
Phylogeny and Evolutionary History of Respiratory Complex I Proteins in Melainabacteria

 ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
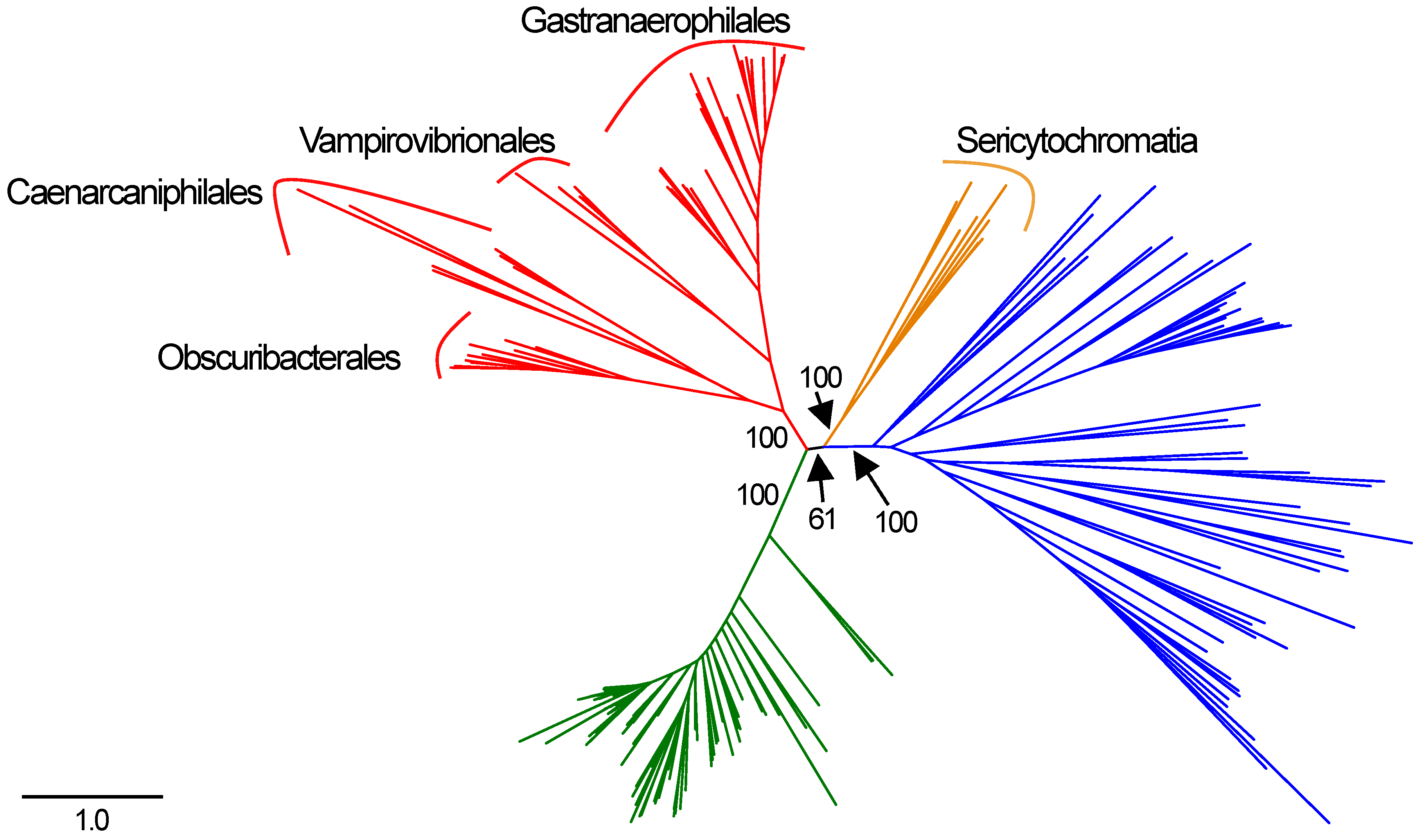
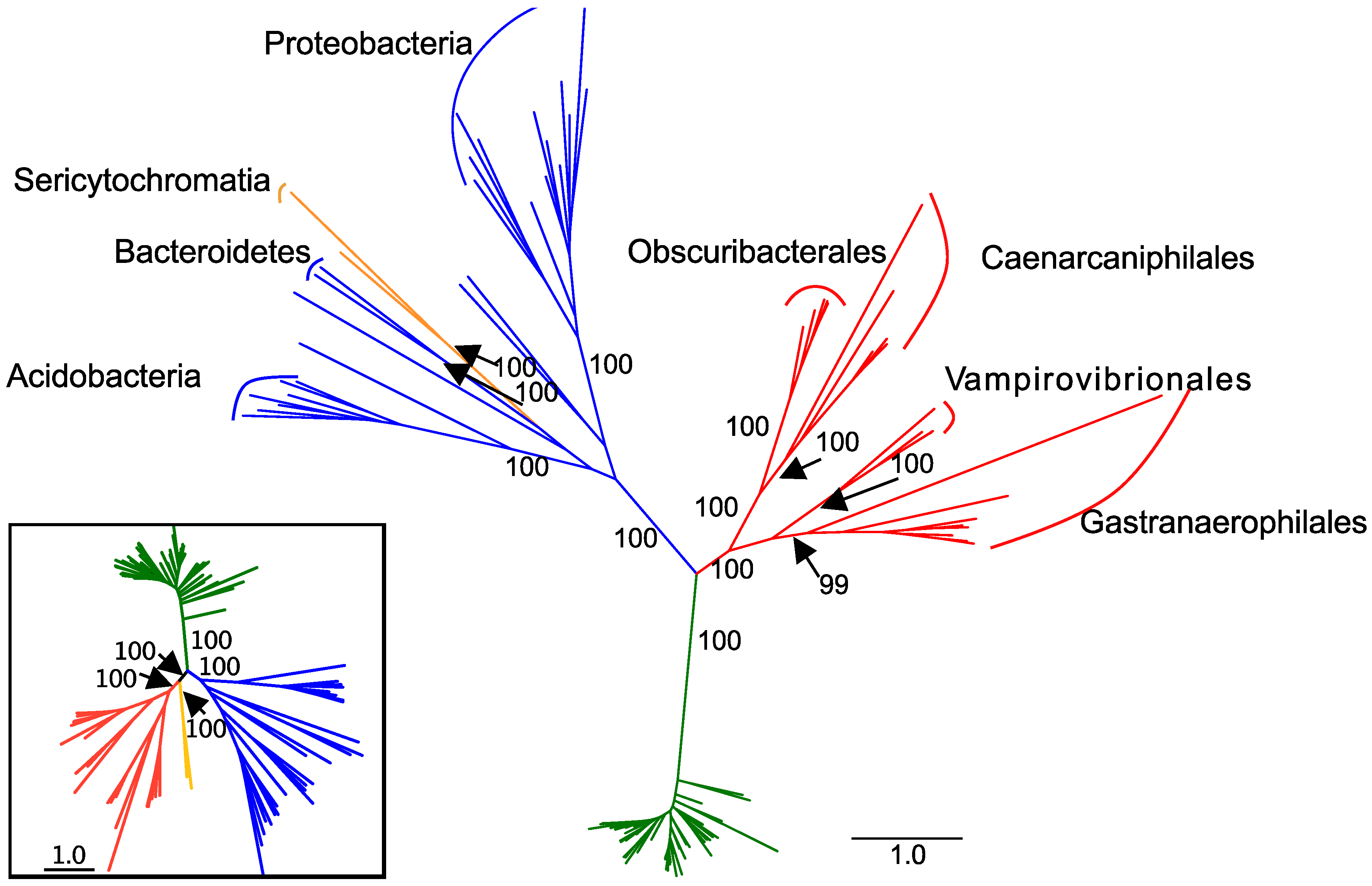
3. Results
Phylogeny of Complex I
4. Discussion
Possible Evolutionary History of Complex I
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shih, P.M.; Hemp, J.; Ward, L.M.; Matzke, N.J.; Fischer, W.W. Crown Group Oxyphotobacteria Postdate the Rise of Oxygen. Geobiology 2017, 15, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Magnabosco, C.; Moore, K.R.; Wolfe, J.M.; Fournier, G.P. Dating Phototrophic Microbial Lineages with Reticulate Gene Histories. Geobiology 2018, 16, 179–189. [Google Scholar] [CrossRef] [Green Version]
- Cardona, T.; Sánchez-Baracaldo, P.; Rutherford, A.W.; Larkum, A.W. Early Archean Origin of Photosystem II. Geobiology 2018, 17, 127–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blankenship, R.E. Early Evolution of Photosynthesis. Plant Physiol. 2010, 154, 434–438. [Google Scholar] [CrossRef] [Green Version]
- Soo, R.; Skennerton, C.; Sekiguchi, Y.; Imelfort, M.; Paech, S.; Dennis, P.; Steen, J.; Parks, D.; Tyson, G.; Hugenholtz, P. An Expanded Genomic Representation of the Phylum Cyanobacteria. Genome Biol. Evol. 2014, 6, 1031–1045. [Google Scholar] [CrossRef] [Green Version]
- Planavsky, N.J.; Asael, D.; Hofmann, A.; Reinhard, C.T.; Lalonde, S.V.; Knudsen, A.; Wang, X.; Ossa, F.O.; Pecoits, E.; Smith, A.J.B.; et al. Evidence for Oxygenic Photosynthesis Half a Billion Years before the Great Oxidation Event. Nat. Geosci. 2014, 7, 283–286. [Google Scholar] [CrossRef] [Green Version]
- Catling, D.C.; Glein, C.R.; Zahnle, K.J.; McKay, C.P. Why O 2 Is Required by Complex Life on Habitable Planets and the Concept of Planetary “Oxygenation Time”. Astrobiology 2005, 5, 415–438. [Google Scholar] [CrossRef] [Green Version]
