

Transcriptional Profiling of Rat Prefrontal Cortex after Acute Inescapable Footshock Stress

 , , , , , , and
, , , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Footshock (FS) Stress Procedure
2.3. RNA Extraction and Purification
2.4. Microarray Procedures
2.5. Microarray Expression Analysis
2.6. Western Blotting: Tissue Processing and Image Analysis
2.7. Statistical Analysis
3. Results
3.1. Gene Expression Analysis in the PFC Immediately after Footshock Stress
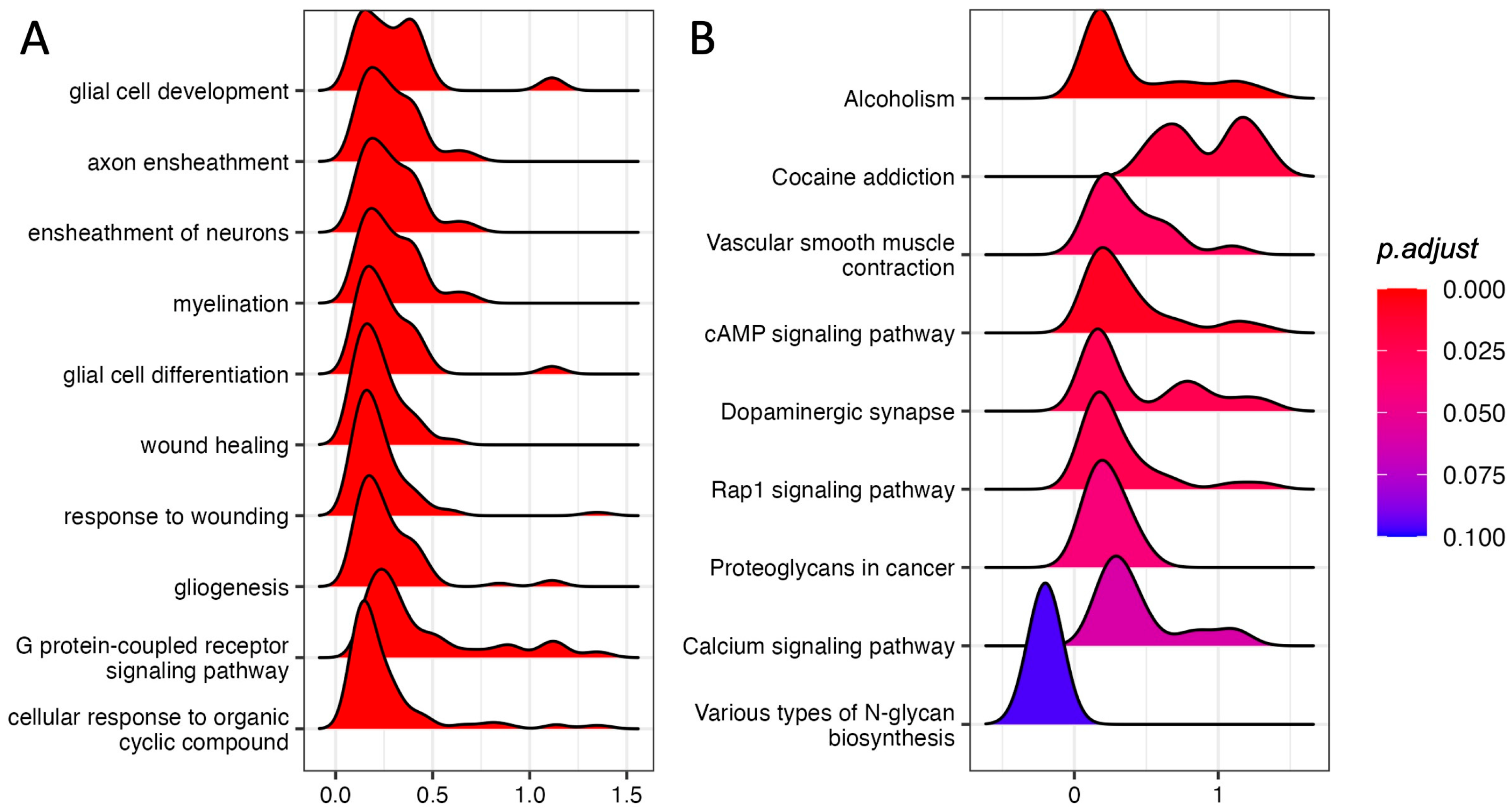
3.2. Gene Set Enrichment Analysis in the PFC of Rats Immediately after Footshock Stress
3.3. Gene Expression Analysis and Gene Set Enrichment Analysis (GSEA) in the PFC of Rats 2 and 24 h after Acute Footshock Stress
3.4. Transcriptional Factor Gene Target Analysis
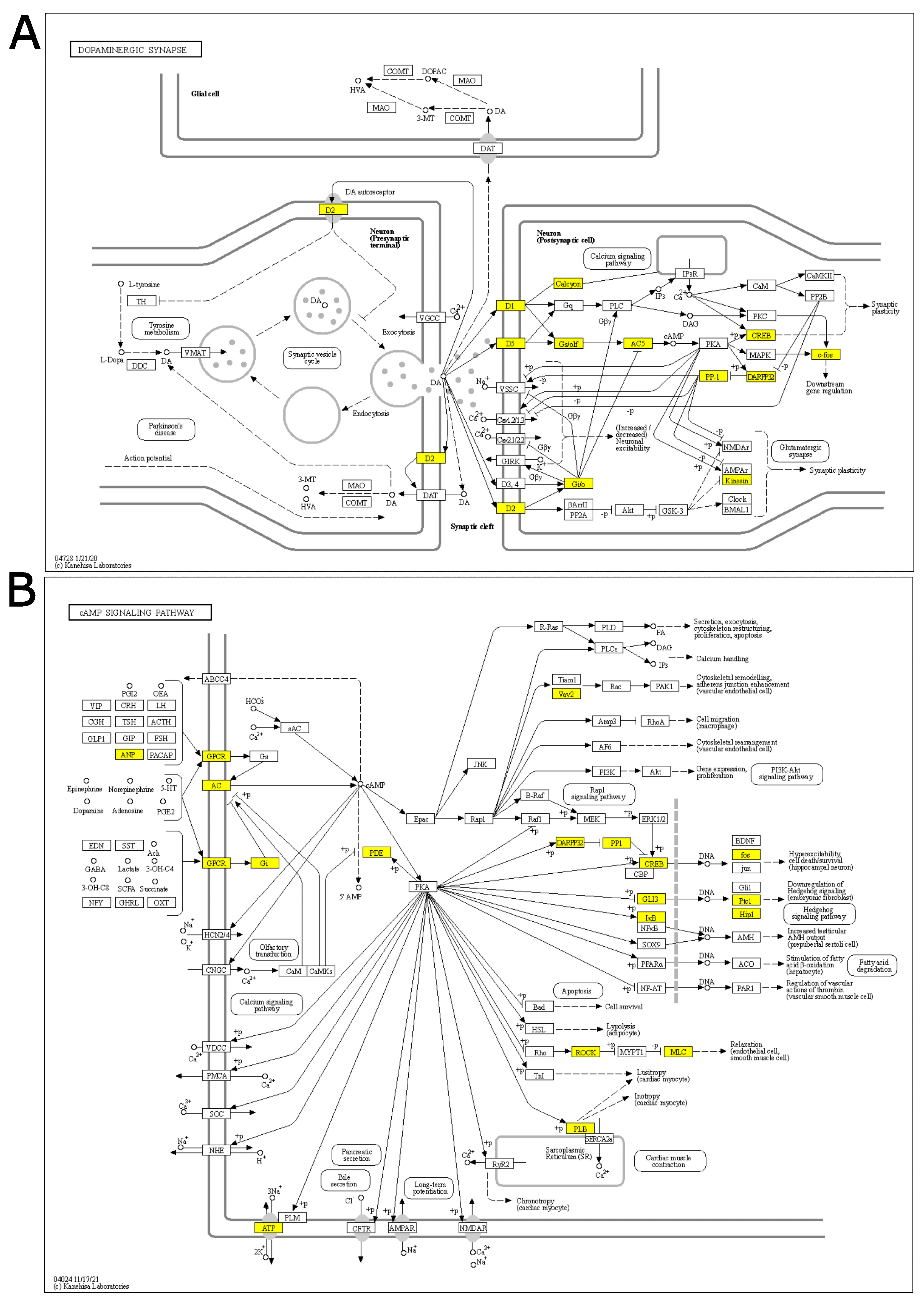
3.5. Protein Expression Analysis of Stress Response Key Effectors in the PFC of Rats Immediately after Acute FS Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanacora, G.; Yan, Z.; Popoli, M. The Stressed Synapse 2.0: Pathophysiological Mechanisms in Stress-Related Neuropsychiatric Disorders. Nat. Rev. Neurosci. 2021, 23, 86–103. [Google Scholar] [CrossRef] [PubMed]
- Popoli, M.; Yan, Z.; McEwen, B.S.; Sanacora, G. The Stressed Synapse: The Impact of Stress and Glucocorticoids on Glutamate Transmission. Nat. Rev. Neurosci. 2011, 13, 22–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musazzi, L.; Tornese, P.; Sala, N.; Popoli, M. Acute Stress Is Not Acute: Sustained Enhancement of Glutamate Release after Acute Stress Involves Readily Releasable Pool Size and Synapsin I Activation. Mol. Psychiatry 2017, 22, 1226–1227. [Google Scholar] [CrossRef] [PubMed]
- Musazzi, L.; Milanese, M.; Farisello, P.; Zappettini, S.; Tardito, D.; Barbiero, V.S.; Bonifacino, T.; Mallei, A.; Baldelli, P.; Racagni, G.; et al. Acute Stress Increases Depolarization-Evoked Glutamate Release in the Rat Prefrontal/Frontal Cortex: The Dampening Action of Antidepressants. PLoS ONE 2010, 5, e8566. [Google Scholar] [CrossRef]
- Treccani, G.; Musazzi, L.; Perego, C.; Milanese, M.; Nava, N.; Bonifacino, T.; Lamanna, J.; Malgaroli, A.; Drago, F.; Racagni, G.; et al. Acute Stress Rapidly Increases the Readily Releasable Pool of Glutamate Vesicles in Prefrontal and Frontal Cortex through Non-Genomic Action of Corticosterone. Mol. Psychiatry 2014, 19, 401. [Google Scholar] [CrossRef]
- Arnsten, A.F.T.; Wang, M.J.; Paspalas, C.D. Neuromodulation of Thought: Flexibilities and Vulnerabilities in Prefrontal Cortical Network Synapses. Neuron 2012, 76, 223–239. [Google Scholar] [CrossRef] [Green Version]
- Musazzi, L.; Tornese, P.; Sala, N.; Popoli, M. Acute or Chronic? A Stressful Question. Trends Neurosci. 2017, 40, 525–535. [Google Scholar] [CrossRef]
