
The Crazy Ovary


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. The Initiation of Follicular Growth: The Hippo Signaling System
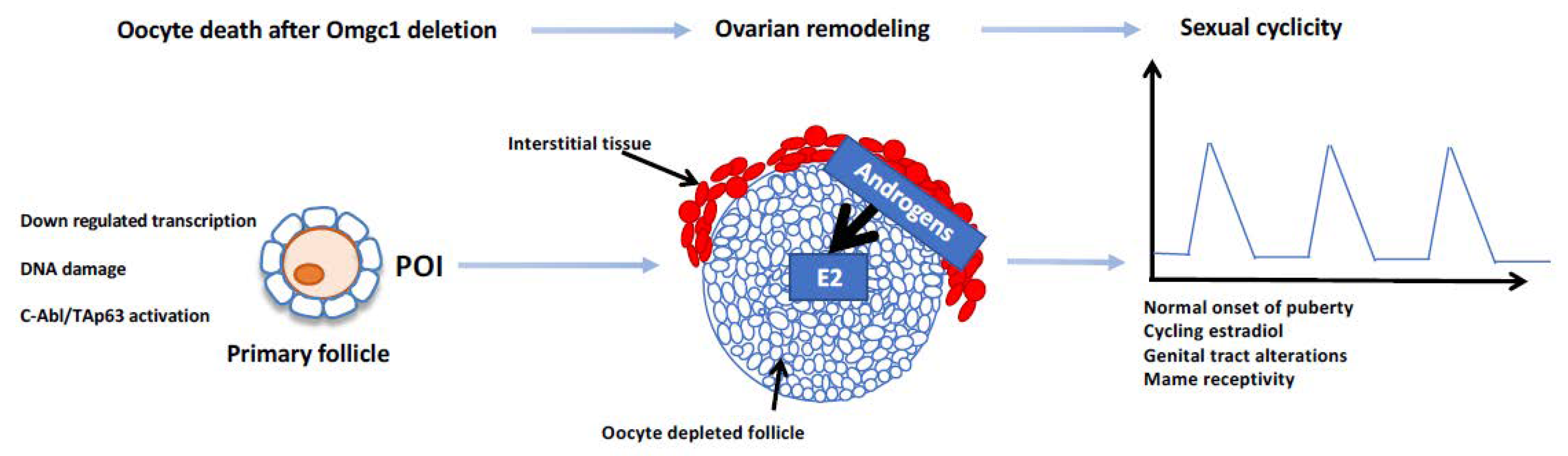
1.2. The Deletion of Omgc1 in Oocytes and Cyclic Ovarian Steroidogenesis in the Interstitium?
