


Identification and Molecular Characterization of RWP-RK Transcription Factors in Soybean


 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Genome-Wide Identification and Physicochemical Properties of Soybean RWP-RK Genes
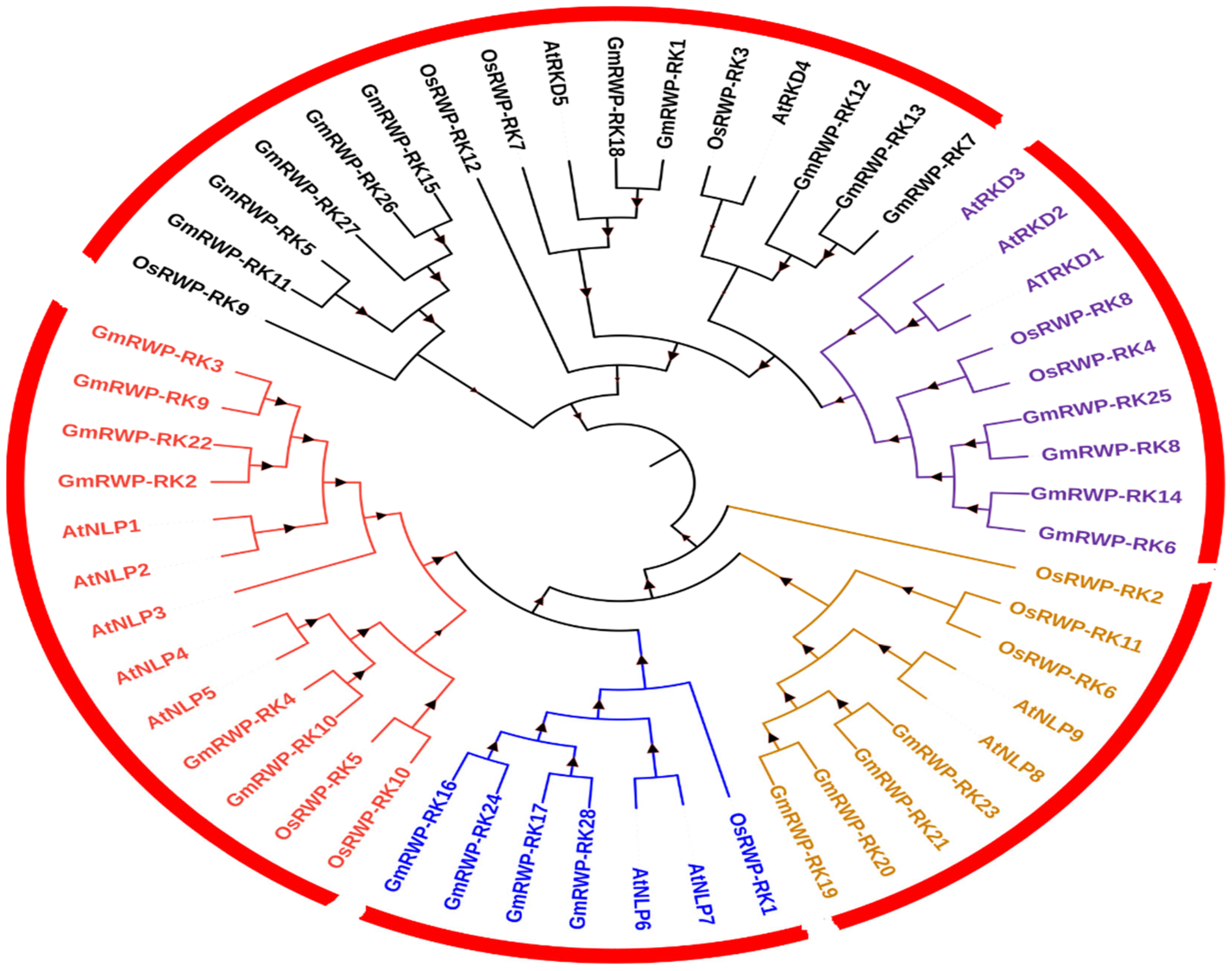
2.2. Phylogenetic Relationships of RWP-RKs and Chromosomal Localization
2.3. Conserved Motif Identification in RWP-RK Proteins
2.4. Analysis of the Cis-Elements in RWP-RK Promoters
2.5. GO Enrichment Analysis of RWP-RK Genes
2.6. Expression Analysis of RWP-RK Genes Family in Different Tissues
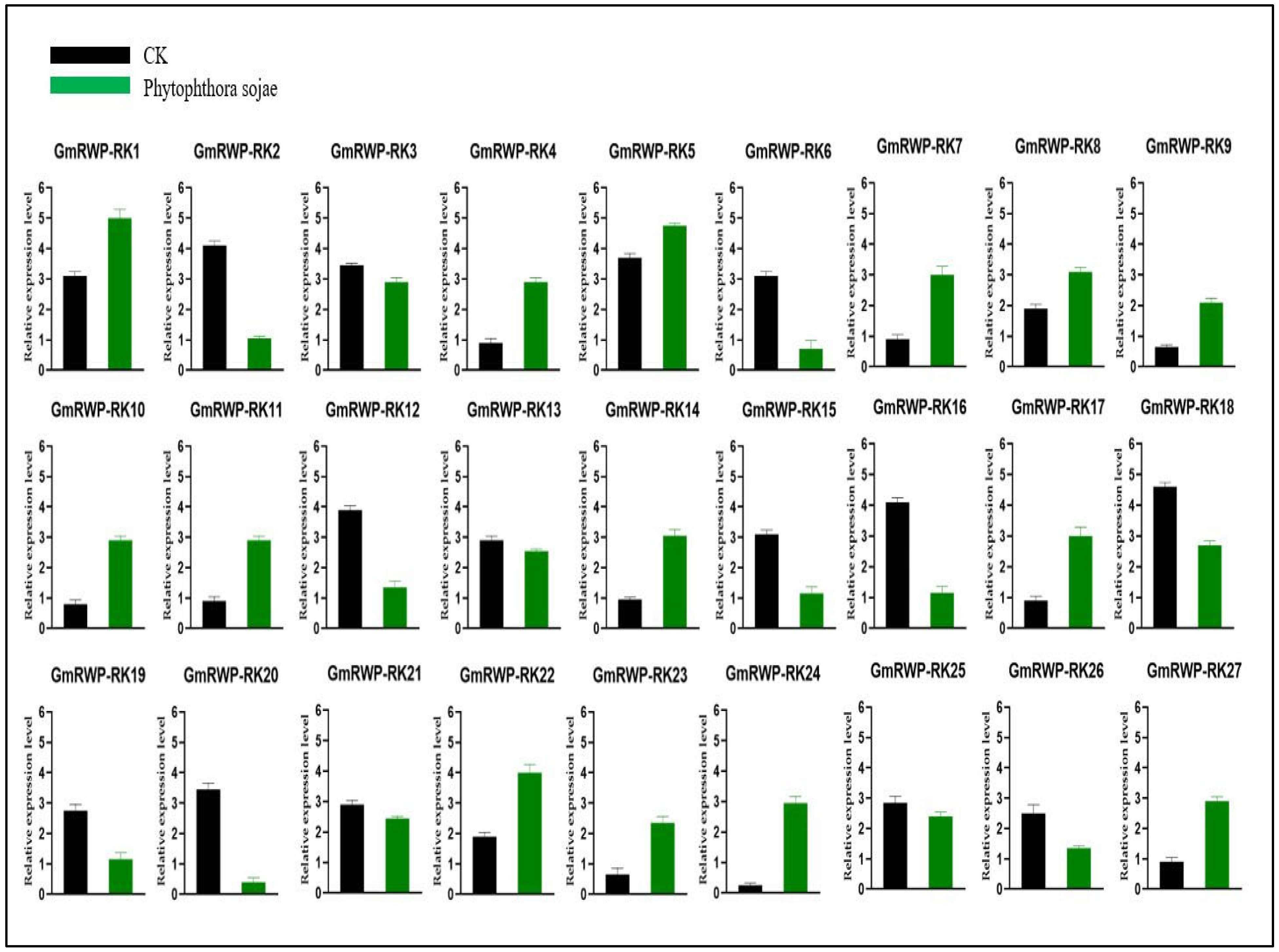
2.7. Expression Analysis of RWP-RK Genes Family under Phytophthora Sojae
2.8. Expression Analysis of RWP-RK Gene Family under Nitrogen Application
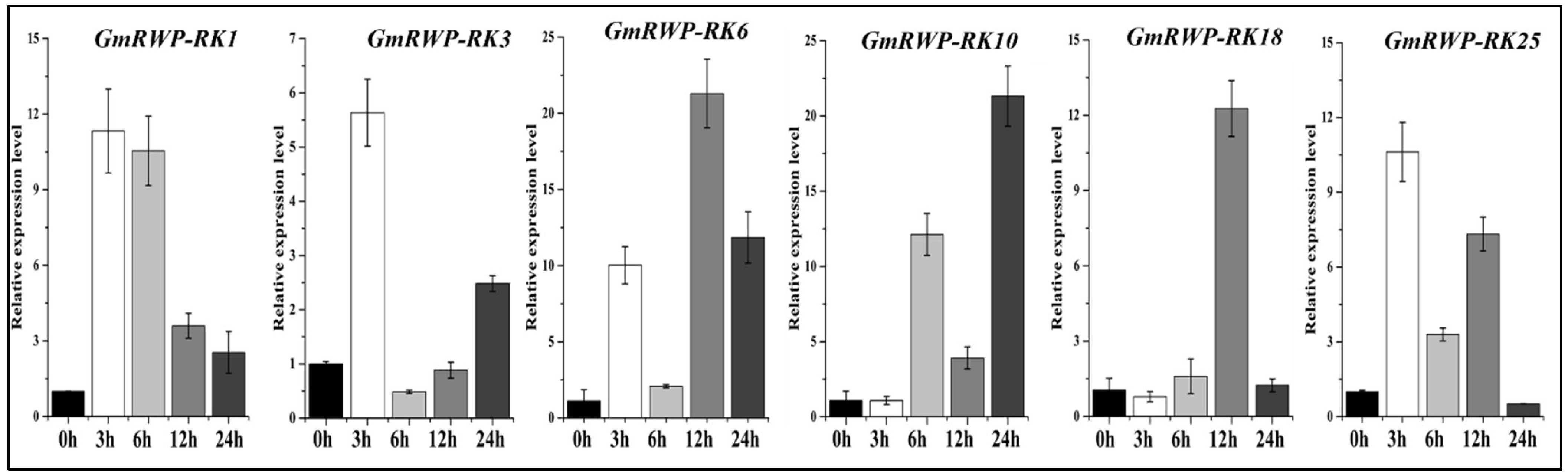
2.9. Expression Analysis of RWP-RK Genes Family under Heat Stress
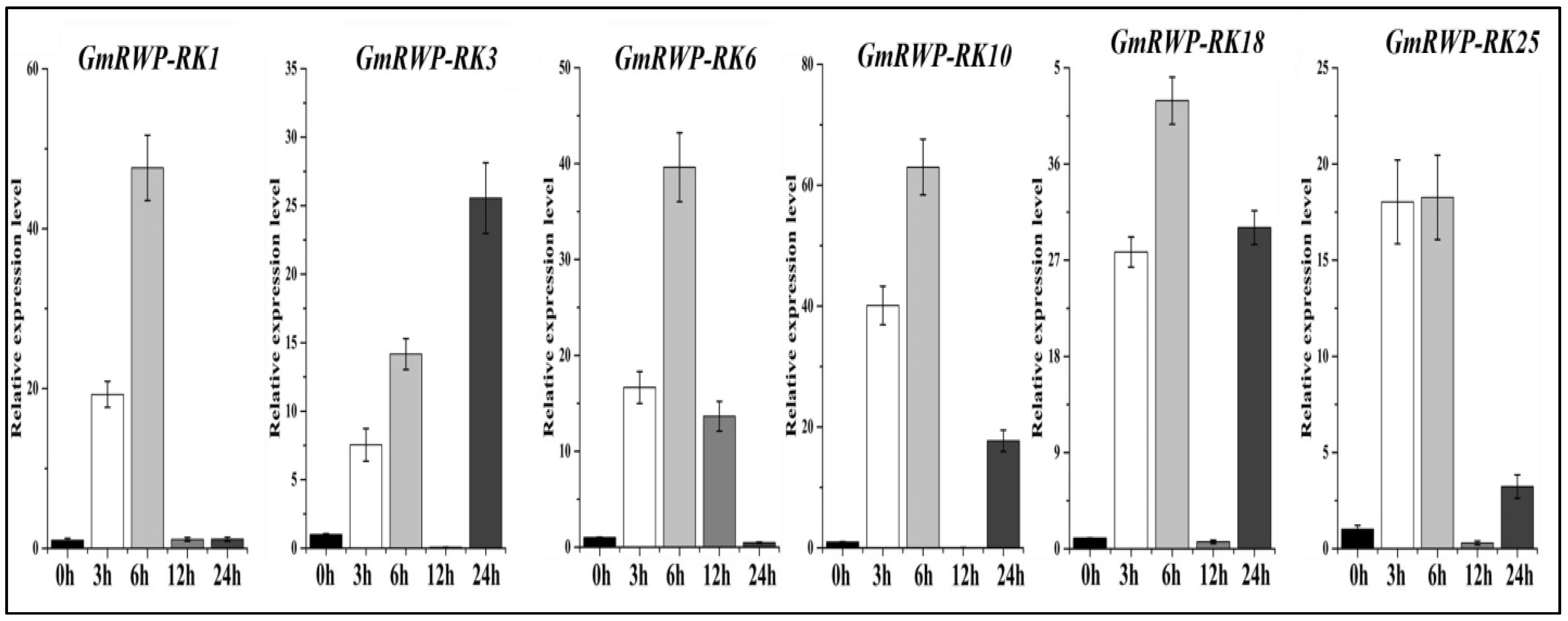
2.10. Expression Analysis of RWP-RK Genes Family under Salt Stress
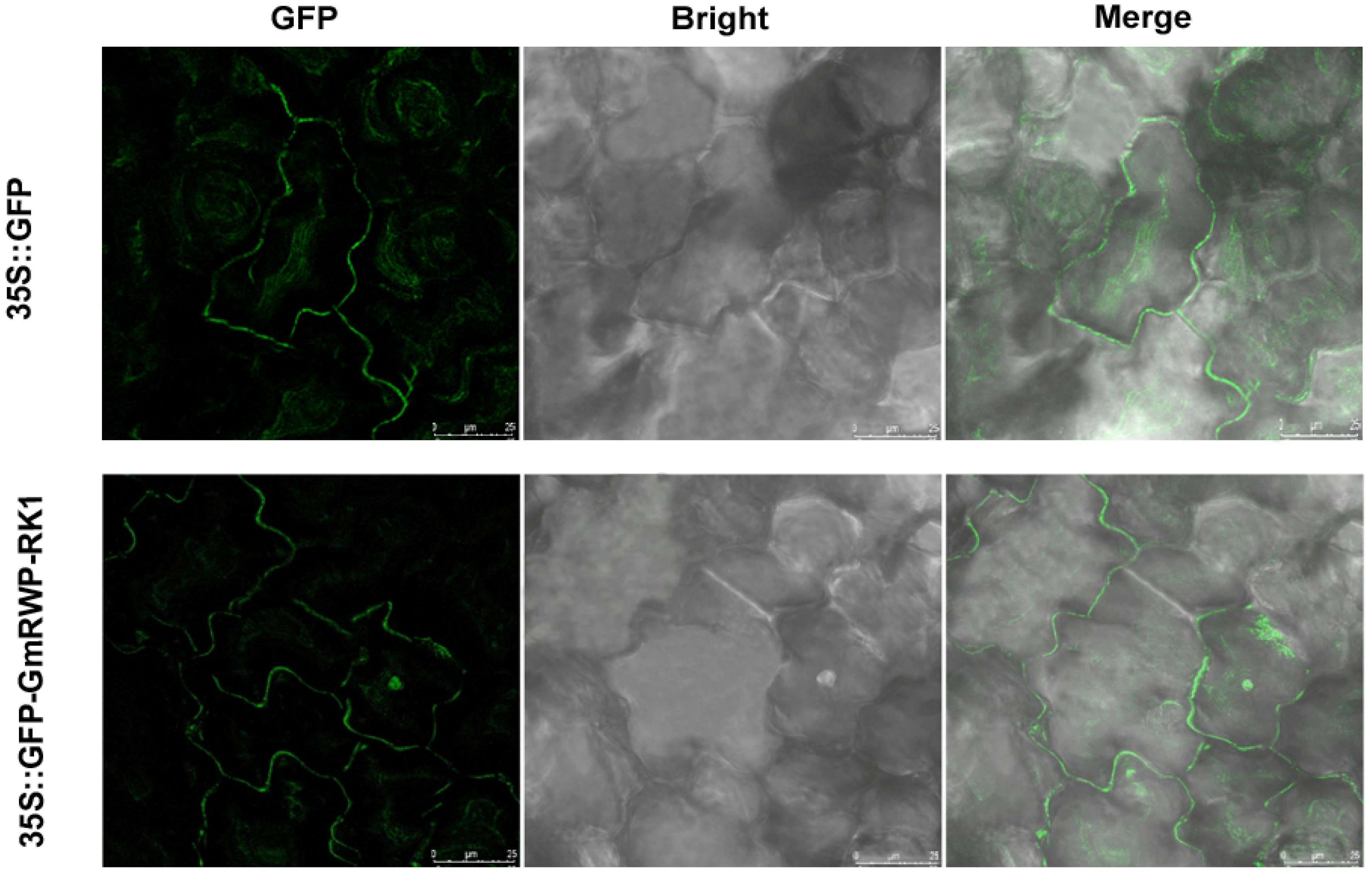
2.11. GmRWP-RK1 Localizations in the Nucleus
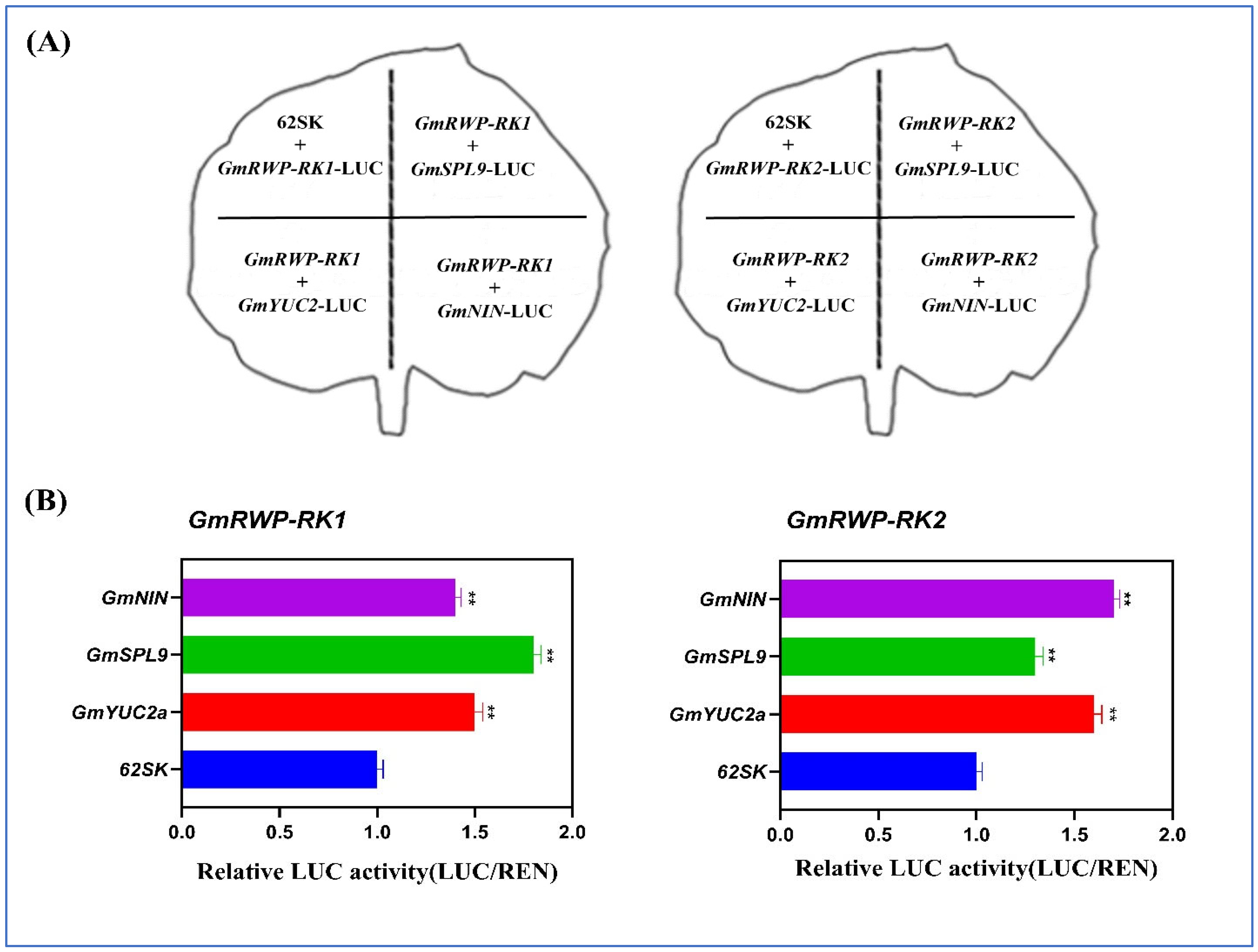
2.12. GmRWP-RK1 and GmRWP-RK2 Promote the Transcription of Nodule Specific Genes
3. Discussion
3.1. Distribution and Features of RWP-RKs in Plants
3.2. RWP-RKs Regulate Soybean Growth and Stress Responses
3.3. The GmRWP-RK Gene Family Binds and Induces the Transcription of Nodule Specific Genes
4. Conclusions
5. Materials and Methods
5.1. Phylogenetic and Physicochemical Characteristics of the Gm-RWP-RK Proteins
5.2. Conserved Protein Motif and Domain Analysis
5.3. Promoter Sequence Analysis
5.4. Plant Materials and Stress Conditions
5.5. Quantitative Real-Time PCR Analysis
5.6. ORF Cloning and Subcellular Localization of GmRWP-RK
5.7. Dual-Luciferase Assay
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gojon, A. Nitrogen nutrition in plants: Rapid progress and new challenges. J. Exp. Bot. 2017, 68, 2457–2462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldroyd, G.E. Speak, friend, and enter: Signalling systems that promote beneficial symbiotic associations in plants. Nat. Rev. Microbiol. 2013, 11, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Good, A. Toward nitrogen-fixing plants. Science 2018, 359, 869–870. [Google Scholar] [CrossRef]
