
Behind the Scene: Exploiting MC1R in Skin Cancer Risk and Prevention

 , ,
, ,  and
and 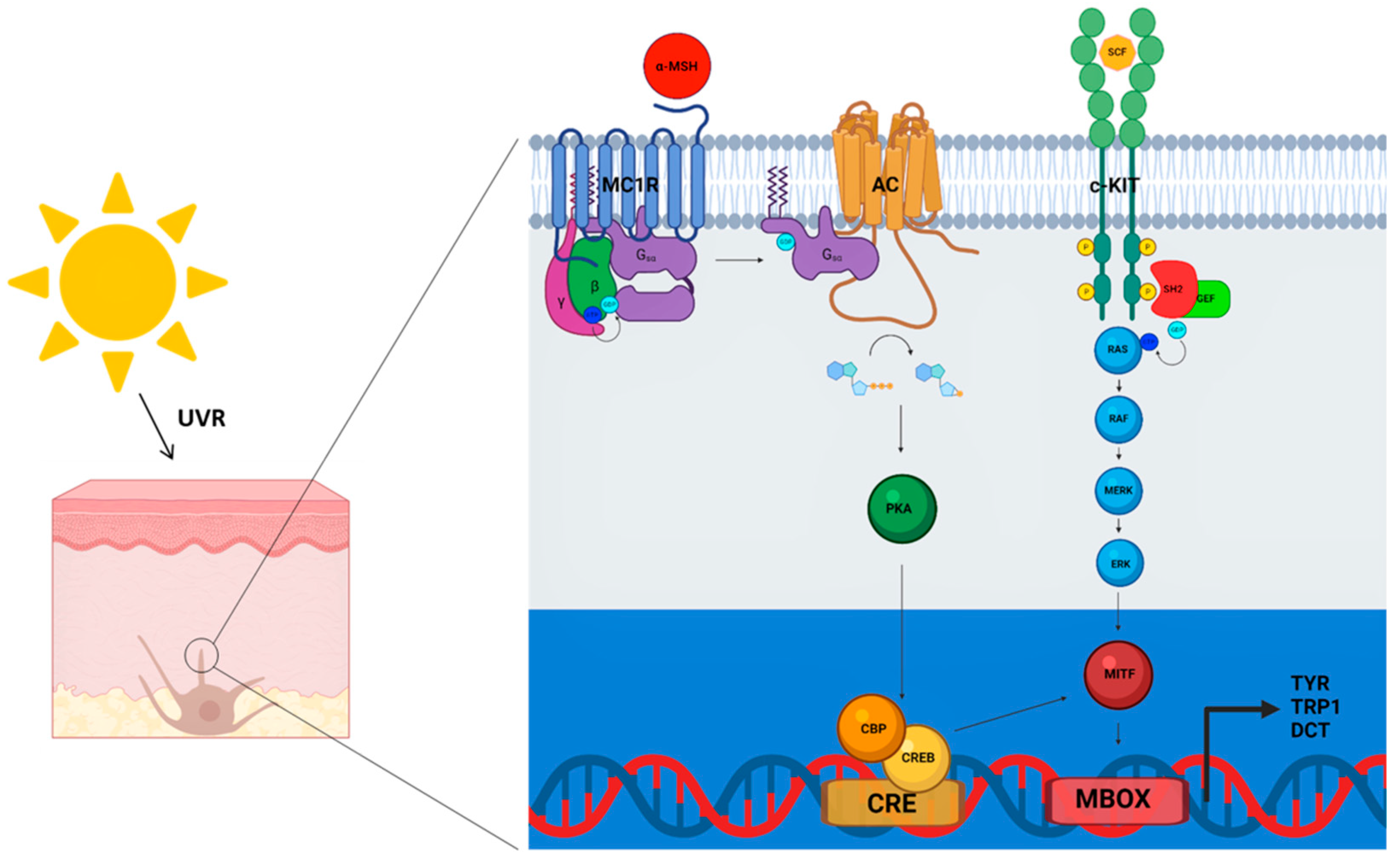{kind=link}
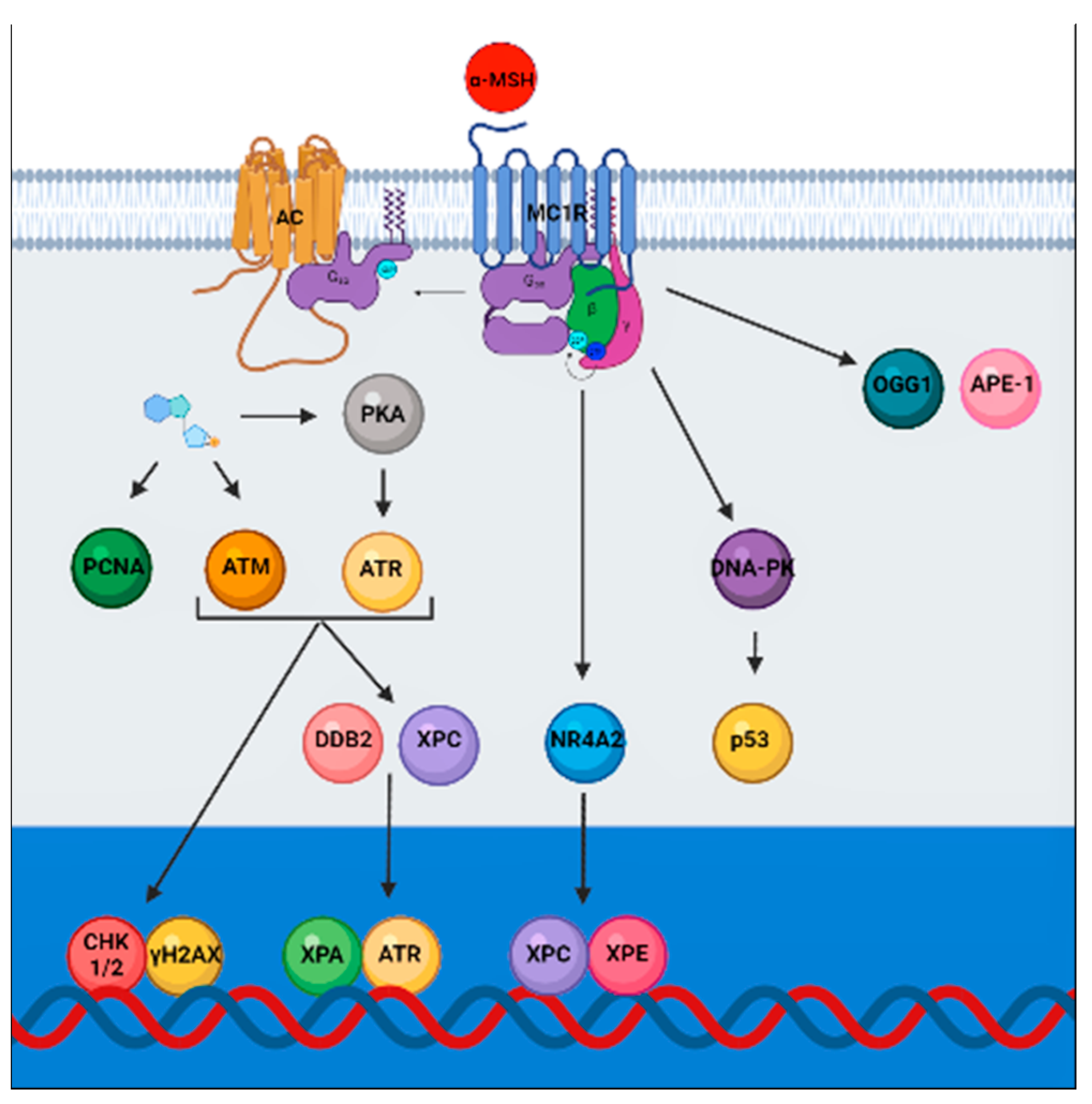{kind=link}
Abstract
:1. Introduction
2. MC1R Structure, Regulation and Functions
2.1. MC1R Structure and Regulation
2.2. MC1R and Pigmentation
2.3. MC1R, Non-Pigmentary Functions and DNA Repair
3. MC1R Polymorphisms
4. Clinical Impact of MC1R Polymorphisms: Hair and Skin Color and Non-Invasive Imaging Features
5. MC1R and Skin Cancers Risk
5.1. MC1R and Melanoma Risk
5.2. MC1R and NMSCs Risk
6. MC1R Association with Melanoma Susceptible Genes
6.1. CDKN2A
6.2. BRAF
6.3. Other Genes
7. MC1R Association with NMSCs Susceptible Genes
8. Epigenetic Regulation of MC1R
9. MC1R and the Impact of Skin Cancer Genetic Testing
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- American Cancer Society. Cancer Facts Figures 2021; American Cancer Society: Atlanta, GA, USA, 2021. [Google Scholar]
- AIOM. I Numeri del Cancro in Italia; AIOM: Milan, Italy, 2020. [Google Scholar]
- Zanna, P.; Maida, I.; Grieco, C.; Guida, S.; Turpin Sevilla, M.C.; De Summa, S.; Tommasi, S.; Vena, G.A.; Filotico, R.; Guida, G. Three novel human sporadic melanoma cell lines: Signaling pathways controlled by MC1R, BRAF and β-catenins. J. Biol. Regul. Homeost. Agents 2013, 27, 131–141. [Google Scholar] [PubMed]
- Guida, S.; De Pace, B.; Ciardo, S.; Farnetani, F.; Pellacani, G. Non-invasive Imaging for Skin Cancers—The European Experience. Curr. Dermatol. Rep. 2019, 8, 172–181. [Google Scholar] [CrossRef]
- Roberts, M.R.; Asgari, M.M.; Toland, A.E. Genome-wide association studies and polygenic risk scores for skin cancer: Clinically useful yet? Br. J. Dermatol. 2019, 181, 1146–1155. [Google Scholar] [CrossRef]
- Caini, S.; Gandini, S.; Botta, F.; Tagliabue, E.; Raimondi, S.; Nagore, E.; Zanna, I.; Maisonneuve, P.; Newton-Bishop, J.; Polsky, D.; et al. MC1R variants and cutaneous melanoma risk according to histological type, body site, and Breslow thickness: A pooled analysis from the M-SKIP project. Melanoma Res. 2020, 30, 500–510. [Google Scholar] [CrossRef]
- Geschwind, I.I. Change in hair color in mice induced by injection of α-MSH. Endocrinology 1966, 79, 1165–1167. [Google Scholar] [CrossRef]
- Sawyer, T.K.; Hruby, V.J.; Wilkes, B.C.; Draelos, M.T.; Hadley, M.E.; Bergsneider, M. Comparative Biological Activities of Highly Potent Active-Site Analogues of α-Melanotropin. J. Med. Chem. 1982, 25, 1022–1027. [Google Scholar] [CrossRef] [PubMed]
- García-Borrón, J.C.; Sánchez-Laorden, B.L.; Jiménez-Cervantes, C. Melanocortin-1 receptor structure and functional regulation. Pigment Cell Res. 2005, 18, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Wolf Horrell, E.M.; Boulanger, M.C.; D’Orazio, J.A. Melanocortin 1 receptor: Structure, function, and regulation. Front. Genet. 2016, 7, 95. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhu, B.; Yin, C.; Liu, W.; Han, C.; Chen, B.; Liu, T.; Li, X.; Chen, X.; Li, C.; et al. Palmitoylation-dependent activation of MC1R prevents melanomagenesis. Nature 2017, 549, 399–403. [Google Scholar] [CrossRef] [Green Version]
- Guida, S.; Guida, G.; Goding, C.R. MC1R functions, expression and implications for targeted therapy. J. Investig. Dermatol. 2021, in press. [Google Scholar]
- Dalziel, M.; Kolesnichenko, M.; Das Neves, R.P.; Iborra, F.; Goding, C.; Furger, A. α-MSH regulates intergenic splicing of MC1R and TUBB3 in human melanocytes. Nucleic Acids Res. 2011, 39, 2378–2392. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Vicente, I.; Abrisqueta, M.; Herraiz, C.; Jiménez-Cervantes, C.; García-Borrón, J.C.; Olivares, C. Functional characterization of a C-terminal splice variant of the human melanocortin 1 receptor. Exp. Dermatol. 2020, 29, 610–615. [Google Scholar] [CrossRef]
- Herraiz, C.; Olivares, C.; Castejón-Griñán, M.; Abrisqueta, M.; Jiménez-Cervantes, C.; García-Borrón, J.C. Functional characterization of MC1R-TUBB3 intergenic splice variants of the human melanocortin 1 receptor. PLoS ONE 2015, 10, e0144757. [Google Scholar] [CrossRef]
- Herraiz, C.; Sánchez-Laorden, B.L.; Jiménez-Cervantes, C.; García-Borrón, J.C. N-glycosylation of the human melanocortin 1 receptor: Occupancy of glycosylation sequons and functional role. Pigment Cell Melanoma Res. 2011, 24, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Laorden, B.L.; Jiménez-Cervantes, C.; García-Borrón, J.C. Regulation of human melanocortin 1 receptor signaling and trafficking by Thr-308 and Ser-316 and its alteration in variant alleles associated with red hair and skin cancer. J. Biol. Chem. 2007, 282, 3241–3251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Laorden, B.L.; Sánchez-Más, J.; Martínez-Alonso, E.; Martínez-Menárguez, J.A.; García-Borrón, J.C.; Jiménez-Cervantes, C. Dimerization of the human melanocortin 1 receptor: Functional consequences and dominant-negative effects. J. Investig. Dermatol. 2006, 126, 172–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Han, C.; Miao, X.; Li, X.; Yin, C.; Zou, J.; Liu, M.; Li, S.; Stawski, L.; Zhu, B.; et al. Targeting MC1R depalmitoylation to prevent melanomagenesis in redheads. Nat. Commun. 2019, 10, 877. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Más, J.; Hahmann, C.; Gerritsen, I.; Garcia-Borrón, J.C.; Jiménez-Cervantes, C. Agonist-independent, high constitutive activity of the human melanocortin 1 receptor. Pigment Cell Res. 2004, 17, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Rouzaud, F.; Annereau, J.P.; Valencia, J.C.; Costin, G.E.; Hearing, V.J. Regulation of melanocortin 1 receptor expression at the mRNA and protein levels by its natural agonist and antagonist. FASEB J. 2003, 17, 1–21. [Google Scholar] [CrossRef]
- Swope, V.B.; Abdel-Malek, Z.A. Significance of the melanocortin 1 and endothelin B receptors in melanocyte homeostasis and prevention of sun-induced genotoxicity. Front. Genet. 2016, 7, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funasaka, Y.; Chakraborty, A.K.; Hayashi, Y.; Komoto, M.; Ohashi, A.; Nagahama, M.; Inoue, Y.; Pawelek, J.; Ichihashi, M. Modulation of melanocyte-stimulating hormone receptor expression on normal human melanocytes: Evidence for a regulatory role of ultraviolet B, interleukin-1α, interleukin-1β, endothelin-1 and tumour necrosis factor-α. Br. J. Dermatol. 1998, 139, 216–224. [Google Scholar] [CrossRef]
- Scott, M.C.; Suzuki, I.; Abdel-Malek, Z.A. Regulation of the human melanocortin 1 receptor expression in epidermal melanocytes by paracrine and endocrine factors and by ultraviolet radiation. Pigment Cell Res. 2002, 15, 433–439. [Google Scholar] [CrossRef]
