

Bta-miR-6531 Regulates Calcium Influx in Bovine Leydig Cells and Is Associated with Sperm Motility
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bta-miR-6531 Target Gene Prediction and Kyoto Encyclopedia of Genes and Genomes (KEGG) Analysis
2.3. Reporter Plasmid Construction and Dual-Luciferase Reporter Assay
2.4. Separation, Culture, and Identification of Bovine Leydig Cells
2.5. Measurement of Calcium in Transfected miR-6531 Bovine Leydig Cells
2.6. Total RNA Isolation, cDNA Synthesis, and qPCR
2.7. Protein Extraction and Western Blotting
2.8. Sperm Samples
2.9. SNP Identification and Genotype Analysis
2.10. Pre-bta-miR-6531 Structure Prediction and bta-miR-6531 Expression Analysis
2.11. Statistics Analysis
3. Results
3.1. miR-6531 Targets the Calcium Signaling Pathway and the cAMP Signaling Pathway
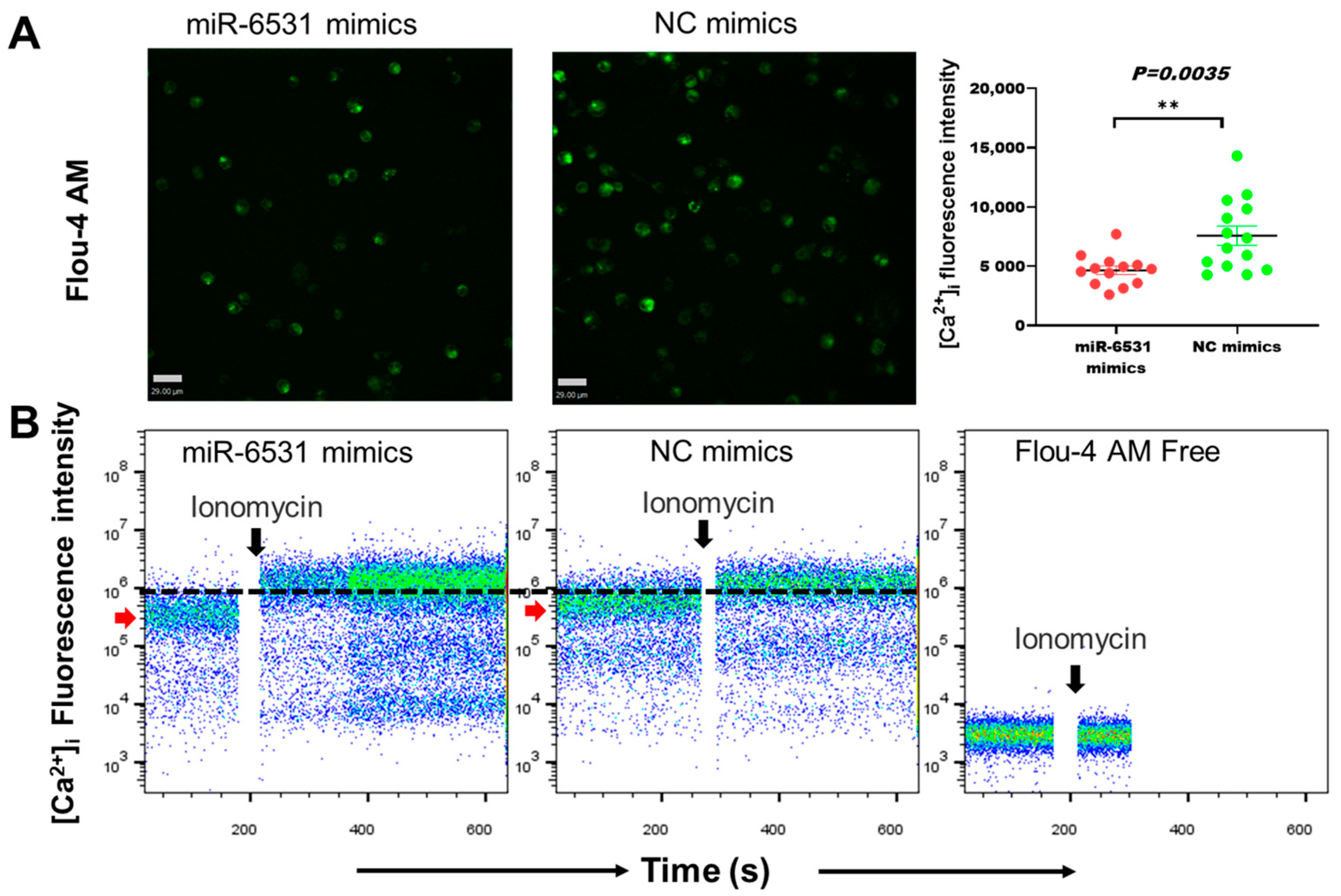
3.2. miR-6531 Regulated Calcium Flex into Cells
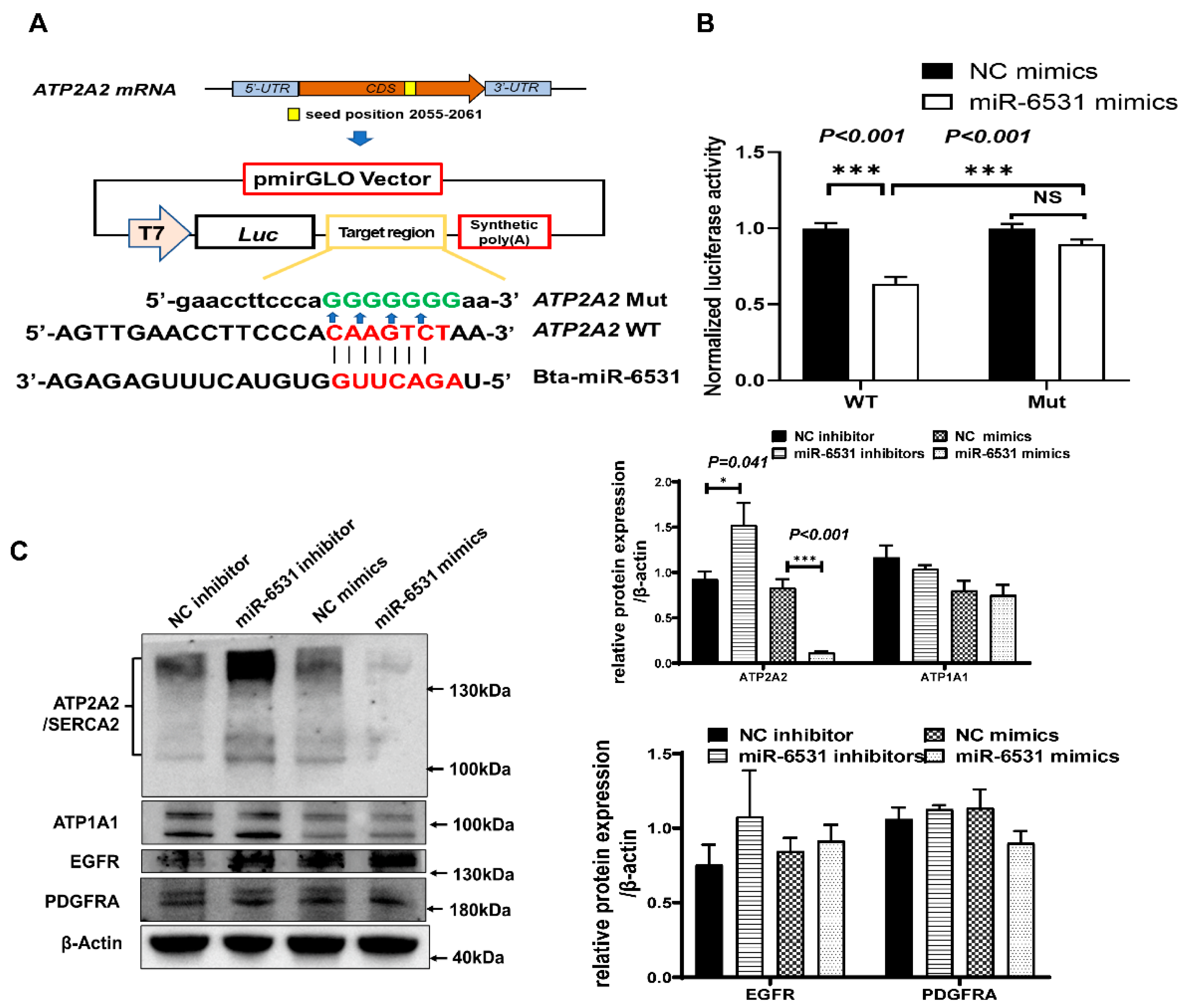
3.3. Bta-miR-6531 Binds and Downregulates ATP2A2 Expression in Bovine Leydig Cells
3.4. ATP2A2 Different Expression Effeceted Calcium Loading into Bovine Leydig Cells
3.5. Sequencing Analysis and SNPs Detection
3.6. Linkage between SNPs of the Pre-bta-miR-6531 is Associated with Sperm Quality
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Capra, E.; Turri, F.; Lazzari, B.; Cremonesi, P.; Gliozzi, T.M.; Fojadelli, I.; Stella, A.; Pizzi, F. Small RNA sequencing of cryopreserved semen from single bull revealed altered miRNAs and piRNAs expression between High- and Low-motile sperm populations. BMC Genom. 2017, 18, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.; Kirby, J.A.; Chu, C.; Gou, L.-T. Small Noncoding RNAs in Reproduction and Infertility. Biomedicines 2021, 9, 1884. [Google Scholar] [CrossRef] [PubMed]