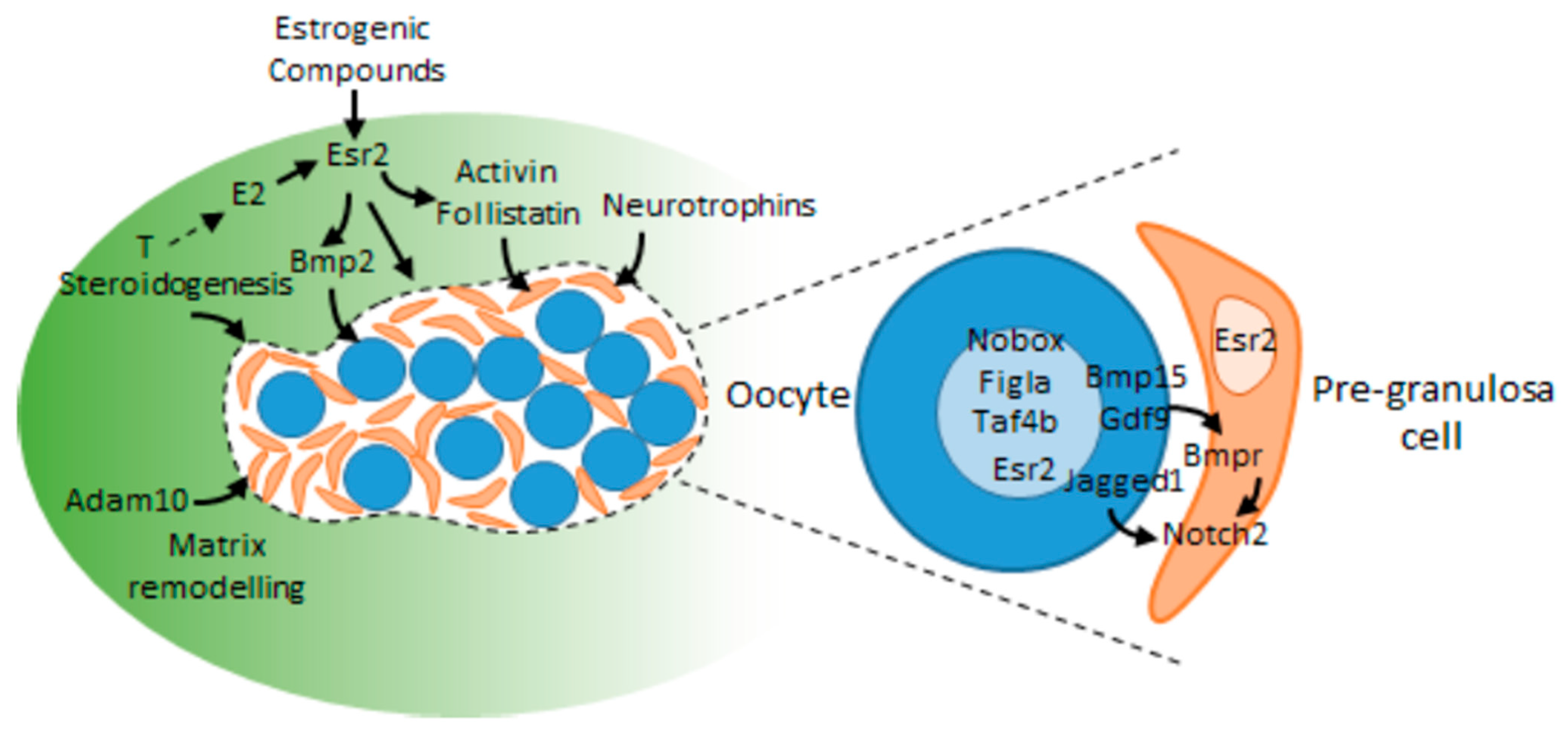
1.3. Depletion of Oocytes before Puberty and the Fate of Granulosa Cells
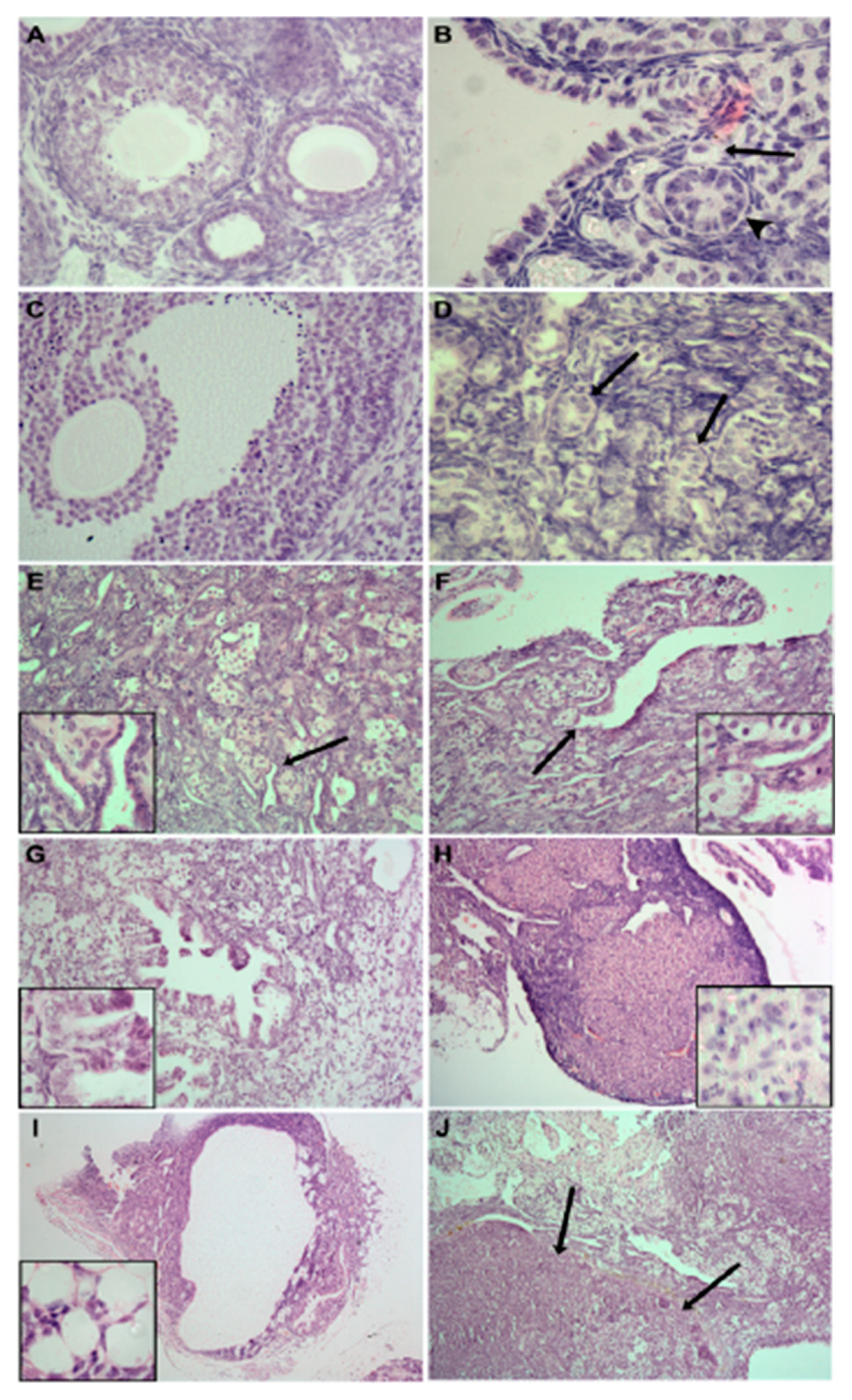
1.4. Poly-Ovular Follicles: From Physiology to Environmentally Induced Pathology