- Chardin, C.; Girin, T.; Roudier, F.; Meyer, C.; Krapp, A. The plant RWP-RK transcription factors: Key regulators of nitrogen responses and of gametophyte development. J. Exp. Bot. 2014, 65, 5577–5587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konishi, M.; Yanagisawa, S. Emergence of a new step towards understanding the molecular mechanisms underlying nitrate-regulated gene expression. J. Exp. Bot. 2014, 65, 5589–5600. [Google Scholar] [CrossRef] [Green Version]
- Konishi, M.; Okitsu, T.; Yanagisawa, S. Nitrate-responsive NIN-like protein transcription factors perform unique and redundant roles in Arabidopsis. J. Exp. Bot. 2021, 72, 5735–5750. [Google Scholar] [CrossRef]
- Yu, L.H.; Wu, J.; Tang, H.; Yuan, Y.; Wang, S.M.; Wang, Y.P.; Zhu, Q.S.; Li, S.G.; Xiang, C.B. Overexpression of Arabidopsis NLP7 improves plant growth under both nitrogen-limiting and sufficient conditions by enhancing nitrogen and carbon assimilation. Sci. Rep. 2016, 6, 27795. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Goodenough, U.W. Gametogenesis in the Chlamydomonas reinhardtii minus mating type is controlled by two genes, MID and MTD1. Genetics 2007, 176, 913–925. [Google Scholar] [CrossRef] [Green Version]
- Konishi, M.; Yanagisawa, S. An NLP-binding site in the 3′ flanking region of the nitrate reductase gene confers nitrate-inducible expression in Arabidopsis thaliana (L.) Heynh. Soil Sci. Plant Nutr. 2013, 59, 612–620. [Google Scholar] [CrossRef]
- Castaings, L.; Camargo, A.; Pocholle, D.; Gaudon, V.; Texier, Y.; Boutet-Mercey, S.; Taconnat, L.; Renou, J.P.; Daniel-Vedele, F.; Fernandez, E.; et al. The nodule inception-like protein 7 modulates nitrate sensing and metabolism in Arabidopsis. Plant J. 2009, 57, 426–435. [Google Scholar] [CrossRef]
- Schauser, L.; Wieloch, W.; Stougaard, J. Evolution of NIN-like proteins in Arabidopsis, rice, and Lotus japonicus. J. Mol. Evol. 2005, 60, 229–237. [Google Scholar] [CrossRef]