- Sellem, E.; Marthey, S.; Rau, A.; Jouneau, L.; Bonnet, A.; Le Danvic, C.; Guyonnet, B.; Kiefer, H.; Jammes, H.; Schibler, L. Dynamics of cattle sperm sncRNAs during maturation, from testis to ejaculated sperm. Epigenetics Chromatin 2021, 14, 24. [Google Scholar] [CrossRef]
- Keles, E.; Malama, E.; Bozukova, S.; Siuda, M.; Wyck, S.; Witschi, U.; Bauersachs, S.; Bollwein, H. The micro-RNA content of unsorted cryopreserved bovine sperm and its relation to the fertility of sperm after sex-sorting. BMC Genom. 2021, 22, 30. [Google Scholar] [CrossRef]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J.; Lee, J.; Provost, P.; Rådmark, O.; Kim, S.; et al. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Druet, T.; Fritz, S.; Sellem, E.; Basso, B.; Gérard, O.; Salas-Cortes, L.; Humblot, P.; Druart, X.; Eggen, A. Estimation of genetic parameters and genome scan for 15 semen characteristics traits of Holstein bulls. J. Anim. Breed. Genet. 2009, 126, 269–277. [Google Scholar] [CrossRef]
- Guo, F.; Yang, B.; Ju, Z.H.; Wang, X.G.; Qi, C.; Zhang, Y.; Wang, C.F.; Liu, H.D.; Feng, M.Y.; Chen, Y.; et al. Alternative splicing, promoter methylation, and functional SNPs of sperm flagella 2 gene in testis and mature spermatozoa of Holstein bulls. Reproduction 2014, 147, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Tong, Y.; Zhang, H.-M.; Wang, K.; Hu, T.; Shan, G.; Sun, J.; Guo, A.-Y. Genome-wide identification of SNPs in microRNA genes and the SNP effects on microRNA target binding and biogenesis. Hum. Mutat. 2012, 33, 254–263. [Google Scholar] [CrossRef]
- Ryan, B.M.; Robles, A.I.; Harris, C.C. Genetic variation in microRNA networks: The implications for cancer research. Nat. Rev. Cancer 2010, 10, 389–402. [Google Scholar] [CrossRef] [Green Version]