- Musazzi, L.; Tornese, P.; Sala, N.; Popoli, M. What Acute Stress Protocols Can Tell Us About PTSD and Stress-Related Neuropsychiatric Disorders. Front. Pharmacol. 2018, 9, 758. [Google Scholar] [CrossRef] [Green Version]
- Musazzi, L.; Treccani, G.; Popoli, M. Functional and Structural Remodeling of Glutamate Synapses in Prefrontal and Frontal Cortex Induced by Behavioral Stress. Front. Psychiatry 2015, 6, 60. [Google Scholar] [CrossRef]
- Bonini, D.; Mora, C.; Tornese, P.; Sala, N.; Filippini, A.; La Via, L.; Milanese, M.; Calza, S.; Bonanno, G.; Racagni, G.; et al. Acute Footshock Stress Induces Time-Dependent Modifications of AMPA/NMDA Protein Expression and AMPA Phosphorylation. Neural Plast. 2016, 2016, 7267865. [Google Scholar] [CrossRef] [Green Version]
- Nava, N.; Treccani, G.; Müller, H.K.; Popoli, M.; Wegener, G.; Elfving, B. The Expression of Plasticity-Related Genes in an Acute Model of Stress Is Modulated by Chronic Desipramine in a Time-Dependent Manner within Medial Prefrontal Cortex. Eur. Neuropsychopharmacol. 2017, 27, 19–28. [Google Scholar] [CrossRef]
- Musazzi, L.; Sala, N.; Tornese, P.; Gallivanone, F.; Belloli, S.; Conte, A.; Di Grigoli, G.; Chen, F.; Ikinci, A.; Treccani, G.; et al. Acute Inescapable Stress Rapidly Increases Synaptic Energy Metabolism in Prefrontal Cortex and Alters Working Memory Performance. Cereb. Cortex 2019, 29, 4948–4957. [Google Scholar] [CrossRef]
- Sala, N.; Paoli, C.; Bonifacino, T.; Mingardi, J.; Schiavon, E.; La Via, L.; Milanese, M.; Tornese, P.; Datusalia, A.K.; Rosa, J.; et al. Acute Ketamine Facilitates Fear Memory Extinction in a Rat Model of PTSD Along With Restoring Glutamatergic Alterations and Dendritic Atrophy in the Prefrontal Cortex. Front. Pharmacol. 2022, 13, 759626. [Google Scholar] [CrossRef]
- Izzi, C.; Barbon, A.; Kretz, R.; Sander, T.; Barlati, S. Sequencing of the GRIK1 Gene in Patients with Juvenile Absence Epilepsy Does Not Reveal Mutations Affecting Receptor Structure. Am. J. Med. Genet. 2002, 114, 354–359. [Google Scholar] [CrossRef]
- Cattane, N.; Minelli, A.; Milanesi, E.; Maj, C.; Bignotti, S.; Bortolomasi, M.; Chiavetto, L.B.; Gennarelli, M. Altered Gene Expression in Schizophrenia: Findings from Transcriptional Signatures in Fibroblasts and Blood. PLoS ONE 2015, 10, e0116686. [Google Scholar] [CrossRef]
- Carvalho, B.S.; Irizarry, R.A. A Framework for Oligonucleotide Microarray Preprocessing. Bioinformatics 2010, 26, 2363–2367. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. ClusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Sales, G.; Calura, E.; Cavalieri, D.; Romualdi, C. Graphite—A Bioconductor Package to Convert Pathway Topology to Gene Network. BMC Bioinform. 2012, 13, 20. [Google Scholar] [CrossRef] [Green Version]
