
Gene-by-Sex Interactions: Genome-Wide Association Study Reveals Five SNPs Associated with Obesity and Overweight in a Male Population

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. DNA Extraction
2.3. Genotyping
2.4. Quality Control (QC)
2.5. Association Analysis and In Silico Functional Annotation
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, K.B.; Smith, M.S. Obesity Statistics. Prim. Care Clin. Off. Pract. 2016, 43, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Caballero, B. Humans against Obesity: Who Will Win? Adv. Nutr. 2019, 10, S4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, Regional, and National Prevalence of Overweight and Obesity in Children and Adults during 1980–2013: A Systematic Analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [Green Version]
- Cawley, J.; Meyerhoefer, C. The Medical Care Costs of Obesity: An Instrumental Variables Approach. J. Health Econ. 2012, 31, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Nicolaidis, S. Environment and Obesity. Metabolism 2019, 100S, 153942. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, M.O. Genetics of Obesity: What Genetic Association Studies Have Taught Us about the Biology of Obesity and Its Complications. Lancet Diabetes Endocrinol. 2018, 6, 223–236. [Google Scholar] [CrossRef]
- Speakman, J.R.; Loos, R.J.F.; O’Rahilly, S.; Hirschhorn, J.N.; Allison, D.B. GWAS for BMI: A Treasure Trove of Fundamental Insights into the Genetic Basis of Obesity. Int. J. Obes. 2018, 42, 1524–1531. [Google Scholar] [CrossRef]
- Müller, M.J.; Geisler, C.; Blundell, J.; Dulloo, A.; Schutz, Y.; Krawczak, M.; Bosy-Westphal, A.; Enderle, J.; Heymsfield, S.B. The Case of GWAS of Obesity: Does Body Weight Control Play by the Rules? Int. J. Obes. 2018, 42, 1395–1405. [Google Scholar] [CrossRef] [PubMed]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic Studies of Body Mass Index Yield New Insights for Obesity Biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Bakshi, A.; Zhu, Z.; Hemani, G.; Vinkhuyzen, A.A.E.; Lee, S.H.; Robinson, M.R.; Perry, J.R.B.; Nolte, I.M.; Van Vliet-Ostaptchouk, J.V.; et al. Genetic Variance Estimation with Imputed Variants Finds Negligible Missing Heritability for Human Height and Body Mass Index. Nat. Genet. 2015, 47, 1114–1120. [Google Scholar] [CrossRef] [Green Version]
