Copy Number Variation in SOX6 Contributes to Chicken Muscle Development

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Experimental Animals
2.3. CNV Population Genetics
2.4. Association Analysis
2.5. The Transformation of the Copy Number Polymorphism Regional Database
2.6. Homology Analysis of Genes’ Coding Sequences and Construction of Phylogenetic Tree
2.7. Bioinformatics of SOX6
2.8. Detection of DNA Copy Number Variations
NR = 2 −∆∆Ct
2.9. Cell Culture
2.10. RNA Interference
2.11. Reverse Transcription and Quantitative Real-Time PCR
2.12. Cell Cycle Distribution Analysis
2.13. Statistical Analysis
3. Results
3.1. Population Genetics of Chicken Copy Number Variations
3.2. Correlation between CNP and Chicken Economically Important Traits
3.3. Candidate Genes in CNPs Related to Chicken Growth Traits
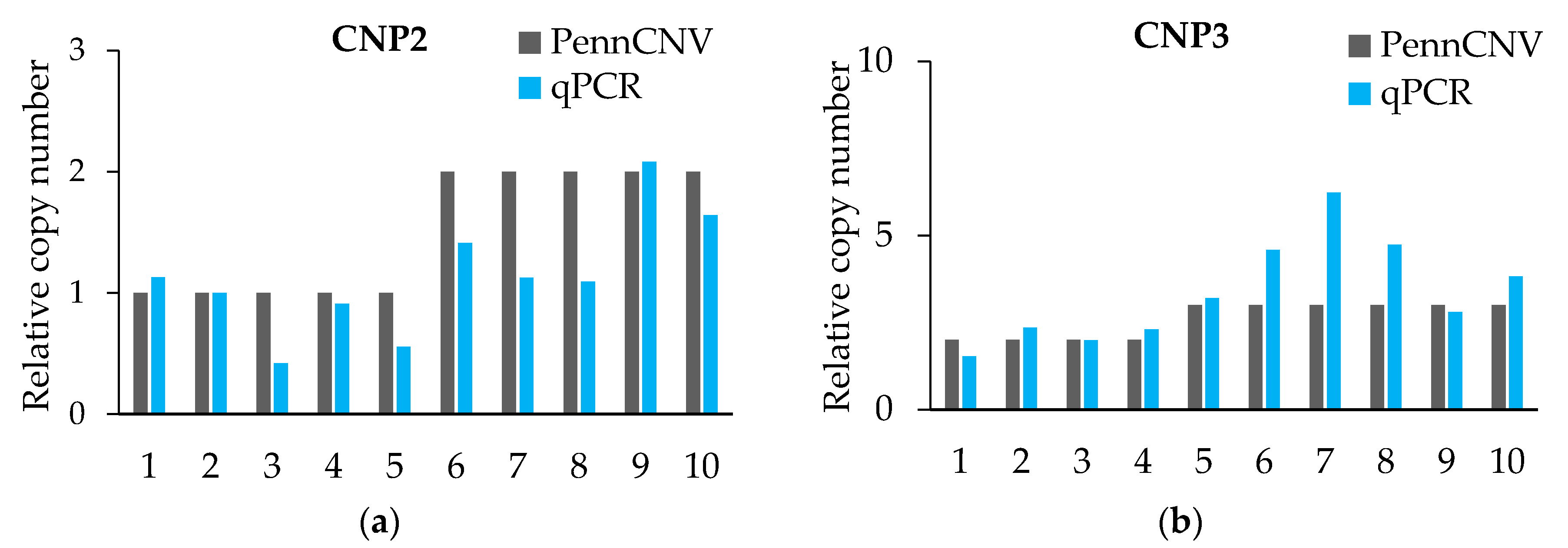
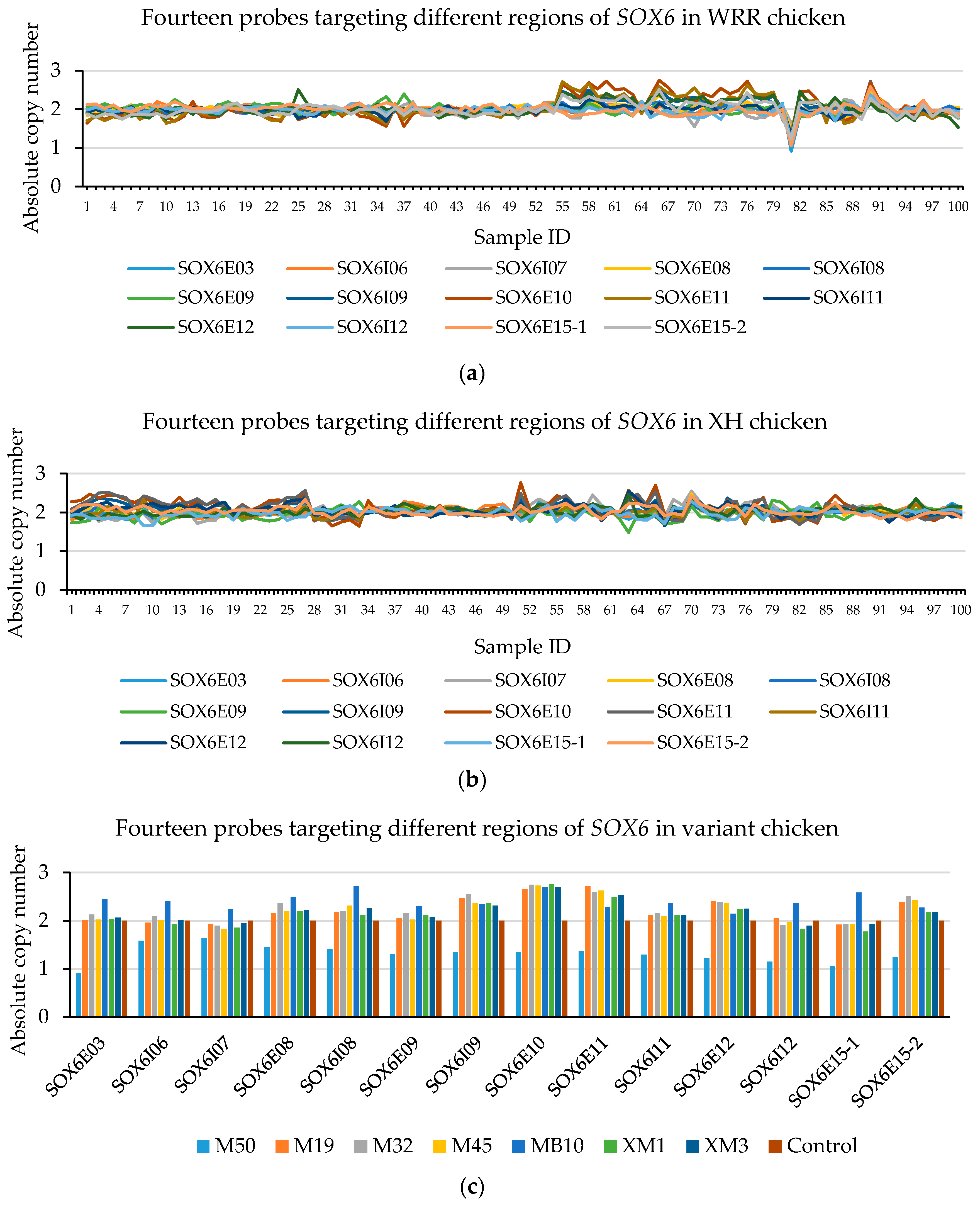
3.4. The CNV in CNPs
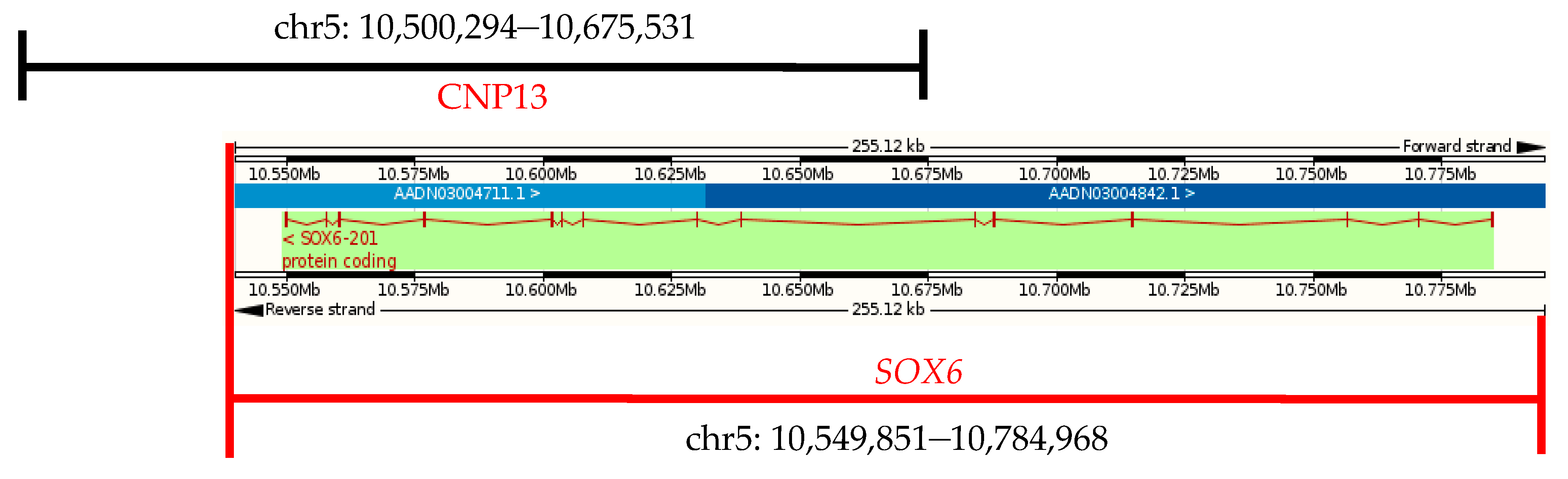
3.5. The CNVs in CNP Structural Mapping
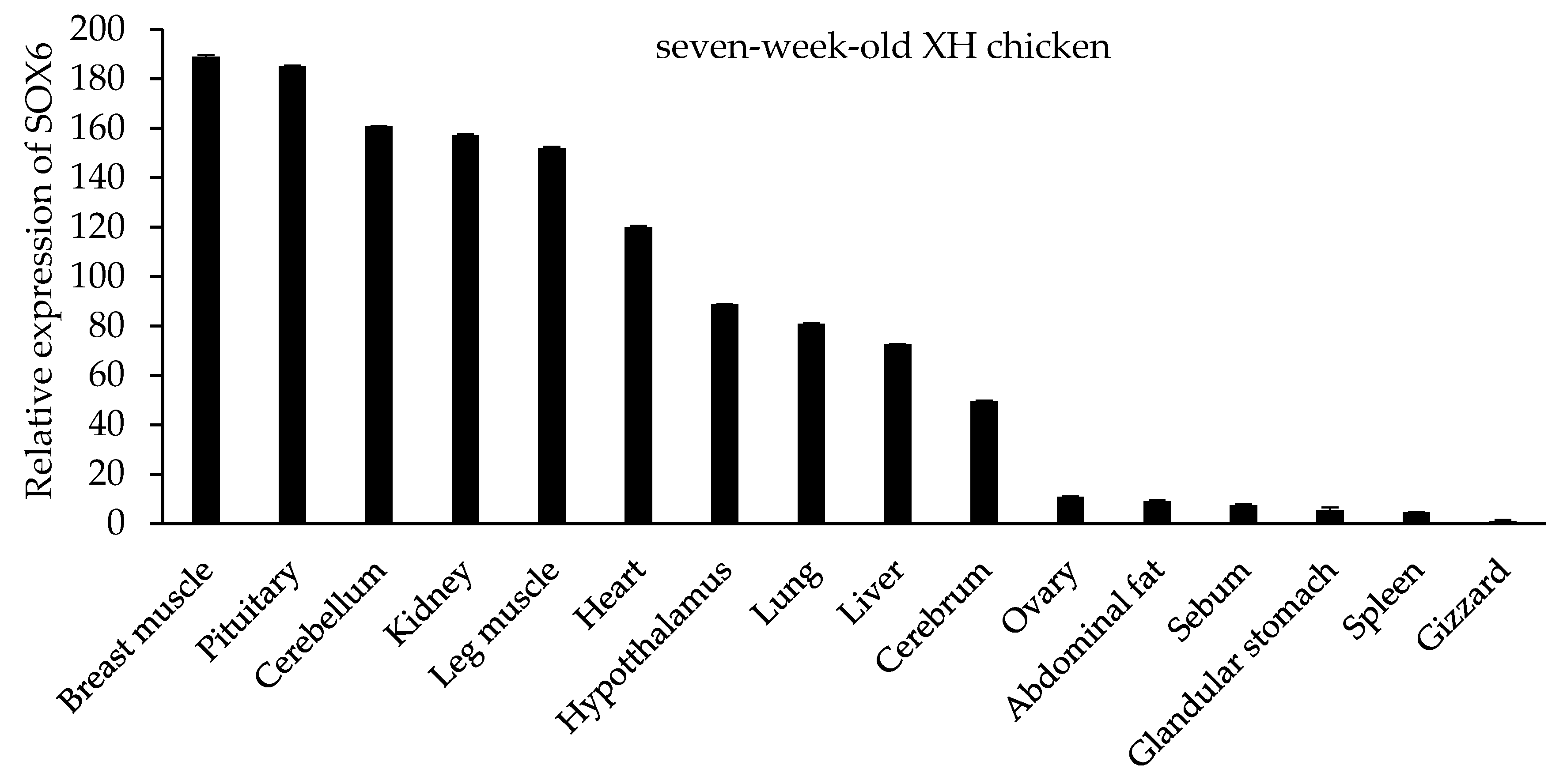
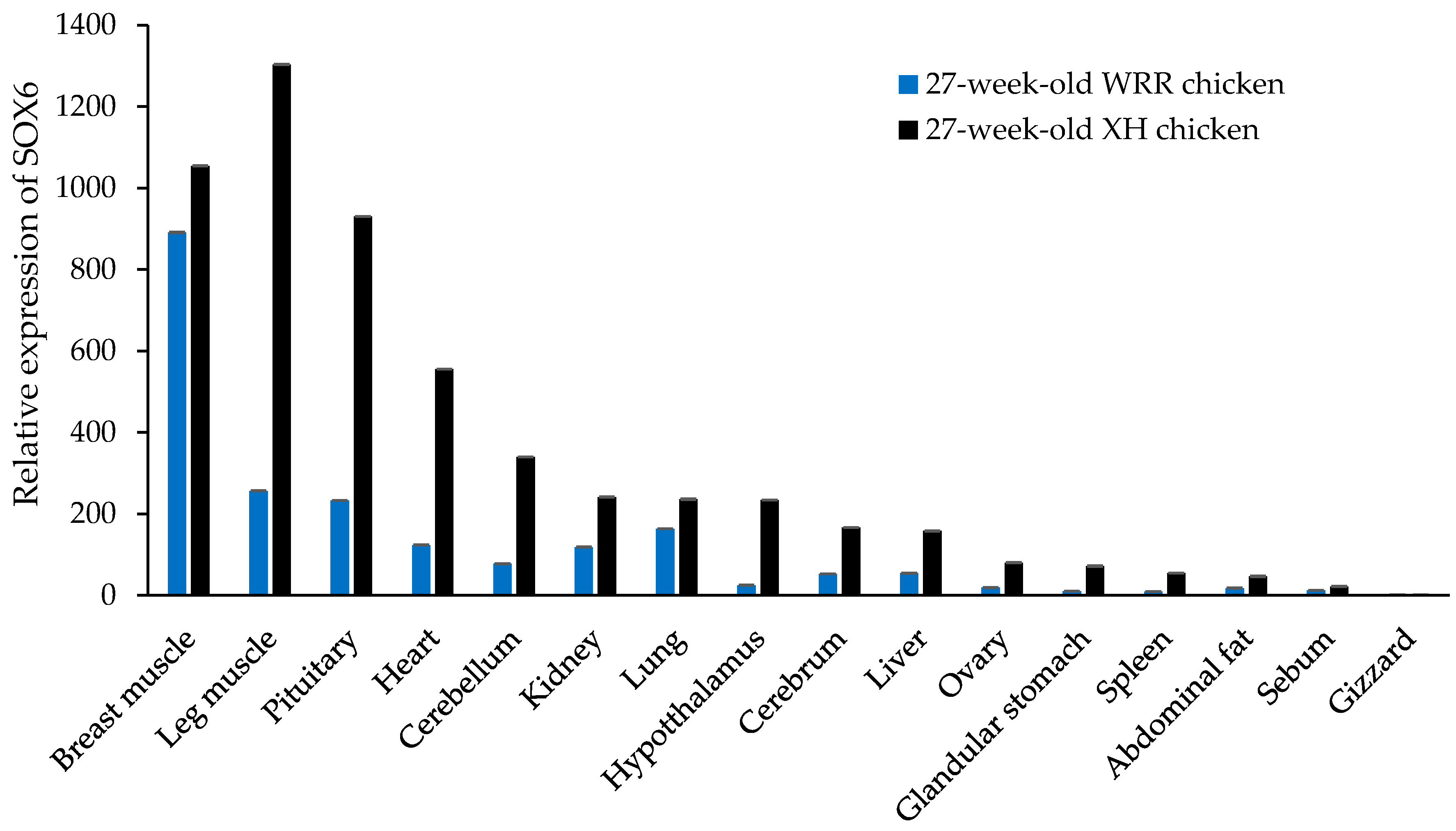
3.6. The Expression Level of SOX6 is Positively Related to Its Copy Number
3.7. Disorder Region of SOX6 Proteins
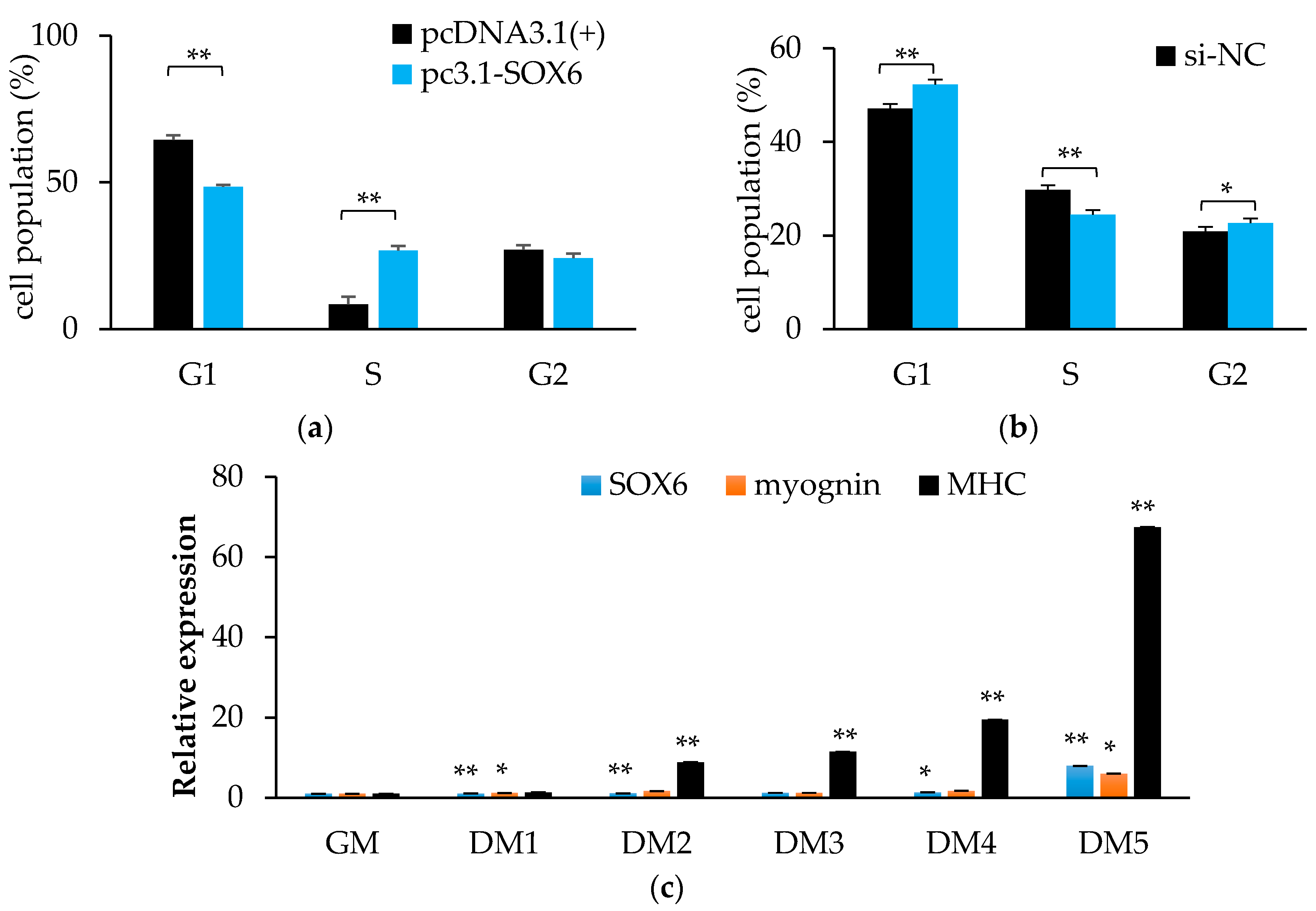
3.8. Chicken SOX6 Promotes Skeletal Cell Proliferation and Differentiation