- Sales, G.; Calura, E.; Romualdi, C. metaGraphite—A New Layer of Pathway Annotation to Get Metabolite Networks. Bioinformatics 2019, 35, 1258–1260. [Google Scholar] [CrossRef]
- Liberzon, A.; Subramanian, A.; Pinchback, R.; Thorvaldsdottir, H.; Tamayo, P.; Mesirov, J.P. Molecular Signatures Database (MSigDB) 3.0. Bioinformatics 2011, 27, 1739–1740. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene Set Variation Analysis for Microarray and RNA-Seq Data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tornese, P.; Sala, N.; Bonini, D.; Bonifacino, T.; La Via, L.; Milanese, M.; Treccani, G.; Seguini, M.; Ieraci, A.; Mingardi, J.; et al. Chronic Mild Stress Induces Anhedonic Behavior and Changes in Glutamate Release, BDNF Trafficking and Dendrite Morphology Only in Stress Vulnerable Rats. The Rapid Restorative Action of Ketamine. Neurobiol. Stress 2019, 10, 100160. [Google Scholar] [CrossRef]
- Hoerndli, F.J.; Wang, R.; Mellem, J.E.; Kallarackal, A.; Brockie, P.J.; Thacker, C.; Madsen, D.M.; Maricq, A.V. Neuronal Activity and CaMKII Regulate Kinesin-Mediated Transport of Synaptic AMPARs. Neuron 2015, 86, 457–474. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Fok, A.H.K.; Fan, R.; Kwan, P.-Y.; Chan, H.-L.; Lo, L.H.-Y.; Chan, Y.-S.; Yung, W.-H.; Huang, J.; Lai, C.S.W.; et al. Specific Depletion of the Motor Protein KIF5B Leads to Deficits in Dendritic Transport, Synaptic Plasticity and Memory. eLife 2020, 9, e53456. [Google Scholar] [CrossRef]
- Yao, W.; Cao, Q.; Luo, S.; He, L.; Yang, C.; Chen, J.; Qi, Q.; Hashimoto, K.; Zhang, J.-C. Microglial ERK-NRBP1-CREB-BDNF Signaling in Sustained Antidepressant Actions of (R)-Ketamine. Mol. Psychiatry 2022, 27, 1618–1629. [Google Scholar] [CrossRef]
- Meijer, O.C.; Buurstede, J.C.; Viho, E.M.G.; Amaya, J.M.; Koning, A.-S.C.A.M.; van der Meulen, M.; van Weert, L.T.C.M.; Paul, S.N.; Kroon, J.; Koorneef, L.L. Transcriptional Glucocorticoid Effects in the Brain: Finding the Relevant Target Genes. J. Neuroendocrinol. 2022, e13213. [Google Scholar] [CrossRef]
- Floriou-Servou, A.; von Ziegler, L.; Waag, R.; Schläppi, C.; Germain, P.-L.; Bohacek, J. The Acute Stress Response in the Multiomic Era. Biol. Psychiatry 2021, 89, 1116–1126. [Google Scholar] [CrossRef]
- Girgenti, M.J.; Duman, R.S. Transcriptome Alterations in Posttraumatic Stress Disorder. Biol. Psychiatry 2018, 83, 840–848. [Google Scholar] [CrossRef]
- Roszkowski, M.; Manuella, F.; von Ziegler, L.; Durán-Pacheco, G.; Moreau, J.-L.; Mansuy, I.M.; Bohacek, J. Rapid Stress-Induced Transcriptomic Changes in the Brain Depend on β-Adrenergic Signaling. Neuropharmacology 2016, 107, 329–338. [Google Scholar] [CrossRef]
- Floriou-Servou, A.; von Ziegler, L.; Stalder, L.; Sturman, O.; Privitera, M.; Rassi, A.; Cremonesi, A.; Thöny, B.; Bohacek, J. Distinct Proteomic, Transcriptomic, and Epigenetic Stress Responses in Dorsal and Ventral Hippocampus. Biol. Psychiatry 2018, 84, 531–541. [Google Scholar] [CrossRef] [Green Version]
