
Sorted Bulls’ X-Chromosome-Bearing Spermatozoa Show Increased GAPDHS Activity Correlating with Motility

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Samples
2.2. Western Blot Analysis of GAPDHS Content in Samples
2.3. GAPDHS Activity Assay
2.4. Spermatozoa Motility Analysis
2.5. Statistical Analysis
3. Results
3.1. The Quantity of GAPDHS Enzyme per Cell in Conventional Semen Samples and Samples Enriched with X-Chromosome-Bearing Spermatozoa Does Not Differ
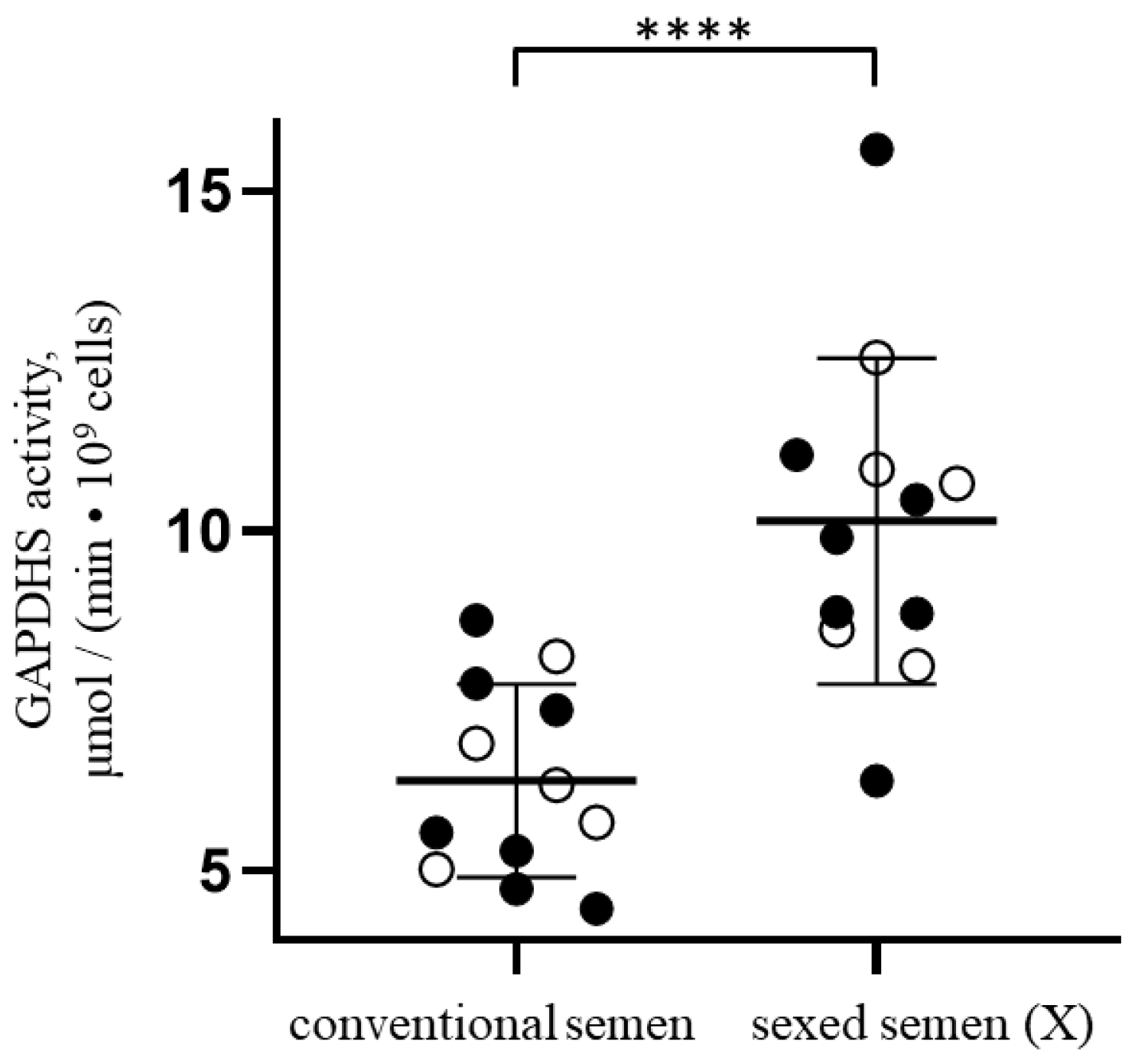
3.2. GAPDHS Enzymatic Activity in Conventional Semen Samples Is Lower Than in Samples Enriched with X-Chromosome-Bearing Spermatozoa
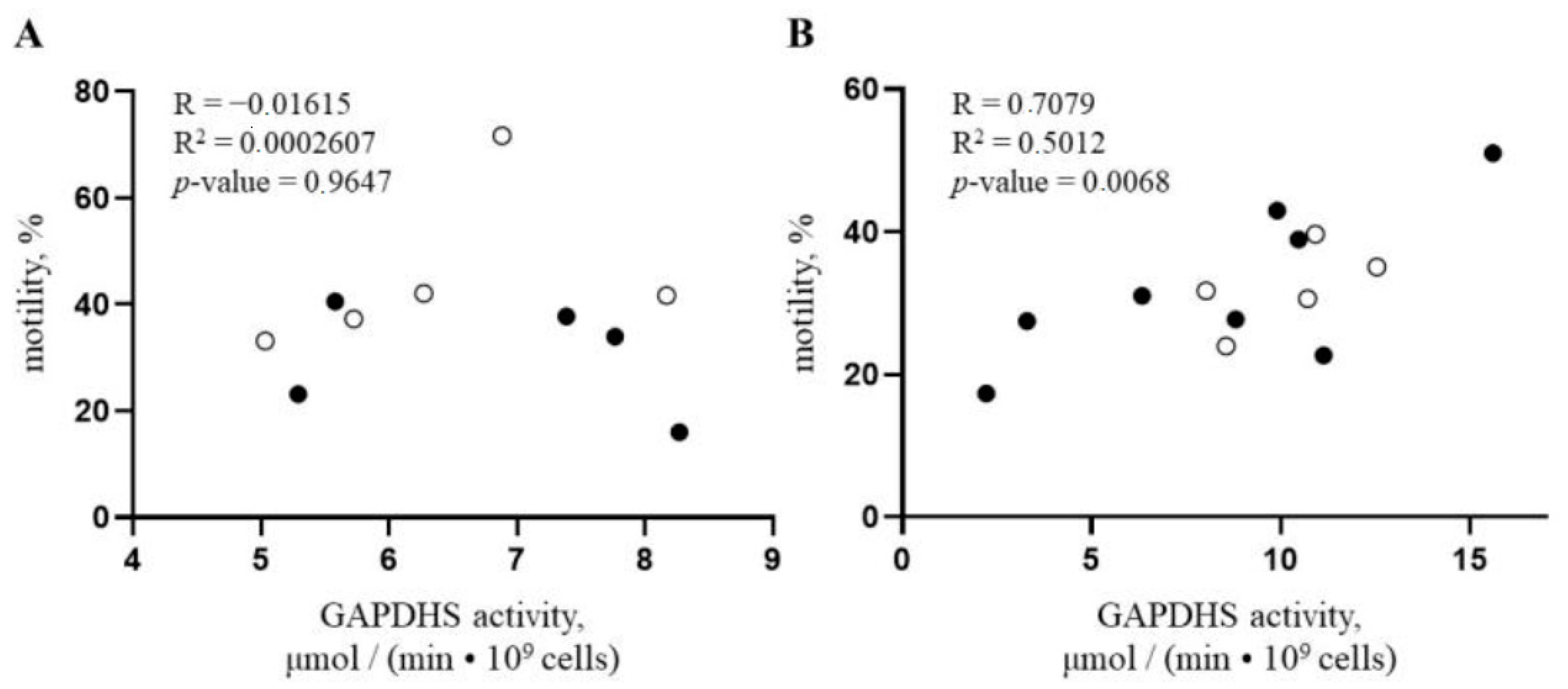
3.3. GAPDHS Enzymatic Activity in Sexed Semen Samples Positively Correlates with Spermatozoa Motility
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Steele, H.; Makri, D.; Maalouf, W.E.; Reese, S.; Kölle, S. Bovine Sperm Sexing Alters Sperm Morphokinetics and Subsequent Early Embryonic Development. Sci. Rep. 2020, 10, 6255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vishwanath, R.; Moreno, J.F. Review: Semen Sexing—Current State of the Art with Emphasis on Bovine Species. Animal 2018, 12, s85–s96. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, J.C.; Evans, K.M. Advances in Flow Cytometry for Sperm Sexing. Theriogenology 2009, 71, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Utt, M.D. Prediction of Bull Fertility. Anim. Reprod Sci. 2016, 169, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Muronetz, V.I.; Kuravsky, M.L.; Barinova, K.V.; Schmalhausen, E.V. Sperm-Specific Glyceraldehyde-3-Phosphate Dehydrogenase—An Evolutionary Acquisition of Mammals. Biochemistry 2015, 80, 1672–1689. [Google Scholar] [CrossRef]
- Mukai, C.; Okuno, M. Glycolysis Plays a Major Role for Adenosine Triphosphate Supplementation in Mouse Sperm Flagellar Movement. Biol. Reprod. 2004, 71, 540–547. [Google Scholar] [CrossRef]