1.5. Differences in Ovarian Follicular Characteristics between Species
1.5.1. From Teleost to Tetrapod
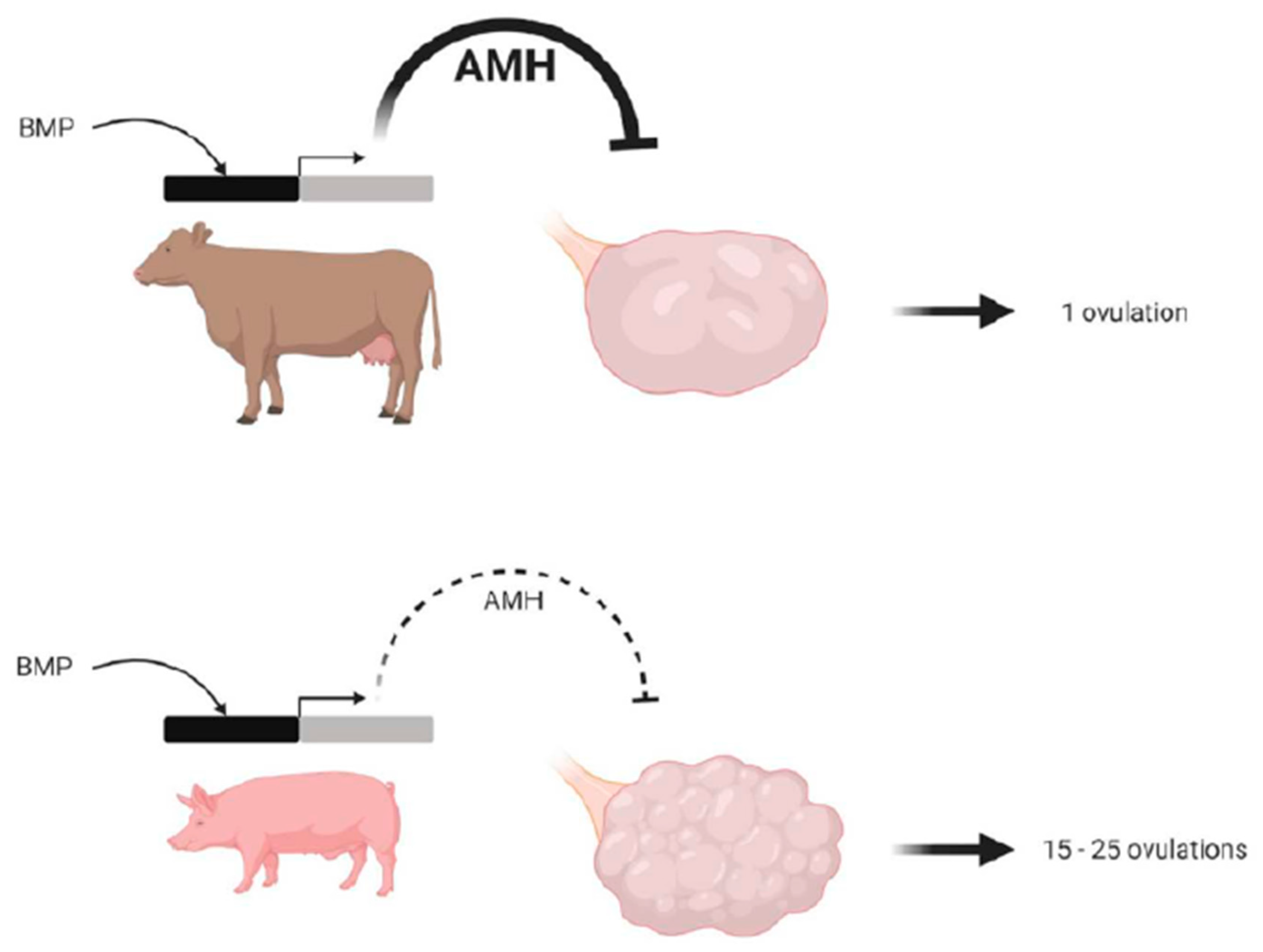
1.5.2. Between Pig and Cattle
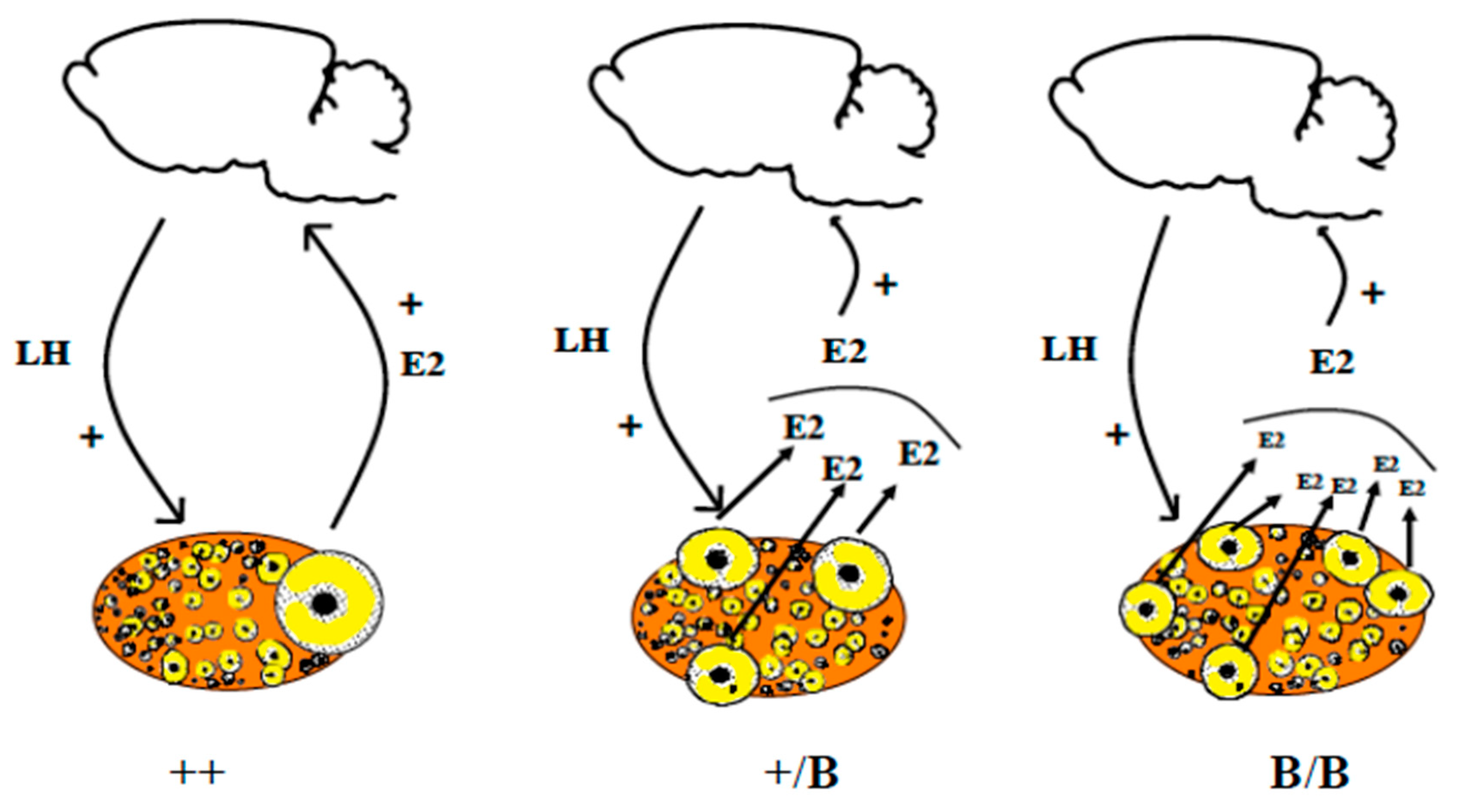