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Redon, R.; Ishikawa, S.; Fitch, K.R.; Feuk, L.; Perry, G.H.; Andrews, T.D.; Fiegler, H.; Shapero, M.H.; Carson, A.R.; Chen, W.; et al. Global variation in copy number in the human genome. Nature 2006, 444, 444–454. [Google Scholar] [CrossRef] [PubMed]
- McCarroll, S.A.; Altshuler, D.M. Copy-number variation and association studies of human disease. Nat. Genet. 2007, 39, S37–S42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Gu, W.; Hurles, M.E.; Lupski, J.R. Copy number variation in human health, disease, and evolution. Annu. Rev. Genom. Hum. Genet. 2009, 10, 451–481. [Google Scholar] [CrossRef] [PubMed]
- Elferink, M.G.; Vallee, A.A.; Jungerius, A.P.; Crooijmans, R.P.; Groenen, M.A. Partial duplication of the PRLR and SPEF2 genes at the late feathering locus in chicken. BMC Genom. 2008, 9, 391. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.; Boije, H.; Meadows, J.R.; Bed’Hom, B.; Gourichon, D.; Vieaud, A.; Tixier-Boichard, M.; Rubin, C.J.; Imsland, F.; Hallbook, F.; et al. Copy number variation in intron 1 of SOX5 causes the Pea-comb phenotype in chickens. PLoS Genet. 2009, 5, e1000512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorshorst, B.; Molin, A.M.; Rubin, C.J.; Johansson, A.M.; Stromstedt, L.; Pham, M.H.; Chen, C.F.; Hallbook, F.; Ashwell, C.; Andersson, L. A complex genomic rearrangement involving the endothelin 3 locus causes dermal hyperpigmentation in the chicken. PLoS Genet. 