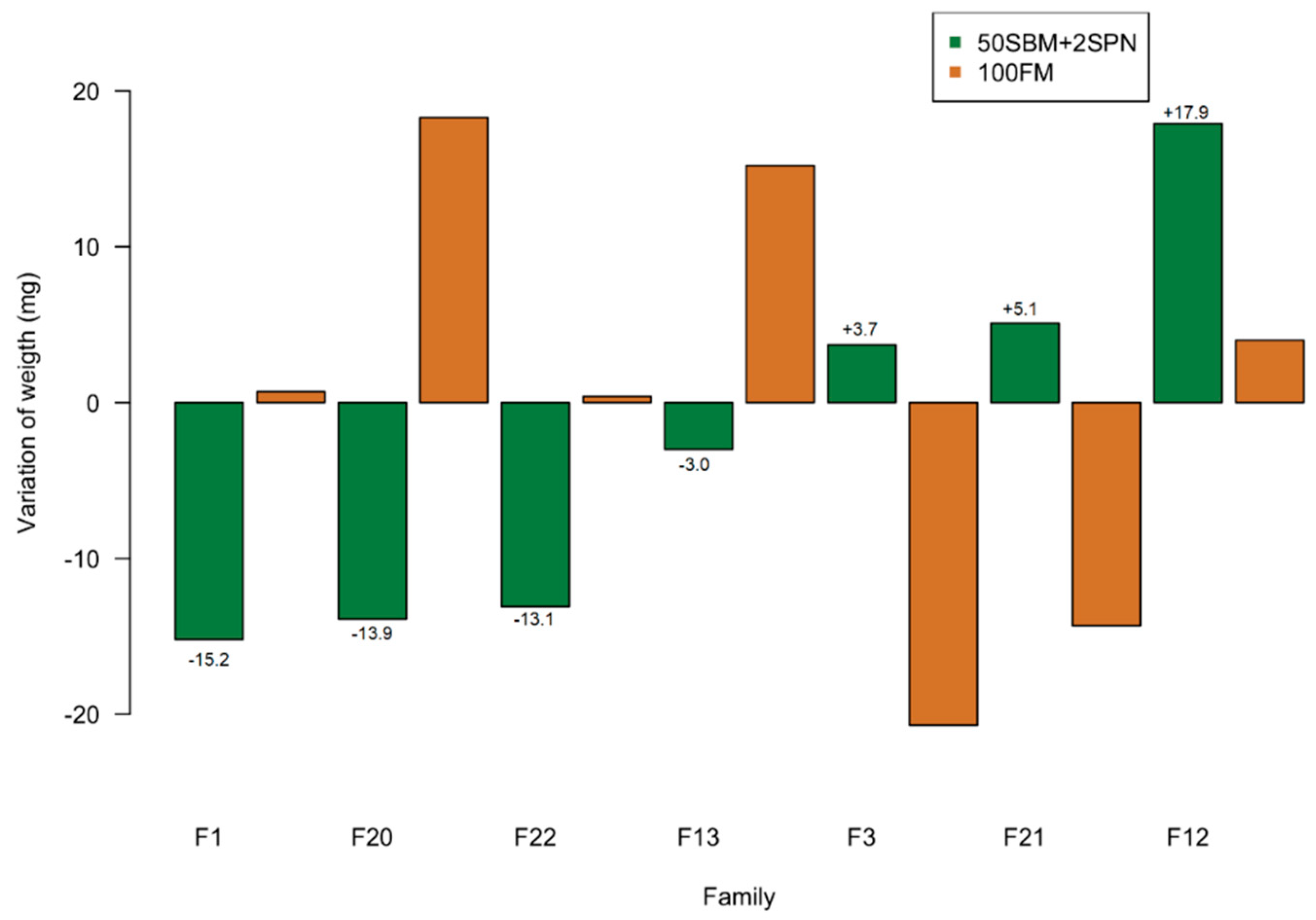
3.1. Growth Performance
3.2. Global Analysis of RNA Sequencing Data
3.3. Differential Gene Eexpression by Phenotype
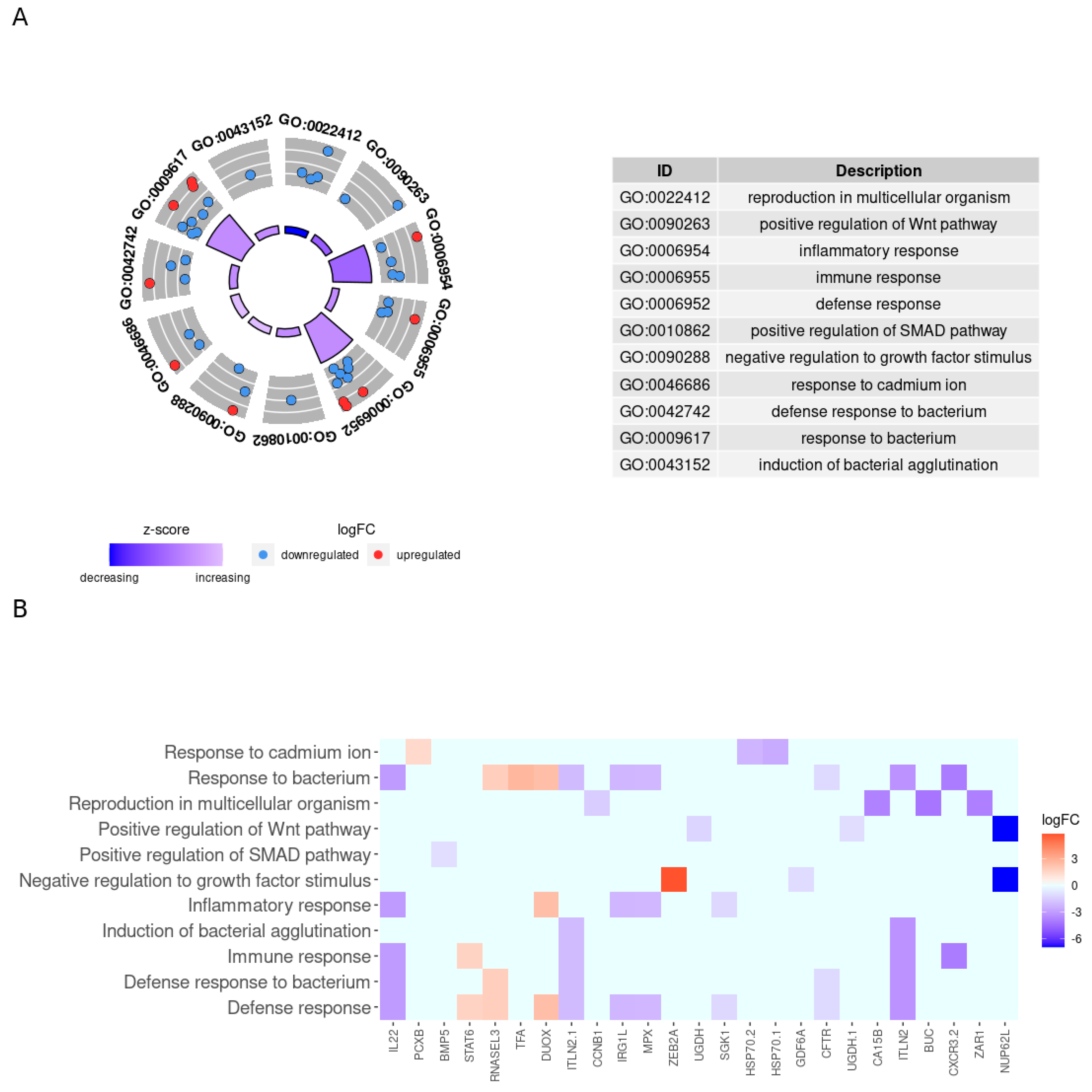
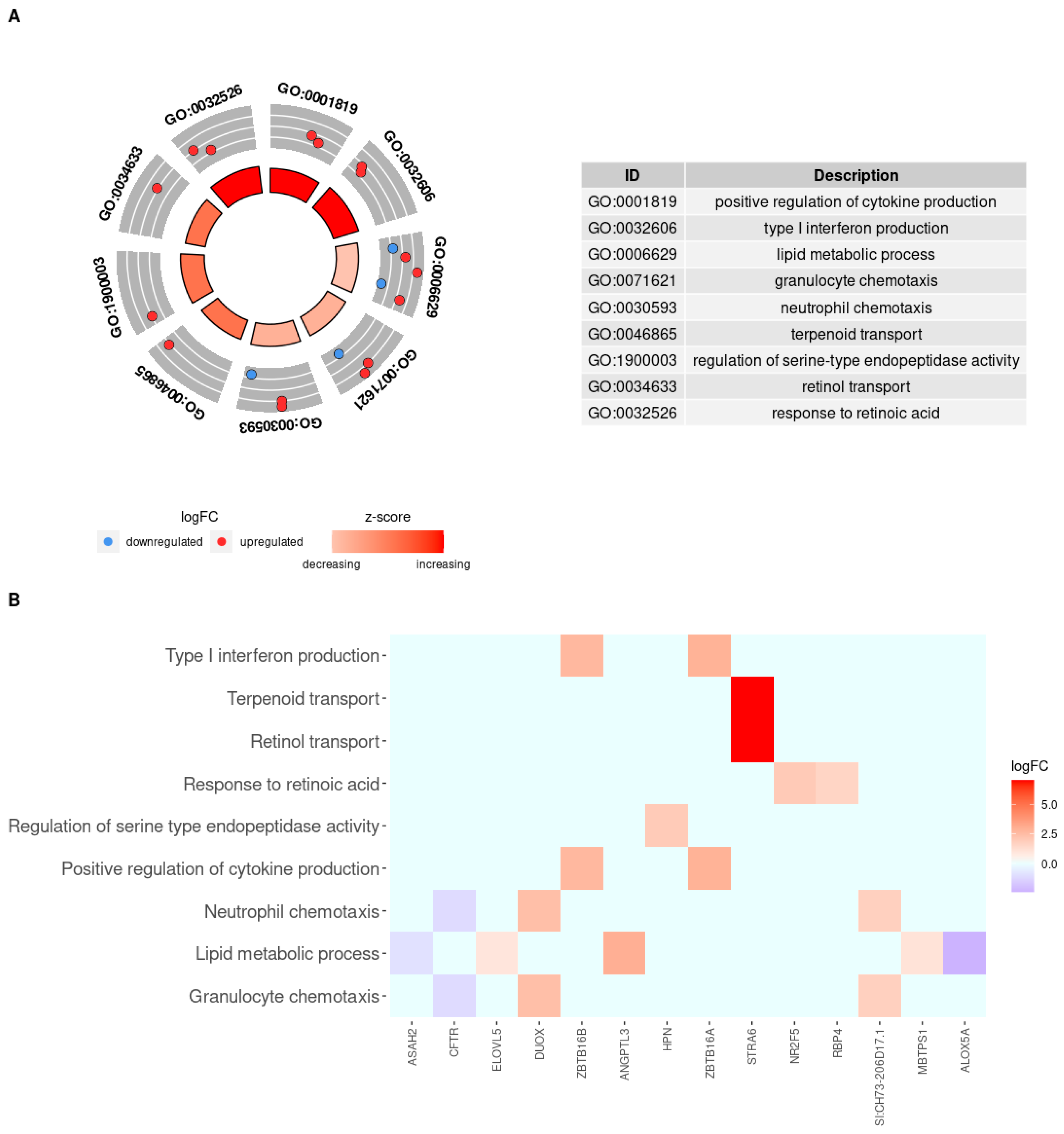
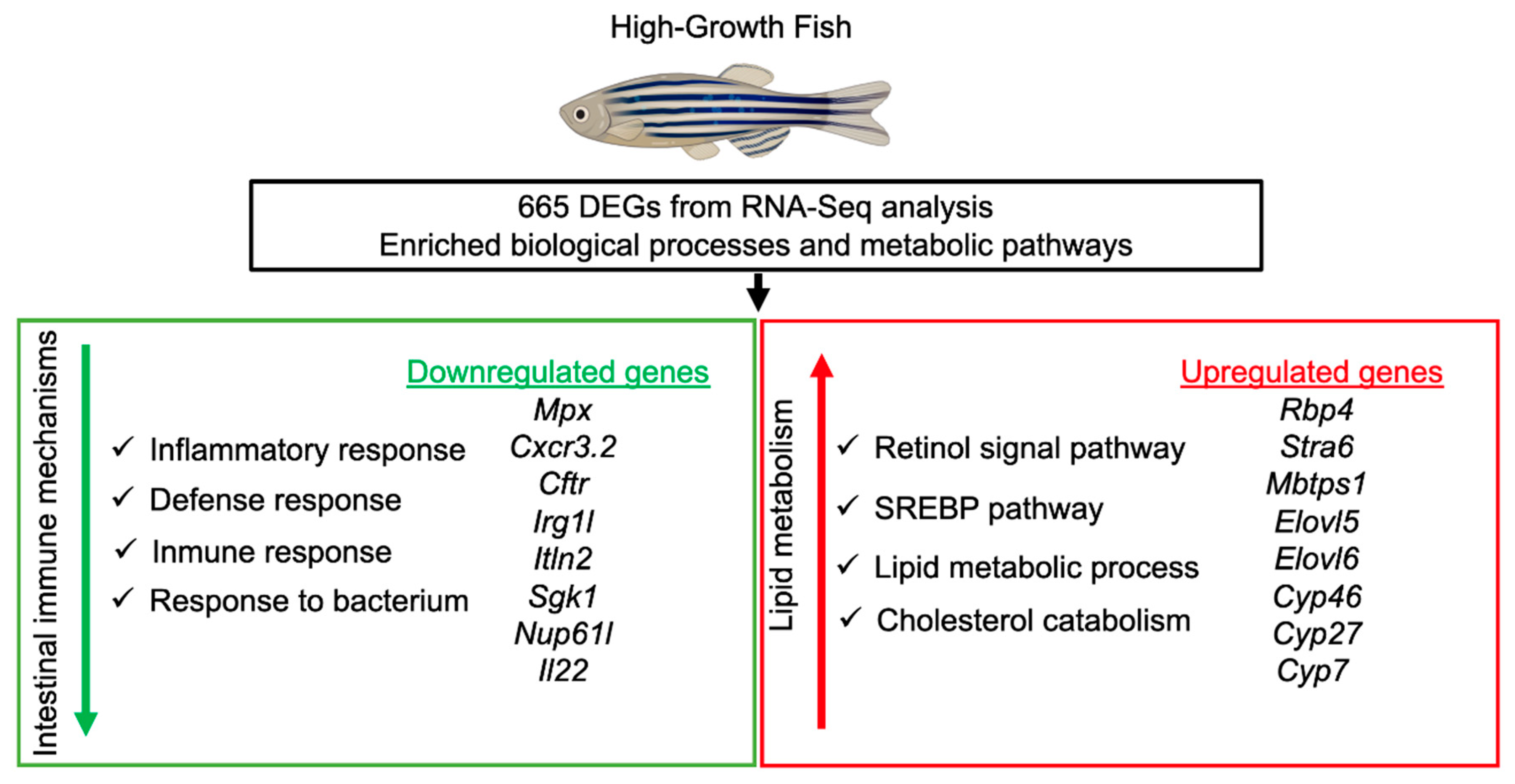
3.4. Enrichment Functional Analysis for Biological Processes
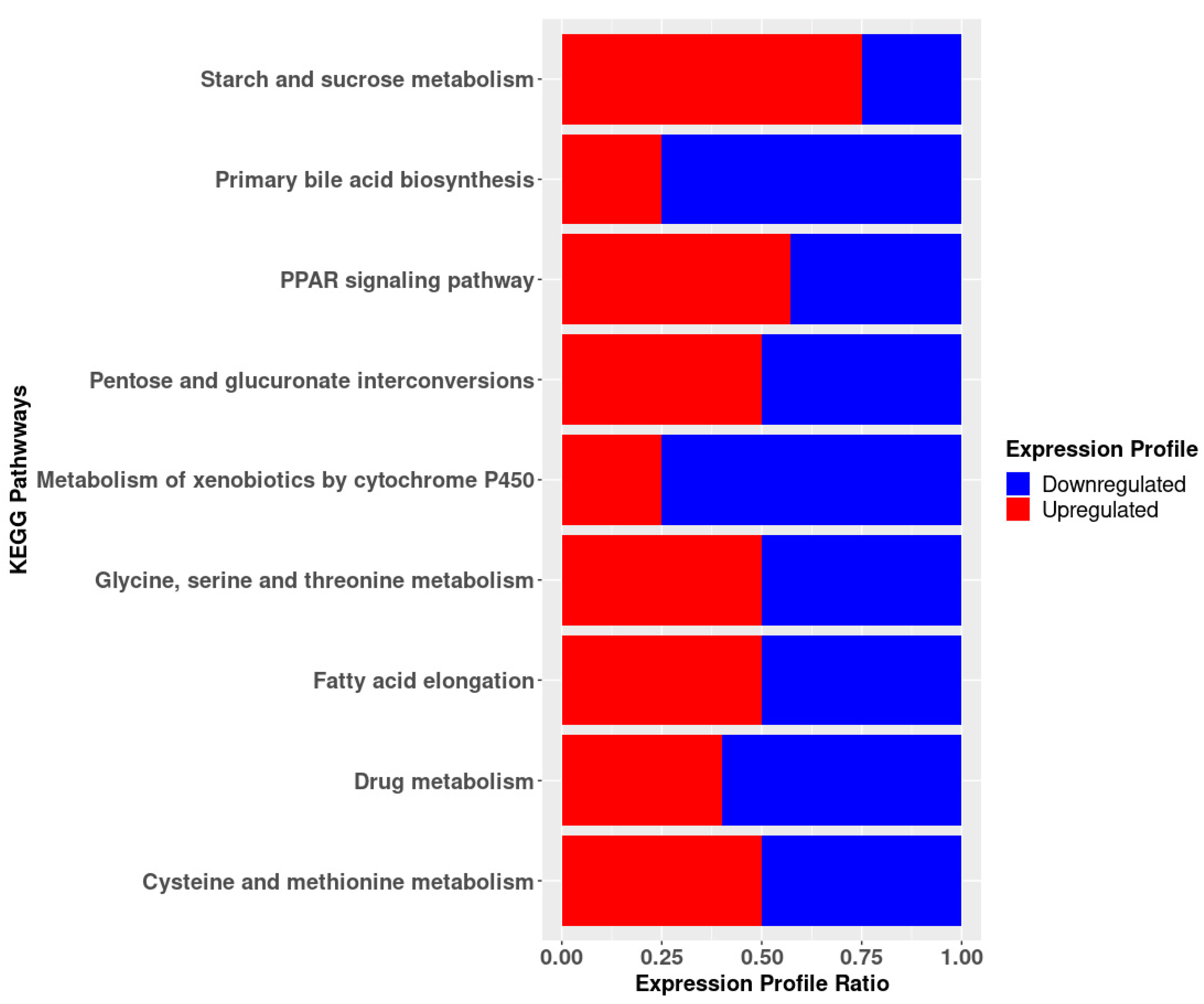
3.5. KEGG Pathway Enrichment Analysis
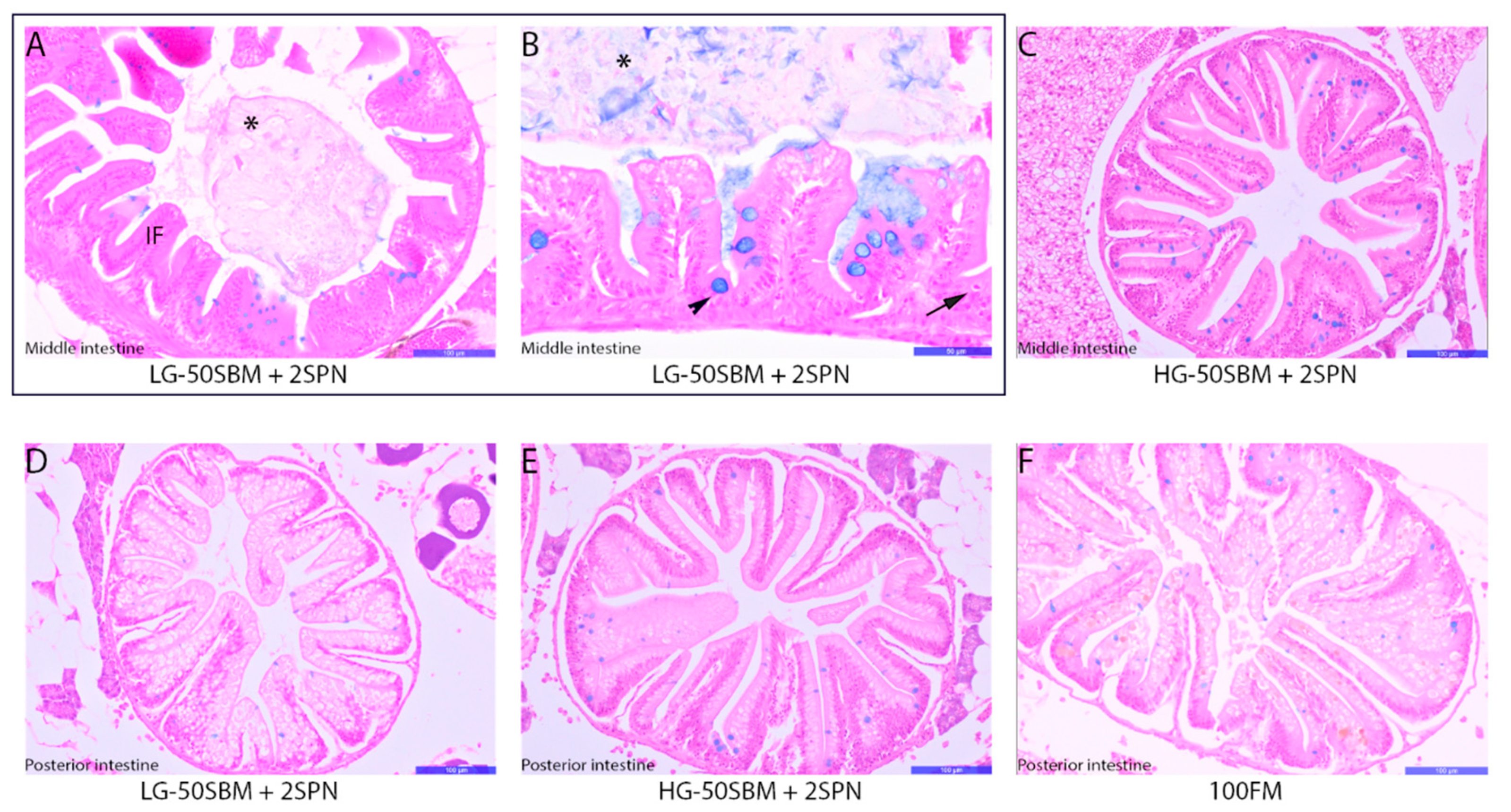
3.6. Intestinal Histology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020; FAO: Rome, Italy, 2020; ISSN 2663-8649. [Google Scholar] [CrossRef]
- Knudsen, D.; Urán, P.; Arnous, A.; Koppe, W.; Frokiaer, H. Saponin-Containing Subfractions of Soybean Molasses Induce Enteritis in the Distal Intestine of Atlantic Salmon. J. Agric. Food Chem. 2007, 55, 2261–2267. [Google Scholar] [CrossRef] [PubMed]
- Merrifield, D.L.; Dimitroglou, A.; Bradley, G.; Baker, R.T.M.; Davies, S.J. Soybean meal alters autochthonous microbial populations, microvilli morphology and compromises intestinal enterocyte integrity of rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish. Dis. 2009, 32, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Urán, P.A.; Goncalves, A.A.; Taverne-Thiele, J.J.; Schrama, J.W.; Verreth, J.A.J.; Rombout, J.H.W.M. Soybean meal induces intestinal inflammation in common carp (Cyprinus carpio L.). Fish Shellfish Immun. 2008, 25, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Hedrera, M.I.; Galdames, J.A.; Jimenez-Reyes, M.F.; Reyes, A.E.; Avendaño-Herrera, R.; Romero, J.; Feijóo, C.G. Soybean meal induces intestinal inflammation in zebrafish larvae. PLoS ONE 2013, 7, e69983. [Google Scholar] [CrossRef] [Green Version]
- Rumsey, G.L.; Siwicki, A.K.; Anderson, D.P.; Bowser, P.R. Effect of soybean protein on serological response, non-specific defense mechanisms, growth, and protein utilization in rainbow trout. Vet. Immunol. Immunopathol. 1994, 41, 323–339. [Google Scholar] [CrossRef]
- Król, E.; Douglas, A.; Tocher, D.R.; Crampton, V.O.; Speakman, J.R.; Secombes, C.J.; Martin, S.A.M. Differential responses of the gut transcriptome to plant protein diets in farmed Atlantic salmon. BMC Genom. 2016, 17, 156. [Google Scholar] [CrossRef] [Green Version]
- Krogdahl, A.; Gajardo, K.; Kortner, T.M.; Penn, M.; Gu, M.; Berge, G.M.; Bakke, A.M. Soya Saponins Induce Enteritis in Atlantic Salmon (Salmo salar L.). Agric. Food Chem. 2015, 63, 3887–3902. [Google Scholar] [CrossRef]
- Kortner, T.M.; Skugor, S.; Penn, M.H.; Mydland, T.L.; Djordjeviv, B.; Hillestad, M.; Krasnov, A.; Krogdahl, A. Dietary soyasaponin supplementation to pea protein concentrate reveals nutrigenomic interactions underlying enteropathy in Atlantic salmon (Salmo salar). BMC Vet. Res. 2012, 8, 101. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Requeni, P.; Mingarro, M.; Calduch-Giner, J.A.; Médale, F.; Martin, S.A.M.; Houlihan, D.F.; Kaushik, S.; Pérez-Sánchez, J. Protein growth performance performance, amino acid utilisation and somatotropic axis responsiveness to fish meal replacement by plant protein sources in gilthead sea bream (Sparus aurata). Aquaculture 2004, 232, 493–510. [Google Scholar] [CrossRef]
