
Molecular Evolution of SARS-CoV-2 during the COVID-19 Pandemic



Abstract
:1. Introduction
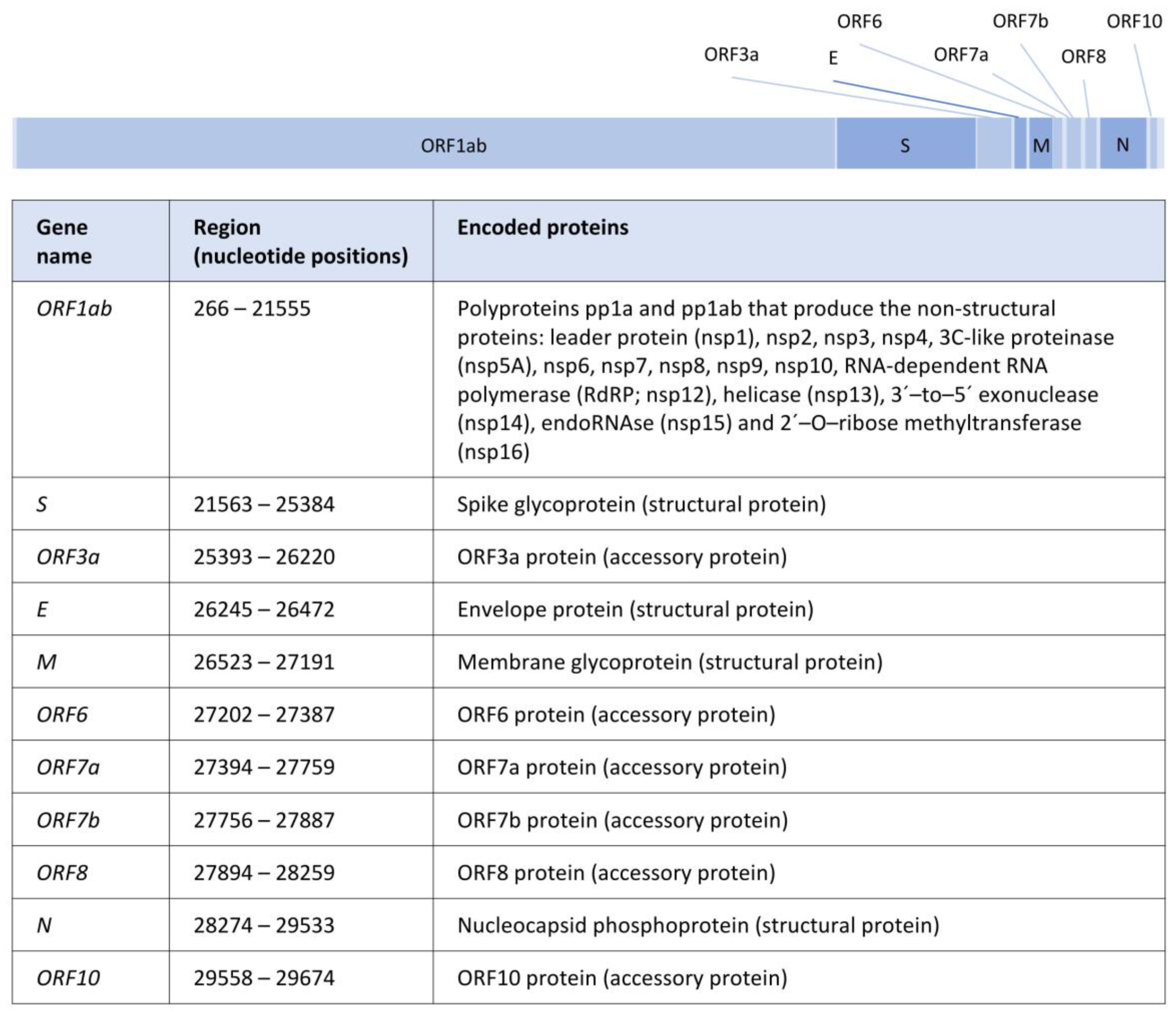
2. The SARS-CoV-2 Genome and Proteins
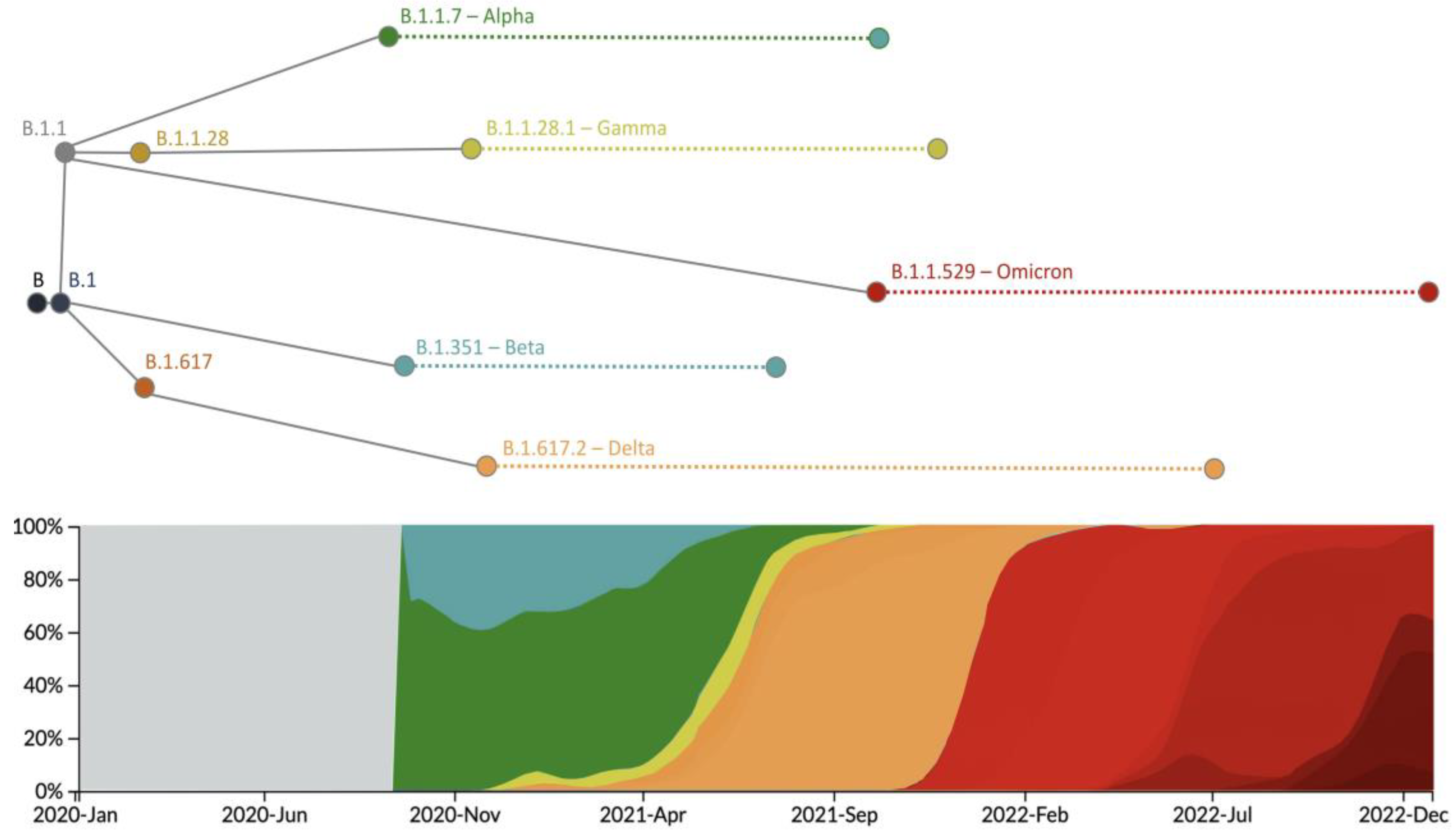
3. The Nomenclature and Evolutionary History of SARS-CoV-2 Lineages
4. Evolutionary Mechanisms of SARS-CoV-2
4.1. The Mutation Process in SARS-CoV-2
4.2. Recombination in the SARS-CoV-2
4.3. The Rate of Molecular Evolution of the SARS-CoV-2
4.4. Molecular Adaptation in the SARS-CoV-2 Genome
Molecular Adaptation Induced by Therapies and Immune Systems
5. Conclusions and Future Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chan-Yeung, M.; Xu, R. SARS: Epidemiology. Respirology 2003, 8, S9–S14. [Google Scholar] [CrossRef] [PubMed]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.M.; Lau, E.H.Y.; Wong, J.Y.; et al. Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus–Infected Pneumonia. N. Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Bonotti, M.; Zech, S.T. The Human, Economic, Social, and Political Costs of COVID-19. In Recovering Civility during COVID-19; Springer: Berlin/Heidelberg, Germany, 2021; pp. 1–36. [Google Scholar] [CrossRef]
- Pan, T.; Chen, R.; He, X.; Yuan, Y.; Deng, X.; Li, R.; Yan, H.; Yan, S.; Liu, J.; Zhang, Y.; et al. Infection of wild-type mice by SARS-CoV-2 B. 1.351 variant indicates a possible novel cross-species transmission route. Signal Transduct. Target. Ther. 2021, 6, 420. [Google Scholar] [CrossRef] [PubMed]
- de Vries, R.D.; Rockx, B.; Haagmans, B.L.; Herfst, S.; Koopmans, M.P.; de Swart, R.L. Animal models of SARS-CoV-2 transmission. Curr. Opin. Virol. 2021, 50, 8–16. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. The Species Severe Acute Respiratory Syndrome-Related Coronavirus: Classifying 2019-NCoV and Naming It SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef]
- Bai, C.; Zhong, Q.; Gao, G.F. Overview of SARS-CoV-2 Genome-Encoded Proteins. Sci. China Life Sci. 2022, 65, 280–294. [Google Scholar] [CrossRef]
- Li, J.; Lai, S.; Gao, G.F.; Shi, W. The Emergence, Genomic Diversity and Global Spread of SARS-CoV-2. Nature 2021, 600, 408–418. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Peacock, S.J.; et al. SARS-CoV-2 Variants, Spike Mutations and Immune Escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Singh, D.; Yi, S.V. On the Origin and Evolution of SARS-CoV-2. Exp. Mol. Med. 2021, 53, 537–547. [Google Scholar] [CrossRef]
- Tao, K.; Tzou, P.L.; Nouhin, J.; Bonilla, H.; Jagannathan, P.; Shafer, R.W. SARS-CoV-2 Antiviral Therapy. Clin. Microbiol. Rev. 2021, 34, e00109-21. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, L.; Poullikkas, T.; Eisenhower, S.; Monsanto, C.; Bakku, R.K.; Chen, M.-H.; Kalra, R.S. Viroinformatics-Based Analysis of SARS-CoV-2 Core Proteins for Potential Therapeutic Targets. Antibodies 2021, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Day, T.; Gandon, S.; Lion, S.; Otto, S.P. On the Evolutionary Epidemiology of SARS-CoV-2. Curr. Biol. 2020, 30, R849–R857. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, F.K. The Proteins of Severe Acute Respiratory Syndrome Coronavirus-2 (SARS CoV-2 or n-COV19), the Cause of COVID-19. Protein J. 2020, 39, 198–216. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, C.; Xu, X.; Xu, W.; Liu, S. Structural and Functional Properties of SARS-CoV-2 Spike Protein: Potential Antivirus Drug Development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef]
- Abavisani, M.; Rahimian, K.; Mahdavi, B.; Tokhanbigli, S.; Mollapour Siasakht, M.; Farhadi, A.; Kodori, M.; Mahmanzar, M.; Meshkat, Z. Mutations in SARS-CoV-2 Structural Proteins: A Global Analysis. Virol. J. 2022, 19, 220. [Google Scholar] [CrossRef]
- Savastano, A.; Ibáñez de Opakua, A.; Rankovic, M.; Zweckstetter, M. Nucleocapsid Protein of SARS-CoV-2 Phase Separates into RNA-Rich Polymerase-Containing Condensates. Nat. Commun. 2020, 11, 6041. [Google Scholar] [CrossRef]
- Candido, K.L.; Eich, C.R.; de Fariña, L.O.; Kadowaki, M.K.; da Conceição Silva, J.L.; Maller, A.; Simão, R.C.G. Spike Protein of SARS-CoV-2 Variants: A Brief Review and Practical Implications. Braz. J. Microbiol. 2022, 53, 1133–1157. [Google Scholar] [CrossRef]
- Zhan, X.Y.; Zhang, Y.; Zhou, X.; Huang, K.; Qian, Y.; Leng, Y.; Yan, L.; Huang, B.; He, Y. Molecular Evolution of SARS-CoV-2 Structural Genes: Evidence of Positive Selection in the Spike Glycoprotein. bioRxiv 2020. [Google Scholar] [CrossRef]
- Mohammad, T.; Choudhury, A.; Habib, I.; Asrani, P.; Mathur, Y.; Umair, M.; Anjum, F.; Shafie, A.; Yadav, D.K.; Hassan, M.I. Genomic Variations in the Structural Proteins of SARS-CoV-2 and Their Deleterious Impact on Pathogenesis: A Comparative Genomics Approach. Front. Cell. Infect. Microbiol. 2021, 11, 951. [Google Scholar] [CrossRef]
- Masters, P.S. Coronavirus Genomic RNA Packaging. Virology 2019, 537, 198–207. [Google Scholar] [CrossRef]