2011, 7, e1002412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Yu, Y.; Mitra, A.; Chang, S.; Zhang, H.; Liu, G.; Yang, N.; Song, J. Genome-wide copy number variant analysis in inbred chickens lines with different susceptibility to Marek’s disease. G3 (Bethesda) 2013, 3, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Pevny, L.H.; Lovell-Badge, R. Sox genes find their feet. Curr. Opin. Genet. Dev. 1997, 7, 338–344. [Google Scholar] [CrossRef]
- Mackie, E.J.; Ahmed, Y.A.; Tatarczuch, L.; Chen, K.S.; Mirams, M. Endochondral ossification: How cartilage is converted into bone in the developing skeleton. Int. J. Biochem. Cell Biol. 2008, 40, 46–62. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Im, G.I. Electroporation-mediated transfer of SOX trio genes (SOX-5, SOX-6, and SOX-9) to enhance the chondrogenesis of mesenchymal stem cells. Stem Cells Dev. 2011, 20, 2103–2114. [Google Scholar] [CrossRef] [PubMed]
- Aza-Carmona, M.; Shears, D.J.; Yuste-Checa, P.; Barca-Tierno, V.; Hisado-Oliva, A.; Belinchon, A.; Benito-Sanz, S.; Rodriguez, J.I.; Argente, J.; Campos-Barros, A.; et al. SHOX interacts with the chondrogenic transcription factors SOX5 and SOX6 to activate the aggrecan enhancer. Hum. Mol. Genet. 2011, 20, 1547–1559. [Google Scholar] [CrossRef] [PubMed]