- Fontaínhas-Fernandes, A.; Gomes, E.; Reis-Henriques, M.A.; Coimbra, J. Replacement of fish meal by plant proteins in the diet of Nile tilapia: Digestibility and growth performance. Aquac. Int. 1999, 7, 57–67. [Google Scholar] [CrossRef]
- Médale, F.; Boujard, T.; Vallee, F.; Blanc, D.; Mambrini, M.; Roem, A.; Kaushik, S.J. Voluntary feed intake, nitrogen and phosphorus losses in rainbow trout (Oncorhynchus mykiss) fed increasing dietary levels of soy protein concentrate. Aquat. Living Resour. 1998, 11, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Vilhelmsson, O.T.; Martin, S.A.M.; Médale, F.; Kaushik, S.J.; Houlihan, D.F. Dietary plant-protein substitution affects hepatic metabolism in rainbow trout (Oncorhynchus mykiss). Br. J. Nutr. 2004, 92, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Mundheim, H.; Aksnes, A.; Hope, B. Feed efficiency and digestibility in salmon (Salmo salar) fed different dietary proportions of vegetable protein sources in combination with two fish meal qualities. Aquaculture 2004, 237, 315–331. [Google Scholar] [CrossRef]
- Ulloa, P.E.; Peña, A.A.; Lizama, C.D.; Araneda, C.; Iturra, P.; Neira, R.; Medrano, J.F. Growth response and expression of muscle growth-related candidate genes in adult zebrafish fed plant and fishmeal protein-based diets. Zebrafish 2013, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Krogdahl, A.; Bakke-McKellep, A.M.; Baeverjord, G. Effects of graded levels of standard soybean meal on intestinal structure, mucosal enzyme activities, and pancreatic response in Atlantic salmon (Salmo salar L.). Aquac. Nutr. 2003, 9, 361–371. [Google Scholar] [CrossRef]
- Overland, M.; Sorensen, M.; Storebakken, T.; Penn, M.; Krogdahl, A.; Skrede, A. Pea protein concentrate substituting fish meal or soybean meal in diets for Atlantic salmon (Salmo salar)—Effect on growth performance, nutrient digestibility, carcass composition, gut health, and physical feed quality. Aquaculture 2009, 288, 305–311. [Google Scholar] [CrossRef]
- Navarrete, P.; Fuentes, P.; De la Fuente, L.; Barros, L.; Magne, F.; Opazo, R.; Ibacache, C.; Espejo, R.; Romero, J. Short-term effects of dietary soybean meal and lactic acid bacteria on the intestinal morphology and microbiota of Atlantic salmon (Salmo salar). Aquac. Nutr. 2013, 19, 827–836. [Google Scholar] [CrossRef]
- Falconer, D.; MacKay, T. Introduction to Quantitative Genetics; Longman Group Ltd.: New York, NY, USA, 1996; ISBN-13: 978-0582243026. [Google Scholar]
- Von Hertell, U.; Hörstgen-Schwarrk, G.; Langholz, H.-J.; Jung, B. Family studies on genetic variability in growth and reproductive performance between and within test fish populations of the zebrafish, Brachydanio rerio. Aquaculture 1990, 85, 307–315. [Google Scholar] [CrossRef]
- Tave, D. Genetics for Fish Hatchery Managers; Springer: New York, NY, USA, 1993; ISBN 978-0-442-00417-0. [Google Scholar]
- Qian, X.; Ba, Y.; Zhuang, Q.; Zhong, G. RNA-seq technology and its application in fish transcriptomics. OMICS 2014, 18, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Sahlmann, C.; Sutherland, B.J.G.; Kortner, T.M.; Koop, B.F.; Krogdahl, A.; Bakke, A.M. Early response of gene expression in the distal intestine of Atlantic salmon (Salmo salar) during the development of soybean meal induced enteritis. Fish Shellfish Immun. 2013, 34, 599–609. [Google Scholar] [CrossRef]