- Bai, Z.; Cao, Y.; Liu, W.; Li, J. The SARS-CoV-2 Nucleocapsid Protein and Its Role in Viral Structure, Biological Functions, and a Potential Target for Drug or Vaccine Mitigation. Viruses 2021, 13, 1115. [Google Scholar] [CrossRef] [PubMed]
- Hillen, H.S. Structure and Function of SARS-CoV-2 Polymerase. Curr. Opin. Virol. 2021, 48, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Holmes, E.C.; O’Toole, Á.; Hill, V.; McCrone, J.T.; Ruis, C.; du Plessis, L.; Pybus, O.G. A Dynamic Nomenclature Proposal for SARS-CoV-2 Lineages to Assist Genomic Epidemiology. Nat. Microbiol. 2020, 5, 1403–1407. [Google Scholar] [CrossRef] [PubMed]
- Alm, E.; Broberg, E.K.; Connor, T.; Hodcroft, E.B.; Komissarov, A.B.; Maurer-Stroh, S.; Melidou, A.; Neher, R.A.; O’Toole, Á.; Pereyaslov, D.; et al. Geographical and Temporal Distribution of SARS-CoV-2 Clades in the WHO European Region, January to June 2020. Eurosurveillance 2020, 25, 2001410. [Google Scholar] [CrossRef]
- Hadfield, J.; Megill, C.; Bell, S.M.; Huddleston, J.; Potter, B.; Callender, C.; Sagulenko, P.; Bedford, T.; Neher, R.A. Nextstrain: Real-Time Tracking of Pathogen Evolution. Bioinformatics 2018, 34, 4121–4123. [Google Scholar] [CrossRef]
- Elbe, S.; Buckland-Merrett, G. Data, Disease and Diplomacy: GISAID’s Innovative Contribution to Global Health. Glob. Chall. 2017, 1, 33–46. [Google Scholar] [CrossRef]
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.B.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; et al. Estimated Transmissibility and Impact of SARS-CoV-2 Lineage B.1.1.7 in England. Science 2021, 372, eabg3055. [Google Scholar] [CrossRef]
- Funk, T.; Pharris, A.; Spiteri, G.; Bundle, N.; Melidou, A.; Carr, M.; Gonzalez, G.; Garcia-Leon, A.; Crispie, F.; O’Connor, L.; et al. Characteristics of SARS-CoV-2 Variants of Concern B.1.1.7, B.1.351 or P.1: Data from Seven EU/EEA Countries, Weeks 38/2020 to 10/2021. Eurosurveillance 2021, 26, 2100348. [Google Scholar] [CrossRef]
- Choi, J.Y.; Smith, D.M. SARS-CoV-2 Variants of Concern. Yonsei Med. J. 2021, 62, 961–968. [Google Scholar] [CrossRef]
- Attwood, S.W.; Hill, S.C.; Aanensen, D.M.; Connor, T.R.; Pybus, O.G. Phylogenetic and Phylodynamic Approaches to Understanding and Combating the Early SARS-CoV-2 Pandemic. Nat. Rev. Genet. 2022, 23, 547–562. [Google Scholar] [CrossRef] [PubMed]
- Naveca, F.G.; Nascimento, V.; de Souza, V.C.; de Lima Corado, A.; Nascimento, F.; Silva, G.; Costa, Á.; Duarte, D.; Pessoa, K.; Mejía, M.; et al. COVID-19 in Amazonas, Brazil, Was Driven by the Persistence of Endemic Lineages and P.1 Emergence. Nat. Med. 2021, 27, 1230–1238. [Google Scholar] [CrossRef] [PubMed]
- Sabino, E.C.; Buss, L.F.; Carvalho, M.P.S.; Prete, C.A.; Crispim, M.A.E.; Fraiji, N.A.; Pereira, R.H.M.; Parag, K.V.; Peixoto, P.; Kraemer, M.U.G.; et al. Resurgence of COVID-19 in Manaus, Brazil, despite High Seroprevalence. Lancet 2021, 397, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.W.X.; Chiew, C.J.; Ang, L.W.; Mak, T.M.; Cui, L.; Toh, M.P.H.; Lim, Y.D.; Lee, P.H.; Lee, T.H.; Chia, P.Y.; et al. Clinical and Virological Features of SARS-CoV-2 Variants of Concern: A Retrospective Cohort Study Comparing B.1.1.7 (Alpha), B.1.315 (Beta), and B.1.617.2 (Delta). Clin. Infect. Dis. 2021, 75, ciab721. [Google Scholar] [CrossRef]
- Kannan, S.; Shaik Syed Ali, P.; Sheeza, A. Omicron (B.1.1.529)–Variant of Concern–Molecular Profile and Epidemiology: A Mini Review. Nucleic Acids Res. 2021, 49, W431–W437. [Google Scholar] [CrossRef]
- Shrestha, L.B.; Foster, C.; Rawlinson, W.; Tedla, N.; Bull, R.A. Evolution of the SARS-CoV-2 omicron variants BA. 1 to BA. 5: Implications for immune escape and transmission. Rev. Med. Virol. 2022, 32, e2381. [Google Scholar] [CrossRef]
- Lauring, A.S.; Tenforde, M.W.; Chappell, J.D.; Gaglani, M.; Ginde, A.A.; McNeal, T.; Ghamande, S.; Douin, D.J.; Talbot, H.K.; Casey, J.D.; et al. Clinical severity of, and effectiveness of mRNA vaccines against, covid-19 from omicron, Delta, and Alpha SARS-CoV-2 variants in the United States: Prospective observational study. BMJ 2022, 376, e069761. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhi, H.; Teng, Y. The outbreak of SARS-CoV-2 Omicron lineages, immune escape, and vaccine effectivity. J. Med. Virol. 2023, 95, e28138. [Google Scholar] [CrossRef]
- Rajpal, V.R.; Sharma, S.; Kumar, A.; Chand, S.; Joshi, L.; Chandra, A.; Babbar, S.; Goel, S.; Raina, S.N.; Shiran, B. ‘Is omicron mild’? Testing this narrative with the mutational landscape of its three lineages and response to existing vaccines and therapeutic antibodies. J. Med. Virol. 2022, 94, 3521–3539. [Google Scholar] [CrossRef]
- Tegally, H.; Moir, M.; Everatt, J.; Giovanetti, M.; Scheepers, C.; Wilkinson, E.; Subramoney, K.; Makatini, Z.; Moyo, S.; Amoako, D.G.; et al. Emergence of SARS-CoV-2 omicron lineages BA. 4 and BA. 5 in South Africa. Nat. Med. 2022, 28, 1785–1790. [Google Scholar] [CrossRef]