- Koi, S.; Hisanaga, T.; Sato, K.; Shimamura, M.; Yamato, K.T.; Ishizaki, K.; Kohchi, T.; Nakajima, K. An evolutionarily conserved plant RKD factor controls germ cell differentiation. Curr. Biol. 2016, 26, 1775–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, M.; Zhang, Z.; Xuan, M.; Feng, H.; Ye, W.; Zheng, X.; Wang, Y. Conserved subgroups of the plant-specific RWP-RK transcription factor family are present in oomycete pathogens. Front. Microbiol. 2020, 11, 1724. [Google Scholar] [CrossRef]
- Kumar, A.; Batra, R.; Gahlaut, V.; Gautam, T.; Kumar, S.; Sharma, M.; Tyagi, S.; Singh, K.P.; Balyan, H.S.; Pandey, R.; et al. Genome-wide identification and characterization of gene family for RWP-RK transcription factors in wheat (Triticum aestivum L.). PLoS ONE 2018, 13, e0208409. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Liu, H.; Huang, W.; Yi, L.; Qin, E.; Yang, T.; Wang, J.; Qin, R. Genome-Wide identification, characterization, and regulation of RWP-RK gene family in the nitrogen-fixing clade. Plants 2020, 9, 1178. [Google Scholar] [CrossRef] [PubMed]
- Konishi, M.; Yanagisawa, S. Arabidopsis NIN-like transcription factors have a central role in nitrate signalling. Nat. Commun. 2013, 4, 1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, S.; Delcourt, C.; Zhang, S.; Arima, H.; Heeley, E.; Zheng, D.; Al-Shahi Salman, R.; Stapf, C.; Tzourio, C.; Robinson, T.; et al. Determinants and Prognostic Significance of Hematoma Sedimentation Levels in Acute Intracerebral Hemorrhage. Cerebrovasc. Dis. 2016, 41, 80–86. [Google Scholar] [CrossRef]
- Mu, X.; Luo, J. Evolutionary analyses of NIN-like proteins in plants and their roles in nitrate signaling. Cell. Mol. Life Sci. 2019, 76, 3753–3764. [Google Scholar] [CrossRef]
- Ahmad, S.; Zhang, J.; Wang, H.; Zhu, H.; Dong, Q.; Zong, S.; Wang, T.; Chen, Y.; Ge, L.Q. The phosphoserine phosphatase alters the free amino acid compositions and fecundity in Cyrtorhinus lividipennis Reuter. Int. J. Mol. Sci. 2022, 23, 15283. [Google Scholar] [CrossRef]
- Suzuki, W.; Konishi, M.; Yanagisawa, S. The evolutionary events necessary for the emergence of symbiotic nitrogen fixation in legumes may involve a loss of nitrate responsiveness of the NIN transcription factor. Plant Signal. Behav. 2013, 8, e25975. [Google Scholar] [CrossRef]