- Aberdam, E.; Bertolotto, C.; Sviderskaya, E.V.; De Thillott, V.; Hemesath, T.J.; Fisher, D.E.; Bennett, D.C.; Ortonne, J.P.; Ballottit, R. Involvement of microphthalmia in the inhibition of melanocyte lineage differentiation and of melanogenesis by agouti signal protein. J. Biol. Chem. 1998, 273, 19560–19565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nix, M.A.; Kaelin, C.B.; Ta, T.; Weis, A.; Morton, G.J.; Barsh, G.S.; Millhauser, G.L. Molecular and functional analysis of human β-defensin 3 action at melanocortin receptors. Chem. Biol. 2013, 20, 784–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swope, V.B.; Jameson, J.A.; McFarland, K.L.; Supp, D.M.; Miller, W.E.; McGraw, D.W.; Patel, M.A.; Nix, M.A.; Millhauser, G.L.; Babcock, G.F.; et al. Defining MC1R regulation in human melanocytes by its agonist α-melanocortin and antagonists agouti signaling protein and Β-defensin 3. J. Investig. Dermatol. 2012, 132, 2255–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, M.C.; Wakamatsu, K.; Ito, S.; Kadekaro, A.L.; Kobayashi, N.; Groden, J.; Kavanagh, R.; Takakuwa, T.; Virador, V.; Hearing, V.J.; et al. Human melanocortin 1 receptor variants, receptor function and melanocyte response to UV radiation. J. Cell Sci. 2002, 115, 2349–2355. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Esparza, M.; Jimenez-Cervantes, C.; Solano, F.; Lozano, J.A.; Garcia-Borron, J.C. Mechanisms of melanogenesis inhibition by tumor necrosis factor-α in B16/F10 mouse melanoma cells. Eur. J. Biochem. 1998, 255, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Esparza, M.; Jiménez-Cervantes, C.; Beermann, F.; Aparicio, P.; Lozano, J.A.; García-Borrón, J.C. Transforming growth factor-β1 inhibits basal melanogenesis in B16/F10 mouse melanoma cells by increasing the rate of degradation of tyrosinase and tyrosinase-related protein-1. J. Biol. Chem. 1997, 272, 3967–3972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Esparza, M.; Solano, F.; García-Borrón, J.C. Independent regulation of tyrosinase by the hypopigmenting cytokines TGF beta1 and TNF α and the melanogenic hormone α-MSH in B16 mouse melanocytes. Cell. Mol. Biol. 1999, 45, 991–1000. [Google Scholar]
- Khaled, M.; Levy, C.; Fisher, D.E. Control of melanocyte differentiation by a MITF-PDE4D3 homeostatic circuit. Genes Dev. 2010, 24, 2276–2281. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Wan, L.; Hacker, E.; Dai, X.; Lenna, S.; Jimenez-Cervantes, C.; Wang, Y.; Leslie, N.R.; Xu, G.X.; Widlund, H.R.; et al. MC1R is a potent regulator of PTEN after UV exposure in melanocytes. Mol. Cell 2013, 51, 409–422. [Google Scholar] [CrossRef] [Green Version]
- Castejón-Griñán, M.; Herraiz, C.; Olivares, C.; Jiménez-Cervantes, C.; García-Borrón, J.C. CAMP-independent non-pigmentary actions of variant melanocortin 1 receptor: AKT-mediated activation of protective responses to oxidative DNA damage. Oncogene 2018, 37, 3631–3646. [Google Scholar] [CrossRef]
- Pérez-Oliva, A.B.; Olivares, C.; Jiménez-Cervantes, C.; García-Borrón, J. Mahogunin ring finger-1 (MGRN1) E3 ubiquitin ligase inhibits signaling from melanocortin receptor by competition with Gαs. J. Biol. Chem. 2009, 284, 31714–31725. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Hemesath, T.J.; Takemoto, C.M.; Horstmann, M.A.; Wells, A.G.; Price, E.R.; Fisher, D.Z.; Fisher, D.E. c-Kit triggers dual phosphorylations, which couple activation and degradation of the essential melanocyte factor Mi. Genes Dev. 2000, 14, 301–312. [Google Scholar] [CrossRef]
- Sánchez-Más, J.; Guillo, L.A.; Zanna, P.; Jiménez-Cervantes, C.; García-Borrón, J.C. Role of G protein-coupled receptor kinases in the homologous desensitization of the human and mouse melanocortin 1 receptors. Mol. Endocrinol. 2005, 19, 1035–1048. [Google Scholar] [CrossRef] [Green Version]
- Abrisqueta, M.; Herraiz, C.; Pérez Oliva, A.B.; Sanchez-Laorden, B.L.; Olivares, C.; Jiménez-Cervantes, C.; García-Borrón, J.C. Differential and competitive regulation of human melanocortin 1 receptor signaling by β-arrestin isoforms. J. Cell Sci. 2013, 126, 3724–3737. [Google Scholar] [CrossRef] [Green Version]
- Levy, C.; Khaled, M.; Fisher, D.E. MITF: Master regulator of melanocyte development and melanoma oncogene. Trends Mol. Med. 2006, 12, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Goding, C.R.; Arnheiter, H. Mitf—The first 25 years. Genes Dev. 2019, 33, 983–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, J.R.; Scurr, L.L.; Becker, T.M.; Kefford, R.F.; Rizos, H. The MAPK pathway functions as a redundant survival signal that reinforces the PI3K cascade in c-Kit mutant melanoma. Oncogene 2014, 33, 236–245. [Google Scholar] [CrossRef] [Green Version]
- Blume-Jensen, P.; Claesson-Welsh, L.; Siegbahn, A.; Zsebo, K.M.; Westermark Heldin, B.C.H. Activation of the human c-kit product by ligand-induced dimerization mediates circular actin reorganization and chemotaxis. EMBO J. 1991, 10, 4121–4128. [Google Scholar] [CrossRef] [PubMed]
- Lev, S.; Yarden, Y.; Givol, D. Dimerization and activation of the kit receptor by monovalent and bivalent binding of the stem cell factor. J. Biol. Chem. 1992, 267, 15970–15977. [Google Scholar] [CrossRef]
- Luo, D.; Chen, H.; Searles, G.; Jimbow, K. Coordinated mRNA expression of c-kit with tyrosinase and TRP-1 in melanin pigmentation of normal and malignant human melanocytes and transient activation of tyrosinase by kit/SCF-R. Melanoma Res. 1995, 5, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Hemesath, T.J.; Price, E.R.; Takemoto, C.; Badalian, T.; Fisher, D.E. MAP kinase links the transcription factor Microphthalmia to c-Kit signalling in melanocytes. Nature 1998, 391, 298–301. [Google Scholar] [CrossRef]
- Price, E.R.; Ding, H.F.; Badalian, T.; Bhattacharya, S.; Takemoto, C.; Yao, T.P.; Hemesath, T.J.; Fisher, D.E. Lineage-specific signaling in melanocytes. c-Kit stimulation recruits p300/CBP to microphthalmia. J. Biol. Chem. 1998, 273, 17983–17986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Gong, L.; Haddad, M.M.; Bischof, O.; Campisi, J.; Yeh, E.T.H.; Medrano, E.E. Regulation of microphthalmia-associated transcription factor MITF protein levels by association with the ubiquitin-conjugating enzyme hUBC9. Exp. Cell Res. 2000, 255, 135–143. [Google Scholar] [CrossRef]
- Alexeev, V.; Yoon, K. Distinctive role of the cKit receptor tyrosine kinase signaling in mammalian melanocytes. J. Investig. Dermatol. 2006, 126, 1102–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikawa, S.; Kusakabe, M.; Yoshinaga, K.; Ogawa, M.; Hayashi, S.I.; Kunisada, T.; Era, T.; Sakakura, T.; Nishikawa, S.I. In utero manipulation of coat color formation by a monoclonal anti-c-kit antibody: Two distinct waves of c-kit-dependency during melanocyte development. EMBO J. 1991, 10, 2111–2118. [Google Scholar] [CrossRef]
- Botchkareva, N.V.; Khlgatian, M.; Jack longley, B.; Botchkarev, V.A.; Gilchrest, B.A. SCF/c-kit signaling is required for cyclic regeneration of the hair pigmentation unit. FASEB J. 2001, 15, 645–658. [Google Scholar] [CrossRef] [Green Version]
- Boissan, M.; Feger, F.; Guillosson, J.J.; Arock, M. c-Kit and c-kit mutations in mastocytosis and other hematological diseases. J. Leukoc. Biol. 2000, 67, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Philo, J.S.; Wen, J.; Wypych, J.; Schwartz, M.G.; Mendiaz, E.A.; Langley, K.E. Human stem cell factor dimer forms a complex with two molecules of the extracellular domain of its receptor, Kit. J. Biol. Chem. 1996, 271, 6895–6902. [Google Scholar] [CrossRef] [Green Version]
- Lemmon, M.A.; Pinchasi, D.; Zhou, M.; Lax, I.; Schlessinger, J. Kit receptor dimerization is driven by bivalent binding of stem cell factor. J. Biol. Chem. 1997, 272, 6311–6317. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, Y.; Hirota, S. Kit as a human oncogenic tyrosine kinase. Cell. Mol. Life Sci. 2004, 61, 2924–2931. [Google Scholar] [CrossRef]
- Kuang, D.; Zhao, X.; Xiao, G.; Ni, J.; Feng, Y.; Wu, R.; Wang, G. Stem cell factor/c-kit signaling mediated cardiac stem cell migration via activation of p38 MAPK. Basic Res. Cardiol. 2008, 103, 265–273. [Google Scholar] [CrossRef]