- Williams, T.M.; Carroll, S.B. Genetic and Molecular Insights into the Development and Evolution of Sexual Dimorphism. Nat. Rev. Genet. 2009, 10, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.C.K. Sexual Dimorphism of Body Composition. Best Pract. Res. Clin. Endocrinol. Metab. 2007, 21, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.W.; Grant, A.M.; Williams, S.M.; Goulding, A. Sex Differences in Regional Body Fat Distribution from Pre- to Postpuberty. Obesity 2010, 18, 1410–1416. [Google Scholar] [CrossRef] [PubMed]
- Kuk, J.L.; Saunders, T.J.; Davidson, L.E.; Ross, R. Age-Related Changes in Total and Regional Fat Distribution. Ageing Res. Rev. 2009, 8, 339–348. [Google Scholar] [CrossRef]
- Pulit, S.L.; Karaderi, T.; Lindgren, C.M. Sexual Dimorphisms in Genetic Loci Linked to Body Fat Distribution. Biosci. Rep. 2017, 37, BSR20160184. [Google Scholar] [CrossRef]
- Winkler, T.W.; Justice, A.E.; Graff, M.; Barata, L.; Feitosa, M.F.; Chu, S.; Czajkowski, J.; Esko, T.; Fall, T.; Kilpeläinen, T.O.; et al. The Influence of Age and Sex on Genetic Associations with Adult Body Size and Shape: A Large-Scale Genome-Wide Interaction Study. PLOS Genet. 2015, 11, e1005378. [Google Scholar] [CrossRef] [Green Version]
- Weyrich, A. Preparation of Genomic DNA from Mammalian Sperm. Curr. Protoc. Mol. Biol. 2012, 1, 2.13.1–2.13.3. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Turner, S.D. Qqman: An R Package for Visualizing GWAS Results Using Q-Q and Manhattan Plots. J. Open Source Softw. 2018, 3, 731. [Google Scholar] [CrossRef] [Green Version]
- Howe, K.L.; Achuthan, P.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Ridwan Amode, M.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J.; et al. Ensembl 2021. Nucleic Acids Res. 2021, 49, D884–D891. [Google Scholar] [CrossRef]
- Oscanoa, J.; Sivapalan, L.; Gadaleta, E.; Dayem Ullah, A.Z.; Lemoine, N.R.; Chelala, C. SNPnexus: A Web Server for Functional Annotation of Human Genome Sequence Variation (2020 Update). Nucleic Acids Res. 2020, 48, W185–W192. [Google Scholar] [CrossRef] [PubMed]
- Lonsdale, J.; Thomas, J.; Salvatore, M.; Phillips, R.; Lo, E.; Shad, S.; Hasz, R.; Walters, G.; Garcia, F.; Young, N.; et al. The Genotype-Tissue Expression (GTEx) Project. Nat. Genet. 2013, 45, 580. [Google Scholar] [CrossRef] [PubMed]