- Alfatih, A.; Zhang, J.; Song, Y.; Jan, S.U.; Zhang, Z.-S.; Xia, J.-Q.; Zhang, Z.-Y.; Nazish, T.; Wu, J.; Zhao, P.-X. Nitrate-dependent salt tolerance mediated by OsNLP4-OsMADS27 module. bioRxiv 2022. [Google Scholar] [CrossRef]
- Driouich, A.; Follet-Gueye, M.-L.; Vicré-Gibouin, M.; Hawes, M. Root border cells and secretions as critical elements in plant host defense. Curr. Opin. Plant Biol. 2013, 16, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Watson, B.S.; Bedair, M.F.; Urbanczyk-Wochniak, E.; Huhman, D.V.; Yang, D.S.; Allen, S.N.; Li, W.; Tang, Y.; Sumner, L.W. Integrated metabolomics and transcriptomics reveal enhanced specialized metabolism in Medicago truncatula root border cells. Plant Physiol. 2015, 167, 1699–1716. [Google Scholar] [PubMed] [Green Version]
- Tubiello, F.N.; Salvatore, M.; Ferrara, A.F.; House, J.; Federici, S.; Rossi, S.; Biancalani, R.; Condor Golec, R.D.; Jacobs, H.; Flammini, A.; et al. The contribution of agriculture, forestry and other land use activities to global warming, 1990–2012. Glob. Chang. Biol. 2015, 21, 2655–2660. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Yang, S.-S.; Liang, Z.; Feng, B.-R.; Liu, L.; Huang, Y.-B.; Tang, Y.-X. Genome-wide analysis of the MYB transcription factor superfamily in soybean. BMC Plant Biol. 2012, 12, 106. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.-M.; Wang, X.; Wang, Y.-W.; Hu, R.-B.; Zhang, X.-M.; Chen, J.-X.; Fu, Y.-F. Genome-wide expression analysis of soybean MADS genes showing potential function in the seed development. PLoS ONE 2013, 8, e62288. [Google Scholar] [CrossRef] [Green Version]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Mochida, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.-S.P. Genome-wide survey and expression analysis of the plant-specific NAC transcription factor family in soybean during development and dehydration stress. DNA Res. 2011, 18, 263–276. [Google Scholar] [CrossRef] [Green Version]