- Liang, J.; Wu, Y.L.; Chen, B.J.; Zhang, W.; Tanaka, Y.; Sugiyama, H. The C-Kit receptor-mediated signal transduction and tumor-related diseases. Int. J. Biol. Sci. 2013, 9, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Herraiz, C.; Journé, F.; Abdel-Malek, Z.; Ghanem, G.; Jiménez-Cervantes, C.; García-Borrón, J.C. Signaling from the human melanocortin 1 receptor to ERK1 and ERK2 mitogen-activated protein kinases involves transactivation of cKIT. Mol. Endocrinol. 2011, 25, 138–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maresca, V.; Flori, E.; Picardo, M. Skin phototype: A new perspective. Pigment Cell Melanoma Res. 2015, 28, 378–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flori, E.; Mastrofrancesco, A.; Kovacs, D.; Bellei, B.; Briganti, S.; Maresca, V.; Cardinali, G.; Picardo, M. The activation of PPARγ by 2,4,6-Octatrienoic acid protects human keratinocytes from UVR-induced damages. Sci. Rep. 2017, 7, 9241. [Google Scholar] [CrossRef] [Green Version]
- Flori, E.; Rosati, E.; Cardinali, G.; Kovacs, D.; Bellei, B.; Picardo, M.; Maresca, V. The α-melanocyte stimulating hormone/peroxisome proliferator activated receptor-γ pathway down-regulates proliferation in melanoma cell lines. J. Exp. Clin. Cancer Res. 2017, 36, 142. [Google Scholar] [CrossRef] [Green Version]
- Herraiz, C.; Martínez-Vicente, I.; Maresca, V. The α-melanocyte-stimulating hormone/melanocortin-1 receptor interaction: A driver of pleiotropic effects beyond pigmentation. Pigment Cell Melanoma Res. 2021, 34, 748–761. [Google Scholar] [CrossRef]
- Hill, R.P.; MacNeil, S.; Haycock, J.W. Melanocyte stimulating hormone peptides inhibit TNF-α signaling in human dermal fibroblast cells. Peptides 2006, 27, 421–430. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J. Neuroendocrinology of the skin. Endocr. Rev. 2000, 21, 457–487. [Google Scholar] [CrossRef]
- Slominski, A.; Fischer, T.W.; Zmijewski, M.A.; Wortsman, J.; Semak, I.; Zbytek, B.; Slominski, R.M.; Tobin, D.J. On the role of melatonin in skin physiology and pathology. Endocrine 2005, 27, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Carreira, S.; Goodall, J.; Denat, L.; Rodriguez, M.; Nuciforo, P.; Hoek, K.S.; Testori, A.; Larue, L.; Goding, C.R. Mitf regulation of Dia1 controls melanoma proliferation and invasiveness. Genes Dev. 2006, 20, 3426–3439. [Google Scholar] [CrossRef] [Green Version]
- Giuliano, S.; Cheli, Y.; Ohanna, M.; Bonet, C.; Beuret, L.; Bille, K.; Loubat, A.; Hofman, V.; Hofman, P.; Ponzio, G.; et al. Microphthalmia-associated transcription factor controls the DNA damage response and a lineage-specific senescence program in melanomas. Cancer Res. 2010, 70, 3813–3822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivas-García, Y.; Falletta, P.; Liebing, J.; Louphrasitthiphol, P.; Feng, Y.; Chauhan, J.; Scott, D.A.; Glodde, N.; Chocarro-Calvo, A.; Bonham, S.; et al. Lineage-Restricted Regulation of SCD and Fatty Acid Saturation by MITF Controls Melanoma Phenotypic Plasticity. Mol. Cell 2020, 77, 120–137.e9. [Google Scholar] [CrossRef] [PubMed]
- Louphrasitthiphol, P.; Ledaki, I.; Chauhan, J.; Falletta, P.; Siddaway, R.; Buffa, F.M.; Mole, D.R.; Soga, T.; Goding, C.R. MITF controls the TCA cycle to modulate the melanoma hypoxia response. Pigment Cell Melanoma Res. 2019, 32, 792–808. [Google Scholar] [CrossRef]
- Abdel-Malek, Z.A.; Ruwe, A.; Kavanagh-Starner, R.; Kadekaro, A.L.; Swope, V.; Haskell-Luevano, C.; Koikov, L.; Knittel, J.J. α-MSH tripeptide analogs activate the melanocortin 1 receptor and reduce UV-induced DNA damage in human melanocytes. Pigment Cell Melanoma Res. 2009, 22, 635–644. [Google Scholar] [CrossRef]
- Scherer, D.; Bermejo, J.L.; Rudnai, P.; Gurzau, E.; Koppova, K.; Hemminki, K.; Kumar, R. MC1R variants associated susceptibility to basal cell carcinoma of skin: Interaction with host factors and XRCC3 polymorphism. Int. J. Cancer 2008, 122, 1787–1793. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, C.; Ter Huurne, J.; Berkhout, M.; Gruis, N.; Bastiaens, M.; Bergman, W.; Willemze, R.; Bouwes Bavinck, J.N. Melanocortin 1 receptor (MC1R) gene variants are associated with an increased risk for cutaneous melanoma which is largely independent of skin type and hair color. J. Investig. Dermatol. 2001, 117, 294–300. [Google Scholar] [CrossRef] [Green Version]
- Kadekaro, A.L.; Chen, J.; Yang, J.; Chen, S.; Jameson, J.; Swope, V.B.; Cheng, T.; Kadakia, M.; Abdel-Malek, Z. α-melanocyte-stimulating hormone suppresses oxidative stress through a p53-mediated signaling pathway in human melanocytes. Mol. Cancer Res. 2012, 10, 778–786. [Google Scholar] [CrossRef] [Green Version]
- Ferretta, A.; Maida, I.; Guida, S.; Azzariti, A.; Porcelli, L.; Tommasi, S.; Zanna, P.; Cocco, T.; Guida, M.; Guida, G. New insight into the role of metabolic reprogramming in melanoma cells harboring BRAF mutations. Biochim. Biophys. Acta—Mol. Cell Res. 2016, 1863, 2710–2718. [Google Scholar] [CrossRef]
- Vazquez, F.; Lim, J.H.; Chim, H.; Bhalla, K.; Girnun, G.; Pierce, K.; Clish, C.B.; Granter, S.R.; Widlund, H.R.; Spiegelman, B.M.; et al. PGC1α Expression Defines a Subset of Human Melanoma Tumors with Increased Mitochondrial Capacity and Resistance to Oxidative Stress. Cancer Cell 2013, 23, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Haq, R.; Shoag, J.; Andreu-Perez, P.; Yokoyama, S.; Edelman, H.; Rowe, G.C.; Frederick, D.T.; Hurley, A.D.; Nellore, A.; Kung, A.L.; et al. Oncogenic BRAF regulates oxidative metabolism via PGC1α and MITF. Cancer Cell 2013, 23, 302–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, A.; Hanawalt, P. Photobiological Origins of the Field of Genomic Maintenance. Photochem. Photobiol. 2016, 92, 52–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, P.; Wyrick, J.J.; Roberts, S.A.; Smerdon, M.J. UV-Induced DNA Damage and Mutagenesis in Chromatin. Photochem. Photobiol. 2017, 93, 216–228. [Google Scholar] [CrossRef]
- Cadet, J.; Douki, T. Formation of UV-induced DNA damage contributing to skin cancer development. Photochem. Photobiol. Sci. 2018, 17, 1816–1841. [Google Scholar] [CrossRef]
- Wolber, R.; Schlenz, K.; Wakamatsu, K.; Smuda, C.; Nakanishi, Y.; Hearing, V.J.; Ito, S. Pigmentation effects of solar-simulated radiation as compared with UVA and UVB radiation. Pigment Cell Melanoma Res. 2008, 21, 487–491. [Google Scholar] [CrossRef]
- Strzałka, W.; Zgłobicki, P.; Kowalska, E.; Bażant, A.; Dziga, D.; Banaś, A.K. The dark side of uv-induced DNA lesion repair. Genes 2020, 11, 1450. [Google Scholar] [CrossRef] [PubMed]
- Besaratinia, A.; Yoon, J.; Schroeder, C.; Bradforth, S.E.; Cockburn, M.; Pfeifer, G.P. Wavelength dependence of ultraviolet radiation-induced DNA damage as determined by laser irradiation suggests that cyclobutane pyrimidine dimers are the principal DNA lesions produced by terrestrial sunlight. FASEB J. 2011, 25, 3079–3091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, Y.H.; Lee, D.H.; Yoon, J.H.; Nakajima, S.; Yasui, A.; Pfeifer, G.P. Cyclobutane Pyrimidine Dimers Are Responsible for the Vast Majority of Mutations Induced by UVB Irradiation in Mammalian Cells. J. Biol. Chem. 2001, 276, 44688–44694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, S.P.; Bartek, J. The DNA-damage response in human biology and disease. Nature 2009, 461, 1071–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Walia, V.; Lin, J.C.; Teer, J.K.; Prickett, T.D.; Gartner, J.; Davis, S.; Stemke-Hale, K.; Davies, M.A.; Gershenwald, J.E.; et al. Exome sequencing identifies GRIN2A as frequently mutated in melanoma. Nat. Genet. 2011, 43, 442–446. [Google Scholar] [CrossRef] [Green Version]
- Pleasance, E.D.; Cheetham, R.K.; Stephens, P.J.; McBride, D.J.; Humphray, S.J.; Greenman, C.D.; Varela, I.; Lin, M.L.; Ordóñez, G.R.; Bignell, G.R.; et al. A comprehensive catalogue of somatic mutations from a human cancer genome. Nature 2010, 463, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Noonan, F.P.; Zaidi, M.R.; Wolnicka-Glubisz, A.; Anver, M.R.; Bahn, J.; Wielgus, A.; Cadet, J.; Douki, T.; Mouret, S.; Tucker, M.A.; et al. Melanoma induction by ultraviolet A but not ultraviolet B radiation requires melanin Pigment. Nat. Commun. 2012, 3, 884. [Google Scholar] [CrossRef] [Green Version]
