
Different Nuclear Architecture in Human Sperm According to Their Morphology

 ,
, 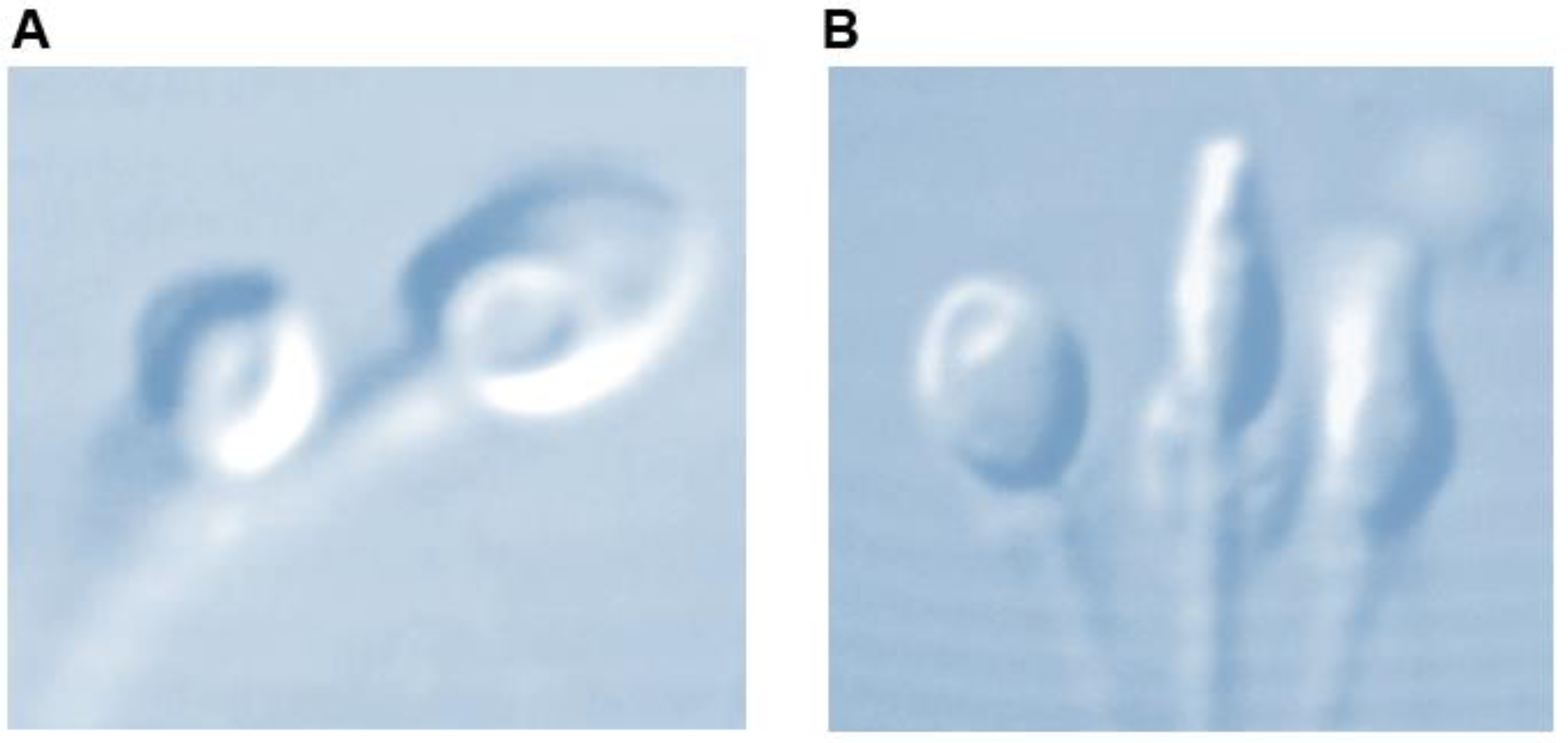{kind=link}
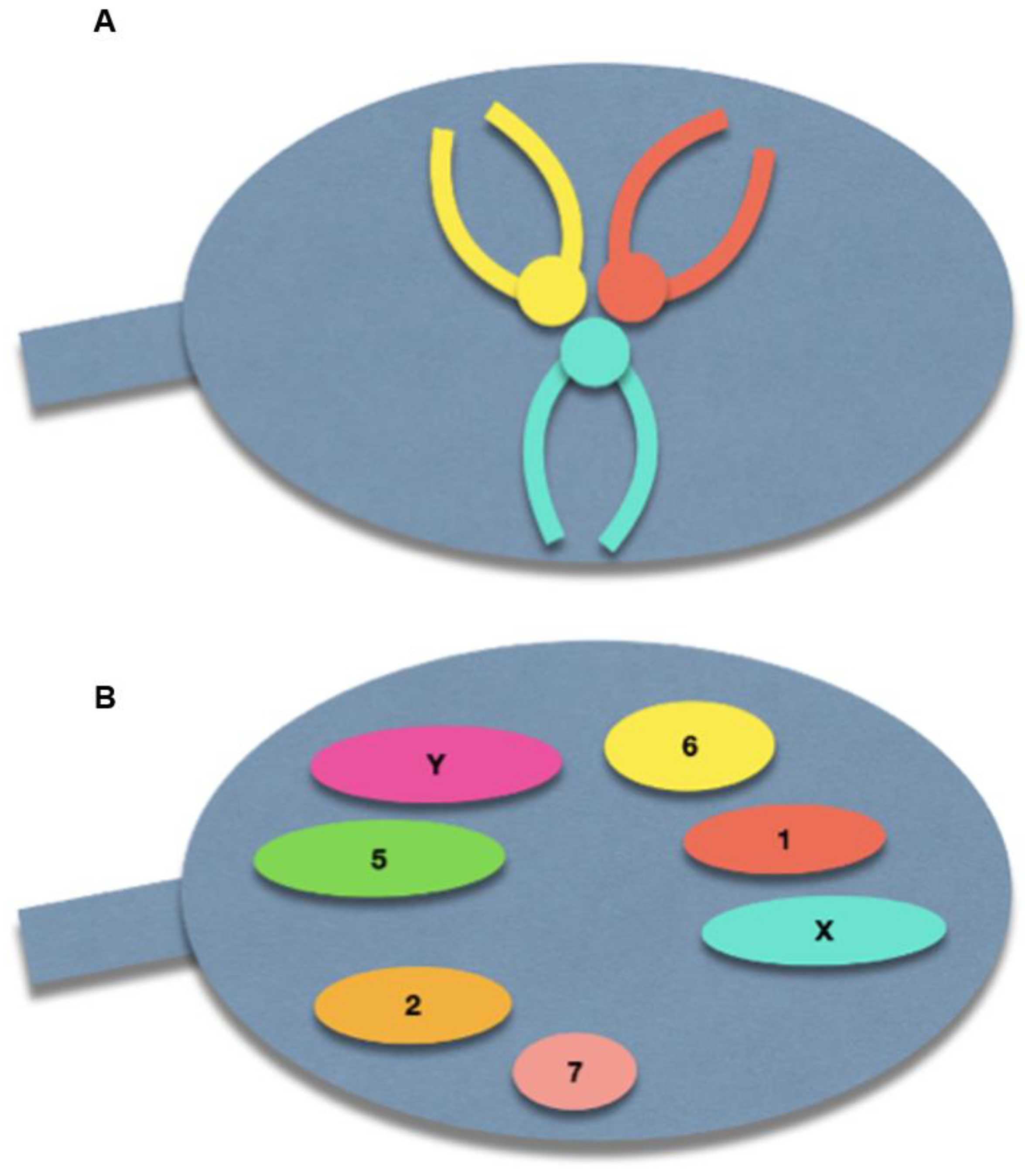{kind=link}
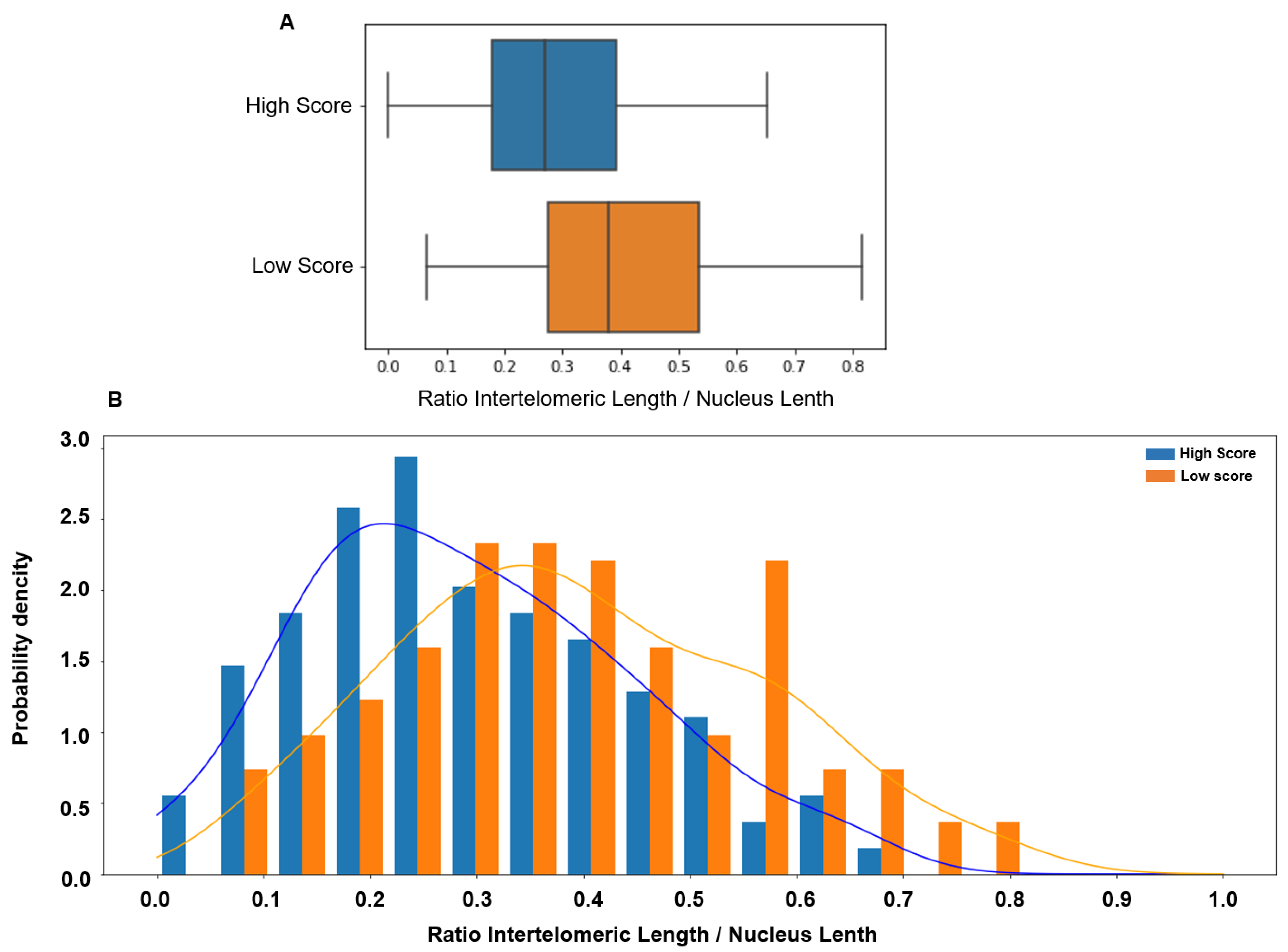{kind=link}
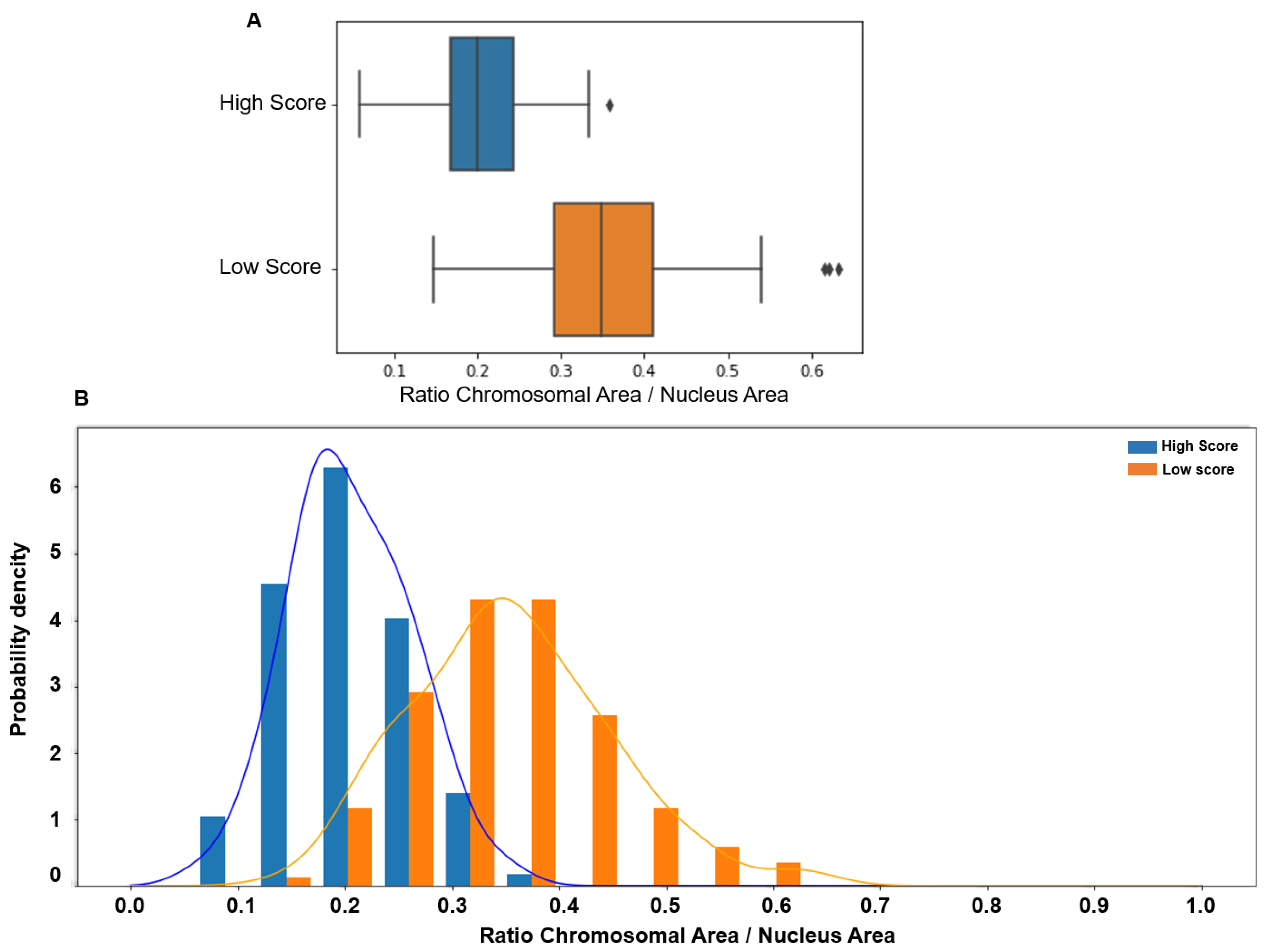{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
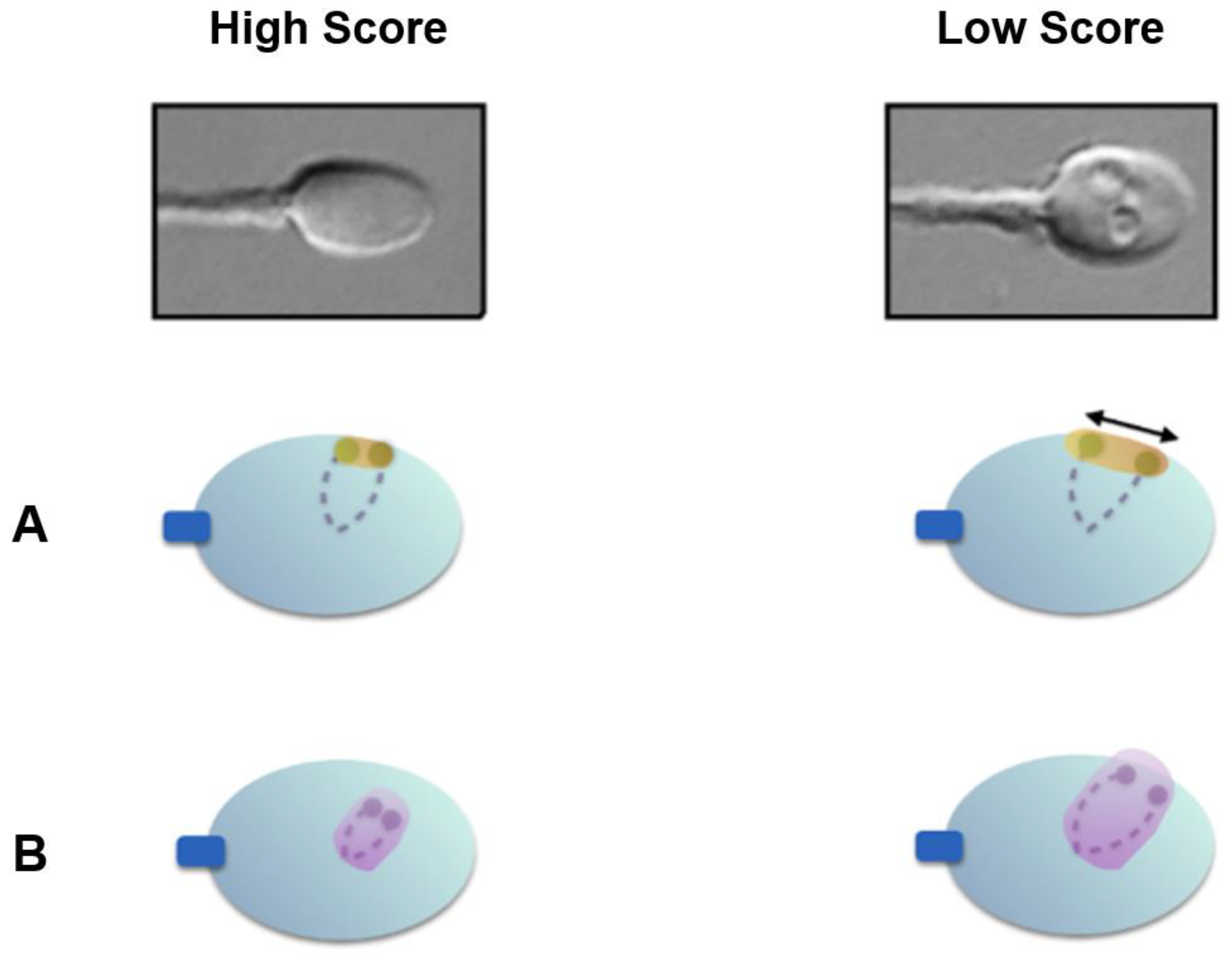
2. Materials and Methods
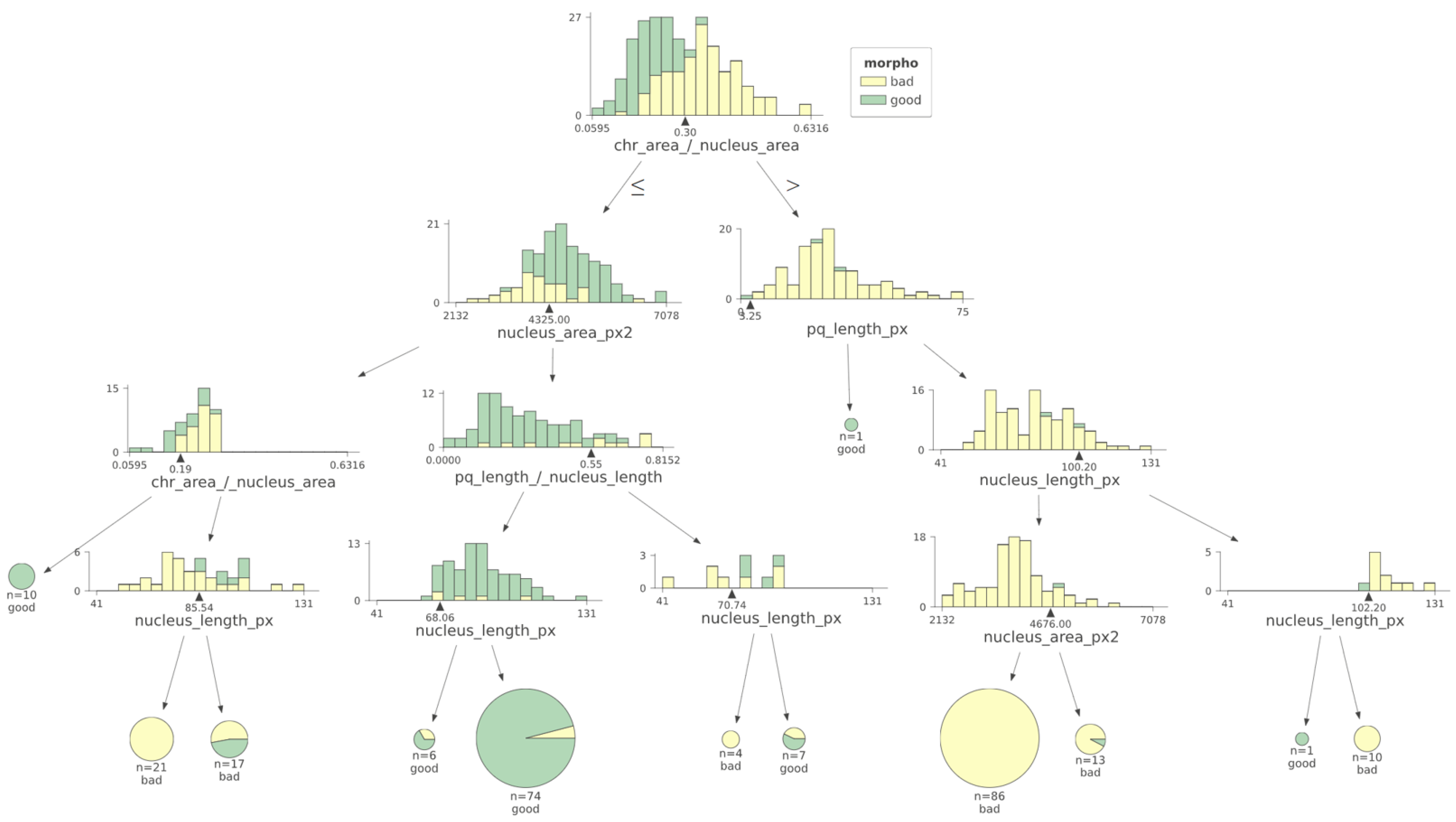
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steptoe, P.C.; Edwards, R.G. Birth after the Reimplantation of a Human Embryo. Lancet 1978, 2, 366. [Google Scholar] [CrossRef]
- Palermo, G.; Joris, H.; Devroey, P.; Van Steirteghem, A.C. Pregnancies after Intracytoplasmic Injection of Single Spermatozoon into an Oocyte. Lancet 1992, 340, 17–18. [Google Scholar] [CrossRef]
- Baldini, D.; Ferri, D.; Baldini, G.M.; Lot, D.; Catino, A.; Vizziello, D.; Vizziello, G. Sperm Selection for ICSI: Do We Have a Winner? Cells 2021, 10, 3566. [Google Scholar] [CrossRef]
- Cassuto, N.G.; Bouret, D.; Plouchart, J.M.; Jellad, S.; Vanderzwalmen, P.; Balet, R.; Larue, L.; Barak, Y. A New Real-Time Morphology Classification for Human Spermatozoa: A Link for Fertilization and Improved Embryo Quality. Fertil. Steril. 2009, 92, 1616–1625. [Google Scholar] [CrossRef]