- Bencke-Malato, M.; Cabreira, C.; Wiebke-Strohm, B.; Bücker-Neto, L.; Mancini, E.; Osorio, M.B.; Homrich, M.S.; Turchetto-Zolet, A.C.; De Carvalho, M.C.; Stolf, R.; et al. Genome-wide annotation of the soybean WRKY family and functional characterization of genes involved in response to Phakopsora pachyrhiziinfection. BMC Plant Biol. 2014, 14, 236. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, L.; ShangGuan, G.; Jia, C.; Deng, S.; Noman, M.; Liu, Y.; Guo, Y.; Han, L.; Zhang, X.; et al. Genome-wide identification and expression analysis of bZIP gene family in Carthamus tinctorius L. Sci. Rep. 2020, 10, 15521. [Google Scholar] [CrossRef]
- Chen, X.; Chen, Z.; Zhao, H.; Zhao, Y.; Cheng, B.; Xiang, Y. Genome-wide analysis of soybean HD-Zip gene family and expression profiling under salinity and drought treatments. PLoS ONE 2014, 9, e87156. [Google Scholar]
- Van Ha, C.; Le, D.T.; Nishiyama, R.; Watanabe, Y.; Sulieman, S.; Tran, U.T.; Mochida, K.; Van Dong, N.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The auxin response factor transcription factor family in soybean: Genome-wide identification and expression analyses during development and water stress. DNA Res. 2013, 20, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Yuan, D.; Liu, T.; Xing, M.; Xu, W.; Zhang, H.; Jin, H.; Cai, C.; Li, S. Characterization and Comparative Analysis of RWP-RK Proteins from Arachis duranensis, Arachis ipaensis, and Arachis hypogaea. Int. J. Genom. 2020, 2020, 2568640. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Xie, C.; Wang, J.; Cao, Z.; Zhang, H.; Chen, P.; Li, Y. Genome wide identification, characterization and expression analysis of HD-ZIP gene family in Cucumis sativus L. under biotic and various abiotic stresses. Int. J. Biol. Macromol. 2020, 158, 502–520. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Liu, P.; Wang, D.; Jin, Z.; Uzair, U.; Yadav, V.; Mujtaba, M.; Chen, P.; Li, Y. Genome-wide characterisation and expression analysis of cellulose synthase genes superfamily under various environmental stresses in Cucumis sativus L. N. Zeal. J. Crop Hort. Sci. 2021, 49, 127–150. [Google Scholar] [CrossRef]