1.5.3. Between Different Sheep Breeds: The Role of the BMP Family Influencing Ovulation Rate
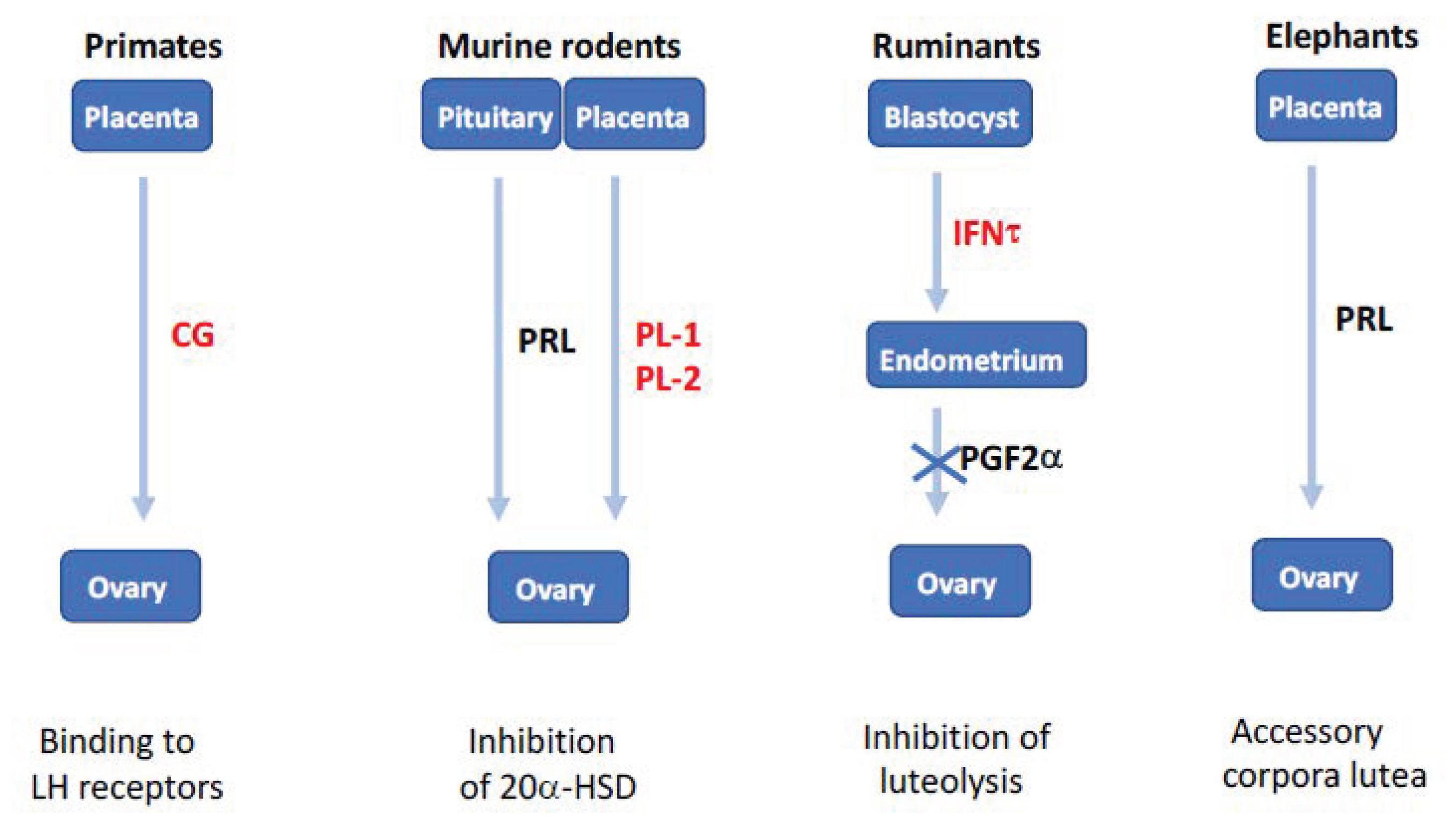
1.6. Why Are There So Many Luteotrophic Factors in Mammals?
- the human chorionic gonadotrophin (hCG) in women, which binds to the LH receptor.