- Auton, A.; Abecasis, G.R.; Altshuler, D.M.; Durbin, R.M.; Bentley, D.R.; Chakravarti, A.; Clark, A.G.; Donnelly, P.; Eichler, E.E.; Flicek, P.; et al. A Global Reference for Human Genetic Variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A Method and Server for Predicting Damaging Missense Mutations. Nat. Methods 2010, 7, 248–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaser, R.; Adusumalli, S.; Ngak Leng, S.; Sikic, M.; Ng, P.C. SIFT missense predictions for genomes. Nat. Protoc. 2015, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Boyle, A.P.; Hong, E.L.; Hariharan, M.; Cheng, Y.; Schaub, M.A.; Kasowski, M.; Karczewski, K.J.; Park, J.; Hitz, B.C.; Weng, S.; et al. Annotation of Functional Variation in Personal Genomes Using RegulomeDB. Genome Res. 2012, 22, 1790–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Quan, C.; Chen, H.; Bo, X.; Zhang, C. 3DSNP: A Database for Linking Human Noncoding SNPs to Their Three-Dimensional Interacting Genes. Nucleic Acids Res. 2017, 45, D643–D649. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.J.; Fu, X.; Xia, M.; Zhang, Q.; Gu, Z.; Guo, A.Y. MiRNASNP-v3: A Comprehensive Database for SNPs and Disease-Related Variations in MiRNAs and MiRNA Targets. Nucleic Acids Res. 2021, 49, D1276–D1281. [Google Scholar] [CrossRef]
- Svensson, P.A.; Jernås, M.; Sjöholm, K.; Hoffmann, J.M.; Nilsson, B.E.; Hansson, M.; Carlsson, L.M.S. Gene Expression in Human Brown Adipose Tissue. Int. J. Mol. Med. 2011, 27, 227–232. [Google Scholar] [CrossRef]
- Feliciangeli, S.; Chatelain, F.C.; Bichet, D.; Lesage, F. The Family of K2P Channels: Salient Structural and Functional Properties. J. Physiol. 2015, 593, 2587–2603. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zeng, X.; Huang, X.; Serag, S.; Woolf, C.J.; Spiegelman, B.M. Crosstalk Between Kcnk3-Mediated Ion Current and Adrenergic Signaling Regulates Adipose Thermogenesis and Obesity. Cell 2017, 171, 836. [Google Scholar] [CrossRef] [Green Version]