- Stankiewicz, A.M.; Goscik, J.; Majewska, A.; Swiergiel, A.H.; Juszczak, G.R. The Effect of Acute and Chronic Social Stress on the Hippocampal Transcriptome in Mice. PLoS ONE 2015, 10, e0142195. [Google Scholar] [CrossRef]
- Sillivan, S.E.; Jones, M.E.; Jamieson, S.; Rumbaugh, G.; Miller, C.A. Bioinformatic Analysis of Long-Lasting Transcriptional and Translational Changes in the Basolateral Amygdala Following Acute Stress. PLoS ONE 2019, 14, e0209846. [Google Scholar] [CrossRef] [Green Version]
- Hohoff, C.; Gorji, A.; Kaiser, S.; Willscher, E.; Korsching, E.; Ambrée, O.; Arolt, V.; Lesch, K.-P.; Sachser, N.; Deckert, J.; et al. Effect of Acute Stressor and Serotonin Transporter Genotype on Amygdala First Wave Transcriptome in Mice. PLoS ONE 2013, 8, e58880. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Xiang, X.-H.; He, F.; Lin, L.-B.; Zhang, R.; Ping, X.-J.; Han, J.-S.; Guo, N.; Zhang, Q.-H.; Cui, C.-L.; et al. Transcriptome Profiling Analysis Reveals Region-Distinctive Changes of Gene Expression in the CNS in Response to Different Moderate Restraint Stress. J. Neurochem. 2010, 113, 1436–1446. [Google Scholar] [CrossRef] [PubMed]
- Nahvi, R.J.; Tanelian, A.; Nwokafor, C.; Godino, A.; Parise, E.; Estill, M.; Shen, L.; Nestler, E.J.; Sabban, E.L. Transcriptome Profiles Associated with Resilience and Susceptibility to Single Prolonged Stress in the Locus Coeruleus and Nucleus Accumbens in Male Sprague-Dawley Rats. Behav. Brain Res. 2023, 439, 114162. [Google Scholar] [CrossRef]
- Hodes, G.E.; Pfau, M.L.; Purushothaman, I.; Ahn, H.F.; Golden, S.A.; Christoffel, D.J.; Magida, J.; Brancato, A.; Takahashi, A.; Flanigan, M.E.; et al. Sex Differences in Nucleus Accumbens Transcriptome Profiles Associated with Susceptibility versus Resilience to Subchronic Variable Stress. J. Neurosci. 2015, 35, 16362–16376. [Google Scholar] [CrossRef] [Green Version]
- Carter, S.D.; Mifsud, K.R.; Reul, J.M.H.M. Acute Stress Enhances Epigenetic Modifications But Does Not Affect the Constitutive Binding of PCREB to Immediate-Early Gene Promoters in the Rat Hippocampus. Front. Mol. Neurosci. 2017, 10, 416. [Google Scholar] [CrossRef]
- Kennedy, C.L.M.; Carter, S.D.; Mifsud, K.R.; Reul, J.M.H.M. Unexpected Effects of Metyrapone on Corticosteroid Receptor Interaction with the Genome and Subsequent Gene Transcription in the Hippocampus of Male Rats. J. Neuroendocrinol. 2020, 32, e12820. [Google Scholar] [CrossRef]
- Musazzi, L.; Tornese, P.; Sala, N.; Lee, F.S.; Popoli, M.; Ieraci, A. Acute Stress Induces an Aberrant Increase of Presynaptic Release of Glutamate and Cellular Activation in the Hippocampus of BDNFVal/Met Mice. J. Cell. Physiol. 2022, 237, 3834–3844. [Google Scholar] [CrossRef] [PubMed]