- the trophoblastin (IFNτ) in ruminants, which impairs the secretion of the PGF2α luteolysis factor by the endometrium.
2. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sawyer, H.R.; Smith, P.; Heath, D.A.; Juengel, J.L.; Wakefield, S.J.; McNatty, K.P. Formation of ovarian follicles during fetal development in sheep. Biol. Reprod. 2002, 66, 1134–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, H.; Byskov, A.G.; Himelstein-Braw, R.; Faber, M. Follicular growth: The basic event in the mouse and human ovary. J. Reprod. Fertil. 1975, 45, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
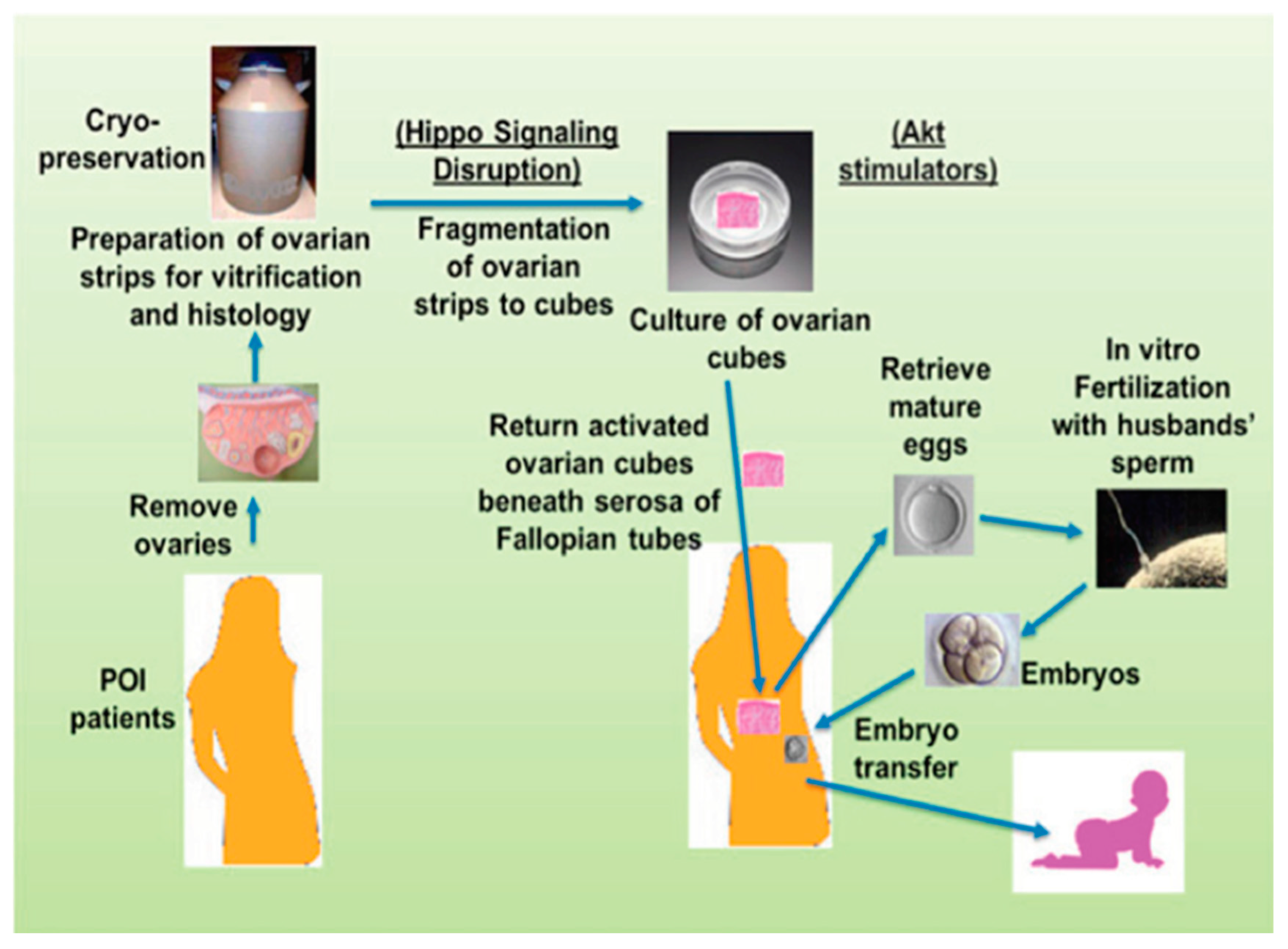
- Kawamura, K.; Cheng, Y.; Suzuki, N.; Deguchi, M.; Sato, Y.; Takae, S.; Ho, C.H.; Kawamura, N.; Tamura, M.; Hashimoto, S.; et al. Hippo signaling disruption and Akt stimulation of ovarian follicles for infertility treatment. Proc. Natl. Acad. Sci. USA 2013, 110, 17474–17479. [Google Scholar] [CrossRef] [Green Version]
- Vandormael-Pournin, S.; Guigon, C.J.; Ishaq, M.; Coudouel, N.; Ave, P.; Huerre, M.; Magre, S.; Cohen-Tannoudji, J.; Cohen-Tannoudji, M. Oocyte-specific inactivation of Omcg1 leads to DNA damage and c-Abl/TAp63-dependent oocyte death associated with dramatic remodeling of ovarian somatic cells. Cell Death Differ. 2015, 22, 108–117. [Google Scholar] [CrossRef] [Green Version]
- Mork, L.; Maatouk, D.M.; McMahon, J.A.; Guo, J.J.; Zhang, P.; McMahon, A.P.; Capel, B. Temporal differences in granulosa cell specification in the ovary reflect distinct follicle fates in mice. Biol. Reprod. 2012, 86, 37. [Google Scholar] [CrossRef] [PubMed]
- Himelstein-Braw, R.; Peters, H.; Faber, M. Influence of irradiation and chemotherapy on the ovaries of children with abdominal tumours. Br. J. Cancer 1977, 36, 269–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houghtaling, S.; Timmers, C.; Noll, M.; Finegold, M.J.; Jones, S.N.; Meyn, M.S.; Grompe, M. Epithelial cancer in Fanconi anemia complementation group D2 (Fancd2) knockout mice. Genes Dev. 2003, 17, 2021–2035. [Google Scholar] [CrossRef] [Green Version]
- Adriaens, I.; Smitz, J.; Jacquet, P. The current knowledge on radiosensitivity of ovarian follicle development stages. Hum. Reprod. Update 2009, 15, 359–377. [Google Scholar] [CrossRef] [Green Version]
- Pitman, J.L.; McNeilly, A.S.; McNeilly, J.R.; Hays, L.E.; Bagby, G.C., Jr.; Sawyer, H.R.; McNatty, K.P. The fate of granulosa cells following premature oocyte loss and the development of ovarian cancers. Int. J. Dev. Biol. 2012, 56, 949–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Albertini, D.F.; Nishimori, K.; Kumar, T.R.; Lu, N.; Matzuk, M.M. Growth differentiation factor-9 is required during early ovarian folliculogenesis. Nature 1996, 383, 531–535. [Google Scholar] [CrossRef]
- Braw-Tal, R.; McNatty, K.P.; Smith, P.; Heath, D.A.; Hudson, N.L.; Phillips, D.J.; McLeod, B.J.; Davis, G.H. Ovaries of ewes homozygous for the X-linked Inverdale gene (FecXI) are devoid of secondary and tertiary follicles but contain many abnormal structures. Biol. Reprod. 1993, 49, 895–907. [Google Scholar] [CrossRef] [Green Version]
- Hartman, C.G. Polynuclear ova and polyovular follicles in the opossum and other mammals, with special reference to the problem of fecundity. Am. J. Anat. 1926, 37, 1–51. [Google Scholar] [CrossRef]
- Telfer, E.; Gosden, R.G. A quantitative cytological study of polyovular follicles in mammalian ovaries with particular reference to the domestic bitch (Canis familiaris). J. Reprod. Fertil. 1987, 81, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Reynaud, K.; Halter, S.; Tahir, Z.; Thoumire, S.; Chebrout, M.; Chastant-Maillard, S. Polyovular follicles. Gynecol. Obstet. Fertil. 2010, 38, 395–397. [Google Scholar] [CrossRef]