- Benvenuto, D.; Angeletti, S.; Giovanetti, M.; Bianchi, M.; Pascarella, S.; Cauda, R.; Ciccozzi, M.; Cassone, A. Evolutionary Analysis of SARS-CoV-2: How Mutation of Non-Structural Protein 6 (NSP6) Could Affect Viral Autophagy. J. Infect. 2020, 81, e24–e27. [Google Scholar] [CrossRef] [PubMed]
- Kistler, K.E.; Huddleston, J.; Bedford, T. Rapid and parallel adaptive mutations in spike S1 drive clade success in SARS-CoV-2. Cell Host Microbe 2022, 30, 545–555. [Google Scholar] [CrossRef]
- McAuley, A.J.; Kuiper, M.J.; Durr, P.A.; Bruce, M.P.; Barr, J.; Todd, S.; Au, G.G.; Blasdell, K.; Tachedjian, M.; Lowther, S.; et al. Experimental and in silico evidence suggests vaccines are unlikely to be affected by D614G mutation in SARS-CoV-2 spike protein. npj Vaccines 2020, 5, 96. [Google Scholar] [CrossRef] [PubMed]
- Jangra, S.; Ye, C.; Rathnasinghe, R.; Stadlbauer, D.; Alshammary, H.; Amoako, A.A.; Awawda, M.H.; Beach, K.F.; Bermúdez-González, M.C.; Chernet, R.L.; et al. SARS-CoV-2 spike E484K mutation reduces antibody neutralisation. Lancet Microbe 2021, 2, e283–e284. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Patil, V.M.; Gupta, M.K. Mutation informatics: SARS-CoV-2 receptor-binding domain of the spike protein. Drug Discov. Today 2022, 27, 103312. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.; Kasry, A.; Amin, M. The new SARS-CoV-2 strain shows a stronger binding affinity to ACE2 due to N501Y mutant. Med. Drug Discov. 2021, 10, 100086. [Google Scholar] [CrossRef]
- Yurkovetskiy, L.; Wang, X.; Pascal, K.E.; Tomkins-Tinch, C.; Nyalile, T.P.; Wang, Y.; Baum, A.; Diehl, W.E.; Dauphin, A.; Carbone, C.; et al. Structural and Functional Analysis of the D614G SARS-CoV-2 Spike Protein Variant. Cell 2020, 183, 739–751. [Google Scholar] [CrossRef]
- Lista, M.J.; Winstone, H.; Wilson, H.D.; Dyer, A.; Pickering, S.; Galao, R.P.; De Lorenzo, G.; Cowton, V.M.; Furnon, W.; Suarez, N.; et al. The P681H Mutation in the Spike Glycoprotein of the Alpha Variant of SARS-CoV-2 Escapes IFITM Restriction and Is Necessary for Type I Interferon Resistance. J. Virol. 2022, 96, e01250-22. [Google Scholar] [CrossRef]
- Fratev, F. N501Y and K417N mutations in the spike protein of SARS-CoV-2 alter the interactions with Both hACE2 and human-derived antibody: A free energy of perturbation retrospective study. J. Chem. Inf. Model. 2021, 61, 6079–6084. [Google Scholar] [CrossRef]
- Yamamoto, M.; Tomita, K.; Hirayama, Y.; Inoue, J.I.; Kawaguchi, Y.; Gohda, J. SARS-CoV-2 Omicron spike H655Y mutation is responsible for enhancement of the endosomal entry pathway and reduction of cell surface entry pathway. bioRxiv 2022. [Google Scholar] [CrossRef]
- Hu, B.; Chan, J.F.W.; Liu, H.; Liu, Y.; Chai, Y.; Shi, J.; Shuai, H.; Hou, Y.; Huang, X.; Yuen, T.T.-T.; et al. Spike mutations contributing to the altered entry preference of SARS-CoV-2 omicron BA.1 and BA.2. Emerg. Microbes Infect. 2022, 11, 2275–2287. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Triche, T.J.; Bard, J.D.; Biegel, J.A.; Judkins, A.R.; Gai, X. Spike Protein NTD mutation G142D in SARS-CoV-2 Delta VOC lineages is associated with frequent back mutations, increased viral loads, and immune evasion. medRxiv 2021. [Google Scholar] [CrossRef]
- Motozono, C.; Toyoda, M.; Zahradnik, J.; Saito, A.; Nasser, H.; Tan, T.S.; Ngare, I.; Kimura, I.; Uriu, K.; Kosugi, Y.; et al. SARS-CoV-2 spike L452R variant evades cellular immunity and increases infectivity. Cell Host Microbe 2021, 29, 1124–1136. [Google Scholar] [CrossRef] [PubMed]
- Di Giacomo, S.; Mercatelli, D.; Rakhimov, A.; Giorgi, F.M. Preliminary report on severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) Spike mutation T478K. J. Med. Virol. 2021, 93, 5638–5643. [Google Scholar] [CrossRef]
- Wu, H.; Xing, N.; Meng, K.; Fu, B.; Xue, W.; Dong, P.; Tang, W.; Xiao, Y.; Liu, G.; Luo, H.; et al. Nucleocapsid mutations R203K/G204R increase the infectivity, fitness, and virulence of SARS-CoV-2. Cell Host Microbe 2021, 29, 1788–1801. [Google Scholar] [CrossRef]
- Rahimi, A.; Mirzazadeh, A.; Tavakolpour, S. Genetics and Genomics of SARS-CoV-2: A Review of the Literature with the Special Focus on Genetic Diversity and SARS-CoV-2 Genome Detection. Genomics 2021, 113, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Rausch, J.W.; Capoferri, A.A.; Katusiime, M.G.; Patro, S.C.; Kearney, M.F. Low Genetic Diversity May Be an Achilles Heel of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 24614–24616. [Google Scholar] [CrossRef]
- Lokman, S.M.; Rasheduzzaman, M.; Salauddin, A.; Barua, R.; Tanzina, A.Y.; Rumi, M.H.; Hossain, M.I.; Siddiki, A.M.A.M.Z.; Mannan, A.; Hasan, M.M. Exploring the Genomic and Proteomic Variations of SARS-CoV-2 Spike Glycoprotein: A Computational Biology Approach. Infect. Genet. Evol. 2020, 84, 104389. [Google Scholar] [CrossRef]