- De Santis, C.; Bartie, K.L.; Olsen, R.E.; Taggart, J.B.; Tocher, D.R. Nutrigenomic profiling of transcriptional processes affected in liver and distal intestine in response to a soybean meal-induced nutritional stress in Atlantic salmon (Salmo salar). Comp. Biochem. Physiol. Part D Genom. Proteom. 2015, 15, 1–11. [Google Scholar] [CrossRef]
- Balasubramanian, M.N.; Panserat, S.; Dupont-Nivet, M.; Quillet, E.; Montfort, J.; Le Cam, A.; Medale, F.; Kaushik, S.J.; Geurden, I. Molecular pathways associated with the nutritional programming of plant-based diet acceptance in rainbow trout following an early feeding exposure. BMC Genom. 2016, 13, 17–449. [Google Scholar] [CrossRef] [Green Version]
- Kemski, M.M.; Rappleye, C.A.; Dabrowski, K.; Bruno, R.S.; Wick, M. Transcriptomic response to soybean meal-based diets as the first formulated feed in juvenile yellow perch (Perca flavescens). Sci Rep. 2020, 4, 3998. [Google Scholar] [CrossRef] [Green Version]
- Ulloa, P.E.; Medrano, J.F.; Feijoo, C.G. Zebrafish as animal model for aquaculture nutrition research. Front. Genet. 2014, 5, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Ribas, L.; Piferrer, F. The zebrafish (Danio rerio) as a model organism, with emphasis on applications for finfish aquaculture research. Rev. Aquac. 2013, 5, 1–32. [Google Scholar] [CrossRef]
- Lee-Estevez, M.; Figueroa, E.; Cosson, J.; Short, S.E.; Valdebenito, I.; Ulloa-Rodriguez, P.; Farias, J. Zebrafish as a useful model for immunological research with potential applications in aquaculture. Rev. Aquac. 2016, 10, 213–223. [Google Scholar] [CrossRef]
- López-Nadal, A.; Ikeda-Ohtsubo, W.; Sipkema, D.; Peggs, D.; McGurk, C.; Forlenza, M.; Wiegertjes, G.F.; Brugman, S. Feed, Microbiota, and Gut Immunity: Using the Zebrafish Model to Understand Fish Health. Front. Immunol. 2020, 5, 11–114. [Google Scholar] [CrossRef] [PubMed]
- Ulloa, P.E.; Iturra, P.; Neira, R.; Araneda, C. Zebrafish as a model organism for nutrition and growth: Towards comparative studies of nutritional genomics applied to aquacultured fishes. Rev. Fish Biol. Fish. 2011, 21, 649–666. [Google Scholar] [CrossRef]
- Ulloa, P.E.; Rincón, G.; Islas-Trejo, A.; Araneda, C.; Iturra, P.; Neira, R.; Medrano, J.F. RNA Sequencing to Study Gene Expression and SNP Variations Associated with Growth in Zebrafish Fed a Plant Protein-Based Diet. Mar. Biotechnol. 2015, 17, 353–363. [Google Scholar] [CrossRef]
- Coronado, M.; Solis, C.J.; Hernandez, P.P.; Feijóo, C.G. Soybean Meal-Induced Intestinal Inflammation in Zebrafish Is T Cell-Dependent and Has a Th17 Cytokine Profile. Front. Immunol. 2019, 10, 610. [Google Scholar] [CrossRef] [PubMed]
- Ulloa, P.E.; Solís, C.J.; De La Paz, J.F.; Alaurent, T.G.S.; Caruffo, M.; Hernández, A.J.; Dantagnan, P.; Feijóo, C.G. Lactoferrin Decreases the Intestinal Inflammation Triggered by a Soybean Meal-Based Diet in Zebrafish. J. Immunol. Res. 2016, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Tello, K.; Ehrenfeld, N.; Solís, C.J.; Ulloa, P.E.; Hedrera, M.; Pizarro-Guajardo, M.; Pizarro-Guajardo, M.; Paredes-Sabja, D.; Feijóo, C.G. Effect of microalgae on intestinal inflammation triggered by soybean meal and bacterial infection in zebrafish. PLoS ONE 2017, 12, e0187696. [Google Scholar] [CrossRef]