- Gardner, D.K.; Schoolcraft, W.B. Culture and Transfer of Human Blastocysts. Curr. Opin. Obstet. Gynecol. 1999, 11, 307–311. [Google Scholar] [CrossRef]
- Balaban, B.; Yakin, K.; Alatas, C.; Oktem, O.; Isiklar, A.; Urman, B. Clinical Outcome of Intracytoplasmic Injection of Spermatozoa Morphologically Selected under High Magnification: A Prospective Randomized Study. Reprod. Biomed. Online 2011, 22, 472–476. [Google Scholar] [CrossRef]
- Setti, A.S.; de Almeida Ferreira Braga, D.P.; Vingris, L.; Serzedello, T.; de Cássia Sávio Figueira, R.; Iaconelli, A.; Borges, E. Sperm Morphological Abnormalities Visualised at High Magnification Predict Embryonic Development, from Fertilisation to the Blastocyst Stage, in Couples Undergoing ICSI. J. Assist. Reprod. Genet. 2014, 31, 1533–1539. [Google Scholar] [CrossRef]
- Vanderzwalmen, P.; Hiemer, A.; Rubner, P.; Bach, M.; Neyer, A.; Stecher, A.; Uher, P.; Zintz, M.; Lejeune, B.; Vanderzwalmen, S.; et al. Blastocyst Development after Sperm Selection at High Magnification Is Associated with Size and Number of Nuclear Vacuoles. Reprod. Biomed. Online 2008, 17, 617–627. [Google Scholar] [CrossRef]
- Boitrelle, F.; Guthauser, B.; Alter, L.; Bailly, M.; Wainer, R.; Vialard, F.; Albert, M.; Selva, J. The Nature of Human Sperm Head Vacuoles: A Systematic Literature Review. Basic Clin. Androl. 2013, 23, 3. [Google Scholar] [CrossRef]