- Crespi, M.; Frugier, F. De novo organ formation from differentiated cells: Root nodule organogenesis. Sci. Signal. 2008, 1, re11. [Google Scholar] [CrossRef]
- Sharif, R.; Su, L.; Chen, X.; Qi, X. Hormonal interactions underlying parthenocarpic fruit formation in horticultural crops. Hortic. Res. 2022, 9, uhab024. [Google Scholar] [CrossRef]
- Ahmad, N.; Hou, L.; Ma, J.; Zhou, X.; Xia, H.; Wang, M.; Leal-Bertioli, S.; Zhao, S.; Tian, R.; Pan, J. Bulk RNA-Seq Analysis Reveals Differentially Expressed Genes Associated with Lateral Branch Angle in Peanut. Genes 2022, 13, 841. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.S.; Li, X.; Luo, Z.; Mysore, K.S.; Wen, J.; Xie, F. NIN interacts with NLPs to mediate nitrate inhibition of nodulation in Medicago truncatula. Nat. Plants 2018, 4, 942–952. [Google Scholar] [CrossRef]
- Movahedi, S.; Van Bel, M.; Heyndrickx, K.S.; Vandepoele, K. Comparative co-expression analysis in plant biology. Plant Cell Environ. 2012, 35, 1787–1798. [Google Scholar] [CrossRef]
- Koszegi, D.; Johnston, A.J.; Rutten, T.; Czihal, A.; Altschmied, L.; Kumlehn, J.; Wüst, S.E.; Kirioukhova, O.; Gheyselinck, J.; Grossniklaus, U.; et al. Members of the RKD transcription factor family induce an egg cell-like gene expression program. Plant J. 2011, 67, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, F.; Rizzo, P.; Rutten, T.; Altschmied, L.; Bäumlein, H. RWP-RK domain-containing transcription factors control cell differentiation during female gametophyte development in Arabidopsis. New Phytol. 2017, 213, 1909–1924. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.F.; Chen, W.H.; Shen, Y.H.; Hsu, W.H.; Mao, W.T.; Yang, C.H. Multifunctional evolution of B and AGL6 MADS box genes in orchids. Nat. Commun. 2021, 12, 902. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Hong, W.J.; Kim, Y.J.; Jung, K.H. Transcriptome Analysis of Triple Mutant for OsMADS62, OsMADS63, and OsMADS68 Reveals the Downstream Regulatory Mechanism for Pollen Germination in Rice (Oryza sativa). Int. J. Mol. Sci. 2021, 23, 239. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, Y.; Xu, L.; Nie, S.; Chen, Y.; Liang, D.; Sun, X.; Karanja, B.K.; Luo, X.; Liu, L. Genome-Wide Characterization of the MADS-Box Gene Family in Radish (Raphanus sativus L.) and Assessment of Its Roles in Flowering and Floral Organogenesis. Front. Plant Sci. 2016, 7, 1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Wei, L.; Wang, W.; Qi, W.; Cao, Z.; Li, H.; Bao, M.; He, Y. Identification, characterization and functional analysis of AGAMOUS subfamily genes associated with floral organs and seed development in Marigold (Tagetes erecta). BMC Plant Biol. 2020, 20, 439. [Google Scholar] [CrossRef]
- Kanno, A.; Saeki, H.; Kameya, T.; Saedler, H.; Theissen, G. Heterotopic expression of class B floral homeotic genes supports a modified ABC model for tulip (Tulipa gesneriana). Plant Mol. Biol. 2003, 52, 831–841. [Google Scholar] [CrossRef]
- Litt, A.; Kramer, E.M. The ABC model and the diversification of floral organ identity. Semin. Cell Dev. Biol. 2010, 21, 129–137. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Q.; Yang, S.; Lin, S.; Bao, M.; Bendahmane, M.; Wu, Q.; Wang, C.; Fu, X. Identification and Characterization of the MADS-Box Genes and Their Contribution to Flower Organ in Carnation (Dianthus caryophyllus L.). Genes 2018, 9, 193. [Google Scholar] [CrossRef] [Green Version]
- Li, B.J.; Zheng, B.Q.; Wang, J.Y.; Tsai, W.C.; Lu, H.C.; Zou, L.H.; Wan, X.; Zhang, D.Y.; Qiao, H.J.; Liu, Z.J.; et al. New insight into the molecular mechanism of colour differentiation among floral segments in orchids. Commun. Biol. 2020, 3, 89. [Google Scholar] [CrossRef] [Green Version]
- Yun, J.; Sun, Z.; Jiang, Q.; Wang, Y.; Wang, C.; Luo, Y.; Zhang, F.; Li, X. The miR156b-GmSPL9d module modulates nodulation by targeting multiple core nodulation genes in soybean. New Phytol. 2022, 233, 1881–1899. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, W.; Zuo, Y.; Zhu, L.; Hastwell, A.H.; Chen, L.; Tian, Y.; Su, C.; Ferguson, B.J.; Li, X. GmYUC2a mediates auxin biosynthesis during root development and nodulation in soybean. J. Exp. Bot. 2019, 70, 3165–3176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.; Zhu, H.W.; Chen, Y.; Xi, C.Y.; Shah, A.Z.; Ge, L.Q. Comprehensive Bioinformatics and Expression Analysis of the TLP Gene Family Revealed Its Role in Regulating the Response of Oryza sativa to Nilaparvata lugens, Laodelphax striatellus, and Jinggangmycin. Agronomy 2022, 12, 1297. [Google Scholar] [CrossRef]