- Ireland, P.A.; Dziedzic, S.Z.; Kearsley, M.W. Saponin content of soya and some commercial soya products by means of highperformance liquid chromatography of the sapogenins. J. Sci. Food Agric. 1986, 37, 694–698. [Google Scholar] [CrossRef]
- [NRC] National Research Council. Nutritional Requirements of Fish; National Academies Press: Washington, DC, USA, 1993; ISBN 978-0-309-04891-0. [Google Scholar]
- Brand, M.; Granato, M.; Nusslein-Volhard, C. Keeping and raising zebrafish. In Zebrafish—A Practical Approach; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Robison, B.D.; Drew, R.E.; Murdoch, G.K.; Powell, M.; Rodnick, K.J.; Settles, M.; Stone, D.; Churchill, D.E.; Hill, R.A.; Papasani, M.; et al. Sexual dimorphism in hepatic gene expression and the response to dietary carbohydrate manipulation in the zebrafish (Danio rerio). Comp. Biochem. Physiol. Part D Genom. Proteom. 2008, 3, 141–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimoda, N.; Knapik, E.W.; Ziniti, J.; Sim, C.; Yamada, E.; Kaplan, S.; Jackson, D.; De Sauvage, F.; Jacob, H.; Fishman, M.C. Zebrafish genetic map with 2000 microsatellite markers. Genomics 1999, 58, 219–232. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Wagner, G.P.; Kin, K.; Lynch, V.J. Measurement of mRNA abundance using RNA-seq data: RPKM measure is inconsistent among samples. Theory Biosci. 2012, 131, 281–285. [Google Scholar] [CrossRef]
- Ferreira, P.M.F.; Caldas, D.W.; Salaro, A.L.; Sartori, S.R.; Oliveira, J.M.; Cardoso, J.S.; Zuanon, A.S. Intestinal and liver morphometry of the Yellow Tail Tetra (Astyanax altiparanae) fed with oregano oil. An. Acad. Brasil. Ciências 2016, 88, 911–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamidian, G.; Zirak, K.; Sheikhzadeh, N.; Khani, O.A.; Shabanzadeh, S.; Divband, B. Intestinal histology and stereology in rainbow trout (Oncorhynchus mykiss) administrated with nanochitosan/zeolite and chitosan/zeolite composites. Aquac. Res. 2018, 49, 1803–1815. [Google Scholar] [CrossRef]
- Nasiadka, A.; Clark, M. Zebrafish Breeding in the Laboratory Environment. ILAR J. 2012, 53, 161–168. [Google Scholar] [CrossRef] [Green Version]
- De Souza, H.S.; Fiocchi, C. Immunopathogenesis of IBD: Current state of the art. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, Z. Inflammatory bowel disease related innate immunity and adaptive immunity. Am. J. Transl. Res. 2006, 8, 2490–2497. [Google Scholar]
- Boeckxstaens, G. Mast cells and inflammatory bowel disease. Curr. Opin. Pharmacol. 2015, 25, 45–49. [Google Scholar] [CrossRef]
- Raphael, I.; Nalawade, S.; Eagar, T.N.; Forsthuber, T.G. T cell subsets and their signature cytokines in autoimmune and inflammatory diseases. Cytokine 2015, 74, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Fang, X.; Ma, N.; Lin, Q.; Huang, Z.; Liu, X.M.; Chen, X.; Zhang, W.; Zhang, Y. Myeloperoxidase-deficient zebrafish show an augmented inflammatory response to challenge with Candida albicans. Fish Shellfish Immun. 2015, 44, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.C.; Boyle, R.H.; Sun, X.; Wicker, S.M.; Misa, J.P.; Krissansen, G.W.; Print, C.G.; Crosier, K.E.; Crosier, P.S. Epidermal cells help coordinate leukocyte migration during inflammation through fatty acid-fuelled matrix metalloproteinase production. Nat. Commun. 2014, 5, 3880. [Google Scholar] [CrossRef] [Green Version]