- Beani, J.C. Ultraviolet A-induced DNA damage: Role in skin cancer. Bull. Acad. Natl. Med. 2014, 198, 273–295. [Google Scholar] [CrossRef] [PubMed]
- Kvam, E.; Tyrrell, R.M. The role of melanin in the induction of oxidative DNA base damage by ultraviolet A irradiation of DNA or melanoma cells. J. Investig. Dermatol. 1999, 113, 209–213. [Google Scholar] [CrossRef] [Green Version]
- Bachelor, M.A.; Bowden, G.T. UVA-mediated activation of signaling pathways involved in skin tumor promotion and progression. Semin. Cancer Biol. 2004, 14, 131–138. [Google Scholar] [CrossRef]
- Cadet, J.; Sage, E.; Douki, T. Ultraviolet radiation-mediated damage to cellular DNA. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2005, 571, 3–17. [Google Scholar] [CrossRef]
- Cadet, J.; Grand, A.; Douki, T. Solar uv radiation-induced DNA bipyrimidine photoproducts: Formation and mechanistic insights. Top. Curr. Chem. 2015, 356, 249–275. [Google Scholar] [CrossRef]
- Kadekaro, A.L.; Kavanagh, R.; Kanto, H.; Terzieva, S.; Hauser, J.; Kobayashi, N.; Schwemberger, S.; Cornelius, J.; Babcock, G.; Shertzer, H.G.; et al. α-melanocortin and endothelin-1 activate antiapoptotic pathways and reduce DNA damage in human melanocytes. Cancer Res. 2005, 65, 4292–4299. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, A.K.; Funasaka, Y.; Pawelek, J.M.; Nagahama, M.; Ito, A.; Ichihashi, M. Enhanced expression of melanocortin-1 receptor (MC1-R) in normal human keratinocytes during differentiation: Evidence for increased expression of POMC peptides near suprabasal layer of epidermis. J. Investig. Dermatol. 1999, 112, 853–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corre, S.; Primot, A.; Sviderskaya, E.; Bennett, D.C.; Vaulont, S.; Goding, C.R.; Galibert, M.D. UV-induced expression of key component of the tanning process, the POMC and MC1R genes, is dependent onp the p-38-activated upstream stimulating factor-1 (USF-1). J. Biol. Chem. 2004, 279, 51226–51233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiller, M.; Brzoska, T.; Böhm, M.; Metze, D.; Scholzen, T.E.; Rougier, A.; Luger, T.A. Solar-Simulated Ultraviolet Radiation-Induced Upregulation of the Melanocortin-1 Receptor, Proopiomelanocortin, and α-Melanocyte- Stimulating Hormone in Human Epidermis In Vivo. J. Investig. Dermatol. 2004, 122, 468–476. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, A.K.; Funasaka, Y.; Slominski, A.; Ermak, G.; Hwang, J.; Pawelek, J.M.; Ichihashi, M. Production and release of proopiomelanocortin (POMC) derived peptides by human melanocytes and keratinocytes in culture: Regulation by ultraviolet B. Biochim. Biophys. Acta—Mol. Cell Res. 1996, 1313, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Im, S.; Moro, O.; Peng, F.; Medrano, E.E.; Cornelius, J.; Babcock, G.; Nordlund, J.J.; Abdel-Malek, Z.A. Activation of the cyclic AMP pathway by α-melanotropin mediates the response of human melanocytes to ultraviolet B radiation. Cancer Res. 1998, 58, 47–54. [Google Scholar] [PubMed]
- Kadekaro, A.L.; Wakamatsu, K.; Ito, S.; Abdel-Malek, Z.A. Cutaneous photoprotection and melanoma susceptibility: Reaching beyond melanin content to the frontiers of DNA repair. Front. Biosci. 2006, 11, 2157–2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schärer, O.D. Nucleotide excision repair in Eukaryotes. Cold Spring Harb. Perspect. Biol. 2013, 5, a012609. [Google Scholar] [CrossRef]
- Sugasawa, K. Molecular mechanisms of DNA damage recognition for mammalian nucleotide excision repair. DNA Repair 2016, 44, 110–117. [Google Scholar] [CrossRef]
- Gillet, L.C.J.; Schärer, O.D. Molecular mechanisms of mammalian global genome nucleotide excision repair. Chem. Rev. 2006, 106, 253–276. [Google Scholar] [CrossRef]
- Spivak, G. Nucleotide excision repair in humans. DNA Repair 2015, 36, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Swope, V.; Alexander, C.; Starner, R.; Schwemberger, S.; Babcock, G.; Abdel-Malek, Z.A. Significance of the melanocortin 1 receptor in the DNA damage response of human melanocytes to ultraviolet radiation. Pigment Cell Melanoma Res. 2014, 27, 601–610. [Google Scholar] [CrossRef]
- Maddodi, N.; Setaluri, V. Role of UV in cutaneous melanoma. Photochem. Photobiol. 2008, 84, 528–536. [Google Scholar] [CrossRef]
- Li, X.; Mao, W.; Chen, J.; Goding, C.R.; Cui, R.; Xu, Z.X.; Miao, X. The protective role of MC1R in chromosome stability and centromeric integrity in melanocytes. Cell Death Discov. 2021, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.L.; Southgate, H.; Tweddle, D.A.; Curtin, N.J. DNA damage checkpoint kinases in cancer. Expert Rev. Mol. Med. 2020, 22, e2. [Google Scholar] [CrossRef] [PubMed]
- Scully, R.; Xie, A. Double strand break repair functions of histone H2AX. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2013, 750, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Jarrett, S.G.; Horrell, E.M.W.; Christian, P.A.; Vanover, J.C.; Boulanger, M.C.; Zou, Y.; D’Orazio, J.A. PKA-Mediated Phosphorylation of ATR Promotes Recruitment of XPA to UV-Induced DNA Damage. Mol. Cell 2014, 54, 999–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, S.; Fischer, E.S.; Yasuda, T.; Dohmae, N.; Iwai, S.; Mori, T.; Nishi, R.; Yoshino, K.I.; Sakai, W.; Hanaoka, F.; et al. Functional regulation of the DNA damage-recognition factor DDB2 by ubiquitination and interaction with xeroderma pigmentosum group C protein. Nucleic Acids Res. 2015, 43, 1700–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleaver, J.E. Cancer in xeroderma pigmentosum and related disorders of DNA repair. Nat. Rev. Cancer 2005, 5, 564–573. [Google Scholar] [CrossRef]
- Puumalainen, M.R.; Rüthemann, P.; Min, J.H.; Naegeli, H. Xeroderma pigmentosum group C sensor: Unprecedented recognition strategy and tight spatiotemporal regulation. Cell. Mol. Life Sci. 2016, 73, 547–566. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.G.; Luk, N.; Newton, R.A.; Roberts, D.W.; Sturm, R.A.; Muscat, G.E.O. Melanocortin-1 receptor signaling markedly induces the expression of the NR4A nuclear receptor subgroup in melanocytic cells. J. Biol. Chem. 2008, 283, 12564–12570. [Google Scholar] [CrossRef] [Green Version]
- Jagirdar, K.; Yin, K.; Harrison, M.; Lim, W.; Muscat, G.E.O.; Sturm, R.A.; Smith, A.G. The NR4A2 nuclear receptor is recruited to novel nuclear foci in response to UV irradiation and participates in nucleotide excision repair. PLoS ONE 2013, 8, e78075. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Malek, Z.A.; Swope, V.B.; Starner, R.J.; Koikov, L.; Cassidy, P.; Leachman, S. Melanocortins and the melanocortin 1 receptor, moving translationally towards melanoma prevention. Arch. Biochem. Biophys. 2014, 563, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, K.A.; Liu, Y.Y.; Sturm, R.A. Chapter 4 The Melanocortin-1 Receptor Gene Polymorphism and Association with Human Skin Cancer. Prog. Mol. Biol. Transl. Sci. 2009, 88, 85–153. [Google Scholar] [CrossRef]
- Gerstenblith, M.R.; Goldstein, A.M.; Fargnoli, M.C.; Peris, K.; Landi, M.T. Comprehensive evaluation of allele frequency differences of MC1R variants across populations. Hum. Mutat. 2007, 28, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Oliva, A.B.P.; Fernéndez, L.P.; de Torre, C.; Herráiz, C.; Martínez-Escribano, J.A.; Benítez, J.; Teruel, J.A.L.; Garcia-Borron, J.C.; Jiménez-Cervantes, C.; Ribas, G. Identification and functional analysis of novel variants of the human melanocortin 1 receptor found in melanoma patients. Hum. Mutat. 2009, 30, 811–822. [Google Scholar] [CrossRef] [PubMed]