- Dikeakos, P.; Theodoropoulos, G.; Rizos, S.; Tzanakis, N.; Zografos, G.; Gazouli, M. Association of the miR-146aC>G, miR-149T>C, and miR-196a2T>C polymorphisms with gastric cancer risk and survival in the Greek population. Mol. Biol. Rep. 2014, 41, 1075–1080. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Zhao, H.; Li, R.; Zhang, Y.; Liu, Y.; Wang, J.; Wang, X.; Ju, Z.; Liu, W.; Hou, M.; et al. In silico genome-wide miRNA-QTL-SNPs analyses identify a functional SNP associated with mastitis in Holsteins. BMC Genet. 2019, 20, 46. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- McGeary, S.E.; Lin, K.S.; Shi, C.Y.; Pham, T.M.; Bisaria, N.; Kelley, G.M.; Bartel, D.P. The biochemical basis of microRNA targeting efficacy. Science 2019, 366, eaav1741. [Google Scholar] [CrossRef]
- Sticht, C.; Carolina, D.; Parveen, A.; Gretz, N.; Campbell, M. miRWalk: An online resource for prediction of microRNA binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef] [PubMed]
- Tsikolia, N.; Merkwitz, C.; Sass, K.; Sakurai, M.; Spanel-Borowski, K.; Ricken, A.M. Characterization of bovine fetal Leydig cells by KIT expression. Histochem. Cell Biol. 2009, 132, 623–632. [Google Scholar] [CrossRef]
- Roszkowski, M.; Mansuy, I.M. High Efficiency RNA Extraction From Sperm Cells Using Guanidinium Thiocyanate Supplemented With Tris(2-Carboxyethyl)Phosphine. Front. Cell Dev. Biol. 2021, 9, 648274. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Cassinello, J.; Abaigar, T.; Gomendio, M.; Roldan, E.R. Characteristics of the semen of three endangered species of gazelles (Gazella dama mhorr, G. dorcas neglecta and G. cuvieri). J. Reprod. Fertil. 1998, 113, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, R.; Bernhart, S.H.; Höner zu Siederdissen, C.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef]
- Ammar, O.; Houas, Z.; Mehdi, M. The association between iron, calcium, and oxidative stress in seminal plasma and sperm quality. Environ. Sci. Pollut. Res. 2019, 26, 14097–14105. [Google Scholar] [CrossRef]
- Suarez, S.S. Control of hyperactivation in sperm. Hum. Reprod. Update 2008, 14, 647–657. [Google Scholar] [CrossRef] [Green Version]