- Torraca, V.; Cui, C.; Boland, R.; Bebelman, J.P.; Van der Sar, A.M.; Smit, M.J.; Siderius, M.; Spaink, H.P.; Meijer, A.H. The CXCR3-CXCL11 signaling axis mediates macrophage recruitment and dissemination of mycobacterial infection. Dis. Model. Mech. 2015, 8, 253–269. [Google Scholar] [CrossRef] [Green Version]
- Navis, A.; Marjoram, L.; Bagnat, M. Cftr controls lumen expansion and function of Kupffer’s vesicle in zebrafish. Development 2013, 140, 1703–1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudakov, J.A.; Hanash, A.M.; van den Brink, M.R.M. “Interleukin-22: Immunobiology and pathology”. Annu. Rev. Immunol. 2015, 33, 747–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleksejeva, E.; Houel, A.; Briolat, V.; Levraud, J.P.; Langevin, C.; Boudinot, P. Zebrafish Plzf transcription factors enhance early type I IFN response induced by two non-enveloped RNA viruses. Dev. Comp. Immunol. 2016, 57, 48–56. [Google Scholar] [CrossRef]
- Flores, M.V.; Crawford, K.C.; Pullin, L.M.; Hall, C.J.; Crosier, K.E.; Crosier, P.S. Dual oxidase in the intestinal epithelium of zebrafish larvae has anti-bacterial properties. Biochem. Biophys. Res. Commun. 2010, 400, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Krogdahl, Å.; Penn, M.; Thorsen, J.; Refstie, S.; Bakke, A.M. Important antinutrients in plant feedstuffs for aquaculture: An update on recent findings regarding responses in salmonids. Aquac. Res. 2010, 41, 333–344. [Google Scholar] [CrossRef]
- Krogdahl, Å.; Kortner, T.M.; Jaramillo-Torres, A.; Gamil, A.A.A.; Chikwati, E.; Li, Y.; Schmidt, M.; Herman, E.; Hymowitz, T.; Teimouri, S.; et al. Removal of three proteinaceous antinutrients from soybean does not mitigate soybean-induced enteritis in Atlantic salmon (Salmo salar, L). Aquaculture 2020, 514, 734495. [Google Scholar] [CrossRef]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 8, 15–73. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Tong, Q.; Chang, J.; Yu, J.; Li, S.; Niu, H.; Ma, D. Gut bacterial communities in the freshwater snail Planorbella trivolvis and their modification by a non-herbivorous diet. PeerJ 2021, 9, e10716. [Google Scholar] [CrossRef]
- Jiang, H.Y.; Ma, J.E.; Li, J.; Zhang, X.J.; Li, L.M.; He, N.; Liu, H.Y.; Luo, S.Y.; Wu, Z.J.; Han, R.C.; et al. Diets Alter the Gut Microbiome of Crocodile Lizards. Front. Microbiol. 2017, 8, 2073. [Google Scholar] [CrossRef]
- Osborne, T.B.; Mendel, L.B. The relation of growth to the chemical constituents of diet. J. Biol. Chem. 1913, 15, 311–326. [Google Scholar] [CrossRef]
- Noy, N. Retinoid-binding proteins: Mediators of retinoid action. Biochem. J. 2000, 348, 481–495. [Google Scholar] [CrossRef]
- Kawaguchi, R.; Yu, J.; Honda, J.; Hu, J.; Whitelegge, J.; Ping, P.; Wiita, P.; Bok, D.; Sun, H. A membrane receptor for retinol binding protein mediates cellular uptake of vitamin A. Science 2007, 315, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.C.; Jin, H.; Majumdar, A.; Noy, N. Signaling by vitamin A and retinol-binding protein regulates gene expression to inhibit insulin responses. Proc. Natl. Acad. Sci. USA 2011, 108, 4340–4345. [Google Scholar] [CrossRef] [Green Version]