- Valverde, P.; Healy, E.; Jackson, I.J.; Rees, J.L.; Thody, A.J. Variants of the melanocyte–stimulating hormone receptor gene are associated with red hair and fair skin in humans. Nat. Genet. 1995, 11, 328–330. [Google Scholar] [CrossRef] [PubMed]
- Box, N.F.; Wyeth, J.R.; O’Gorman, L.E.; Martin, N.G.; Sturm, R.A. Characterization of melanocyte stimulating hormone receptor variant alleles in twins with red hair. Hum. Mol. Genet. 1997, 6, 1891–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frändberg, P.-A.; Doufexis, M.; Kapas, S.; Chhajlani, V. Human pigmentation phenotype: A point mutation generates nonfunctional MSH receptor. Biochem. Biophys. Res. Commun. 1998, 245, 490–492. [Google Scholar] [CrossRef]
- Herraiz, C.; Garcia-Borron, J.C.; Jiménez-Cervantes, C.; Olivares, C. MC1R signaling. Intracellular partners and pathophysiological implications. Biochim. Biophys. Acta. Mol. Basis Dis. 2017, 1863, 2448–2461. [Google Scholar] [CrossRef]
- Beaumont, K.A.; Shekar, S.L.; Newton, R.A.; James, M.R.; Stow, J.L.; Duffy, D.L.; Sturm, R.A. Receptor function, dominant negative activity and phenotype correlations for MC1R variant alleles. Hum. Mol. Genet. 2007, 16, 2249–2260. [Google Scholar] [CrossRef]
- Herraiz, C.; Journé, F.; Ghanem, G.; Jiménez-Cervantes, C.; García-Borrón, J.C. Functional status and relationships of melanocortin 1 receptor signaling to the cAMP and extracellular signal-regulated protein kinases 1 and 2 pathways in human melanoma cells. Int. J. Biochem. Cell Biol. 2012, 44, 2244–2252. [Google Scholar] [CrossRef]
- Pasquali, E.; García-Borrón, J.C.; Fargnoli, M.C.; Gandini, S.; Maisonneuve, P.; Bagnardi, V.; Specchia, C.; Liu, F.; Kayser, M.; Nijsten, T.; et al. MC1R variants increased the risk of sporadic cutaneous melanoma in darker-pigmented Caucasians: A pooled-analysis from the M-SKIP project. Int. J. Cancer 2015, 136, 618–631. [Google Scholar] [CrossRef] [Green Version]
- Beaumont, K.A.; Newton, R.A.; Smit, D.J.; Leonard, J.H.; Stow, J.L.; Sturm, R.A. Altered cell surface expression of human MC1R variant receptor alleles associated with red hair and skin cancer risk. Hum. Mol. Genet. 2005, 14, 2145–2154. [Google Scholar] [CrossRef]
- Sánchez-Laorden, B.L.; Sánchez-MáS, J.; Turpín, M.C.; García-Borrón, J.C.; Jiménez-Cervantes, C. Variant amino acids in different domains of the human melanocortin 1 receptor impair cell surface expression. Cell. Mol. Biol. 2006, 52, 39–46. [Google Scholar] [CrossRef]
- Schiöth, H.B.; Phillips, S.R.; Rudzish, R.; Birch-Machin, M.A.; Wikberg, J.E.S.; Rees, J.L. Loss of function mutations of the human melanocortin 1 receptor are common and are associated with red hair. Biochem. Biophys. Res. Commun. 1999, 260, 488–491. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, N.; Healy, E.; Ray, A.; Philips, S.; Todd, C.; Jackson, I.J.; Birch-Machin, M.A.; Rees, J.L. Pleiotropic effects of the melanocortin 1 receptor (MC1R) gene on human pigmentation. Hum. Mol. Genet. 2000, 9, 2531–2537. [Google Scholar] [CrossRef] [Green Version]
- Morgan, M.D.; Pairo-Castineira, E.; Rawlik, K.; Canela-Xandri, O.; Rees, J.; Sims, D.; Tenesa, A.; Jackson, I.J. Genome-wide study of hair colour in UK Biobank explains most of the SNP heritability. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, J.L. The melanocortin 1 receptor (MC1R): More than just red hair. Pigment Cell Res. 2000, 13, 135–140. [Google Scholar] [CrossRef]
- Bastiaens, M.T.; Ter Huurne, J.A.C.; Kielich, C.; Gruis, N.A.; Westendorp, R.G.J.; Vermeer, B.J.; Bavinck, J.N.B.; Van Amsterdam, N.; Bergman, W.; Berkhout, M.; et al. Melanocortin-1 receptor gene variants determine the risk of nonmelanoma skin cancer independently of fair skin and red hair. Am. J. Hum. Genet. 2001, 68, 884–894. [Google Scholar] [CrossRef] [Green Version]
- Healy, E.; Flannagan, N.; Ray, A.; Todd, C.; Jackson, I.J.; Matthews, J.N.S.; Birch-Machin, M.A.; Rees, J.L. Melanocortin-1-receptor gene and sun sensitivity in individuals without red hair. Lancet 2000, 355, 1072–1073. [Google Scholar] [CrossRef]
- Guida, S.; Ciardo, S.; De Pace, B.; De Carvalho, N.; Peccerillo, F.; Manfredini, M.; Farnetani, F.; Chester, J.; Kaleci, S.; Manganelli, M.; et al. The influence of MC1R on dermal morphological features of photo-exposed skin in women revealed by reflectance confocal microscopy and optical coherence tomography. Exp. Dermatol. 2019, 28, 1321–1327. [Google Scholar] [CrossRef] [PubMed]
- Guida, S.; Ciardo, S.; De Pace, B.; De Carvalho, N.; Farnetani, F.; Pezzini, C.; Chester, J.; Shaniko, K.; Manganelli, M.; Guida, G.; et al. Atrophic and hypertrophic skin photoaging and melanocortin-1 receptor (MC1R): The missing link. J. Am. Acad. Dermatol. 2021, 84, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Fargnoli, M.C.; Sera, F.; Suppa, M.; Piccolo, D.; Landi, M.T.; Chiarugi, A.; Pellegrini, C.; Seidenari, S.; Peris, K. Dermoscopic features of cutaneous melanoma are associated with clinical characteristics of patients and tumours and with MC1R genotype. J. Eur. Acad. Dermatol. Venereol. 2014, 28, 1768–1775. [Google Scholar] [CrossRef] [PubMed]
- Vallone, M.G.; Tell-Marti, G.; Potrony, M.; Rebollo-Morell, A.; Badenas, C.; Puig-Butille, J.A.; Gimenez-Xavier, P.; Carrera, C.; Malvehy, J.; Puig, S. Melanocortin 1 receptor (MC1R) polymorphisms’ influence on size and dermoscopic features of nevi. Pigment Cell Melanoma Res. 2018, 31, 39–50. [Google Scholar] [CrossRef]
- Bassoli, S.; Maurichi, A.; Rodolfo, M.; Casari, A.; Frigerio, S.; Pupelli, G.; Farnetani, F.; Pelosi, G.; Santinami, M.; Pellacani, G. CDKN2A and MC1R variants influence dermoscopic and confocal features of benign melanocytic lesions in multiple melanoma patients. Exp. Dermatol. 2013, 22, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Amos, C.I.; Wang, L.-E.; Lee, J.E.; Gershenwald, J.E.; Chen, W.V.; Fang, S.; Kosoy, R.; Zhang, M.; Qureshi, A.A.; Vattathil, S.; et al. Genome-wide association study identifies novel loci predisposing to cutaneous melanoma. Hum. Mol. Genet. 2011, 20, 5012–5023. [Google Scholar] [CrossRef] [Green Version]
- Chatzinasiou, F.; Lill, C.M.; Kypreou, K.; Stefanaki, I.; Nicolaou, V.; Spyrou, G.; Evangelou, E.; Roehr, J.T.; Kodela, E.; Katsambas, A.; et al. Comprehensive field synopsis and systematic meta-analyses of genetic association studies in cutaneous melanoma. J. Natl. Cancer Inst. 2011, 103, 1227–1235. [Google Scholar] [CrossRef]
- Raimondi, S.; Sera, F.; Gandini, S.; Iodice, S.; Caini, S.; Maisonneuve, P.; Fargnoli, M.C. MC1R variants, melanoma and red hair color phenotype: A meta-analysis. Int. J. Cancer 2008, 122, 2753–2760. [Google Scholar] [CrossRef]
- Williams, P.F.; Olsen, C.M.; Hayward, N.K.; Whiteman, D.C. Melanocortin 1 receptor and risk of cutaneous melanoma: A meta-analysis and estimates of population burden. Int. J. Cancer 2011, 129, 1730–1740. [Google Scholar] [CrossRef]
- Dwyer, T.; Stankovich, J.M.; Blizzard, L.; FitzGerald, L.M.; Dickinson, J.L.; Reilly, A.; Williamson, J.; Ashbolt, R.; Berwick, M.; Sale, M.M. Does the addition of information on genotype improve prediction of the risk of melanoma and nonmelanoma skin cancer beyond that obtained from skin phenotype? Am. J. Epidemiol. 2004, 159, 826–833. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Kraft, P.; Colditz, G.A.; Wong, J.; Hunter, D.J. Melanocortin 1 receptor variants and skin cancer risk. Int. J. Cancer 2006, 119, 1976–1984. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.S.; Duffy, D.L.; Box, N.F.; Aitken, J.F.; O’Gorman, L.E.; Green, A.C.; Hayward, N.K.; Martin, N.G.; Sturm, R.A. Melanocortin-1 receptor polymorphisms and risk of melanoma: Is the association explained solely by pigmentation phenotype? Am. J. Hum. Genet. 2000, 66, 176–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, D.; Luo, X.; Morgan, A.; Wang, J.; Hoang, M.P.; Lo, J.; Guerrero, C.R.; Lennerz, J.K.; Mihm, M.C.; Wargo, J.A.; et al. An ultraviolet-radiation-independent pathway to melanoma carcinogenesis in the red hair/fair skin background. Nature 2012, 491, 449–453. [Google Scholar] [CrossRef] [PubMed]