- Franco, J.G.; Baruffi, R.L.R.; Mauri, A.L.; Petersen, C.G.; Oliveira, J.B.A.; Vagnini, L. Significance of Large Nuclear Vacuoles in Human Spermatozoa: Implications for ICSI. Reprod. Biomed. Online 2008, 17, 42–45. [Google Scholar] [CrossRef]
- Fekonja, N.; Štrus, J.; Tušek Žnidarič, M.; Knez, K.; Vrtacnik Bokal, E.; Verdenik, I.; Virant-Klun, I. Clinical and Structural Features of Sperm Head Vacuoles in Men Included in the in Vitro Fertilization Programme. Biomed. Res. Int. 2014, 2014, 927841. [Google Scholar] [CrossRef] [PubMed]
- Boitrelle, F.; Ferfouri, F.; Petit, J.M.; Segretain, D.; Tourain, C.; Bergere, M.; Bailly, M.; Vialard, F.; Albert, M.; Selva, J. Large Human Sperm Vacuoles Observed in Motile Spermatozoa under High Magnification: Nuclear Thumbprints Linked to Failure of Chromatin Condensation. Hum. Reprod. 2011, 26, 1650–1658. [Google Scholar] [CrossRef] [PubMed]
- Cassuto, N.G.; Piquemal, D.; Boitrelle, F.; Larue, L.; Lédée, N.; Hatem, G.; Ruoso, L.; Bouret, D.; Siffroi, J.-P.; Rouen, A.; et al. Molecular Profiling of Spermatozoa Reveals Correlations between Morphology and Gene Expression: A Novel Biomarker Panel for Male Infertility. Biomed. Res. Int. 2021, 2021, 1434546. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.-L.; Wu, Y.-B.; Yu, L.; Zheng, Y.; Yang, T.-T.; Wang, Y.-Y.; Zhou, B.; Zhang, L.; Li, F.