- Grygiel-Górniak, B. Peroxisome proliferator-activated receptors and their ligands: Nutritional and clinical implications—A review. Nutr. J. 2014, 13, 17. [Google Scholar] [CrossRef] [Green Version]
- Christofides, A.; Konstantinidou, E.; Jani, C.; Boussiotis, V.A. The role of peroxisome proliferator-activated receptors (PPAR) in immune responses. Metabolism 2020, 114, 154338. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.K. Lipids, inflammasomes, metabolism, and disease. Immunol. Rev. 2020, 297, 108–122. [Google Scholar] [CrossRef]
- Raghow, R.; Yellaturu, C.; Deng, X.; Park, E.A.; Elam, M.B. SREBPs: The cross-roads of physiological and pathological lipid homeostasis. Trends Endocrinol. Metab. 2008, 19, 65e73. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.S.; Goldstein, J.L. The SREBP pathway: Regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 1997, 89, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Sakai, J.; Nohturfft, A.; Goldstein, J.L.; Brown, M.S. Cleavage of sterol regulatory element-binding proteins (SREBPs) at site-1 requires interaction with SREBP cleavage-activating protein. Evidence from in vivo competition studies. J. Biol. Chem. 1998, 273, 5785–5793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, P.T.; Kuah, M.K.; Chew, Y.S.; The, H.Y.; Shu-Chien, A.C. The requirements for sterol regulatory element-binding protein (Srebp) and stimulatory protein 1 (Sp1)-binding elements in the transcriptional activation of two freshwater fish Channa striata and Danio rerio elovl5 elongase. Fish Physiol. Biochem. 2020, 46, 1349–1359. [Google Scholar] [CrossRef]
- Zheng, X.; Tocher, D.R.; Dickson, C.A.; Bell, J.G.; Teale, A.J. Effects of diets containing vegetable oil on expression of genes involved in highly unsaturated fatty acid biosynthesisin liver of Atlantic salmon (Salmo salar). Aquaculture 2004, 236, 467–483. [Google Scholar] [CrossRef]
- Zheng, X.; Torstensen, B.E.; Tocher, D.R.; Dick, J.R.; Henderson, R.J.; Bell, J.G. Environmental and dietary influences of highly unsaturated fatty acid biosynthesis and expression offatty acyl desaturase and elongase genes in liver of Atlantic salmon (Salmo salar). Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1734, 13–24. [Google Scholar] [CrossRef]
- Moon, Y.A.; Ochoa, C.R.; Mitsche, M.A.; Hammer, R.E.; Horton, J.D. Deletion of ELOVL6 blocks the synthesis of oleic acid but does not prevent the development of fatty liver or insulin resistance. J. Lipid Res. 2014, 55, 2597–2605. [Google Scholar] [CrossRef] [Green Version]
- Oishi, Y.; Spann, N.J.; Link, V.M.; Muse, E.D.; Strid, T.; Edillor, C.; Kolar, M.J.; Matsuzaka, T.; Hayakawa, S.; Tao, J.; et al. SREBP1 Contributes to Resolution of Pro-inflammatory TLR4 Signaling by Reprogramming Fatty Acid Metabolism. Cell Metab. 2017, 25, 412–427. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.; Finlay, D.K. Diverse Immunoregulatory Roles of Oxysterols—The Oxidized Cholesterol Metabolites. Metabolites 2020, 10, 384. [Google Scholar] [CrossRef] [PubMed]