- Shungin, D.; Winkler, T.; Croteau-Chonka, D.C.; Ferreira, T.; Locke, A.E.; Mägi, R.; Strawbridge, R.J.; Pers, T.H.; Fischer, K.; Justice, A.E.; et al. New Genetic Loci Link Adipose and Insulin Biology to Body Fat Distribution. Nature 2015, 518, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Park, E.; Pan, Z.; Zhang, Z.; Lin, L.; Xing, Y. The Expanding Landscape of Alternative Splicing Variation in Human Populations. Am. J. Hum. Genet. 2018, 102, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Kwan, T.; Benovoy, D.; Dias, C.; Gurd, S.; Provencher, C.; Beaulieu, P.; Hudson, T.J.; Sladek, R.; Majewski, J. Genome-Wide Analysis of Transcript Isoform Variation in Humans. Nat. Genet. 2008, 40, 225–231. [Google Scholar] [CrossRef]
- Coulombe-Huntington, J.; Lam, K.C.L.; Dias, C.; Majewski, J. Fine-Scale Variation and Genetic Determinants of Alternative Splicing across Individuals. PLoS Genet. 2009, 5, e1000766. [Google Scholar] [CrossRef]
- Cooper, D.N. Functional Intronic Polymorphisms: Buried Treasure Awaiting Discovery within Our Genes. Hum. Genom. 2010, 4, 284. [Google Scholar] [CrossRef] [PubMed]
- Sauna, Z.E.; Kimchi-Sarfaty, C. Understanding the Contribution of Synonymous Mutations to Human Disease. Nat. Rev. Genet. 2011, 12, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Chamary, J.V.; Parmley, J.L.; Hurst, L.D. Hearing Silence: Non-Neutral Evolution at Synonymous Sites in Mammals. Nat. Rev. Genet. 2006, 7, 98–108. [Google Scholar] [CrossRef]
- Hunt, R.; Sauna, Z.E.; Ambudkar, S.V.; Gottesman, M.M.; Kimchi-Sarfaty, C. Silent (Synonymous) SNPs: Should We Care about Them? Methods Mol. Biol. 2009, 578, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Edwards, N.C.; Hing, Z.A.; Perry, A.; Blaisdell, A.; Kopelman, D.B.; Fathke, R.; Plum, W.; Newell, J.; Allen, C.E.; Geetha, S.; et al. Characterization of Coding Synonymous and Non-Synonymous Variants in ADAMTS13 Using Ex Vivo and In Silico Approaches. PLoS ONE 2012, 7, 38864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lekholm, E.; Perland, E.; Eriksson, M.M.; Hellsten, S.V.; Lindberg, F.A.; Rostami, J.; Fredriksson, R. Putative Membrane-Bound Transporters MFSD14A and MFSD14B Are Neuronal and Affected by Nutrient Availability. Front. Mol. Neurosci. 2017, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, S.; Martin, T.L.; Surwit, R.S.; Robidoux, J. Genetic Vulnerability to Diet-Induced Obesity in the C57BL/6J Mouse: Physiological and Molecular Characteristics. Physiol. Behav. 2004, 81, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Ceder, M.M.; Lekholm, E.; Klaesson, A.; Tripathi, R.; Schweizer, N.; Weldai, L.; Patil, S.; Fredriksson, R. Glucose Availability Alters Gene and Protein Expression of Several Newly Classified and Putative Solute Carriers in Mice Cortex Cell Culture and D. Melanogaster. Front. Cell Dev. Biol. 2020, 8, 579. [Google Scholar] [CrossRef] [PubMed]