-P. Normal Sperm Head Morphometric Reference Values in Fertile Asian Males. Asian J. Androl. 2023. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Castillo, J.; Estanyol, J.M.; Ballescá, J.L.; Oliva, R. Human Sperm Chromatin Epigenetic Potential: Genomics, Proteomics, and Male Infertility. Asian J. Androl. 2015, 17, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Zalenskaya, I.A.; Bradbury, E.M.; Zalensky, A.O. Chromatin Structure of Telomere Domain in Human Sperm. Biochem. Biophys. Res. Commun. 2000, 279, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Zalensky, A.; Zalenskaya, I. Organization of Chromosomes in Spermatozoa: An Additional Layer of Epigenetic Information? Biochem. Soc. Trans. 2007, 35, 609–611. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, D.; Tempest, H.G. Human Sperm Chromosomes: To Form Hairpin-Loops, Or Not to Form Hairpin-Loops, That Is the Question. Genes 2019, 10, 504. [Google Scholar] [CrossRef] [PubMed]
- Solov’eva, L.; Svetlova, M.; Bodinski, D.; Zalensky, A.O. Nature of Telomere Dimers and Chromosome Looping in Human Spermatozoa. Chromosome Res. 2004, 12, 817–823. [Google Scholar] [CrossRef]
- Zalensky, A.O.; Tomilin, N.V.; Zalenskaya, I.A.; Teplitz, R.L.; Bradbury, E.M. Telomere-Telomere Interactions and Candidate Telomere Binding Protein(s) in Mammalian Sperm Cells. Exp. Cell Res. 1997, 232, 29–41. [Google Scholar] [CrossRef]