- Miki, K.; Qu, W.; Goulding, E.H.; Willis, W.D.; Bunch, D.O.; Strader, L.F.; Perreault, S.D.; Eddy, E.M.; O’Brien, D.A. Glyceraldehyde 3-Phosphate Dehydrogenase-S, a Sperm-Specific Glycolytic Enzyme, Is Required for Sperm Motility and Male Fertility. Proc. Natl. Acad. Sci. USA. 2004, 101, 16501–16506. [Google Scholar] [CrossRef] [Green Version]
- De Canio, M.; Soggiu, A.; Piras, C.; Bonizzi, L.; Galli, A.; Urbani, A.; Roncada, P. Differential Protein Profile in Sexed Bovine Semen: Shotgun Proteomics Investigation. Mol. Biosyst. 2014, 10, 1264–1271. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Kuravsky, M.L.; Schmalhausen, E.V.; Pozdnyakova, N.V.; Muronetz, V.I. Isolation of Antibodies against Different Protein Conformations Using Immunoaffinity Chromatography. Anal. Biochem. 2012, 426, 47–53. [Google Scholar] [CrossRef]
- Welch, J.E.; Brown, P.L.; O’Brien, D.A.; Magyar, P.L.; Bunch, D.O.; Mori, C.; Eddy, E.M. Human Glyceraldehyde 3-Phosphate Dehydrogenase-2 Gene Is Expressed Specifically in Spermatogenic Cells. J. Androl. 2000, 21, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Morrell, J.M. Artificial insemination: Current and future trends. In Artificial Insemination in Farm Animals; Manafi, M., Ed.; InTech: London, UK, 2011; ISBN 978-953-307-312-5. [Google Scholar]
- Gürler, H.; Malama, E.; Heppelmann, M.; Calisici, O.; Leiding, C.; Kastelic, J.P.; Bollwein, H. Effects of Cryopreservation on Sperm Viability, Synthesis of Reactive Oxygen Species, and DNA Damage of Bovine Sperm. Theriogenology 2016, 86, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Reese, S.; Pirez, M.C.; Steele, H.; Kölle, S. The Reproductive Success of Bovine Sperm after Sex-Sorting: A Meta-Analysis. Sci. Rep. 2021, 11, 17366. [Google Scholar] [CrossRef] [PubMed]
- Bucci, D.; Galeati, G.; Tamanini, C.; Vallorani, C.; Rodriguez-Gil, J.E.; Spinaci, M. Effect of Sex Sorting on CTC Staining, Actin Cytoskeleton and Tyrosine Phosphorylation in Bull and Boar Spermatozoa. Theriogenology 2012, 77, 1206–1216. [Google Scholar] [CrossRef]
- Mostek, A.; Janta, A.; Ciereszko, A. Proteomic Comparison of Non-Sexed and Sexed (X-Bearing) Cryopreserved Bull Semen. Anim. Reprod. Sci. 2020, 221, 106552. [Google Scholar] [CrossRef]