- Owein, G.-L.; Mutemberezi, V.; Muccioli, G.G. Oxysterols in Metabolic Syndrome: From Bystander Molecules to Bioactive Lipids. Trends Mol. Med. 2016, 22, 594–614. [Google Scholar] [CrossRef]
- Mutemberezi, V.; Guillemot-Legris, O.; Muccioli, G.G. Oxysterols: From cholesterol metabolites to key mediators. Prog. Lipid Res. 2016, 64, 152–169. [Google Scholar] [CrossRef]
- Soroosh, P.; Wu, J.; Xue, X.; Song, J.; Sutton, S.W.; Sablad, M.; Yu, J.; Nelen, M.I.; Liu, X.; Castro, G. Oxysterols are agonist ligands of RORγt and drive Th17 cell differentiation. Proc. Natl. Acad. Sci. USA 2014, 111, 12163–12168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abernathy, J.; Brezas, A.; Snekvik, K.R.; Hardy, R.W.; Overturf, K. Integrative functional analyses using rainbow trout selected for tolerance to plant diets reveal nutrigenomic signatures for soy utilization without the concurrence of enteritis. PLoS ONE 2017, 12, e0180972. [Google Scholar] [CrossRef]
- Ding, Y.; Ai, C.; Mu, Y.; Ao, J.; Chen, X. Molecular characterization and evolution analysis of five interleukin-17 receptor genes in large yellow croaker Larimichthys crocea. Fish Shellfish Immunol. 2016, 58, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Lazzarotto, V.; Médale, F.; Larroquet, L.; Corraze, G. Long-term dietary replacement of fishmeal and fish oil in diets for rainbow trout (Oncorhynchus mykiss): Effects on growth, whole body fatty acids and intestinal and hepatic gene expression. PLoS ONE 2018, 13, e0190730. [Google Scholar] [CrossRef] [Green Version]
- Geay, F.; Ferraresso, S.; Zambonino-Infante, J.L.; Bargelloni, L.; Quentel, C.; Vandeputte, M.; Kaushik, S.; Cahu, C.L.; Mazurais, D. Effects of the total replacement of fish-based diet with plant-based diet on the hepatic transcriptome of two European sea bass (Dicentrarchus labrax) half-sibfamilies showing different growth rates with the plant-based diet. BMC Genom. 2011, 23, 12–522. [Google Scholar] [CrossRef] [Green Version]
- Maita, M.; Maekawa, J.; Satoh, K.I.; Futami, K.; Satoh, S. Disease resistance and hypocholesterolemia in yellowtail Seriola quinqueradiata fed a non-fishmeal diet. Fish. Sci. 2006, 72, 513–519. [Google Scholar] [CrossRef]
- De Santis, C.; Crampton, V.O.; Bicskei, B.; Tocher, D.R. Replacement of dietary soy- with air classified faba bean protein concentrate alters the hepatic transcriptome in Atlantic salmon (Salmo salar) parr. Comp. Biochem. Physiol. D Genom. Proteom. 2015, 16, 48–58. [Google Scholar] [CrossRef]
- Sasabe, J.; Suzuki, M. Emerging role of d-amino acid metabolism in the innate defense. Front. Microbiol. 2018, 9, 933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, N.; Solt, L.A.; Wang, Y.; Rogers, P.M.; Bhattacharyya, G.; Kamenecka, T.M.; Stayrook, K.R.; Crumbley, C.; Floyd, E.Z.; Gimble, J.M.; et al. Regulation of Adipogenesis by Natural and Synthetic REV-ERB Ligands. Endocrinology 2010, 151, 3015–3025. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Navarrete, J.M.; Rodríguez, A.; Ortega, F.; Becerril, S.; Girones, J.; Sabater-Masdeu, M.; Latorre, J.; Ricart, W.; Frühbeck, G.; Fernández-Real, J.M. Heme biosynthetic pathway is functionally linked to adipogenesis via mitochondrial respiratory activity. Obesity 2017, 25, 1723–173375. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Chen, H.-M.; Qian, X.-Q.; Gui, J.-F. Transcriptome analysis of grass carp (Ctenopharyngodon idella) between fast-and slow-growing fish. Comp. Biochem. Phys. D Genom. Proteom. 2020, 35, 100688. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients, G/Kg | Control Diet | Experimental Diet |