- Rouen, A.; Lavillaureix, A.; Hyon, C.; Heide, S.; Clède, S.; Balet, R.; Kott, E.; Cassuto, N.G.; Siffroi, J.-P. Nuclear Volume Differences between Balanced and Unbalanced Spermatozoa in Chromosomal Translocation Carriers. Reprod. Biomed. Online 2015, 30, 290–295. [Google Scholar] [CrossRef]
- Mebrek, M.L.; Clède, S.; de Chalus, A.; Heide, S.; Ruoso, L.; Rogers, E.; Lédée, N.; Prat-Ellenberg, L.; Cassuto, N.G.; Siffroi, J.-P.; et al. Simple FISH-Based Evaluation of Spermatic Nuclear Architecture Shows an Abnormal Chromosomal Organization in Balanced Chromosomal Rearrangement Carriers. J. Assist. Reprod. Genet. 2020, 37, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Rouen, A.; Carlier, L.; Heide, S.; Egloff, M.; Marzin, P.; Ader, F.; Schwartz, M.; Rogers, E.; Joyé, N.; Balet, R.; et al. Potential Selection of Genetically Balanced Spermatozoa Based on the Hypo-Osmotic Swelling Test in Chromosomal Rearrangement Carriers. Reprod. Biomed. Online 2017, 35, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Awaysheh, A.; Wilcke, J.; Elvinger, F.; Rees, L.; Fan, W.; Zimmerman, K.L. Review of Medical Decision Support and Machine-Learning Methods. Vet. Pathol. 2019, 56, 512–525. [Google Scholar] [CrossRef]