- Hamano, K. Sex Preselection in Bovine by Separation of X- and Y-Chromosome Bearing Spermatozoa. J. Reprod. Dev. 2007, 53, 27–38. [Google Scholar] [CrossRef] [Green Version]
- King, W.A.; Yadav, B.R.; Xu, K.P.; Picard, L.; Sirard, M.A.; Verini Supplizi, A.; Betteridge, K.J. The Sex Ratios of Bovine Embryos Produced in Vivo and in Vitro. Theriogenology 1991, 36, 779–788. [Google Scholar] [CrossRef]
- Chen, X.; Yue, Y.; He, Y.; Zhu, H.; Hao, H.; Zhao, X.; Qin, T.; Wang, D. Identification and Characterization of Genes Differentially Expressed in X and Y Sperm Using Suppression Subtractive Hybridization and CDNA Microarray. Mol. Reprod. Dev. 2014, 81, 908–917. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, J.; Sun, W.; Ding, R.; Li, X.; Shangguan, A.; Zhou, Y.; Worku, T.; Hao, X.; Khan, F.A.; et al. Differences in Small Noncoding RNAs Profile between Bull X and Y Sperm. PeerJ 2020, 8, e9822. [Google Scholar] [CrossRef]
- Chen, X.; Zhu, H.; Wu, C.; Han, W.; Hao, H.; Zhao, X.; Du, W.; Qin, T.; Liu, Y.; Wang, D. Identification of Differentially Expressed Proteins between Bull X and Y Spermatozoa. J. Proteom. 2012, 77, 59–67. [Google Scholar] [CrossRef]
- Scott, C.; de Souza, F.F.; Aristizabal, V.H.V.; Hethrington, L.; Krisp, C.; Molloy, M.; Baker, M.A.; Dell’Aqua, J.A. Proteomic Profile of Sex-Sorted Bull Sperm Evaluated by SWATH-MS Analysis. Anim. Reprod. Sci. 2018, 198, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, L.; Li, Y.; Zhao, N.; Zhen, L.; Fu, J.; Yang, Q. Calcium Regulates Motility and Protein Phosphorylation by Changing CAMP and ATP Concentrations in Boar Sperm In Vitro. Anim. Reprod. Sci. 2016, 172, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-Y.; Zhao, Y.-H.; Hao, H.-S.; Wang, H.-Y.; Huang, J.-M.; Yan, C.-L.; Du, W.-H.; Pang, Y.-W.; Zhang, P.-P.; Liu, Y.; et al. Resveratrol Significantly Improves the Fertilisation Capacity of Bovine Sex-Sorted Semen by Inhibiting Apoptosis and Lipid Peroxidation. Sci. Rep. 2018, 8, 7603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenz, R.; Moreno, J.; Vishwanath, R. Compositions and Methods for Improving the Quality of Processed Sperm 2017. U.S. Patent 9781919B2, 10 October 2017. [Google Scholar]