- De Summa, S.; Lasorella, A.; Strippoli, S.; Giudice, G.; Guida, G.; Elia, R.; Nacchiero, E.; Azzariti, A.; Silvestris, N.; Guida, M.; et al. The Genetic Germline Background of Single and Multiple Primary Melanomas. Front. Mol. Biosci. 2021, 7, 555630. [Google Scholar] [CrossRef] [PubMed]
- Landi, M.T.; Kanetsky, P.A.; Tsang, S.; Gold, B.; Munroe, D.; Rebbeck, T.; Swoyer, J.; Ter-Minassian, M.; Hedayati, M.; Grossman, L.; et al. MC1R, ASIP, and DNA repair in sporadic and familial melanoma in a mediterranean population. J. Natl. Cancer Inst. 2005, 97, 998–1007. [Google Scholar] [CrossRef] [Green Version]
- Concetta Fargnoli, M.; Altobelli, E.; Keller, G.; Chimenti, S.; Höfler, H.; Peris, K. Contribution of melanocortin-1 receptor gene variants to sporadic cutaneous melanoma risk in a population in central Italy: A case-control study. Melanoma Res. 2006, 16, 175–182. [Google Scholar] [CrossRef]
- Fernandez, L.P.; Milne, R.L.; Bravo, J.; Lopez, J.M.; Avilés, J.A.; Longo, M.I.; Benítez, J.; Lázaro, P.; Ribas, G. MC1R: Three novel variants identified in a malignant melanoma association study in the Spanish population. Carcinogenesis 2007, 28, 1659–1664. [Google Scholar] [CrossRef]
- Ghiorzo, P.; Bonelli, L.; Pastorino, L.; Bruno, W.; Barile, M.; Andreotti, V.; Nasti, S.; Battistuzzi, L.; Grosso, M.; Bianchi-Scarrà, G.; et al. MC1R variation and melanoma risk in relation to host/clinical and environmental factors in CDKN2A positive and negative melanoma patients. Exp. Dermatol. 2012, 21, 718–720. [Google Scholar] [CrossRef]
- Kanetsky, P.A.; Panossian, S.; Elder, D.E.; Guerry, D.P.; Ming, M.E.; Schuchter, L.; Rebbeck, T.R. Does MC1R genotype convey information about melanoma risk beyond risk phenotypes? Cancer 2010, 116, 2416–2428. [Google Scholar] [CrossRef] [Green Version]
- Guida, S.; Bartolomeo, N.; Zanna, P.T.; Grieco, C.; Maida, I.; De Summa, S.; Tommasi, S.; Guida, M.; Azzariti, A.; Foti, C.; et al. Sporadic melanoma in South-Eastern Italy: The impact of melanocortin 1 receptor (MC1R) polymorphism analysis in low-risk people and report of three novel variants. Arch. Dermatol. Res. 2015, 307, 495–503. [Google Scholar] [CrossRef]
- Wendt, J.; Rauscher, S.; Burgstaller-Muehlbacher, S.; Fae, I.; Fischer, G.; Pehamberger, H.; Okamoto, I. Human determinants and the role of melanocortin-1 receptor variants in melanoma risk independent of UV Radiation Exposure. JAMA Dermatol. 2016, 152, 776–782. [Google Scholar] [CrossRef] [Green Version]
- Fargnoli, M.C.; Gandini, S.; Peris, K.; Maisonneuve, P.; Raimondi, S. MC1R variants increase melanoma risk in families with CDKN2A mutations: A meta-analysis. Eur. J. Cancer 2010, 46, 1413–1420. [Google Scholar] [CrossRef] [PubMed]
- Dȩbniak, T.; Scott, R.; Masojc, B.; Serrano-Fernández, P.; Huzarski, T.; Byrski, T.; Dȩbniak, B.; Górski, B.; Cybulski, C.; Mȩdrek, K.; et al. MC1R common variants, CDKN2A and their association with melanoma and breast cancer risk. Int. J. Cancer 2006, 119, 2597–2602. [Google Scholar] [CrossRef]
- Robles-Espinoza, C.D.; Roberts, N.D.; Chen, S.; Leacy, F.P.; Alexandrov, L.B.; Pornputtapong, N.; Halaban, R.; Krauthammer, M.; Cui, R.; Timothy Bishop, D.; et al. Germline MC1R status influences somatic mutation burden in melanoma. Nat. Commun. 2016, 7, 12064. [Google Scholar] [CrossRef]
- Johansson, P.A.; Pritchard, A.L.; Patch, A.-M.; Wilmott, J.S.; Pearson, J.V.; Waddell, N.; Scolyer, R.A.; Mann, G.J.; Hayward, N.K. Mutation load in melanoma is affected by MC1R genotype. Pigment Cell Melanoma Res. 2017, 30, 255–258. [Google Scholar] [CrossRef]
- Tagliabue, E.; Gandini, S.; Bellocco, R.; Maisonneuve, P.; Newton-Bishop, J.; Polsky, D.; Lazovich, D.; Kanetsky, P.A.; Ghiorzo, P.; Gruis, N.A.; et al. MC1R variants as melanoma risk factors independent of at-risk phenotypic characteristics: A pooled analysis from the M-SKIP project. Cancer Manag. Res. 2018, 10, 1143–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, M.R.; Eliades, P.; Gupta, S.; Grant-Kels, J.M.; Tsao, H. Cutaneous melanoma in women. Int. J. Women’s Dermatol. 2017, 3, S11–S15. [Google Scholar] [CrossRef]
- Lu, C.; Zhang, J.; Nagahawatte, P.; Easton, J.; Lee, S.; Liu, Z.; Ding, L.; Wyczalkowski, M.A.; Valentine, M.; Navid, F.; et al. The genomic landscape of childhood and adolescent melanoma. J. Investig. Dermatol. 2015, 135, 816–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniotti, M.; Ferrari, A.; Frigerio, S.; Casieri, P.; Miselli, F.; Zucca, E.; Collini, P.; Della Torre, G.; Manoukian, S.; Peissel, B.; et al. Cutaneous melanoma in childhood and adolescence shows frequent loss of INK4A and gain of KIT. J. Investig. Dermatol. 2009, 129, 1759–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabbie, R.; Rashid, M.; Arance, A.M.; Sánchez, M.; Tell-Marti, G.; Potrony, M.; Conill, C.; van Doorn, R.; Dentro, S.; Gruis, N.A.; et al. Genomic analysis and clinical management of adolescent cutaneous melanoma. Pigment Cell Melanoma Res. 2017, 30, 307–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrini, C.; Botta, F.; Massi, D.; Martorelli, C.; Facchetti, F.; Gandini, S.; Maisonneuve, P.; Avril, M.F.; Demenais, F.; Bressac-de Paillerets, B.; et al. MC1R variants in childhood and adolescent melanoma: A retrospective pooled analysis of a multicentre cohort. Lancet Child. Adolesc. Heal. 2019, 3. [Google Scholar] [CrossRef] [Green Version]
- Preston, D.S.; Stern, R.S. Nonmelanoma Cancers of the Skin. N. Engl. J. Med. 1992, 327, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Kricker, A.; Armstrong, B.K.; English, D.R. Sun exposure and non-melanocytic skin cancer. Cancer Causes Control 1994, 5, 367–392. [Google Scholar] [CrossRef] [PubMed]
- Kricker, A.; Armstrong, B.K.; English, D.R.; Heenan, P.J. Pigmentary and cutaneous risk factors for non-melanocytic skin cancer—A case-control study. Int. J. Cancer 1991, 48, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Tagliabue, E.; Fargnoli, M.C.; Gandini, S.; Maisonneuve, P.; Liu, F.; Kayser, M.; Nijsten, T.; Han, J.; Kumar, R.; Gruis, N.A.; et al. MC1R gene variants and non-melanoma skin cancer: A pooled-analysis from the M-SKIP project. Br. J. Cancer 2015, 113, 354–363. [Google Scholar] [CrossRef] [Green Version]
- Box, N.F.; Duffy, D.L.; Irving, R.E.; Russell, A.; Chen, W.; Griffyths, L.R.; Parsons, P.G.; Green, A.C.; Sturm, R.A. Melanocortin-1 receptor genotype is a risk factor for basal and squamous cell carcinoma. J. Investig. Dermatol. 2001, 116, 224–229. [Google Scholar] [CrossRef]
- Puig, S.; Malvehy, J.; Badenas, C.; Ruiz, A.; Jimenez, D.; Cuellar, F.; Azon, A.; Gonzàlez, U.; Castel, T.; Campoy, A.; et al. Role of the CDKN2A locus in patients with multiple primary melanomas. J. Clin. Oncol. 2005, 23, 3043–3051. [Google Scholar] [CrossRef]
- Liontos, M.; Pateras, I.S.; Evangelou, K.; G. Gorgoulis, V. The Tumor Suppressor Gene ARF as a Sensor of Oxidative Stress. Curr. Mol. Med. 2012, 12, 704–715. [Google Scholar] [CrossRef]