- Ahmad, S.; Chen, Y.; Shah, A.Z.; Wang, H.; Xi, C.; Zhu, H.; Ge, L. The Homeodomain-Leucine Zipper Genes Family Regulates the Jinggangmycin Mediated Immune Response of Oryza sativa to Nilaparvata lugens, and Laodelphax striatellus. Bioengineering 2022, 9, 398. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Ahmad, S.; Jiang, L.; Zheng, S.; Chen, Y.; Zhang, J.Y.; Stanley, D.; Miao, H.; Ge, L.Q. Silencing of a putative alanine aminotransferase (ALT) gene influences free amino acid composition in hemolymph and fecundity of the predatory bug, Cyrtorhinus lividipennis Reuter. Arch. Insect Biochem. Physiol. 2021, 108, e21836. [Google Scholar] [CrossRef]
- Ahmad, S.; Chen, Y.; Zhang, J.; Stanley, D.; Song, Q.; Ge, L. Octopamine signaling is involved in the female postmating state in Nilaparvata lugens Stål (Hemiptera: Delphacidae). Arch. Insect Biochem. Physiol. 2021, 107, e21825. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amin, N.; Ahmad, N.; Khalifa, M.A.S.; Du, Y.; Mandozai, A.; Khattak, A.N.; Piwu, W. Identification and Molecular Characterization of RWP-RK Transcription Factors in Soybean. Genes 2023, 14, 369. https://doi.org/10.3390/genes14020369
Amin N, Ahmad N, Khalifa MAS, Du Y, Mandozai A, Khattak AN, Piwu W. Identification and Molecular Characterization of RWP-RK Transcription Factors in Soybean. Genes. 2023; 14(2):369. https://doi.org/10.3390/genes14020369
Chicago/Turabian StyleAmin, Nooral, Naveed Ahmad, Mohamed A. S. Khalifa, Yeyao Du, Ajmal Mandozai, Aimal Nawaz Khattak, and Wang Piwu. 2023. "Identification and Molecular Characterization of RWP-RK Transcription Factors in Soybean" Genes 14, no. 2: 369. https://doi.org/10.3390/genes14020369
APA StyleAmin, N., Ahmad, N., Khalifa, M. A. S., Du, Y., Mandozai, A., Khattak, A. N., & Piwu, W. (2023). Identification and Molecular Characterization of RWP-RK Transcription Factors in Soybean. Genes, 14(2), 369. https://doi.org/10.3390/genes14020369