- Tang, X.; Wu, C.; Li, X.; Song, Y.; Yao, X.; Wu, X.; Duan, Y.; Zhang, H.; Wang, Y.; Qian, Z.; et al. On the Origin and Continuing Evolution of SARS-CoV-2. Natl. Sci. Rev. 2020, 7, 1012–1023. [Google Scholar] [CrossRef] [Green Version]
- Amicone, M.; Borges, V.; Alves, M.J.; Isidro, J.; Zé-Zé, L.; Duarte, S.; Vieira, L.; Guiomar, R.; Gomes, J.P.; Gordo, I. Mutation Rate of SARS-CoV-2 and Emergence of Mutators during Experimental Evolution. Evol. Med. Public Health 2022, 10, 142–155. [Google Scholar] [CrossRef]
- Chaw, S.M.; Tai, J.H.; Chen, S.L.; Hsieh, C.H.; Chang, S.Y.; Yeh, S.H.; Yang, W.S.; Chen, P.J.; Wang, H.Y. The Origin and Underlying Driving Forces of the SARS-CoV-2 Outbreak. J. Biomed. Sci. 2020, 27, 73. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, X.; Zhou, J.; Dong, Y.; Jiang, W.; Jiang, W. Rampant C-to-U Deamination Accounts for the Intrinsically High Mutation Rate in SARS-CoV-2 Spike Gene. RNA 2022, 28, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.A.; VanInsberghe, D.; Koelle, K. Insights from SARS-CoV-2 Sequences. Science 2021, 371, 466–467. [Google Scholar] [CrossRef]
- Ye, C.; Thornlow, B.; Kramer, A.; McBroome, J.; Hinrichs, A.; Corbett-Detig, R.; Turakhia, Y. Pandemic-Scale Phylogenetics. bioRxiv 2021. [Google Scholar] [CrossRef]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Puech, J.; et al. Reduced Sensitivity of SARS-CoV-2 Variant Delta to Antibody Neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Planas, D.; Saunders, N.; Maes, P.; Guivel-Benhassine, F.; Planchais, C.; Buchrieser, J.; Bolland, W.H.; Porrot, F.; Staropoli, I.; Lemoine, F.; et al. Considerable Escape of SARS-CoV-2 Omicron to Antibody Neutralization. Nature 2022, 602, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Priya, P.; Shanker, A. Coevolutionary forces shaping the fitness of SARS-CoV-2 spike glycoprotein against human receptor ACE2. Infect. Genet. Evol. 2021, 87, 104646. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Losada, M.; Arenas, M.; Galán, J.C.; Palero, F.; González-Candelas, F. Recombination in Viruses: Mechanisms, Methods of Study, and Evolutionary Consequences. Infect. Genet. Evol. 2015, 30, 296–307. [Google Scholar] [CrossRef]
- Schierup, M.H.; Hein, J. Consequences of Recombination on Traditional Phylogenetic Analysis. Genetics 2000, 156, 879–891. [Google Scholar] [CrossRef]
- Arenas, M.; Posada, D. The Effect of Recombination on the Reconstruction of Ancestral Sequences. Genetics 2010, 184, 1133–1139. [Google Scholar] [CrossRef]
- Arenas, M.; Posada, D. The Influence of Recombination on the Estimation of Selection from Coding Sequence Alignments. In Natural Selection: Methods and Applications; Fares, M.A., Ed.; CRC Press: Boca Raton, FL, USA, 2014; Chapter 6; pp. 112–125. [Google Scholar]
- Simon-Loriere, E.; Holmes, E.C. Why Do RNA Viruses Recombine? Nat. Rev. Microbiol. 2011, 9, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [PubMed]
- VanInsberghe, D.; Neish, A.; Lowen, A.C.; Koelle, K. Identification of SARS-CoV-2 Recombinant Genomes. biorXiv 2020. [Google Scholar] [CrossRef]
- Zhu, Z.; Meng, K.; Meng, G. Genomic recombination events may reveal the evolution of coronavirus and the origin of SARS-CoV-2. Sci. Rep. 2020, 10, 21617. [Google Scholar] [CrossRef]
- Li, X.; Giorgi, E.E.; Marichannegowda, M.H.; Foley, B.; Xiao, C.; Kong, X.-P.; Chen, Y.; Gnanakaran, S.; Korber, B.; Gao, F. Emergence of SARS-CoV-2 through Recombination and Strong Purifying Selection. Sci. Adv. 2020, 6, eabb9153. [Google Scholar] [CrossRef] [PubMed]
- Posada, D. Evaluation of Methods for Detecting Recombination from DNA Sequences: Empirical Data. Mol. Biol. Evol. 2002, 19, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. Evaluation of Methods for Detecting Recombination from DNA Sequences: Computer Simulations. Proc. Natl. Acad. Sci. USA 2001, 98, 13757–13762. [Google Scholar] [CrossRef]
- Arenas, M. Computational Analysis of Recombination in Viral Nucleotide Sequences. In Encyclopedia of Virology, 4th ed.; Bamford, D.H., Zuckerman, M., Eds.; Academic Press: Oxford, UK, 2021; Volume 1, pp. 108–115. [Google Scholar]
- Nie, Q.; Li, X.; Chen, W.; Liu, D.; Chen, Y.; Li, H.; Li, D.; Tian, M.; Tan, W.; Zai, J. Phylogenetic and Phylodynamic Analyses of SARS-CoV-2. Virus Res. 2020, 287, 198098. [Google Scholar] [CrossRef]
- Richard, D.; Owen, C.J.; van Dorp, L.; Balloux, F. No Detectable Signal for Ongoing Genetic Recombination in SARS-CoV-2. bioRxiv 2020. [Google Scholar] [CrossRef]
- Jackson, B.; Rambaut, A.; Pybus, O.G.; Robertson, D.L.; Connor, T.; Loman, N.J. COG-UK Consortium (2021) Recombinant SARS-CoV-2 Genomes Involving Lineage B.1.1.7 in the UK. Virological. Available online: https://virological.org/t/recombinant-sars-cov-2-genomes-involving-lineage-b-1-1-7-in-the-uk/658 (accessed on 22 November 2022).