- Soo, R.; Hemp, J.; Parks, D.; Fischer, W.; Hugenholtz, P. On the Origins of Oxygenic Photosynthesis and Aerobic Respiration in Cyanobacteria. Science 2017, 355, 1436–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soo, R.M.; Hemp, J.; Hugenholtz, P. The Evolution of Photosynthesis and Aerobic Respiration in the Cyanobacteria. Free Radic. Bio. Med. 2019. [Google Scholar] [CrossRef]
- Brochier-Armanet, C.; Talla, E.; Gribaldo, S. The Multiple Evolutionary Histories of Dioxygen Reductases: Implications for the Origin and Evolution of Aerobic Respiration. Mol. Biol. Evol. 2008, 26, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Strous, M. Denitrification and Aerobic Respiration, Hybrid Electron Transport Chains and Co-Evolution. Biochim. Biophys. Acta BBA Bioenerg. 2013, 1827, 136–144. [Google Scholar] [CrossRef] [Green Version]
- Saraste, M.; Castresana, J. Cytochrome Oxidase Evolved by Tinkering with Denitrification Enzymes. FEBS Lett. 1994, 341, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Ducluzeau, A.-L.; van Lis, R.; Duval, S.; Schoepp-Cothenet, B.; Russell, M.J.; Nitschke, W. Was Nitric Oxide the First Deep Electron Sink? Trends Biochem. Sci. 2009, 34, 9–15. [Google Scholar] [CrossRef]
- Garcia-Pichel, F.; Zehr, J.P.; Bhattacharya, D.; Pakrasi, H.B. What’s in a Name? The Case of Cyanobacteria. J. Phycol. 2020, 56, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Nayfach, S.; Roux, S.; Seshadri, R.; Udwary, D.; Varghese, N.; Schulz, F.; Wu, D.; Paez-Espino, D.; Chen, I.-M.; Huntemann, M.; et al. A Genomic Catalog of Earth’s Microbiomes. Nat. Biotechnol. 2021, 39, 499–509. [Google Scholar] [CrossRef]
- Chen, I.-M.A.; Chu, K.; Palaniappan, K.; Ratner, A.; Huang, J.; Huntemann, M.; Hajek, P.; Ritter, S.; Varghese, N.; Seshadri, R.; et al. The IMG/M Data Management and Analysis System v.6.0: New Tools and Advanced Capabilities. Nucleic Acids Res. 2020, 49, gkaa939. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the Quality of Microbial Genomes Recovered from Isolates, Single Cells, and Metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. EggNOG 5.0: A Hierarchical, Functionally and Phylogenetically Annotated Orthology Resource Based on 5090 Organisms and 2502 Viruses. Nucleic Acids Res. 2018, 47, D309–D314. [Google Scholar] [CrossRef] [Green Version]
- Eren, A.M.; Esen, Ö.C.; Quince, C.; Vineis, J.H.; Morrison, H.G.; Sogin, M.L.; Delmont, T.O. Anvi’o: An Advanced Analysis and Visualization Platform for ‘omics Data. PeerJ 2015, 3, e1319. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. TrimAl: A Tool for Automated Alignment Trimming in Large-Scale Phylogenetic Analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A New and Scalable Tool for the Selection of DNA and Protein Evolutionary Models. Mol. Biol. Evol. 2019, 37, 291–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v3: An Online Tool for the Display and Annotation of Phylogenetic and Other Trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Battchikova, N.; Eisenhut, M.; Aro, E.-M. Cyanobacterial NDH-1 Complexes: Novel Insights and Remaining Puzzles. Biochim. Biophys. Acta BBA Bioenerg. 2011, 1807, 935–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathiesen, C.; Hägerhäll, C. The ‘Antiporter Module’ of Respiratory Chain Complex I Includes the MrpC/NuoK Subunit—A Revision of the Modular Evolution Scheme. FEBS Lett. 2003, 549, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Grettenberger, C.L.; Sumner, D.Y.; Eisen, J.A.; Jungblut, A.D.; Mackey, T.J. Supplemental Data for Phylogeny of Complex I in Melainabacteria. Available online: https://osf.io/b72ym/?view_only=ca528ff7c74e42babfefa82ab25b9b50.
- Eren, M. A Tutorial on Assembly-Based Metagenomics. Available online: http://merenlab.org/tutorials/assembly-based-metagenomics/ (accessed on 1 June 2020).