- Ward, W.S. Function of Sperm Chromatin Structural Elements in Fertilization and Development. Mol. Hum. Reprod. 2010, 16, 30–36. [Google Scholar] [CrossRef]
- Liu, L.; Blasco, M.; Trimarchi, J.; Keefe, D. An Essential Role for Functional Telomeres in Mouse Germ Cells during Fertilization and Early Development. Dev. Biol. 2002, 249, 74–84. [Google Scholar] [CrossRef]
- Darmishonnejad, Z.; Zarei-Kheirabadi, F.; Tavalaee, M.; Zarei-Kheirabadi, M.; Zohrabi, D.; Nasr-Esfahani, M.H. Relationship between Sperm Telomere Length and Sperm Quality in Infertile Men. Andrologia 2020, 52, e13546. [Google Scholar] [CrossRef]
- Okada, Y. Sperm Chromatin Structure: Insights from in Vitro to in Situ Experiments. Curr. Opin. Cell Biol. 2022, 75, 102075. [Google Scholar] [CrossRef]
- Mudrak, O.; Tomilin, N.; Zalensky, A. Chromosome Architecture in the Decondensing Human Sperm Nucleus. J. Cell Sci. 2005, 118, 4541–4550. [Google Scholar] [CrossRef]
- Zalensky, A.O.; Allen, M.J.; Kobayashi, A.; Zalenskaya, I.A.; Balhórn, R.; Bradbury, E.M. Well-Defined Genome Architecture in the Human Sperm Nucleus. Chromosoma 1995, 103, 577–590. [Google Scholar] [CrossRef]
- Zalenskaya, I.A.; Zalensky, A.O. Non-Random Positioning of Chromosomes in Human Sperm Nuclei. Chromosome Res. 2004, 12, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Mudrak, O.S.; Nazarov, I.B.; Jones, E.L.; Zalensky, A.O. Positioning of Chromosomes in Human Spermatozoa Is Determined by Ordered Centromere Arrangement. PLoS ONE 2012, 7, e52944. [Google Scholar] [CrossRef]