- Kovács, L.Á.; Schiessl, J.A.; Nafz, A.E.; Csernus, V.; Gaszner, B. Both Basal and Acute Restraint Stress-Induced c-Fos Expression Is Influenced by Age in the Extended Amygdala and Brainstem Stress Centers in Male Rats. Front. Aging Neurosci. 2018, 10, 248. [Google Scholar] [CrossRef] [PubMed]
- Marrocco, J.; Petty, G.H.; Ríos, M.B.; Gray, J.D.; Kogan, J.F.; Waters, E.M.; Schmidt, E.F.; Lee, F.S.; McEwen, B.S. A Sexually Dimorphic Pre-Stressed Translational Signature in CA3 Pyramidal Neurons of BDNF Val66Met Mice. Nat. Commun. 2017, 8, 808. [Google Scholar] [CrossRef] [Green Version]
- Häusl, A.S.; Brix, L.M.; Hartmann, J.; Pöhlmann, M.L.; Lopez, J.-P.; Menegaz, D.; Brivio, E.; Engelhardt, C.; Roeh, S.; Bajaj, T.; et al. The Co-Chaperone Fkbp5 Shapes the Acute Stress Response in the Paraventricular Nucleus of the Hypothalamus of Male Mice. Mol. Psychiatry 2021, 26, 3060–3076. [Google Scholar] [CrossRef]
- Flati, T.; Gioiosa, S.; Chillemi, G.; Mele, A.; Oliverio, A.; Mannironi, C.; Rinaldi, A.; Castrignanò, T. A Gene Expression Atlas for Different Kinds of Stress in the Mouse Brain. Sci. Data 2020, 7, 437. [Google Scholar] [CrossRef]
- Von Ziegler, L.M.; Floriou-Servou, A.; Waag, R.; Das Gupta, R.R.; Sturman, O.; Gapp, K.; Maat, C.A.; Kockmann, T.; Lin, H.-Y.; Duss, S.N.; et al. Multiomic Profiling of the Acute Stress Response in the Mouse Hippocampus. Nat. Commun. 2022, 13, 1824. [Google Scholar] [CrossRef]
- Zhang, X.; Alnafisah, R.S.; Hamoud, A.-R.A.; Shukla, R.; McCullumsmith, R.E.; O’Donovan, S.M. Astrocytes in Neuropsychiatric Disorders: A Review of Postmortem Evidence. Adv. Neurobiol. 2021, 26, 153–172. [Google Scholar] [CrossRef]
- Scuderi, C.; Verkhratsky, A.; Parpura, V.; Li, B. Neuroglia in Psychiatric Disorders. Adv. Neurobiol. 2021, 26, 3–19. [Google Scholar] [CrossRef]
- Weber, M.D.; McKim, D.B.; Niraula, A.; Witcher, K.G.; Yin, W.; Sobol, C.G.; Wang, Y.; Sawicki, C.M.; Sheridan, J.F.; Godbout, J.P. The Influence of Microglial Elimination and Repopulation on Stress Sensitization Induced by Repeated Social Defeat. Biol. Psychiatry 2019, 85, 667–678. [Google Scholar] [CrossRef]
- Saur, L.; Baptista, P.P.A.; Bagatini, P.B.; Neves, L.T.; de Oliveira, R.M.; Vaz, S.P.; Ferreira, K.; Machado, S.A.; Mestriner, R.G.; Xavier, L.L. Experimental Post-Traumatic Stress Disorder Decreases Astrocyte Density and Changes Astrocytic Polarity in the CA1 Hippocampus of Male Rats. Neurochem. Res. 2016, 41, 892–904. [Google Scholar] [CrossRef]