- Gougeon, A. Frequent occurrence of multiovular follicles and multinuclear oocytes in the adult human ovary. Fertil. Steril. 1981, 35, 417–422. [Google Scholar] [CrossRef]
- Ben-Rafael, Z.; Mastroianni, L., Jr.; Kopf, G.S. In vitro fertilization and cleavage of a single egg from a binovular follicle containing two individual eggs surrounded by a single zona pellucida. Fertil. Steril. 1987, 47, 707–709. [Google Scholar] [CrossRef]
- Ron-El, R.; Nachum, H.; Golan, A.; Herman, A.; Yigal, S.; Caspi, E. Binovular human ovarian follicles associated with in vitro fertilization: Incidence and outcome. Fertil. Steril. 1990, 54, 869–872. [Google Scholar] [CrossRef]
- Coban, O.; Serdarogullari, M.; Pervaiz, R.; Soykok, A.; Bankeroglu, H. Fertilization and development of oocytes with separated and conjoined zona pellucida recovered from polyovular follicles: Description of two cases and a literature review. Zygote 2021, 20, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, L. Binovular follicles in the adult human ovary. Fertil. Steril. 1978, 29, 342–350. [Google Scholar] [CrossRef]
- Muretto, P.; Chilosi, M.; Rabitti, C.; Tommasoni, S.; Colato, C. Biovularity and “coalescence of primary follicles” in ovaries with mature teratomas. Int. J. Surg. Pathol. 2001, 9, 121–125. [Google Scholar] [CrossRef]
- Magdi, Y. Dizygotic twin from conjoined oocytes: A case report. J. Assist. Reprod. Genet. 2020, 37, 1367–1370. [Google Scholar] [CrossRef]
- Soyal, S.M.; Amleh, A.; Dean, J. FIGalpha, a germ cell-specific transcription factor required for ovarian follicle formation. Development 2000, 127, 4645–4654. [Google Scholar] [CrossRef] [PubMed]
- Lechowska, A.; Bilinski, S.; Choi, Y.; Shin, Y.; Kloc, M.; Rajkovic, A. Premature ovarian failure in nobox-deficient mice is caused by defects in somatic cell invasion and germ cell cyst breakdown. J. Assist. Reprod. Genet. 2011, 28, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Grive, K.J.; Seymour, K.A.; Mehta, R.; Freiman, R.N. TAF4b promotes mouse primordial follicle assembly and oocyte survival. Dev. Biol. 2014, 392, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.; Wang, P.; DeMayo, J.; DeMayo, F.J.; Elvin, J.A.; Carino, C.; Prasad, S.V.; Skinner, S.S.; Dunbar, B.S.; Dube, J.L.; et al. Synergistic roles of bone morphogenetic protein 15 and growth differentiation factor 9 in ovarian function. Mol. Endocrinol. 2001, 15, 854–866. [Google Scholar] [CrossRef]
- Bristol-Gould, S.K.; Kreeger, P.K.; Selkirk, C.G.; Kilen, S.M.; Cook, R.W.; Kipp, J.L.; Shea, L.D.; Mayo, K.E.; Woodruff, T.K. Postnatal regulation of germ cells by activin: The establishment of the initial follicle pool. Dev. Biol. 2006, 298, 132–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, B.; Garcia-Rudaz, C.; Dorfman, M.; Paredes, A.; Ojeda, S.R. NTRK1 and NTRK2 receptors facilitate follicle assembly and early follicular development in the mouse ovary. Reproduction 2009, 138, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Kimura, F.; Bonomi, L.M.; Schneyer, A.L. Follistatin regulates germ cell nest breakdown and primordial follicle formation. Endocrinology 2011, 152, 697–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, L.; Wang, Y.; Cai, H.; Sun, G.; Niu, W.; Xin, Q.; Tang, X.; Zhang, J.; Wang, C.; Zhang, H.; et al. ADAM10-Notch signaling governs the recruitment of ovarian pregranulosa cells and controls folliculogenesis in mice. J. Cell Sci. 2016, 129, 2202–2212. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Gridley, T. Notch2 is required in somatic cells for breakdown of ovarian germ-cell nests and formation of primordial follicles. BMC Biol. 2013, 11, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Du, X.; Huang, K.; Zhang, T.; Teng, Z.; Niu, W.; Wang, C.; Xia, G. Rac1 modulates the formation of primordial follicles by facilitating STAT3-directed Jagged1, GDF9 and BMP15 transcription in mice. Sci. Rep. 2016, 6, 23972. [Google Scholar] [CrossRef] [Green Version]