- Turner, K.J.; Watson, E.M.; Skinner, B.M.; Griffin, D.K. Telomere Distribution in Human Sperm Heads and Its Relation to Sperm Nuclear Morphology: A New Marker for Male Factor Infertility? Int. J. Mol. Sci. 2021, 22, 7599. [Google Scholar] [CrossRef]
- Gómez-Torres, M.J.; Luna-Romero, J.; Fernández-Colom, P.J.; Aizpurua, J.; Avilés, M.; Romero, A. Human Sperm Head Vacuoles Are Related to Nuclear-Envelope Invaginations. Int. J. Mol. Sci. 2023, 24, 10027. [Google Scholar] [CrossRef]
- Bloch, A.; Rogers, E.J.; Nicolas, C.; Martin-Denavit, T.; Monteiro, M.; Thomas, D.; Morel, H.; Lévy, R.; Siffroi, J.-P.; Dupont, C.; et al. Detailed Cell-Level Analysis of Sperm Nuclear Quality among the Different Hypo-Osmotic Swelling Test (HOST) Classes. J. Assist. Reprod. Genet. 2021, 38, 2491–2499. [Google Scholar] [CrossRef]
- Brugnon, F.; Janny, L.; Communal, Y.; Darcha, C.; Szczepaniak, C.; Pellestor, F.; Vago, P.; Pons-Rejraji, H.; Artonne, C.; Grizard, G. Apoptosis and Meiotic Segregation in Ejaculated Sperm from Robertsonian Translocation Carrier Patients. Hum. Reprod. 2010, 25, 1631–1642. [Google Scholar] [CrossRef] [PubMed]