- Han, F.; Xiao, B.; Wen, L. Loss of Glial Cells of the Hippocampus in a Rat Model of Post-Traumatic Stress Disorder. Neurochem. Res. 2015, 40, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Gryz, M.; Lehner, M.; Wisłowska-Stanek, A.; Płaźnik, A. Dopaminergic System Activity under Stress Condition—Seeking Individual Differences, Preclinical Studies. Psychiatr. Pol. 2018, 52, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Caffino, L.; Calabrese, F.; Giannotti, G.; Barbon, A.; Verheij, M.M.M.; Racagni, G.; Fumagalli, F. Stress Rapidly Dysregulates the Glutamatergic Synapse in the Prefrontal Cortex of Cocaine-Withdrawn Adolescent Rats. Addict. Biol. 2015, 20, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Caffino, L.; Mottarlini, F.; Mingardi, J.; Zita, G.; Barbon, A.; Fumagalli, F. Anhedonic-like Behavior and BDNF Dysregulation Following a Single Injection of Cocaine during Adolescence. Neuropharmacology 2020, 175, 108161. [Google Scholar] [CrossRef]
- Plattner, F.; Hayashi, K.; Hernández, A.; Benavides, D.R.; Tassin, T.C.; Tan, C.; Day, J.; Fina, M.W.; Yuen, E.Y.; Yan, Z.; et al. The Role of Ventral Striatal CAMP Signaling in Stress-Induced Behaviors. Nat. Neurosci. 2015, 18, 1094–1100. [Google Scholar] [CrossRef] [Green Version]
- Conroy, L.R.; Hawkinson, T.R.; Young, L.E.A.; Gentry, M.S.; Sun, R.C. Emerging Roles of N-Linked Glycosylation in Brain Physiology and Disorders. Trends Endocrinol. Metab. 2021, 32, 980–993. [Google Scholar] [CrossRef]
- Brachet, A.; Lario, A.; Fernández-Rodrigo, A.; Heisler, F.F.; Gutiérrez, Y.; Lobo, C.; Kneussel, M.; Esteban, J.A. A Kinesin 1-Protrudin Complex Mediates AMPA Receptor Synaptic Removal during Long-Term Depression. Cell Rep. 2021, 36, 109499. [Google Scholar] [CrossRef]
- Sarantis, K.; Matsokis, N.; Angelatou, F. Synergistic Interactions of Dopamine D1 and Glutamate NMDA Receptors in Rat Hippocampus and Prefrontal Cortex: Involvement of ERK1/2 Signaling. Neuroscience 2009, 163, 1135–1145. [Google Scholar] [CrossRef]
- Kim, J.-E.; Lee, D.-S.; Kim, T.-H.; Park, H.; Kim, M.-J.; Kang, T.-C. PLPP/CIN-Mediated DARPP-32 Serine 97 Dephosphorylation Delays the Seizure Onset in Response to Kainic Acid in the Mouse Hippocampus. Neuropharmacology 2022, 219, 109238. [Google Scholar] [CrossRef]
- de Carvalho, C.R.; Lopes, M.W.; Constantino, L.C.; Hoeller, A.A.; de Melo, H.M.; Guarnieri, R.; Linhares, M.N.; Bortolotto, Z.A.; Prediger, R.D.; Latini, A.; et al. The ERK Phosphorylation Levels in the Amygdala Predict Anxiety Symptoms in Humans and MEK/ERK Inhibition Dissociates Innate and Learned Defensive Behaviors in Rats. Mol. Psychiatry 2021, 26, 7257–7269. [Google Scholar] [CrossRef]
- Elhussiny, M.E.A.; Carini, G.; Mingardi, J.; Tornese, P.; Sala, N.; Bono, F.; Fiorentini, C.; La Via, L.; Popoli, M.; Musazzi, L.; et al. Modulation by Chronic Stress and Ketamine of Ionotropic AMPA/NMDA and Metabotropic Glutamate Receptors in the Rat Hippocampus. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 104, 110033. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.-Y.; Wang, Y.; Zou, H.-W.; Li, Z.-L.; Liu, Y.-J.; Li, L.-F. MGlu2/3 Receptor in the Prelimbic Cortex Is Implicated in Stress Resilience and Vulnerability in Mice. Eur. J. Pharmacol. 2021, 906, 174231. [Google Scholar] [CrossRef]