- Lacek, K.A.; Rambo-Martin, B.L.; Batra, D.; Zheng, X.; Sakaguchi, H.; Peacock, T.; Keller, M.; Wilson, M.M.; Sheth, M.; Davis, M.L.; et al. Identification of a Novel SARS-CoV-2 Delta-Omicron Recombinant Virus in the United States. bioRxiv 2022. [Google Scholar] [CrossRef]
- Lopes, J.S.; Arenas, M.; Posada, D.; Beaumont, M.A. Coestimation of Recombination, Substitution and Molecular Adaptation Rates by Approximate Bayesian Computation. Heredity 2014, 112, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Arenas, M.; Lorenzo-Redondo, R.; Lopez-Galindez, C. Influence of Mutation and Recombination on HIV-1 in Vitro Fitness Recovery. Mol. Phylogenet. Evol. 2016, 94, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Castelhano, N.; Araujo, N.M.; Arenas, M. Heterogeneous Recombination among Hepatitis B Virus Genotypes. Infect. Genet. Evol. 2017, 54, 486–490. [Google Scholar] [CrossRef]
- Wertheim, J.O.; Wang, J.C.; Leelawong, M.; Martin, D.P.; Havens, J.L.; Chowdhury, M.A.; Pekar, J.E.; Amin, H.; Arroyo, A.; Awandare, G.A.; et al. Detection of SARS-CoV-2 Intra-Host Recombination during Superinfection with Alpha and Epsilon Variants in New York City. Nat. Commun. 2022, 13, 3645. [Google Scholar] [CrossRef] [PubMed]
- Combes, P.; Bisseux, M.; Bal, A.; Marin, P.; Latour, J.; Archimbaud, C.; Brebion, A.; Chabrolles, H.; Regagnon, C.; Lafolie, J.; et al. Evidence of Co-Infections during Delta and Omicron SARS-CoV-2 Variants Co-Circulation through Prospective Screening and Sequencing. Clin. Microbiol. Infect. 2022, 28, 1503. [Google Scholar] [CrossRef] [PubMed]
- Simon-Loriere, E.; Montagutelli, X.; Lemoine, F.; Donati, F.; Touret, F.; Bourret, J.; Prot, M.; Munier, S.; Attia, M.; Conquet, L.; et al. Rapid characterization of a Delta-Omicron SARS-CoV-2 recombinant detected in Europe. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Colson, P.; Fournier, P.E.; Delerce, J.; Million, M.; Bedotto, M.; Houhamdi, L.; Yahi, N.; Bayette, J.; Levasseur, A.; Fantini, J.; et al. Culture and Identification of a “Deltamicron” SARS-CoV-2 in a Three Cases Cluster in Southern France. J. Med. Virol. 2022, 94, 3739–3749. [Google Scholar] [CrossRef]
- Bolze, A.; Basler, T.; White, S.; Rossi, A.D.; Wyman, D.; Roychoudhury, P.; Greninger, A.L.; Hayashibara, K.; Beatty, M.; Shah, S.; et al. Evidence for SARS-CoV-2 Delta and Omicron Co-Infections and Recombination. Med 2022, 3, 848–859. [Google Scholar] [CrossRef]
- Karbalaei, M.; Keikha, M. Deltacron Is a Recombinant Variant of SARS-CoV-2 but Not a Laboratory Mistake. Ann. Med. Surg. 2022, 79, 104032. [Google Scholar] [CrossRef]
- Wang, L.; Gao, G.F. The “Wolf” Is Indeed Coming: Recombinant “Deltacron” SARS-CoV-2 Detected. China CDC Wkly. 2022, 4, 285–287. [Google Scholar] [CrossRef]
- Kreier, F. Deltacron: The Story of the Variant That Wasn’t. Nature 2022, 602, 19. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Bhattacharya, M.; Sharma, A.R.; Dhama, K. Recombinant SARS-CoV-2 Variants XD, XE, and XF: The Emergence of Recombinant Variants Requires an Urgent Call for Research–Correspondence. Int. J. Surg. 2022, 102, 106670. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, R.K.; Kandi, V.; Tuli, H.S.; Chakraborty, C.; Dhama, K. The Recombinant Variants of SARS-CoV-2: Concerns Continues amid COVID-19 Pandemic. J. Med. Virol. 2022, 94, 3506–3508. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.; Lan, W.; Wu, X.; Zhao, T.; Duan, B.; Yang, P.; Ren, Y.; Quan, L.; Zhao, W.; Seto, D.; et al. Tracking SARS-CoV-2 Omicron diverse spike gene mutations identifies multiple inter-variant recombination events. Signal Transduct. Target. Ther. 2020, 7, 138. [Google Scholar] [CrossRef]
- Burel, E.; Colson, P.; Lagier, J.C.; Levasseur, A.; Bedotto, M.; Lavrard-Meyer, P.; Fournier, P.E.; La Scola, B.; Raoult, D. Sequential Appearance and Isolation of a SARS-CoV-2 Recombinant between Two Major SARS-CoV-2 Variants in a Chronically Infected Immunocompromised Patient. Viruses 2022, 14, 1266. [Google Scholar] [CrossRef] [PubMed]
- Escalera-Zamudio, M.; Castelán-Sánchez, H.G.; Delaye-Arredondo, L.J.; Gutiérrez, B. RE: Proposal to Redesignate B.1.631 as Recombinant Lineage XB. Virological. Available online: https://virological.org/t/re-proposal-to-redesignate-b-1-631-as-recombinant-lineage-xb/746 (accessed on 24 November 2022).
- Lindh, E.; Smura, T.; Blomqvist, S.; Liitsola, K.; Vauhkonen, H.; Savolainen, L.; Ikonen, J.; Ronkainen, J.; Taskila, J.; Taskila, T.; et al. Genomic and Epidemiological Report of the Recombinant XJ Lineage SARS-CoV-2 Variant, Detected in Northern Finland, January 2022. Eurosurveillance 2022, 27, 2200257. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Ng, D.Y.M.; Liu, G.Y.Z.; Cheng, S.S.M.; Krishnan, P.; Chang, L.D.J.; Cheuk, S.S.Y.; Hui, M.M.Y.; Lam, T.T.Y.; Peiris, M.; et al. Recombinant BA.1/BA.2 SARS-CoV-2 Virus in Arriving Travelers, Hong Kong, February 2022. Emerg. Infect. Dis. 2022, 28, 1276–1278. [Google Scholar] [CrossRef]
- Liu, X.; Xiong, J.; Sun, Z.; Hu, J.; Thilakavathy, K.; Chen, M.; Zhao, Q.; Feng, Y.; Jiang, Q.; Xiong, C. Omicron: A chimera of two early SARS-CoV-2 lineages. Signal Transduct. Target. Ther. 2022, 7, 90. [Google Scholar] [CrossRef]
- Turkahia, Y.; Thornlow, B.; Hinrichs, A.; McBroome, J.; Ayala, N.; Ye, C.; Maio, N.D.; Haussler, D.; Lanfear, R.; Corbett-Detig, R. Pandemic-Scale Phylogenomics Reveals Elevated Recombination Rates in the SARS-CoV-2 Spike Region. bioRxiv 2021. [Google Scholar] [CrossRef]
- Gutierrez, B.; Castelán Sánchez, H.G.; da Silva Candido, D.; Jackson, B.; Fleishon, S.; Houzet, R.; Ruis, C.; Delaye, L.; Faria, N.R.; Rambaut, A.; et al. Emergence and Widespread Circulation of a Recombinant SARS-CoV-2 Lineage in North America. Cell Host Microbe 2022, 30, 1112–1123. [Google Scholar] [CrossRef]
- Ignatieva, A.; Hein, J.; Jenkins, P.A. Ongoing Recombination in SARS-CoV-2 Revealed through Genealogical Reconstruction. Mol. Biol. Evol. 2022, 39, msac028. [Google Scholar] [CrossRef] [PubMed]
- Varabyou, A.; Pockrandt, C.; Salzberg, S.L.; Pertea, M. Rapid Detection of Inter-Clade Recombination in SARS-CoV-2 with Bolotie. Genetics 2021, 218, iyab074. [Google Scholar] [CrossRef] [PubMed]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B.; et al. Tracking Changes in SARS-CoV-2 Spike: Evidence That D614G Increases Infectivity of the COVID-19 Virus. Cell 2020, 182, 812–827. [Google Scholar] [CrossRef] [PubMed]
- Taghizadeh, P.; Salehi, S.; Heshmati, A.; Houshmand, S.M.; InanlooRahatloo, K.; Mahjoubi, F.; Sanati, M.H.; Yari, H.; Alavi, A.; Jamehdar, S.A.; et al. Study on SARS-CoV-2 Strains in Iran Reveals Potential Contribution of Co-Infection with and Recombination between Different Strains to the Emergence of New Strains. Virology 2021, 562, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Bukin, Y.S.; Bondaryuk, A.N.; Kulakova, N.V.; Balakhonov, S.V.; Dzhioev, Y.P.; Zlobin, V.I. Phylogenetic Reconstruction of the Initial Stages of the Spread of the SARS-CoV-2 Virus in the Eurasian and American Continents by Analyzing Genomic Data. Virus Res. 2021, 305, 198551. [Google Scholar] [CrossRef]