- Rey, F.; Urrata, V.; Gilardini, L.; Bertoli, S.; Calcaterra, V.; Zuccotti, G.V.; Cancello, R.; Carelli, S. Role of Long Non-coding RNAs in Adipogenesis: State of the Art and Implications in Obesity and Obesity-associated Diseases. Obes. Rev. 2021, 22, e13203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Xu, S.; Liu, J.; Xie, Y.; Xiaobo, S. Long Noncoding RNAs: Novel Important Players in Adipocyte Lipid Metabolism and Derivative Diseases. Front. Physiol. 2021, 12, 840. [Google Scholar] [CrossRef]
- Wijesinghe, S.N.; Nicholson, T.; Tsintzas, K.; Jones, S.W. Involvements of Long Noncoding RNAs in Obesity-Associated Inflammatory Diseases. Obes. Rev. 2021, 22, e13156. [Google Scholar] [CrossRef]
- Nakano, T.; Goto, K. Diacylglycerol Kinase ε in Adipose Tissues: A Crosstalk Between Signal Transduction and Energy Metabolism. Front. Physiol. 2022, 13, 49. [Google Scholar] [CrossRef]
- Itani, S.I.; Ruderman, N.B.; Schmieder, F.; Boden, G. Lipid-Induced Insulin Resistance in Human Muscle Is Associated with Changes in Diacylglycerol, Protein Kinase C, and IkappaB-α. Diabetes 2002, 51, 2005–2011. [Google Scholar] [CrossRef] [Green Version]
- Kanoh, H.; Yamada, K.; Sakane, F. Diacylglycerol Kinases: Emerging Downstream Regulators in Cell Signaling Systems. J. Biochem. 2002, 131, 629–633. [Google Scholar] [CrossRef]
- Jiang, L.Q.; de Castro Barbosa, T.; Massart, J.; Deshmukh, A.S.; Löfgren, L.; Duque-Guimaraes, D.E.; Ozilgen, A.; Osler, M.E.; Chibalin, A.V.; Zierath, J.R. Diacylglycerol Kinase-δ Regulates AMPK Signaling, Lipid Metabolism, and Skeletal Muscle Energetics. Am. J. Physiol.—Endocrinol. Metab. 2016, 310, E51. [Google Scholar] [CrossRef] [Green Version]
- Chibalin, A.V.; Leng, Y.; Vieira, E.; Krook, A.; Björnholm, M.; Long, Y.C.; Kotova, O.; Zhong, Z.; Sakane, F.; Steiler, T.; et al. Downregulation of Diacylglycerol Kinase Delta Contributes to Hyperglycemia-Induced Insulin Resistance. Cell 2008, 132, 375–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, R.E.; Kuchenbaecker, K.; Walters, R.K.; Chen, C.Y.; Popejoy, A.B.; Periyasamy, S.; Lam, M.; Iyegbe, C.; Strawbridge, R.J.; Brick, L.; et al. Genome-Wide Association Studies in Ancestrally Diverse Populations: Opportunities, Methods, Pitfalls, and Recommendations. Cell 2019, 179, 589–603. [Google Scholar] [CrossRef] [PubMed]