- Ling, X.; Cui, H.; Chen, Q.; Yang, W.; Zou, P.; Yang, H.; Zhou, N.; Deng, J.; Liu, J.; Cao, J.; et al. Sperm Telomere Length Is Associated with Sperm Nuclear DNA Integrity and Mitochondrial DNA Abnormalities among Healthy Male College Students in Chongqing, China. Hum. Reprod. 2023, 38, 1036–1046. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cassuto, N.-G.; Ogal, N.; Assou, S.; Ruoso, L.; Rogers, E.-J.; Monteiro, M.-J.; Thomas, D.; Siffroi, J.-P.; Rouen, A. Different Nuclear Architecture in Human Sperm According to Their Morphology. Genes 2024, 15, 464. https://doi.org/10.3390/genes15040464
Cassuto N-G, Ogal N, Assou S, Ruoso L, Rogers E-J, Monteiro M-J, Thomas D, Siffroi J-P, Rouen A. Different Nuclear Architecture in Human Sperm According to Their Morphology. Genes. 2024; 15(4):464. https://doi.org/10.3390/genes15040464
Chicago/Turabian StyleCassuto, Nino-Guy, Nesrine Ogal, Said Assou, Lea Ruoso, Eli-Jonathan Rogers, Miguel-José Monteiro, Daniel Thomas, Jean-Pierre Siffroi, and Alexandre Rouen. 2024. "Different Nuclear Architecture in Human Sperm According to Their Morphology" Genes 15, no. 4: 464. https://doi.org/10.3390/genes15040464
APA StyleCassuto, N. -G., Ogal, N., Assou, S., Ruoso, L., Rogers, E. -J., Monteiro, M. -J., Thomas, D., Siffroi, J. -P., & Rouen, A. (2024). Different Nuclear Architecture in Human Sperm According to Their Morphology. Genes, 15(4), 464. https://doi.org/10.3390/genes15040464