- Monniaux, D. Factors influencing establishment of the ovarian reserve and their effects on fertility. Anim. Reprod. 2018, 15, 635–647. [Google Scholar] [CrossRef]
- Billiar, R.B.; Zachos, N.C.; Burch, M.G.; Albrecht, E.D.; Pepe, G.J. Up-regulation of α-inhibin expression in the fetal ovary of estrogen-suppressed baboons is associated with impaired fetal ovarian folliculogenesis. Biol. Reprod. 2003, 68, 1989–1996. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Roy, S.K. Stimulation of primordial follicle assembly by estradiol-17beta requires the action of bone morphogenetic protein-2 (BMP2). Sci. Rep. 2017, 7, 15581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillette, L.J., Jr.; Gross, T.S.; Masson, G.R.; Matter, J.M.; Percival, H.F.; Woodward, A.R. Developmental abnormalities of the gonad and abnormal sex hormone concentrations in juvenile alligators from contaminated and control lakes in Florida. Environ. Health Perspect. 1994, 102, 680–688. [Google Scholar] [CrossRef]
- Jefferson, W.; Newbold, R.; Padilla-Banks, E.; Pepling, M. Neonatal genistein treatment alters ovarian differentiation in the mouse: Inhibition of oocyte nest breakdown and increased oocyte survival. Biol. Reprod. 2006, 74, 161–168. [Google Scholar] [CrossRef]
- Karavan, J.R.; Pepling, M.E. Effects of estrogenic compounds on neonatal oocyte development. Reprod. Toxicol. 2012, 34, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Li, L.; Qin, X.S.; Zhou, Y.; Zhang, X.F.; Wang, L.Q.; De Felici, M.; Chen, H.; Qin, G.Q.; Shen, W. Di-(2-ethylhexyl) phthalate and bisphenol A exposure impairs mouse primordial follicle assembly in vitro. Environ. Mol. Mutagen. 2014, 55, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Liao, X.; Chen, X.; Li, Y.; Wang, M.; Shen, C.; Zhang, X.; Wang, Y.; Liu, X.; He, J. DEHP exposure impairs mouse oocyte cyst breakdown and primordial follicle assembly through estrogen receptor-dependent and independent mechanisms. J. Hazard. Mater. 2015, 298, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Meslin, C.; Mugnier, S.; Callebaut, I.; Laurin, M.; Pascal, G.; Poupon, A.; Goudet, G.; Monget, P. Evolution of genes involved in gamete interaction: Evidence for positive selection, duplications and losses in vertebrates. PLoS ONE 2012, 7, e44548. [Google Scholar] [CrossRef]
- Goudet, G.; Mugnier, S.; Callebaut, I.; Monget, P. Phylogenetic analysis and identification of pseudogenes reveal a progressive loss of zona pellucida genes during evolution of vertebrates. Biol. Reprod. 2008, 78, 796–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josso, N.; Racine, C.; di Clemente, N.; Rey, R.; Xavier, F. The role of anti-Mullerian hormone in gonadal development. Mol. Cell. Endocrinol. 1998, 145, 3–7. [Google Scholar] [CrossRef]
- Visser, J.A.; Themmen, A.P. Anti-Mullerian hormone and folliculogenesis. Mol. Cell. Endocrinol. 2005, 234, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Visser, J.A.; Themmen, A.P. Role of anti-Mullerian hormone and bone morphogenetic proteins in the regulation of FSH sensitivity. Mol. Cell. Endocrinol. 2014, 382, 460–465. [Google Scholar] [CrossRef]
- Estienne, A.; Jarrier, P.; Staub, C.; Venturi, E.; Le Vern, Y.; Clemente, N.; Monniaux, D.; Monget, P. Anti-Mullerian hormone production in the ovary: A comparative study in bovine and porcine granulosa cellsdagger. Biol. Reprod. 2020, 103, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Juengel, J.L.; McNatty, K.P. The role of proteins of the transforming growth factor-β superfamily in the intraovarian regulation of follicular development. Hum. Reprod. Update 2005, 11, 143–160. [Google Scholar] [CrossRef]