- Duchene, S.; Featherstone, L.; Haritopoulou-Sinanidou, M.; Rambaut, A.; Lemey, P.; Baele, G. Temporal Signal and the Phylodynamic Threshold of SARS-CoV-2. Virus Evol. 2020, 6, veaa061. [Google Scholar] [CrossRef]
- Engelbrecht, S.; Delaney, K.; Kleinhans, B.; Wilkinson, E.; Tegally, H.; Stander, T.; van Zyl, G.; Preiser, W.; de Oliveira, T. Multiple Early Introductions of SARS-CoV-2 to Cape Town, South Africa. Viruses 2021, 13, 526. [Google Scholar] [CrossRef]
- Ghafari, M.; du Plessis, L.; Raghwani, J.; Bhatt, S.; Xu, B.; Pybus, O.G.; Katzourakis, A. Purifying Selection Determines the Short-Term Time Dependency of Evolutionary Rates in SARS-CoV-2 and PH1N1 Influenza. Mol. Biol. Evol. 2022, 39, msac009. [Google Scholar] [CrossRef]
- Lai, A.; Bergna, A.; Acciarri, C.; Galli, M.; Zehender, G. Early Phylogenetic Estimate of the Effective Reproduction Number of SARS-CoV-2. J. Med. Virol. 2020, 92, 675–679. [Google Scholar] [CrossRef] [Green Version]
- Pekar, J.; Worobey, M.; Moshiri, N.; Scheffler, K.; Wertheim, J.O. Timing the SARS-CoV-2 Index Case in Hubei Province. Science 2021, 372, 412–417. [Google Scholar] [CrossRef]
- Pipes, L.; Wang, H.; Huelsenbeck, J.P.; Nielsen, R. Assessing Uncertainty in the Rooting of the SARS-CoV-2 Phylogeny. Mol. Biol. Evol. 2021, 38, 1537–1543. [Google Scholar] [CrossRef] [PubMed]
- Sallam, M.; Ababneh, N.A.; Dababseh, D.; Bakri, F.G.; Mahafzah, A. Temporal Increase in D614G Mutation of SARS-CoV-2 in the Middle East and North Africa. Heliyon 2021, 7, e06035. [Google Scholar] [CrossRef] [PubMed]
- Tay, J.H.; Porter, A.F.; Wirth, W.; Duchene, S. The Emergence of SARS-CoV-2 Variants of Concern Is Driven by Acceleration of the Substitution Rate. Mol. Biol. Evol. 2022, 39, msac013. [Google Scholar] [CrossRef] [PubMed]
- Bromham, L.; Penny, D. The Modern Molecular Clock. Nat. Rev. Genet. 2003, 4, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Del Amparo, R.; Branco, C.; Arenas, J.; Vicens, A.; Arenas, M. Analysis of Selection in Protein-Coding Sequences Accounting for Common Biases. Brief. Bioinform. 2021, 22, bbaa431. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Zhao, S.; Liu, Q.; Zhang, X.; Sha, T.; Su, Y.; Zhao, W.; Bao, Y.; Xue, Y.; Chen, H. Ongoing Positive Selection Drives the Evolution of SARS-CoV-2 Genomes. Genom. Proteom. Bioinform. 2022, in press. [CrossRef]
- Rochman, N.D.; Wolf, Y.I.; Faure, G.; Mutz, P.; Zhang, F.; Koonin, E.V. Ongoing Global and Regional Adaptive Evolution of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2021, 118, e2104241118. [Google Scholar] [CrossRef]
- Velazquez-Salinas, L.; Zarate, S.; Eberl, S.; Gladue, D.P.; Novella, I.; Borca, M.V. Positive Selection of ORF1ab, ORF3a, and ORF8 Genes Drives the Early Evolutionary Trends of SARS-CoV-2 During the 2020 COVID-19 Pandemic. Front. Microbiol. 2020, 11, 550674. [Google Scholar] [CrossRef]
- Koçhan, N.; Eskier, D.; Suner, A.; Karakülah, G.; Oktay, Y. Different Selection Dynamics of S and RdRp between SARS-CoV-2 Genomes with and without the Dominant Mutations. Infect. Genet. Evol. 2021, 91, 104796. [Google Scholar] [CrossRef]
- Dearlove, B.; Lewitus, E.; Bai, H.; Li, Y.; Reeves, D.B.; Joyce, M.G.; Scott, P.T.; Amare, M.F.; Vasan, S.; Michael, N.L.; et al. A SARS-CoV-2 Vaccine Candidate Would Likely Match All Currently Circulating Variants. Proc. Natl. Acad. Sci. USA 2020, 117, 23652–23662. [Google Scholar] [CrossRef]
- Kumar, R.; Verma, H.; Singhvi, N.; Sood, U.; Gupta, V.; Singh, M.; Kumari, R.; Hira, P.; Nagar, S.; Talwar, C.; et al. Comparative Genomic Analysis of Rapidly Evolving SARS-CoV-2 Reveals Mosaic Pattern of Phylogeographical Distribution. mSystems 2020, 5, e00505-20. [Google Scholar] [CrossRef] [PubMed]
- Chiara, M.; Horner, D.S.; Gissi, C.; Pesole, G. Comparative Genomics Suggests Limited Variability and Similar Evolutionary Patterns between Major Clades of SARS-CoV-2. bioRxiv 2020. [Google Scholar] [CrossRef]
- Lythgoe, K.A.; Hall, M.; Ferretti, L.; Cesare, M.; MacIntyre-Cockett, G.; Trebes, A.; Andersson, M.; Otecko, N.; Wise, E.L.; Moore, N.; et al. Within-Host Genomics of SARS-CoV-2. bioRxiv 2020. [Google Scholar] [CrossRef]
- Lo Presti, A.; Rezza, G.; Stefanelli, P. Selective Pressure on SARS-CoV-2 Protein Coding Genes and Glycosylation Site Prediction. Heliyon 2020, 6, e05001. [Google Scholar] [CrossRef] [PubMed]
- Berrio, A.; Gartner, V.; Wray, G.A. Positive Selection within the Genomes of SARS-CoV-2 and Other Coronaviruses Independent of Impact on Protein Function. PeerJ 2020, 8, e10234. [Google Scholar] [CrossRef]
- Tegally, H.; Wilkinson, E.; Giovanetti, M.; Iranzadeh, A.; Fonseca, V.; Giandhari, J.; Doolabh, D.; Pillay, S.; San, E.J.; Msomi, N.; et al. Detection of a SARS-CoV-2 Variant of Concern in South Africa. Nature 2021, 592, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Volz, E.; Hill, V.; McCrone, J.T.; Price, A.; Jorgensen, D.; O’Toole, Á.; Southgate, J.; Johnson, R.; Jackson, B.; Nascimento, F.F.; et al. Evaluating the Effects of SARS-CoV-2 Spike Mutation D614G on Transmissibility and Pathogenicity. Cell 2021, 184, 64–75. [Google Scholar] [CrossRef]
- Wibmer, C.K.; Ayres, F.; Hermanus, T.; Madzivhandila, M.; Kgagudi, P.; Oosthuysen, B.; Lambson, B.E.; de Oliveira, T.; Vermeulen, M.; van der Berg, K.; et al. SARS-CoV-2 501Y.V2 escapes neutralization by South African COVID-19 donor plasma. Nat. Med. 2021, 27, 622–625. [Google Scholar] [CrossRef]
- Martin, D.P.; Weaver, S.; Tegally, H.; San, J.E.; Shank, S.D.; Wilkinson, E.; Lucaci, A.G.; Giandhari, J.; Naidoo, S.; Pillay, Y.; et al. The Emergence and Ongoing Convergent Evolution of the SARS-CoV-2 N501Y Lineages. Cell 2021, 184, 5189–5200. [Google Scholar] [CrossRef]
- McCallum, M.; De Marco, A.; Lempp, F.A.; Tortorici, M.A.; Pinto, D.; Walls, A.C.; Beltramello, M.; Chen, A.; Liu, Z.; Zatta, F.; et al. N-terminal Domain Antigenic Mapping Reveals a Site of Vulnerability for SARS-CoV-2. Cell 2021, 184, 2332–2347. [Google Scholar] [CrossRef]
- Upadhyay, V.; Patrick, C.; Lucas, A.; Mallela, K.M.G. Convergent Evolution of Multiple Mutations Improves the Viral Fitness of SARS-CoV-2 Variants by Balancing Positive and Negative Selection. Biochemistry 2022, 61, 963–980. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Nair, M.S.; Liu, L.; Iketani, S.; Luo, Y.; Guo, Y.; Wang, M.; Yu, J.; Zhang, B.; Kwong, P.D.; et al. Antibody Resistance of SARS-CoV-2 Variants B.1.351 and B.1.1.7. Nature 2021, 593, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Schmidt, F.; Weisblum, Y.; Muecksch, F.; Barnes, C.O.; Finkin, S.; Schaefer-Babajew, D.; Cipolla, M.; Gaebler, C.; Lieberman, J.A.; et al. MRNA Vaccine-Elicited Antibodies to SARS-CoV-2 and Circulating Variants. Nature 2021, 592, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Lubinski, B.; Fernandes, M.H.; Frazier, L.; Tang, T.; Daniel, S.; Diel, D.G.; Jaimes, J.A.; Whittaker, G.R. Functional evaluation of the P681H mutation on the proteolytic activation of the SARS-CoV-2 variant B.1.1.7 (Alpha) spike. Iscience 2022, 25, 103589. [Google Scholar] [CrossRef] [PubMed]