- Stryjecki, C.; Alyass, A.; Meyre, D. Ethnic and Population Differences in the Genetic Predisposition to Human Obesity. Obes. Rev. 2018, 19, 62–80. [Google Scholar] [CrossRef] [PubMed]
- Young, K.L.; Graff, M.; Fernandez-Rhodes, L.; North, K.E. Genetics of Obesity in Diverse Populations. Curr. Diab. Rep. 2018, 18, 145. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
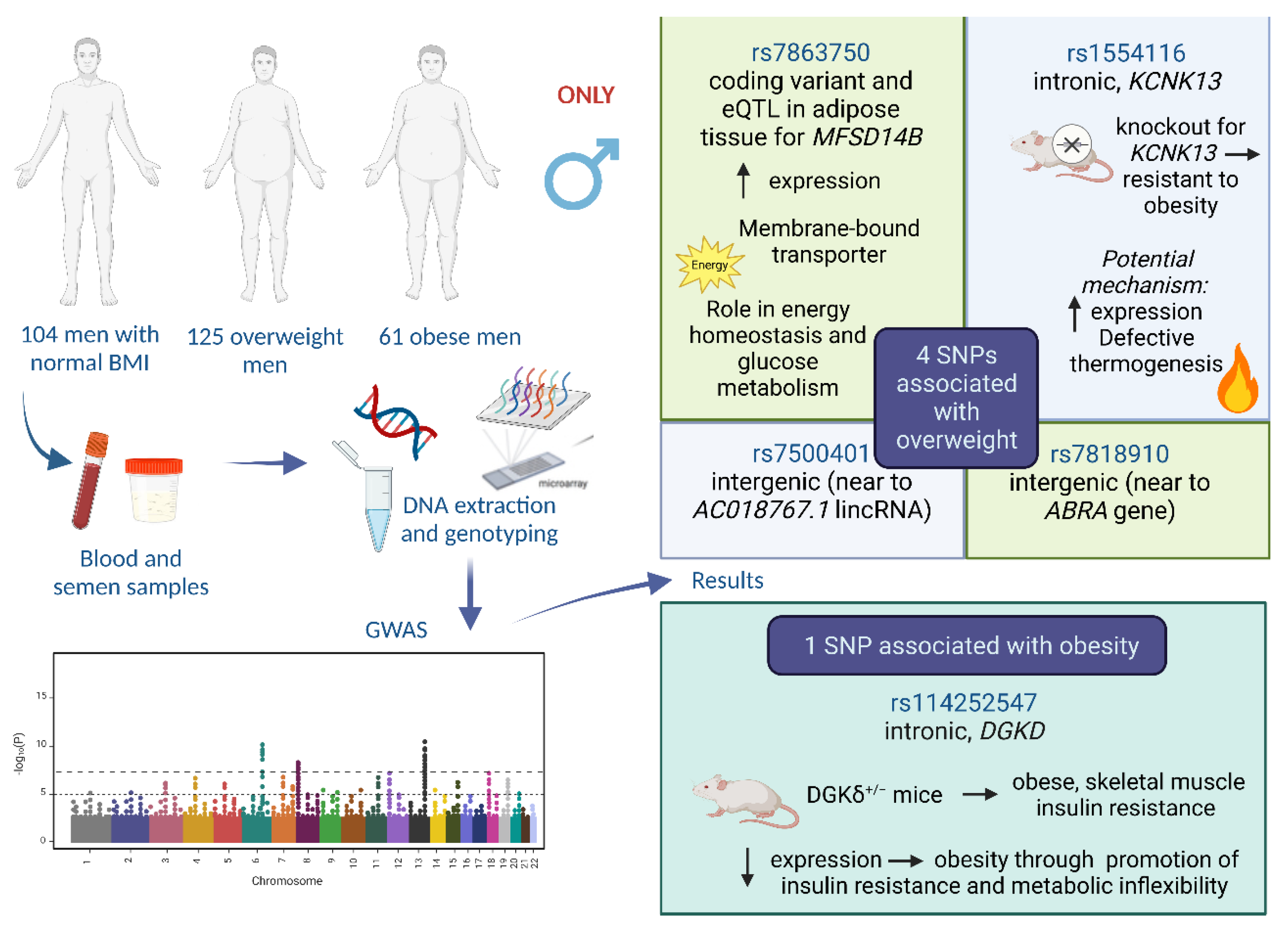
| Normal (n = 104) | Overweight (n = 125) | Obese (n = 61) | |
|---|---|---|---|
| BMI (m/kg2) mean, SD | 19.1–24.9 23.14 (1.48) | 25–29.9 27.05 (1.41) | 30.1–47.6 33.12 (2.94) |
| Age (years) mean, SD | 19–52 32.3 (7.19) | 22–49 36.7 (5.37) | 26–53 39.34 (5.65) |
| Smoking (Yes/No) | No, n = 59 (56.7%) Yes, n = 45 (43.3%) | No, n = 64 (51.2%) Yes, n = 61 (48.8%) | No, n = 39 (63.9%) Yes, n = 22 (36.1%) |
| Alcohol Consumption (<2 drinks/week, 2 drinks/week, >2 drinks/week) | <2 drinks/week, n = 64 (61.6%) 2 drinks/week, n = 20 (19.2%) >2 drinks/week, n = 20 (19.2%) | <2 drinks/week, n = 67 (53.6%) 2 drinks/week, n = 34 (27.2%) >2 drinks/week, n = 24 (19.2%) | <2 drinks/week, n = 41 (67.2%) 2 drinks/week, n = 13 (21.3%) >2 drinks/week, n = 7 (11.5%) |
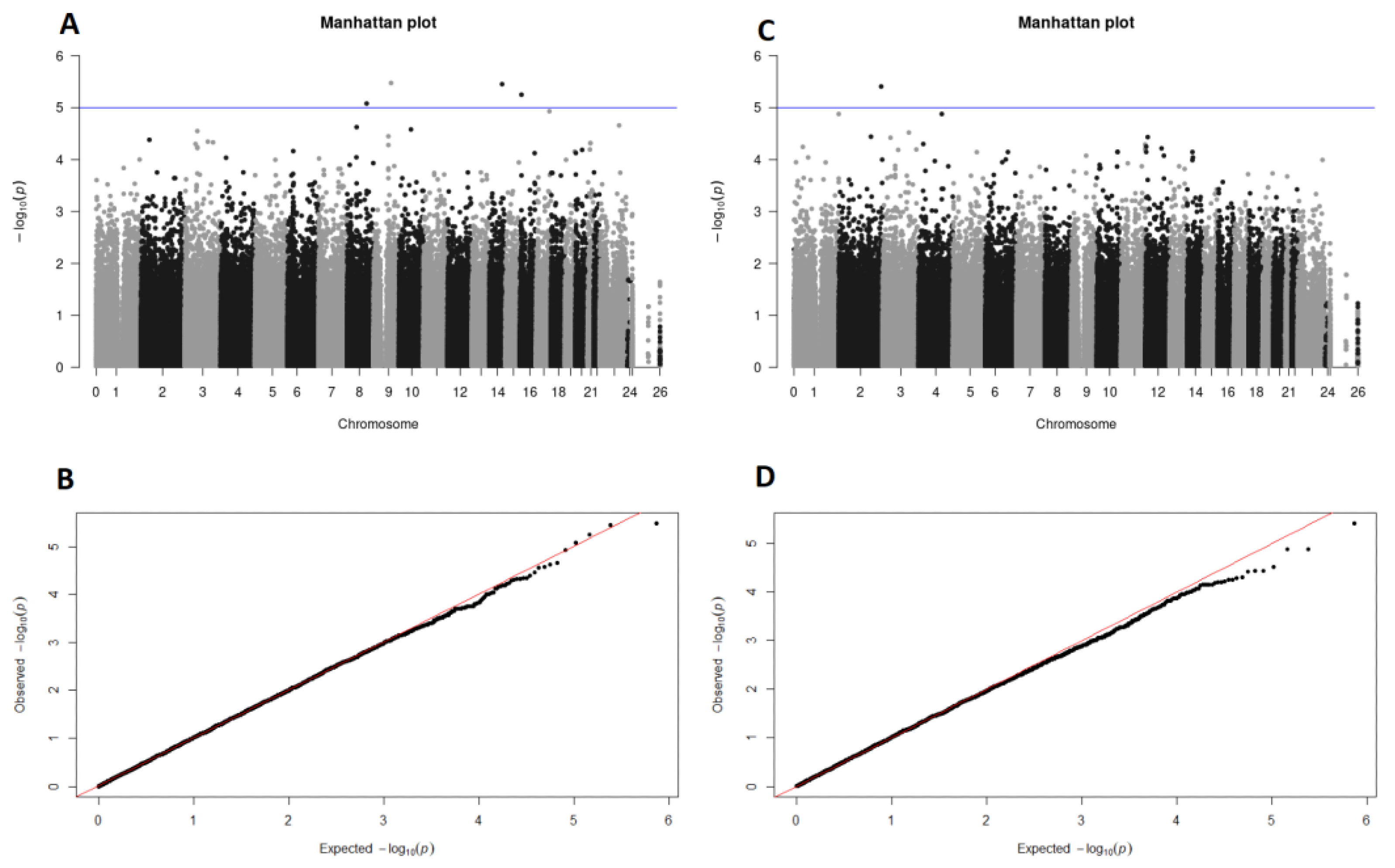
| CHR | SNP | Position | Ref/Alt Allele | Frequency Cases | Frequency Controls | p-Value | OR |
|---|---|---|---|---|---|---|---|
| SNPs associated with overweight | |||||||
| 8 | rs7818910 | 107979407 | A/C | 0.3686 | 0.172 | 8.344 × 10−6 | 2.81 |
| 9 | rs7863750 | 97221463 | G/A | 0.07203 | 0.2312 | 3.356 × 10−6 | 0.2582 |
| 14 | rs1554116 | 90570376 | C/A | 0.572 | 0.3457 | 3.534 × 10−6 | 2.529 |
| 16 | rs7500401 | 8495998 | C/T | 0.1441 | 0.3298 | 5.64 × 10−6 | 0.3421 |
| SNP associated with obesity | |||||||
| 2 | rs114252547 | 234311271 | T/G | 0.1404 | 0.01064 | 3.921 × 10−6 | 15.18 |
| Variant ID | Ref/Alt Allele | MAF | EAS | AMR | AFR | EUR | SAS |
|---|---|---|---|---|---|---|---|
| SNPs associated with overweight | |||||||
| rs7818910 | A/C | 0.48 | 0.378 | 0.653 | 0.272 | 0.719 | 0.522 |