- Fabre, S.; Pierre, A.; Mulsant, P.; Bodin, L.; Di Pasquale, E.; Persani, L.; Monget, P.; Monniaux, D. Regulation of ovulation rate in mammals: Contribution of sheep genetic models. Reprod. Biol. Endocr. 2006, 4, 20. [Google Scholar] [CrossRef] [Green Version]
- Juengel, J.L.; Davis, G.H.; McNatty, K.P. Using sheep lines with mutations in single genes to better understand ovarian function. Reproduction 2013, 146, R111–R123. [Google Scholar] [CrossRef]
- Davis, G.H. Fecundity genes in sheep. Anim. Reprod. Sci. 2004, 82–83, 247–253. [Google Scholar] [CrossRef]
- Davis, G.H.; Galloway, S.M.; O’Connell, A.; Farquhar, P.A.; McNatty, K.P.; Juengel, J.L. Hyper-prolific ewes carrying copies of three major genes: A model for studying genes controlling ovulation rate. Biol. Reprod. 2008, 78, 110. [Google Scholar] [CrossRef]
- Christoforou, E.R.; Pitman, J.L. Intrafollicular growth differentiation factor 9: Bone morphogenetic 15 ratio determines litter size in mammalsdagger. Biol. Reprod. 2019, 100, 1333–1343. [Google Scholar] [CrossRef] [PubMed]
- McNatty, K.P.; Lun, S.; Heath, D.A.; Ball, K.; Smith, P.; Hudson, N.L.; McDiarmid, J.; Gibb, M.; Henderson, K.M. Differences in ovarian activity between booroola X merino ewes which were homozygous, heterozygous and non-carriers of a major gene influencing their ovulation rate. J. Reprod. Fertil. 1986, 77, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Shackell, G.H.; Hudson, N.L.; Heath, D.A.; Lun, S.; Shaw, L.; Condell, L.; Blay, L.R.; McNatty, K.P. Plasma gonadotropin concentrations and ovarian characteristics in Inverdale ewes that are heterozygous for a major gene (FecX1) on the X chromosome that influences ovulation rate. Biol. Reprod. 1993, 48, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- McNatty, K.P.; Heath, D.A.; Clark, Z.; Reader, K.; Juengel, J.L.; Pitman, J.L. Ovarian characteristics in sheep with multiple fecundity genes. Reproduction 2017, 153, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Niswender, G.D.; McNatty, K.P.; Smith, P.; Niswender, K.D.; Farin, C.E.; Sawyer, H.R. Numbers of steroidogenic luteal cells in Booroola Merino ewes. J. Reprod. Fertil. 1990, 90, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Juengel, J.L.; Hudson, N.L.; Berg, M.; Hamel, K.; Smith, P.; Lawrence, S.B.; Whiting, L.; McNatty, K.P. Effects of active immunization against growth differentiation factor 9 and/or bone morphogenetic protein 15 on ovarian function in cattle. Reproduction 2009, 138, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.S.; Zhao, Z.Z.; Hoekstra, C.; Hayward, N.K.; Webb, P.M.; Whiteman, D.C.; Martin, N.G.; Boomsma, D.I.; Duffy, D.L.; Montgomery, G.W. Novel variants in growth differentiation factor 9 in mothers of dizygotic twins. J. Clin. Endocr. Metab. 2006, 91, 4713–4716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, A.M. Evolution of placental function in mammals: The molecular basis of gas and nutrient transfer, hormone secretion, and immune responses. Physiol. Rev. 2012, 92, 1543–1576. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monget, P.; McNatty, K.; Monniaux, D. The Crazy Ovary. Genes 2021, 12, 928. https://doi.org/10.3390/genes12060928
Monget P, McNatty K, Monniaux D. The Crazy Ovary. Genes. 2021; 12(6):928. https://doi.org/10.3390/genes12060928
Chicago/Turabian StyleMonget, Philippe, Ken McNatty, and Danielle Monniaux. 2021. "The Crazy Ovary" Genes 12, no. 6: 928. https://doi.org/10.3390/genes12060928
APA StyleMonget, P., McNatty, K., & Monniaux, D. (2021). The Crazy Ovary. Genes, 12(6), 928. https://doi.org/10.3390/genes12060928