- Hussussian, C.J.; Struewing, J.P.; Goldstein, A.M.; Higgins, P.A.T.; Ally, D.S.; Sheahan, M.D.; Clark, W.H.; Tucker, M.A.; Dracopoli, N.C. Germline p16 mutations in familial melanoma. Nat. Genet. 1994, 8, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Bishop, D.T.; Demenais, F.; Goldstein, A.M.; Bergman, W.; Bishop, J.N.; Bressac-De Paillerets, B.; Chompret, A.; Ghiorzo, P.; Gruis, N.; Hansson, J.; et al. Geographical variation in the penetrance of CDKN2A mutations for melanoma. J. Natl. Cancer Inst. 2002, 94, 894–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, A.M.; Chan, M.; Harland, M.; Hayward, N.K.; Demenais, F.; Bishop, D.T.; Azizi, E.; Bergman, W.; Bianchi-Scarra, G.; Bruno, W.; et al. Features associated with germline CDKN2A mutations: A GenoMEL study of melanoma-prone families from three continents. J. Med. Genet. 2007, 44, 99–106. [Google Scholar] [CrossRef]
- Lang, J.; Boxer, M.; MacKie, R.M. CDKN2A mutations in Scottish families with cutaneous melanoma: Results from 32 newly identified families. Br. J. Dermatol. 2005, 153, 1121–1125. [Google Scholar] [CrossRef]
- Borges, A.L.; Cuéllar, F.; Puig-Butillé, J.A.; Scarone, M.; Delgado, L.; Badenas, C.; Milà, M.; Malvehy, J.; Barquet, V.; Núñez, J.; et al. CDKN2A mutations in melanoma families from Uruguay. Br. J. Dermatol. 2009, 161, 536–541. [Google Scholar] [CrossRef]
- Yakobson, E.; Eisenberg, S.; Isacson, R.; Halle, D.; Levy-Lahad, E.; Catane, R.; Safro, M.; Sobolev, V.; Huot, T.; Peters, G.; et al. A single Mediterranean, possibly Jewish, origin for the Val59Gly CDKN2A mutation in four melanoma-prone families. Eur. J. Hum. Genet. 2003, 11, 288–296. [Google Scholar] [CrossRef] [Green Version]
- Majore, S.; De Simone, P.; Crisi, A.; Eibenschutz, L.; Binni, F.; Antigoni, I.; De Bernardo, C.; Catricalà, C.; Grammatico, P. CDKN2A/CDK4 molecular study on 155 Italian subjects with familial and/or primary multiple melanoma. Pigment Cell Melanoma Res. 2008, 21, 209–211. [Google Scholar] [CrossRef]
- Eliason, M.J.; Larson, A.A.; Florell, S.R.; Zone, J.J.; Cannon-Albright, L.A.; Samlowski, W.E.; Leachman, S.A. Population-based prevalence of CDKN2A mutations in Utah melanoma families. J. Investig. Dermatol. 2006, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Box, N.F.; Duffy, D.L.; Chen, W.; Stark, M.; Martin, N.G.; Sturm, R.A.; Hayward, N.K. MC1R genotype modifies risk of melanoma in families segregating CDKN2A mutations. Am. J. Hum. Genet. 2001, 69, 660–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sturm, R.A. Skin colour and skin cancer-MC1R, the genetic link. Melanoma Res. 2002, 12, 405–416. [Google Scholar] [CrossRef]
- Chaudru, V.; Laud, K.; Avril, M.F.; Minière, A.; Chompret, A.; Bressac-De Paillerets, B.; Demenais, F. Melanocortin-1 receptor (MC1R) gene variants and dysplastic nevi modify penetrance of CDKN2A mutations in French melanoma-prone pedigrees. Cancer Epidemiol. Biomarkers Prev. 2005, 14, 2384–2390. [Google Scholar] [CrossRef] [Green Version]
- Helsing, P.; Nymoen, D.A.; Rootwelt, H.; Vårdal, M.; Akslen, L.A.; Molven, A.; Andresen, P.A. MC1R, ASIP, TYR, and TYRP1 gene variants in a population-based series of multiple primary melanomas. Genes Chromosom. Cancer 2012, 51, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Demenais, F.; Mohamdi, H.; Chaudru, V.; Goldstein, A.M.; Bishop, J.A.N.; Bishop, D.T.; Kanetsky, P.A.; Hayward, N.K.; Gillanders, E.; Elder, D.E.; et al. Association of MC1R variants and host phenotypes with melanoma risk in CDKN2A mutation carriers: A GenoMEL study. J. Natl. Cancer Inst. 2010, 102, 1568–1583. [Google Scholar] [CrossRef]
- Van der Velden, P.A.; Sandkuijl, L.A.; Bergman, W.; Pavel, S.; Van Mourik, L.; Frants, R.R.; Gruis, N.A. Melanocortin-1 receptor variant R151C modifies melanoma risk in Dutch families with melanoma. Am. J. Hum. Genet. 2001, 69, 774–779. [Google Scholar] [CrossRef] [Green Version]
- Aoude, L.G.; Wadt, K.A.W.; Pritchard, A.L.; Hayward, N.K. Genetics of familial melanoma: 20 years after CDKN2A. Pigment Cell Melanoma Res. 2015, 28, 148–160. [Google Scholar] [CrossRef]
- Fargnoli, M.C.; Pike, K.; Pfeiffer, R.M.; Tsang, S.; Rozenblum, E.; Munroe, D.J.; Golubeva, Y.; Calista, D.; Seidenari, S.; Massi, D.; et al. MC1R variants increase risk of melanomas harboring BRAF mutations. J. Investig. Dermatol. 2008, 128, 2485–2490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landi, M.T.; Bauer, J.; Pfeiffer, R.M.; Elder, D.E.; Hulley, B.; Minghetti, P.; Calista, D.; Kanetsky, P.A.; Pinkel, D.; Bastian, B.C. MC1R germline variants confer risk for BRAF-mutant melanoma. Science 2006, 313, 521–522. [Google Scholar] [CrossRef] [PubMed]
- Hacker, E.; Olsen, C.M.; Kvaskoff, M.; Pandeya, N.; Yeo, A.; Green, A.C.; Williamson, R.M.; Triscott, J.; Wood, D.; Mortimore, R.; et al. Histologic and Phenotypic Factors and MC1R Status Associated with BRAFV600E, BRAFV600K, and NRAS Mutations in a Community-Based Sample of 414 Cutaneous Melanomas. J. Investig. Dermatol. 2016, 136, 829–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrini, C.; Di Nardo, L.; Cipolloni, G.; Martorelli, C.; De Padova, M.; Antonini, A.; Maturo, M.G.; Del Regno, L.; Strafella, S.; Micantonio, T.; et al. Heterogeneity of BRAF, NRAS, and TERT Promoter Mutational Status in Multiple Melanomas and Association with MC1R Genotype: Findings from Molecular and Immunohistochemical Analysis. J. Mol. Diagn. 2018, 20, 110–122. [Google Scholar] [CrossRef] [Green Version]
- William, B.; Claudia, M.; Bruna, D.; Virginia, A.; Lorenza, P.; Francesco, C.; Marina, G.; Francesco, S.; Alberto, B.; Paola, Q.; et al. Combining molecular and immunohistochemical analyses of key drivers in primary melanomas: Interplay between germline and somatic variations. Oncotarget 2018, 9, 5691–5702. [Google Scholar] [CrossRef] [Green Version]
- García-Casado, Z.; Traves, V.; Bañuls, J.; Niveiro, M.; Gimeno-Carpio, E.; Jimenez-Sanchez, A.I.; Moragõn, M.; Onrubia, J.A.; Oliver, V.; Kumar, R.; et al. BRAF, NRAS and MC1R status in a prospective series of primary cutaneous melanoma. Br. J. Dermatol. 2015, 172, 1128–1131. [Google Scholar] [CrossRef] [PubMed]
- Scherer, D.; Rachakonda, P.S.; Angelini, S.; Mehnert, F.; Sucker, A.; Egberts, F.; Hauschild, A.; Hemminki, K.; Schadendorf, D.; Kumar, R. Association between the germline MC1R variants and somatic BRAF/NRAS mutations in melanoma tumors. J. Investig. Dermatol. 2010, 130, 2844–2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacker, E.; Hayward, N.K.; Dumenil, T.; James, M.R.; Whiteman, D.C. The association between MC1R genotype and BRAF mutation status in cutaneous melanoma: Findings from an australian population. J. Investig. Dermatol. 2010, 130, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Thomas, N.E.; Kanetsky, P.A.; Edmiston, S.N.; Alexander, A.; Begg, C.B.; Groben, P.A.; Hao, H.; Busam, K.; Ollila, D.W.; Berwick, M.; et al. Relationship between germline MC1R variants and BRAF-mutant melanoma in a North Carolina population-based study. J. Investig. Dermatol. 2010, 130, 1463–1465. [Google Scholar] [CrossRef] [Green Version]
- Hacker, E.; Nagore, E.; Cerroni, L.; Woods, S.L.; Hayward, N.K.; Chapman, B.; Montgomery, G.W.; Soyer, H.P.; Whiteman, D.C. NRAS and BRAF mutations in cutaneous melanoma and the association with MC1R genotype: Findings from Spanish and Austrian populations. J. Investig. Dermatol. 2013, 133, 1027–1033. [Google Scholar] [CrossRef]
- Kosiniak-Kamysz, A.; Marczakiewicz-Lustig, A.; Marcińska, M.; Skowron, M.; Wojas-Pelc, A.; Pośpiech, E.; Branicki, W. Increased risk of developing cutaneous malignant melanoma is associated with variation in pigmentation genes and VDR, and may involve epistatic effects. Melanoma Res. 2014, 24, 388–396. [Google Scholar] [CrossRef]