- Jackson, H.E.; Ono, Y.; Wang, X.; Elworthy, S.; Cunliffe, V.T.; Ingham, P.W. The role of Sox6 in zebrafish muscle fiber type specification. Skelet. Muscle 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, N.; Yeh, M.; Liu, A. Sox6 is required for normal fiber type differentiation of fetal skeletal muscle in mice. Dev. Dyn. 2007, 236, 2062–2076. [Google Scholar] [CrossRef] [PubMed]
- An, C.I.; Dong, Y.; Hagiwara, N. Genome-wide mapping of Sox6 binding sites in skeletal muscle reveals both direct and indirect regulation of muscle terminal differentiation by Sox6. BMC Dev. Biol. 2011, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.S.; Li, J.; Zhang, R.; Lin, X.R.; Xu, J.G.; Xie, L.; Xu, Z.Q.; Wang, L.; Gan, J.K.; Xie, X.J.; et al. Copy number variation identification and analysis of the chicken genome using a 60K SNP BeadChip. Poult. Sci. 2016, 95, 1750–1756. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Luo, C.; Zhang, C.; Zhang, R.; Tang, J.; Nie, Q.; Ma, L.; Hu, X.; Li, N.; Da, Y.; et al. Genome-wide association study identified a narrow chromosome 1 region associated with chicken growth traits. PLoS ONE 2012, 7, e30910. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Ji, C.; Zhang, Y.; Zhang, Z.; Nie, Q.; Xu, J.; Zhang, D.; Zhang, X. Combination analysis of genome-wide association and transcriptome sequencing of residual feed intake in quality chickens. BMC Genom. 