|---|---|---|
| 50SBM + 2SPN | 100FM | |
| Fishmeal | 610 | 250 |
| Soybean Meal | 0 | 500 |
| Wheat Meal | 255 | 113 |
| Starch | 45 | 45 |
| Fish Oil | 30 | 60 |
| Vitamins 1 | 15 | 15 |
| Minerals 1 | 15 | 15 |
| Cellulose | 30 | 0 |
| Soy Saponin | 0 | 2 |
| Total | 1000 | 1000 |
| Analytical Composition * (Dry Basis) | ||
| Dry Material (%) | 95.3 | 93.5 |
| Digestible Protein (%) | 46.4 | 43.5 |
| Digestible Lipids (%) | 7.8 | 8.4 |
| Fiber (%) | 2.5 | 1.1 |
| Ash (%) | 12.6 | 9.7 |
| Non-Nitrogenous Extract (%) | 30.5 | 37.1 |
| Gross Energy (MJ/Kg) ** | 20.0 | 20.3 |
| Family | Familial Mean Weight Gain (Mg) | Individuals Selected For RNA-Seq | Weight Gain (Mg) | Mean Weight Gain of Individuals Selected (Mg) | Phenotype |
|---|---|---|---|---|---|
| F3 (n = 12) | 124.2 ± 32 | F3-7 | 162.3 | 171.5 ± 18.1 | Higher growth (HG) |
| F3-9 | 183.0 | ||||
| F12 (n = 10) | 137.6 ± 31 | F12-8 | 196.5 | ||
| F21 (n = 7) | 125.7 ± 35 | F21-1 | 150.6 | ||
| F21-7 | 165.5 | ||||
| F1 (n = 9) | 105.8 ± 33 | F1-8 | 79.4 | 76.4 ± 10.3 | Lower growth (LG) |
| F20 (n = 7) | 106.8 ± 48 | F20-4 | 62.6 | ||
| F22 (n = 6) | 108 ± 30 | F22-3 | 87.3 | ||
| F22-5 | 62.7 | ||||
| F13 (n = 8) | 117 ± 28 | F13-1 | 77.9 | ||
| F13-6 | 88.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valenzuela, L.; Pacheco, S.; Rincón, G.; Pavez, L.; Lam, N.; Hernández, A.J.; Dantagnan, P.; González, F.; Jilberto, F.; Ravanal, M.C.; et al. Intestinal Transcriptome Analysis Reveals Enrichment of Genes Associated with Immune and Lipid Mechanisms, Favoring Soybean Meal Tolerance in High-Growth Zebrafish (Danio Rerio). Genes 2021, 12, 700. https://doi.org/10.3390/genes12050700
Valenzuela L, Pacheco S, Rincón G, Pavez L, Lam N, Hernández AJ, Dantagnan P, González F, Jilberto F, Ravanal MC, et al. Intestinal Transcriptome Analysis Reveals Enrichment of Genes Associated with Immune and Lipid Mechanisms, Favoring Soybean Meal Tolerance in High-Growth Zebrafish (Danio Rerio). Genes. 2021; 12(5):700. https://doi.org/10.3390/genes12050700
Chicago/Turabian StyleValenzuela, Luis, Sebastian Pacheco, Gonzalo Rincón, Leonardo Pavez, Natalia Lam, Adrián J. Hernández, Patricio Dantagnan, Felipe González, Felipe Jilberto, M. Cristina Ravanal, and et al. 2021. "Intestinal Transcriptome Analysis Reveals Enrichment of Genes Associated with Immune and Lipid Mechanisms, Favoring Soybean Meal Tolerance in High-Growth Zebrafish (Danio Rerio)" Genes 12, no. 5: 700. https://doi.org/10.3390/genes12050700
APA StyleValenzuela, L., Pacheco, S., Rincón, G., Pavez, L., Lam, N., Hernández, A. J., Dantagnan, P., González, F., Jilberto, F., Ravanal, M. C., Ramos, C., Garcia, H., Araneda, C., & Ulloa, P. E. (2021). Intestinal Transcriptome Analysis Reveals Enrichment of Genes Associated with Immune and Lipid Mechanisms, Favoring Soybean Meal Tolerance in High-Growth Zebrafish (Danio Rerio). Genes, 12(5), 700. https://doi.org/10.3390/genes12050700