- Lawson, C.; Dorval, V.; Goupil, S.; Leclerc, P. Identification and localisation of SERCA 2 isoforms in mammalian sperm. Mol. Hum. Reprod. 2007, 13, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Henningsen, M.B.; McWhan, K.; Dam, V.S.; Mele, M.; Hauerslev, K.R.; Voss, N.C.S.; Dabir, P.D.; Balling, E.; Pedersen, H.L.; Vahl, P.; et al. Amplified Ca2+ dynamics and accelerated cell proliferation in breast cancer tissue during purinergic stimulation. Int. J. Cancer 2022, 151, 1150–1165. [Google Scholar] [CrossRef]
- Darszon, A.; Nishigaki, T.; Wood, C.; Treviño, C.L.; Felix, R.; Beltrán, C. Calcium channels and Ca2+ fluctuations in sperm physiology. Int. Rev. Cytol. 2005, 243, 79–172. [Google Scholar] [CrossRef]
- Mata-Martínez, E.; Sánchez-Cárdenas, C.; Chávez, J.C.; Guerrero, A.; Treviño, C.L.; Corkidi, G.; Montoya, F.; Hernandez-Herrera, P.; Buffone, M.G.; Balestrini, P.A.; et al. Role of calcium oscillations in sperm physiology. Biosystems 2021, 209, 104524. [Google Scholar] [CrossRef]
- Kotaja, N. MicroRNAs and spermatogenesis. Fertil. Steril. 2014, 101, 1552–1562. [Google Scholar] [CrossRef]
- Wang, M.; Du, Y.; Gao, S.; Wang, Z.; Qu, P.; Gao, Y.; Wang, J.; Liu, Z.; Zhang, J.; Zhang, Y.; et al. Sperm-borne miR-202 targets SEPT7 and regulates first cleavage of bovine embryos via cytoskeletal remodeling. Development 2021, 148, dev189670. [Google Scholar] [CrossRef]
- Alves, M.B.R.; Celeghini, E.C.C.; Belleannée, C. From Sperm Motility to Sperm-Borne microRNA Signatures: New Approaches to Predict Male Fertility Potential. Front. Cell Dev. Biol. 2020, 8, 791. [Google Scholar] [CrossRef]
- Liu, X.; Ju, Z.; Wang, L.; Zhang, Y.; Huang, J.; Li, Q.; Li, J.; Zhong, J.; An, L.; Wang, C. Six novel single-nucleotide polymorphisms in SPAG11 gene and their association with sperm quality traits in Chinese Holstein bulls. Anim. Reprod. Sci. 2011, 129, 14–21. [Google Scholar] [CrossRef]