2016, 17, 594. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol. 2013, 30, W2725–W2729. [Google Scholar] [CrossRef] [PubMed]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Gallo Cassarino, T.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef] [PubMed]
- Majczak, G.; Lilla, S.; Garay-Malpartida, M.; Markovic, J.; Medrano, F.J.; de Nucci, G.; Belizário, J.E. Prediction and biochemical characterization of intrinsic disorder in the structure of proteolysis-inducing factor/dermcidin. Genet. Mol. Res. 2007, 6, 1000–1011. [Google Scholar] [PubMed]
- Du, R.; Lu, C.; Jiang, Z.; Li, S.; Ma, R.; An, H.; Xu, M.; An, Y.; Xia, Y.; Jin, L.; et al. Efficient typing of copy number variations in a segmental duplication-mediated rearrangement hotspot using multiplex competitive amplification. J. Hum. Genet. 2012, 57, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xu, Y.; Liu, D.; Geng, J.; Chen, S.; Jiang, Z.; Fu, Q.; Sun, K. A modified multiplex ligation-dependent probe amplification method for the detection of 22q11.2 copy number variations in patients with congenital heart disease. BMC Genom. 2015, 16, 364. [Google Scholar] [CrossRef] [PubMed]
- Derveaux, S.; Vandesompele, J.; Hellemans, J. How to do successful gene expression analysis using real-time PCR. Methods 2010, 50, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, J.S.; Estivill, X.; Antonarakis, S.E. Copy number variants and genetic traits: Closer to the resolution of phenotypic to genotypic variability. Nat. Rev. Genet. 2007, 8, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Crooijmans, R.P.; Fife, M.S.; Fitzgerald, T.W.; Strickland, S.; Cheng, H.H.; Kaiser, P.; Redon, R.; Groenen, M.A. Large scale variation in DNA copy number in chicken breeds. BMC Genom. 2013, 14, 398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheong, H.I.; Kang, H.G.; Schlondorff, J. GLCCI1 single nucleotide polymorphisms in pediatric nephrotic syndrome. Pediatr. Nephrol. 2012, 27, 1595–1599. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.F.; Lefebvre, V. The transcription factors SOX9 and SOX5/SOX6 cooperate genome-wide through super-enhancers to drive chondrogenesis. Nucleic Acids Res. 2015, 43, 8183–8203. [Google Scholar] [CrossRef] [PubMed]
- Baroti, T.; Zimmermann, Y.; Schillinger, A.; Liu, L.; Lommes, P.; Wegner, M.; Stolt, C.C. Transcription factors Sox5 and Sox6 exert direct and indirect influences on oligodendroglial migration in spinal cord and forebrain. Glia 2016, 64, 122–138. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J.L.; Perry, G.H.; Feuk, L.; Redon, R.; McCarroll, S.A.; Altshuler, D.M.; Aburatani, H.; Jones, K.W.; Tyler-Smith, C.; Hurles, M.E.; et al. Copy number variation: New insights in genome diversity. Genome Res. 2006, 16, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Hunter, T.; Pines, J. Cyclins and cancer. II: Cyclin D and CDK inhibitors come of age. Cell 1994, 79, 573–582. [Google Scholar] [CrossRef]
- Sherr, C.J. The Pezcoller lecture: Cancer cell cycles revisited. Cancer Res. 2000, 60, 3689–3695. [Google Scholar] [PubMed]
- Liu, B.; Yang, L.; Huang, B.; Cheng, M.; Wang, H.; Li, Y.; Huang, D.; Zheng, J.; Li, Q.; Zhang, X.; et al. A functional copy-number variation in MAPKAPK2 predicts risk and prognosis of lung cancer. Am. J. Hum. Genet. 2012, 91, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.; Peng, X.; Zhou, M.; Luo, C.; Nie, Q.; Zhang, X. Polymorphisms of the IGF1R gene and their genetic effects on chicken early growth and carcass traits. BMC Genet. 2008, 9, 70. [Google Scholar] [CrossRef] [PubMed]
- Tosca, L.; Chabrolle, C.; Crochet, S.; Tesseraud, S.; Dupont, J. IGF-1 receptor signaling pathways and effects of AMPK activation on IGF-1-induced progesterone secretion in hen granulosa cells. Domest. Anim. Endocrinol. 2008, 34, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Ling, B.M.; Bharathy, N.; Chung, T.K.; Kok, W.K.; Li, S.; Tan, Y.H.; Rao, V.K.; Gopinadhan, S.; Sartorelli, V.; Walsh, M.J.; et al. Lysine methyltransferase G9a methylates the transcription factor MyoD and regulates