- Marques-Pereira, C.; Pires, M.N.; Gouveia, R.P.; Pereira, N.N.; Caniceiro, A.B.; Rosário-Ferreira, N.; Moreira, I.S. SARS-CoV-2 Membrane Protein: From Genomic Data to Structural New Insights. Int. J. Mol. Sci. 2022, 23, 2986. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Tian, C.; Liu, P.; Guo, D.; Zheng, W.; Huang, X.; Zhang, Y.; Liu, L. Effects of SARS-CoV-2 Mutations on Protein Structures and Intraviral Protein-Protein Interactions. J. Med. Virol. 2021, 93, 2132–2140. [Google Scholar] [CrossRef] [PubMed]
- Omotoso, O.E. Contributory Role of SARS-CoV-2 Genomic Variations and Life Expectancy in COVID-19 Transmission and Low Fatality Rate in Africa. Egypt. J. Med. Hum. Genet. 2020, 21, 72. [Google Scholar] [CrossRef]
- Morales, A.C.; Rice, A.M.; Ho, A.T.; Mordstein, C.; Mühlhausen, S.; Watson, S.; Cano, L.; Young, B.; Kudla, G.; Hurst, L.D. Causes and Consequences of Purifying Selection on SARS-CoV-2. Genome Biol. Evol. 2021, 13, evab196. [Google Scholar] [CrossRef]
- Azkur, A.K.; Akdis, M.; Azkur, D.; Sokolowska, M.; van de Veen, W.; Brüggen, M.C.; O’Mahoni, L.; Gao, Y.; Nadeau, K.; Akdis, C.A. Immune response to SARS-CoV-2 and mechanisms of immunopathological changes in COVID-19. Allergy 2020, 75, 1564–1581. [Google Scholar] [CrossRef]
- Thomson, E.C.; Rosen, L.E.; Shepherd, J.G.; Spreafico, R.; da Silva Filipe, A.; Wojcechowskyj, J.A.; Davis, C.; Piccoli, L.; Pascall, D.J.; Dillen, J.; et al. Circulating SARS-CoV-2 spike N439K variants maintain fitness while evading antibody-mediated immunity. Cell 2021, 184, 1171–1187. [Google Scholar] [CrossRef]
- Greaney, A.J.; Starr, T.N.; Gilchuk, P.; Zost, S.J.; Binshtein, E.; Loes, A.N.; Hilton, S.K.; Huddleston, J.; Eguia, R.; Crawford, K.H.D.; et al. Complete mapping of mutations to the SARS-CoV-2 spike receptor-binding domain that escape antibody recognition. Cell Host Microbe 2021, 29, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Greaney, A.J.; Loes, A.N.; Crawford, K.H.D.; Starr, T.N.; Malone, K.D.; Chu, H.Y.; Bloom, J.D. Comprehensive Mapping of Mutations in the SARS-CoV-2 Receptor-Binding Domain That Affect Recognition by Polyclonal Human Plasma Antibodies. Cell Host Microbe 2021, 29, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Starr, T.N.; Greaney, A.J.; Addetia, A.; Hannon, W.W.; Choudhary, M.C.; Dingens, A.S.; Li, J.Z.; Bloom, J.D. Prospective Mapping of Viral Mutations That Escape Antibodies Used to Treat COVID-19. Science 2021, 371, 850–854. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Arora, P.; Groß, R.; Seidel, A.; Hörnich, B.F.; Hahn, A.S.; Krüger, N.; Graichen, L.; Hofmann-Winkler, H.; Kempf, A.; et al. SARS-CoV-2 Variants B.1.351 and P.1 Escape from Neutralizing Antibodies. Cell 2021, 184, 2384–2393. [Google Scholar] [CrossRef]
- Mahase, E. Covid-19: Novavax Vaccine Efficacy Is 86% against UK Variant and 60% against South African Variant. BMJ 2021, 372, n296. [Google Scholar] [CrossRef]
- Tada, T.; Zhou, H.; Samanovic, M.I.; Dcosta, B.M.; Cornelius, A.; Herati, R.S.; Mulligan, M.J.; Landau, N.R. Neutralization of SARS-CoV-2 Variants by mRNA and Adenoviral Vector Vaccine-Elicited Antibodies. Front. Immunol. 2022, 13, 797589. [Google Scholar] [CrossRef]
- Tada, T.; Zhou, H.; Dcosta, B.M.; Samanovic, M.I.; Chivukula, V.; Herati, R.S.; Hubbard, S.R.; Mulligan, M.J.; Landau, N.R. Increased resistance of SARS-CoV-2 Omicron variant to neutralization by vaccine-elicited and therapeutic antibodies. EBioMedicine 2022, 78, 103944. [Google Scholar] [CrossRef]
- Focosi, D.; Maggi, F.; McConnell, S.; Casadevall, A. Very Low Levels of Remdesivir Resistance in SARS-COV-2 Genomes after 18 Months of Massive Usage during the COVID19 Pandemic: A GISAID Exploratory Analysis. Antivir. Res. 2022, 198, 105247. [Google Scholar] [CrossRef]
- Agostini, M.L.; Andres, E.L.; Sims, A.C.; Graham, R.L.; Sheahan, T.P.; Lu, X.; Smith, E.C.; Case, J.B.; Feng, J.Y.; Jordan, R.; et al. Coronavirus Susceptibility to the Antiviral Remdesivir (GS-5734) Is Mediated by the Viral Polymerase and the Proofreading Exoribonuclease. mBio 2018, 9, e00221-18. [Google Scholar] [CrossRef] [Green Version]
- Szemiel, A.M.; Merits, A.; Orton, R.J.; MacLean, O.A.; Pinto, R.M.; Wickenhagen, A.; Lieber, G.; Turnbull, M.L.; Wang, S.; Furnon, W.; et al. In Vitro Selection of Remdesivir Resistance Suggests Evolutionary Predictability of SARS-CoV-2. PLoS Pathog. 2021, 17, e1009929. [Google Scholar] [CrossRef]
- Gandhi, S.; Klein, J.; Robertson, A.J.; Peña-Hernández, M.A.; Lin, M.J.; Roychoudhury, P.; Lu, P.; Fournier, J.; Ferguson, D.; Mohamed Bakhash, S.A.K.; et al. De Novo Emergence of a Remdesivir Resistance Mutation during Treatment of Persistent SARS-CoV-2 Infection in an Immunocompromised Patient: A Case Report. Nat. Commun. 2022, 13, 1547. [Google Scholar] [CrossRef]
- Huang, Y.Z.; Kuan, C.C. Vaccination to reduce severe COVID-19 and mortality in COVID-19 patients: A systematic review and meta-analysis. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 1770–1776. [Google Scholar] [CrossRef] [PubMed]
- Watson, O.J.; Barnsley, G.; Toor, J.; Hogan, A.B.; Winskill, P.; Ghani, A.C. Global impact of the first year of COVID-19 vaccination: A mathematical modelling study. Lancet Infect. Dis. 2022, 22, 1293–1302. [Google Scholar] [CrossRef] [PubMed]
- Wells, C.R.; Galvani, A.P. The global impact of disproportionate vaccination coverage on COVID-19 mortality. Lancet Infect. Dis. 2022, 22, 1254–1255. [Google Scholar] [CrossRef] [PubMed]
- Morel, B.; Barbera, P.; Czech, L.; Bettisworth, B.; Hübner, L.; Lutteropp, S.; Serdari, D.; Kostaki, E.G.; Mamais, I.; Kozlov, A.M.; et al. Phylogenetic Analysis of SARS-CoV-2 Data Is Difficult. Mol. Biol. Evol. 2021, 38, 1777–1791. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| VOC (PANGO Lineage) | Detected Mutations that Define VOCs (Except Omicron, Table 2) at Each Gene | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ORF1a | ORF1b | S | ORF3a | E | M | ORF6 | ORF7a | ORF7b | ORF8 | N | |
| Alpha (B.1.1.7) | Aa: T1001I A1708D I2230T Del: 3675–3677 | Aa: P314L | Aa: P9L V483F E484K F486I Q498R N501Y A570D D614G P681H T716I S982A D1118H Del: H69 V70 | Aa: Q27* R52I Y73C S84L | Aa: D3L R203K G204R S235F | ||||||
| Beta (B.1.351) | Aa: T265I K1655N K3353R Del: 3675–3677 | Aa: P314L | Aa: S13I D80A D215G, K417N E484K N501Y D614G A701V A879S Del: L242 A243 L244 | Aa: Q57H S171L | Aa: P71L | Aa: S84L | Aa: T205I | ||||
| Gamma (P.1) | Aa: S1188L K1795Q Del: 3675–3677 | Aa: P314L E1264D | Aa: L18F T20N P26S R78M D138Y R190S K417T E484K N501Y D614G H655Y T1027I V1176F | Aa: S253P | Aa: S84L E92K | Aa: P80R R203K G204R | |||||