- Nagore, E.; Reyes-Garcia, D.; Heidenreich, B.; Garcia-Casado, Z.; Requena, C.; Kumar, R. TERT promoter mutations associate with MC1R variants in melanoma patients. Pigment Cell Melanoma Res. 2017, 30, 273–275. [Google Scholar] [CrossRef]
- Duffy, D.L.; Zhao, Z.Z.; Sturm, R.A.; Hayward, N.K.; Martin, N.G.; Montgomery, G.W. Multiple pigmentation gene polymorphisms account for a substantial proportion of risk of cutaneous malignant melanoma. J. Investig. Dermatol. 2010, 130, 520–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liboutet, M.; Portela, M.; Delestaing, G.; Vilmer, C.; Dupin, N.; Gorin, I.; Saiag, P.; Lebbé, C.; Kerob, D.; Dubertret, L.; et al. MC1R and PTCH gene polymorphism in French patients with basal cell carcinomas. J. Investig. Dermatol. 2006, 126, 1510–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gujar, H.; Weisenberger, D.J.; Liang, G. The roles of human DNA methyltransferases and their isoforms in shaping the epigenome. Genes 2019, 10, 172. [Google Scholar] [CrossRef] [Green Version]
- Cappetta, M.; Berdasco, M.; Hochmann, J.; Bonilla, C.; Sans, M.; Hidalgo, P.C.; Artagaveytia, N.; Kittles, R.; Martínez, M.; Esteller, M.; et al. Effect of genetic ancestry on leukocyte global DNA methylation in cancer patients. BMC Cancer 2015, 15, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grönniger, E.; Weber, B.; Heil, O.; Peters, N.; Stäb, F.; Wenck, H.; Korn, B.; Winnefeld, M.; Lyko, F. Aging and chronic sun exposure cause distinct epigenetic changes in human skin. PLoS Genet. 2010, 6, e1000971. [Google Scholar] [CrossRef] [Green Version]
- Holzscheck, N.; Söhle, J.; Schläger, T.; Falckenhayn, C.; Grönniger, E.; Kolbe, L.; Wenck, H.; Terstegen, L.; Kaderali, L.; Winnefeld, M.; et al. Concomitant DNA methylation and transcriptome signatures define epidermal responses to acute solar UV radiation. Sci. Rep. 2020, 10, 12918. [Google Scholar] [CrossRef]
- Vandiver, A.R.; Irizarry, R.A.; Hansen, K.D.; Garza, L.A.; Runarsson, A.; Li, X.; Chien, A.L.; Wang, T.S.; Leung, S.G.; Kang, S.; et al. Age and sun exposure-related widespread genomic blocks of hypomethylation in nonmalignant skin. Genome Biol. 2015, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Nikolouzakis, T.K.; Falzone, L.; Lasithiotakis, K.; Krüger-Krasagakis, S.; Kalogeraki, A.; Sifaki, M.; Spandidos, D.A.; Chrysos, E.; Tsatsakis, A.; Tsiaoussis, J. Current and Future Trends in Molecular Biomarkers for Diagnostic, Prognostic, and Predictive Purposes in Non-Melanoma Skin Cancer. J. Clin. Med. 2020, 9, 2868. [Google Scholar] [CrossRef]
- Jones, P.A.; Baylin, S.B. The fundamental role of epigenetic events in cancer. Nat. Rev. Genet. 2002, 3, 415–428. [Google Scholar] [CrossRef]
- Riefolo, M.; Porcellini, E.; Dika, E.; Broseghini, E.; Ferracin, M. Interplay between small and long non-coding RNAs in cutaneous melanoma: A complex jigsaw puzzle with missing pieces. Mol. Oncol. 2019, 13, 74–98. [Google Scholar] [CrossRef]
- Sang, Y.; Deng, Y. Current insights into the epigenetic mechanisms of skin cancer. Dermatol. Ther. 2019, 32, e12964. [Google Scholar] [CrossRef]
- Budden, T.; Bowden, N.A. MC1R CpG island regulates MC1R expression and is methylated in a subset of melanoma tumours. Pigment Cell Melanoma Res. 2019, 32, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martínez, S.; Márquez, R.; Inácio, Â.; Galván, I. Changes in melanocyte RNA and DNA methylation favour pheomelanin synthesis and may avoid systemic oxidative stress after dietary cysteine supplementation in birds. Mol. Ecol. 2019, 28, 1030–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Luo, M.; Wang, L.; Yin, H.; Zhu, W.; Fu, J. MicroRNA-206 Regulation of Skin Pigmentation in Koi Carp (Cyprinus carpio L.). Front. Genet. 2020, 11, 47. [Google Scholar] [CrossRef] [Green Version]
- Aspinwall, L.G.; Leaf, S.L.; Dola, E.R.; Kohlmann, W.; Leachman, S.A. CDKN2A/p16 genetic test reporting improves early detection intentions and practices in high-risk melanoma families. Cancer Epidemiol. Biomark. Prev. 2008, 17, 1510–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glanz, K.; Volpicelli, K.; Kanetsky, P.A.; Ming, M.E.; Schuchter, L.M.; Jepson, C.; Domchek, S.M.; Armstrong, K. Melanoma genetic testing, counseling, and adherence to skin cancer prevention and detection behaviors. Cancer Epidemiol. Biomark. Prev. 2013, 22, 607–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hay, J.L.; Zielaskowski, K.; White, K.M.; Kaphingst, K.; Robers, E.; Guest, D.; Sussman, A.; Talamantes, Y.; Schwartz, M.; Rodríguez, V.M.; et al. Interest and uptake of MC1R testing for melanoma risk in a diverse primary care population a randomized clinical trial. JAMA Dermatol. 2018, 154, 684–693. [Google Scholar] [CrossRef] [Green Version]
- Kaphingst, K.A.; Khan, E.; White, K.M.; Sussman, A.; Guest, D.; Schofield, E.; Dailey, Y.T.; Robers, E.; Schwartz, M.R.; Li, Y.; et al. Effects of health literacy skills, educational attainment, and level of melanoma risk on responses to personalized genomic testing. Patient Educ. Couns. 2021, 104, 12–19. [Google Scholar] [CrossRef]
- Guida, M.; Strippoli, S.; Ferretta, A.; Bartolomeo, N.; Porcelli, L.; Maida, I.; Azzariti, A.; Tommasi, S.; Grieco, C.; Guida, S.; et al. Detrimental effects of melanocortin-1 receptor (MC1R) variants on the clinical outcomes of BRAF V600 metastatic melanoma patients treated with BRAF inhibitors. Pigment Cell Melanoma Res. 2016, 29, 679–687. [Google Scholar] [CrossRef]
- Koikov, L.; Starner, R.J.; Swope, V.B.; Upadhyay, P.; Hashimoto, Y.; Freeman, K.T.; Knittel, J.J.; Haskell-Luevano, C.; Abdel-Malek, Z.A. Development of hMC1R Selective Small Agonists for Sunless Tanning and Prevention of Genotoxicity of UV in Melanocytes. J. Investig. Dermatol. 2021, 141, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Bautista, R.M.; Carter, K.M.; Jarrett, S.G.; Napier, D.; Wakamatsu, K.; Ito, S.; D’Orazio, J.A. Cutaneous pharmacologic cAMP induction induces melanization of the skin and improves recovery from ultraviolet injury in melanocortin 1 receptor-intact or heterozygous skin. Pigment Cell Melanoma Res. 2020, 33, 30–40. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, J.A.; Nobuhisa, T.; Cui, R.; Arya, M.; Spry, M.; Wakamatsu, K.; Igras, V.; Kunisada, T.; Granter, S.R.; Nishimura, E.K.; et al. Topical drug rescue strategy and skin protection based on the role of Mc1r in UV-induced tanning. Nature 2006, 443, 340–344. [Google Scholar] [CrossRef]
- Langendonk, J.G.; Balwani, M.; Anderson, K.E.; Bonkovsky, H.L.; Anstey, A.V.; Bissell, D.M.; Bloomer, J.; Edwards, C.; Neumann, N.J.; Parker, C.; et al. Afamelanotide for Erythropoietic Protoporphyria. N. Engl. J. Med. 2015, 373, 48–59. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manganelli, M.; Guida, S.; Ferretta, A.; Pellacani, G.; Porcelli, L.; Azzariti, A.; Guida, G. Behind the Scene: Exploiting MC1R in Skin Cancer Risk and Prevention. Genes 2021, 12, 1093. https://doi.org/10.3390/genes12071093
Manganelli M, Guida S, Ferretta A, Pellacani G, Porcelli L, Azzariti A, Guida G. Behind the Scene: Exploiting MC1R in Skin Cancer Risk and Prevention. Genes. 2021; 12(7):1093. https://doi.org/10.3390/genes12071093
Chicago/Turabian StyleManganelli, Michele, Stefania Guida, Anna Ferretta, Giovanni Pellacani, Letizia Porcelli, Amalia Azzariti, and Gabriella Guida. 2021. "Behind the Scene: Exploiting MC1R in Skin Cancer Risk and Prevention" Genes 12, no. 7: 1093. https://doi.org/10.3390/genes12071093
APA StyleManganelli, M., Guida, S., Ferretta, A., Pellacani, G., Porcelli, L., Azzariti, A., & Guida, G. (2021). Behind the Scene: Exploiting MC1R in Skin Cancer Risk and Prevention. Genes, 12(7), 1093. https://doi.org/10.3390/genes12071093