- Preskill, C.; Weidhaas, J.B. SNPs in microRNA binding sites as prognostic and predictive cancer biomarkers. Crit. Rev. Oncog. 2013, 18, 327–340. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Allele Frequency | Ejaculate Volume (mL) | Fresh Sperm Motility (%) | Density (108 mL−1) | Frozen Sperm Motility (%) | Deformity Rate (%) |
|---|---|---|---|---|---|---|
| TCTC/ATCT (n = 37) | 6.46 ± 0.23 | 69.16 ± 0.55 ab | 12.79 ± 0.44 ab | 43.76 ± 1.05 | 16.45 ± 0.50 ab | |
| ATCT (n = 12) | 0.54 | 6.05 ± 0.55 | 67.02 ± 1.00 b | 11.67 ± 0.62 b | 43.26 ± 1.41 | 17.90 ± 1.15 a |
| TCTC (n = 7) | 0.46 | 6.80 ± 0.50 | 70.28 ± 0.66 a | 14.37 ± 0.70 a | 44.51 ± 2.24 | 14.37 ± 0.57 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, Q.; Ding, X.; Xia, S.; Zhao, F.; Chen, K.; Qian, Y.; Cao, S.; Lin, Z.; Gao, Y.; Wang, H.; et al. Bta-miR-6531 Regulates Calcium Influx in Bovine Leydig Cells and Is Associated with Sperm Motility. Genes 2022, 13, 1788. https://doi.org/10.3390/genes13101788
Ding Q, Ding X, Xia S, Zhao F, Chen K, Qian Y, Cao S, Lin Z, Gao Y, Wang H, et al. Bta-miR-6531 Regulates Calcium Influx in Bovine Leydig Cells and Is Associated with Sperm Motility. Genes. 2022; 13(10):1788. https://doi.org/10.3390/genes13101788
Chicago/Turabian StyleDing, Qiang, Xiuhu Ding, Shuwen Xia, Fang Zhao, Kunlin Chen, Yong Qian, Shaoxian Cao, Zhiping Lin, Yundong Gao, Huili Wang, and et al. 2022. "Bta-miR-6531 Regulates Calcium Influx in Bovine Leydig Cells and Is Associated with Sperm Motility" Genes 13, no. 10: 1788. https://doi.org/10.3390/genes13101788
APA StyleDing, Q., Ding, X., Xia, S., Zhao, F., Chen, K., Qian, Y., Cao, S., Lin, Z., Gao, Y., Wang, H., & Zhong, J. (2022). Bta-miR-6531 Regulates Calcium Influx in Bovine Leydig Cells and Is Associated with Sperm Motility. Genes, 13(10), 1788. https://doi.org/10.3390/genes13101788