| Delta (B.617.2) | Aa: P314L G662S P1000L | Aa: T19R T95I G142D E156G L452R T478K R567I D614G H655Y P681R D950N T1117I Del: F157 R158 | Aa: S26L | Aa: I82T | Aa: V82A, T120I | Aa: S84L Del: 119–120 | Aa: D63G R203M D377Y | ||||
| Omicron (PANGO Lineage) | Detected Mutations that Define Omicron Lineages at Each Gene | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ORF1a | ORF1b | S | ORF3a | E | M | ORF6 | ORF7b | ORF8 | N | |
| BA.1 | Aa: K856R L2084I A2710T T3255I P3395H I3758V Del: S2083 3674-3676 | Aa: P314L I1566V | Aa: A67V T95I G142D L212I G339D S371L S373P S375F K417N N440K G446S S477N T478K E484A Q493R G496S Q498R N501Y Y505H T547K D614G H655Y N679K P681H N764K D796Y N856K Q954H N969K L981F Del: 69–70 143–145 211 Ins: 214EPE | Aa: T9I | Aa: D3G Q19E A63T | Aa: S84L | Aa: P13L R203K G204R Del: 31–33 | |||
| BA.2 | Aa: S135R T842I G1307S L3027F T3090I L3201F T3255I P3395H Del: 3675–3677 | Aa: P314L R1315C I1566V T2163I | Aa: T19I L24S G142D V213G G339D S371F S373P S375F T376A D405N R408S K417N N440K S477N T478K E484A Q493R Q498R N501Y Y505H D614G H655Y N679K P681H N764K D796Y Q954H N969K Del: 25–27 | Aa: T223I | Aa: T9I | Aa: Q19E A63T | Aa: D61L | Aa: S84L | Aa: P13L R203K G204R S413R Del: 31–33 | |
| BA.3 | Aa: S153R G1307S T3090I T3255I P3395H A3657V Del: 3675–3677 | Aa: P314L I1566V | Aa: A67V T95I G142D L212I G339D S371F S373P S375F D405N K417N N440K G446S S477N T478K E484A Q493R Q498R N501Y Y505H D614G H655Y N679K P681H N764K D796Y Q954H N969K Del: 69–70 143–145 211 | Aa: T223I | Aa: T9I | Aa: Q19E A63T | Aa: S84L | Aa: P13L R203K G204R S413R Del: 31–33 | ||
| BA.4 | Aa: S135R T842I G1307S L3027F T3090I T3255I P3395H Del: 141–143 3675–3677 Nt: G12160A | Aa: P314L R1315C I1566V T2163I | Aa: T19I L24S G142D V213G G339D S371F S373P S375F T376A D405N R408S K417N N440K G446S L452R S477N T478K E484A F486V Q498R N501Y Y505H T547K D614G H655Y N679K P681H N764K D796Y Q954H N969K Del: 25–27 69–70 | Aa: T223I | Aa: T9I | Aa: Q19E A63T | Aa: D61L | Aa: L11F | Aa: P13L P151S R203K G204R S413R Del: 31–33 | |
| BA.5 | Aa: S135R T842I G1307S L1507F L3027F T3090I T3255I P3395H Del: 3675–3677 Nt: G12160A | Aa: P314L R1315C I1566V T2163I | Same mutations as BA.4 | Aa: T223I | Aa: T9I | Aa: D3N Q19E A63T | Aa: P13L R203K G204R S413R Del: 31–33 | |||
| Gene | Mutation (s) | Consequences | References |
|---|---|---|---|
| ORF1a | Del: 3675–3677 | Deletion located in the protein nsp6, which is important for the synthesis of RNA. This deletion removes amino acids from a transmembrane loop A very similar deletion was observed in the VOC Omicron BA.1 (deletion 3674–3676), which was associated with a favoring effect about increasing mutability | [42,43] |
| ORF1b | P314L | This mutation is in linkage disequilibrium with the D614G mutation of the S gene | [44] |
| S | E484K | This mutation induces escape to monoclonal antibodies and reduces the neutralizing capacity of convalescent and post-vaccination polyclonal sera | [45] |
| Q498R | This mutation increases the affinity between SARS-CoV-2 RBD and human ACE2 | [46] | |
| N501Y | This mutation increases the affinity between SARS-CoV-2 RBD and human ACE2 | [47] | |
| D614G | This mutation increases infectivity by changing the formation of the spike protein to a competent state for binding with ACE2 | [48] | |
| P681H | This mutation confers resistance to type I interferons and reduces dependence on endosomal cathepsins favoring cell entry | [49] | |
| K417N | This mutation reduces the activity of human and commercial antibodies but also reduces the affinity of the spike protein with human ACE2 | [50] | |
| H655Y | This mutation increases the fusogenicity with human cell membrane by using cathepsin-mediated entry and reduces the entry using the serine transmembrane protease 2 | [51,52] | |
| T95I | - | - | |
| G142D | Mutation associated with immune evasion, back mutations, and increased viral load | [53] | |
| L452R | This mutation increases the infectivity and fusogenicity and promotes viral replication | [54] | |
| T478K | This mutation increases the electrostatic potential of the spike protein at human ACE2 binding and can also play a role in immune escape | [55] | |
| ORF8 | S84L | - | - |
| N | R203K, G204R | These mutations increase infectivity, fitness and virulence | [56] |
| Variants Involved | Breakpoint Interval/Gene or Genes Involved | Additional Information | References |
|---|---|---|---|
| Alpha (VOC) and Epsilon (VOI) | Inside the genes S, N, and ORF8 [88] | - | [88] |
| Delta (VOC) and Omicron (VOC) | 19220–21618/ORF1ab and S [89] 22034–22194/S [89,90,91] 22035–22577/S [84] 22204–22578/S [92] 22218–22586/S [89] 25469–25584/ORF3a [90,91] | Recombinant named “Deltacron” or “Deltamicron”. The concern about this chimera lies in the combination of the high transmissibility of Omicron and the virulence and severe disease caused by Delta [90,93] | [84,89,90,91,92,93,94,95,96,97,98] |
| Alpha (VOC) and B.1.160 | 17109–18877/ORF1ab [90,91] 25710–27972/ORF3a, E, M, ORF6, ORF7a, ORF7b [90,91] | Recombination occurred among parents of these variants | [90,91,99] |
| Various lineages and Alpha (VOC) | 21255–21764/ORF1ab, S [83] 6528–6954/ORF1ab [83] 24914–28651/S, ORF3a, E, M, ORF6, ORF7a, ORF7b, ORF8, N [83] 21575–23063/S [83] 11396–21991/ORF1ab, S [83] Window nt 3267–5388 gene ORF1ab [83] 12534–21765/ORF1ab, S [83] 26801–27972/M, ORF6, ORF7a, ORF7b, ORF8 [83] 6954–10870/ORF1ab [83] 22775–22778/S [100] | - | [83,100] |
| Various lineages and Omicron (VOC) | 13296–15240/ORF1ab [101] 20055–21618/ORF1ab, S [102] | - | [101,102] |
| Omicron parents | 21593–23118/S [103] | - | [98,103] |
| Not involving VOCs | Inside the gene S [104] 22775–22778/S [105] 19408–19411/ORF1ab [105] | - | [76,104,105,106,107,108,109] |
| Rate of Evolution (Substitutions/Site/Year) | Statistical Confidence (95%) | References |
|---|---|---|
| 7.31 × 10−4 | 5.95 × 10−4–8.68 × 10−4 (CI) | [11,110] |
| 1.1 × 10−3 | 7.03 × 10−4–1.5 × 10−3 (CI) | [111] |
| 8 × 10−4 | – | [112] |
| 1 × 10−4–1.4 × 10−3 | – | [113] |
| 7.8 × 10−4 | 1.1 × 10−4–1.5 × 10−3 (HPDI) | [114] |
| 9.9 × 10−4 | 6.29 × 10−4–1.35 × 10−3 (BCI) | [81] |
| 7.9 × 10−4 | 6.64 × 10−4–9.27 × 10−4 (HPDI) | [115] |
| 3.547 × 10−4 | 1.112 × 10−4–5.969 × 10−4 (CI) | [116] |
| 6.5 × 10−3 | 4.9 × 10−3–8.0 × 10−3 (HPDI) | [117] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Vázquez, L.D.; Arenas, M. Molecular Evolution of SARS-CoV-2 during the COVID-19 Pandemic. Genes 2023, 14, 407. https://doi.org/10.3390/genes14020407
González-Vázquez LD, Arenas M. Molecular Evolution of SARS-CoV-2 during the COVID-19 Pandemic. Genes. 2023; 14(2):407. https://doi.org/10.3390/genes14020407
Chicago/Turabian StyleGonzález-Vázquez, Luis Daniel, and Miguel Arenas. 2023. "Molecular Evolution of SARS-CoV-2 during the COVID-19 Pandemic" Genes 14, no. 2: 407. https://doi.org/10.3390/genes14020407
APA StyleGonzález-Vázquez, L. D., & Arenas, M. (2023). Molecular Evolution of SARS-CoV-2 during the COVID-19 Pandemic. Genes, 14(2), 407. https://doi.org/10.3390/genes14020407