- Highland, J.N.; Zanos, P.; Georgiou, P.; Gould, T.D. Group II Metabotropic Glutamate Receptor Blockade Promotes Stress Resilience in Mice. Neuropsychopharmacology 2019, 44, 1788–1796. [Google Scholar] [CrossRef]
- Nasca, C.; Bigio, B.; Zelli, D.; de Angelis, P.; Lau, T.; Okamoto, M.; Soya, H.; Ni, J.; Brichta, L.; Greengard, P.; et al. Role of the Astroglial Glutamate Exchanger XCT in Ventral Hippocampus in Resilience to Stress. Neuron 2017, 96, 402–413.e5. [Google Scholar] [CrossRef] [Green Version]
- Nasca, C.; Zelli, D.; Bigio, B.; Piccinin, S.; Scaccianoce, S.; Nisticò, R.; McEwen, B.S. Stress Dynamically Regulates Behavior and Glutamatergic Gene Expression in Hippocampus by Opening a Window of Epigenetic Plasticity. Proc. Natl. Acad. Sci. USA 2015, 112, 14960–14965. [Google Scholar] [CrossRef] [Green Version]
- Nasca, C.; Xenos, D.; Barone, Y.; Caruso, A.; Scaccianoce, S.; Matrisciano, F.; Battaglia, G.; Mathé, A.A.; Pittaluga, A.; Lionetto, L.; et al. L-Acetylcarnitine Causes Rapid Antidepressant Effects through the Epigenetic Induction of MGlu2 Receptors. Proc. Natl. Acad. Sci. USA 2013, 110, 4804–4809. [Google Scholar] [CrossRef] [Green Version]
- Musazzi, L. Targeting Metabotropic Glutamate Receptors for Rapid-Acting Antidepressant Drug Discovery. Expert Opin. Drug Discov. 2021, 16, 147–157. [Google Scholar] [CrossRef]
- Chaki, S. MGlu2/3 Receptor as a Novel Target for Rapid Acting Antidepressants. In Advances in Pharmacology; Elsevier: Amsterdam, The Netherlands, 2020; Volume 89, pp. 289–309. ISBN 9780128201893. [Google Scholar]
- Lowes, D.C.; Harris, A.Z. Stressed and Wired: The Effects of Stress on the VTA Circuits Underlying Motivated Behavior. Curr. Opin. Endocr. Metab. Res. 2022, 26, 100388. [Google Scholar] [CrossRef]
- Vaessen, T.; Hernaus, D.; Myin-Germeys, I.; van Amelsvoort, T. The Dopaminergic Response to Acute Stress in Health and Psychopathology: A Systematic Review. Neurosci. Biobehav. Rev. 2015, 56, 241–251. [Google Scholar] [CrossRef]
- MacDougall, M.J.; Howland, J.G. Acute Stress, but Not Corticosterone, Disrupts Short- and Long-Term Synaptic Plasticity in Rat Dorsal Subiculum via Glucocorticoid Receptor Activation. Cereb. Cortex 2013, 23, 2611–2619. [Google Scholar] [CrossRef] [Green Version]