- Baty, J.W.; Hampton, M.B.; Winterbourn, C.C. Proteomic Detection of Hydrogen Peroxide-Sensitive Thiol Proteins in Jurkat Cells. Biochem. J. 2005, 389, 785–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, R.M. Moving to the beat: A review of mammalian sperm motility regulation. Reprod. Fertil. Dev. 2006, 18, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Elkina, Y.L.; Atroshchenko, M.M.; Bragina, E.E.; Muronetz, V.I.; Schmalhausen, E.V. Oxidation of Glyceraldehyde-3-Phosphate Dehydrogenase Decreases Sperm Motility. Biochemistry (Moscow) 2011, 76, 268–272. [Google Scholar] [CrossRef]
- Hashemitabar, M.; Sabbagh, S.; Orazizadeh, M.; Ghadiri, A.; Bahmanzadeh, M. A Proteomic Analysis on Human Sperm Tail: Comparison between Normozoospermia and Asthenozoospermia. J. Assist. Reprod. Genet. 2015, 32, 853–863. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapitonova, A.A.; Muronetz, V.I.; Pozdyshev, D.V. Sorted Bulls’ X-Chromosome-Bearing Spermatozoa Show Increased GAPDHS Activity Correlating with Motility. Genes 2023, 14, 235. https://doi.org/10.3390/genes14010235
Kapitonova AA, Muronetz VI, Pozdyshev DV. Sorted Bulls’ X-Chromosome-Bearing Spermatozoa Show Increased GAPDHS Activity Correlating with Motility. Genes. 2023; 14(1):235. https://doi.org/10.3390/genes14010235
Chicago/Turabian StyleKapitonova, Anna A., Vladimir I. Muronetz, and Denis V. Pozdyshev. 2023. "Sorted Bulls’ X-Chromosome-Bearing Spermatozoa Show Increased GAPDHS Activity Correlating with Motility" Genes 14, no. 1: 235. https://doi.org/10.3390/genes14010235
APA StyleKapitonova, A. A., Muronetz, V. I., & Pozdyshev, D. V. (2023). Sorted Bulls’ X-Chromosome-Bearing Spermatozoa Show Increased GAPDHS Activity Correlating with Motility. Genes, 14(1), 235. https://doi.org/10.3390/genes14010235