| rs7863750 | G/A | 0.04 | 0.001 | 0.032 | 0.006 | 0.084 | 0.071 |
| rs1554116 | C/A | 0.47 | 0.535 | 0.491 | 0.399 | 0.528 | 0.424 |
| rs7500401 | C/T | 0.36 | 0.489 | 0.429 | 0.194 | 0.259 | 0.510 |
| SNP associated with obesity | |||||||
| rs114252547 | T/G | 0.0157748 | 0 | 0.0159 | 0.0008 | 0.0328 | 0.0348 |
| SNP | Closest Gene | SNP–Gene Distance | Annotation | eQTL | RegulomeDB Score | 3DSNP Score |
|---|---|---|---|---|---|---|
| SNPs associated with overweight | ||||||
| rs7818910 | ABRA | 196 kb | Intergenic | No | Rank = 7, Score = 0.18412 | 1.69 |
| rs7863750 | MFSD14B | 0 kb | Coding | Yes (Adipose Tissue—Visceral and Subcutaneous) | Rank = 5, Score = 0.13454 | 0 |
| rs1554116 | KCNK13 | 0 kb | Intronic | No | Rank = 4, Score = 0.60906 | 1.59 |
| rs7500401 | AC018767.1 (lincRNA) | 70 kb | Intergenic | No | Rank = 5, Score = 0.13454 | 0.79 |
| SNP associated with obesity | ||||||
| rs114252547 | DGKD | 0 kb | Intronic | No | Rank = 5, Score = 0.13454 | 10.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyrgiafini, M.-A.; Sarafidou, T.; Giannoulis, T.; Chatziparasidou, A.; Christoforidis, N.; Mamuris, Z. Gene-by-Sex Interactions: Genome-Wide Association Study Reveals Five SNPs Associated with Obesity and Overweight in a Male Population. Genes 2023, 14, 799. https://doi.org/10.3390/genes14040799
Kyrgiafini M-A, Sarafidou T, Giannoulis T, Chatziparasidou A, Christoforidis N, Mamuris Z. Gene-by-Sex Interactions: Genome-Wide Association Study Reveals Five SNPs Associated with Obesity and Overweight in a Male Population. Genes. 2023; 14(4):799. https://doi.org/10.3390/genes14040799
Chicago/Turabian StyleKyrgiafini, Maria-Anna, Theologia Sarafidou, Themistoklis Giannoulis, Alexia Chatziparasidou, Nikolaos Christoforidis, and Zissis Mamuris. 2023. "Gene-by-Sex Interactions: Genome-Wide Association Study Reveals Five SNPs Associated with Obesity and Overweight in a Male Population" Genes 14, no. 4: 799. https://doi.org/10.3390/genes14040799
APA StyleKyrgiafini, M. -A., Sarafidou, T., Giannoulis, T., Chatziparasidou, A., Christoforidis, N., & Mamuris, Z. (2023). Gene-by-Sex Interactions: Genome-Wide Association Study Reveals Five SNPs Associated with Obesity and Overweight in a Male Population. Genes, 14(4), 799. https://doi.org/10.3390/genes14040799