- McKlveen, J.M.; Myers, B.; Flak, J.N.; Bundzikova, J.; Solomon, M.B.; Seroogy, K.B.; Herman, J.P. Role of Prefrontal Cortex Glucocorticoid Receptors in Stress and Emotion. Biol. Psychiatry 2013, 74, 672–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramanan, N.; Shen, Y.; Sarsfield, S.; Lemberger, T.; Schütz, G.; Linden, D.J.; Ginty, D.D. SRF Mediates Activity-Induced Gene Expression and Synaptic Plasticity but Not Neuronal Viability. Nat. Neurosci. 2005, 8, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Vialou, V.; Maze, I.; Renthal, W.; LaPlant, Q.C.; Watts, E.L.; Mouzon, E.; Ghose, S.; Tamminga, C.A.; Nestler, E.J. Serum Response Factor Promotes Resilience to Chronic Social Stress through the Induction of DeltaFosB. J. Neurosci. 2010, 30, 14585–14592. [Google Scholar] [CrossRef] [Green Version]
- Zimprich, A.; Mroz, G.; Meyer Zu Reckendorf, C.; Anastasiadou, S.; Förstner, P.; Garrett, L.; Hölter, S.M.; Becker, L.; Rozman, J.; Prehn, C.; et al. Serum Response Factor (SRF) Ablation Interferes with Acute Stress-Associated Immediate and Long-Term Coping Mechanisms. Mol. Neurobiol. 2017, 54, 8242–8262. [Google Scholar] [CrossRef]
- Wang, Z.-H.; Xia, Y.; Wu, Z.; Kang, S.S.; Zhang, J.; Liu, P.; Liu, X.; Song, W.; Huin, V.; Dhaenens, C.-M.; et al. Neuronal ApoE4 Stimulates C/EBPβ Activation, Promoting Alzheimer’s Disease Pathology in a Mouse Model. Prog. Neurobiol. 2022, 209, 102212. [Google Scholar] [CrossRef]
- Aghanoori, M.-R.; Agarwal, P.; Gauvin, E.; Nagalingam, R.S.; Bonomo, R.; Yathindranath, V.; Smith, D.R.; Hai, Y.; Lee, S.; Jolivalt, C.G.; et al. CEBPβ Regulation of Endogenous IGF-1 in Adult Sensory Neurons Can Be Mobilized to Overcome Diabetes-Induced Deficits in Bioenergetics and Axonal Outgrowth. Cell. Mol. Life Sci. 2022, 79, 193. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martini, P.; Mingardi, J.; Carini, G.; Mattevi, S.; Ndoj, E.; La Via, L.; Magri, C.; Gennarelli, M.; Russo, I.; Popoli, M.; et al. Transcriptional Profiling of Rat Prefrontal Cortex after Acute Inescapable Footshock Stress. Genes 2023, 14, 740. https://doi.org/10.3390/genes14030740
Martini P, Mingardi J, Carini G, Mattevi S, Ndoj E, La Via L, Magri C, Gennarelli M, Russo I, Popoli M, et al. Transcriptional Profiling of Rat Prefrontal Cortex after Acute Inescapable Footshock Stress. Genes. 2023; 14(3):740. https://doi.org/10.3390/genes14030740
Chicago/Turabian StyleMartini, Paolo, Jessica Mingardi, Giulia Carini, Stefania Mattevi, Elona Ndoj, Luca La Via, Chiara Magri, Massimo Gennarelli, Isabella Russo, Maurizio Popoli, and et al. 2023. "Transcriptional Profiling of Rat Prefrontal Cortex after Acute Inescapable Footshock Stress" Genes 14, no. 3: 740. https://doi.org/10.3390/genes14030740
APA StyleMartini, P., Mingardi, J., Carini, G., Mattevi, S., Ndoj, E., La Via, L., Magri, C., Gennarelli, M., Russo, I., Popoli, M., Musazzi, L., & Barbon, A. (2023). Transcriptional Profiling of Rat Prefrontal Cortex after Acute Inescapable Footshock Stress. Genes, 14(3), 740. https://doi.org/10.3390/genes14030740