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum Likelihood-Based Phylogenetic Analyses with Thousands of Taxa and Mixed Models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. Gatew. Comput. Environ. Work GCE 2010, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Cornet, L.; Meunier, L.; Vlierberghe, M.V.; Léonard, R.R.; Durieu, B.; Lara, Y.; Misztak, A.; Sirjacobs, D.; Javaux, E.J.; Philippe, H.; et al. Consensus Assessment of the Contamination Level of Publicly Available Cyanobacterial Genomes. PLoS ONE 2018, 13, e0200323. [Google Scholar] [CrossRef] [Green Version]
- Shibata, M.; Ohkawa, H.; Kaneko, T.; Fukuzawa, H.; Tabata, S.; Kaplan, A.; Ogawa, T. Distinct Constitutive and Low-CO2-Induced CO2 Uptake Systems in Cyanobacteria: Genes Involved and Their Phylogenetic Relationship with Homologous Genes in Other Organisms. Proc. Natl. Acad. Sci. USA 2001, 98, 11789–11794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, T.; Böttcher, B. The Gross Structure of the Respiratory Complex I: A Lego System. Biochim. Biophys. Acta BBA Bioenerg. 2004, 1608, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grettenberger, C.; Sumner, D.Y.; Eisen, J.A.; Jungblut, A.D.; Mackey, T.J. Phylogeny and Evolutionary History of Respiratory Complex I Proteins in Melainabacteria. Genes 2021, 12, 929. https://doi.org/10.3390/genes12060929
Grettenberger C, Sumner DY, Eisen JA, Jungblut AD, Mackey TJ. Phylogeny and Evolutionary History of Respiratory Complex I Proteins in Melainabacteria. Genes. 2021; 12(6):929. https://doi.org/10.3390/genes12060929
Chicago/Turabian StyleGrettenberger, Christen, Dawn Y. Sumner, Jonathan A. Eisen, Anne D. Jungblut, and Tyler J. Mackey. 2021. "Phylogeny and Evolutionary History of Respiratory Complex I Proteins in Melainabacteria" Genes 12, no. 6: 929. https://doi.org/10.3390/genes12060929
APA StyleGrettenberger, C., Sumner, D. Y., Eisen, J. A., Jungblut, A. D., & Mackey, T. J. (2021). Phylogeny and Evolutionary History of Respiratory Complex I Proteins in Melainabacteria. Genes, 12(6), 929. https://doi.org/10.3390/genes12060929