skeletal muscle differentiation. Proc. Natl. Acad. Sci. USA 2012, 109, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Patel, S.P.; McCarthy, J.J.; Rabchevsky, A.G.; Goldhamer, D.J.; Esser, K.A. A non-canonical E-box within the MyoD core enhancer is necessary for circadian expression in skeletal muscle. Nucleic Acids Res. 2012, 40, 3419–3430. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Cinar, M.U.; Phatsara, C.; Tesfaye, D.; Tholen, E.; Looft, C.; Schellander, K. Molecular mechanism underlying the differential MYF6 expression in postnatal skeletal muscle of Duroc and Pietrain breeds. Gene 2011, 486, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Te Pas, M.F.; Verburg, F.J.; Gerritsen, C.L.; de Greef, K.H. Messenger ribonucleic acid expression of the MyoD gene family in muscle tissue at slaughter in relation to selection for porcine growth rate. J. Anim. Sci. 2000, 78, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Rudrabhatla, P.; Utreras, E.; Jaffe, H.; Kulkarni, A.B. Regulation of Sox6 by cyclin dependent kinase 5 in brain. PLoS ONE 2014, 9, e89310. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.K.; deLeeuw, R.J.; Dosanjh, N.S.; Kimm, L.R.; Cheng, Z.; Horsman, D.E.; MacAulay, C.; Ng, R.T.; Brown, C.J.; Eichler, E.E.; et al. A comprehensive analysis of common copy-number variations in the human genome. Am. J. Hum. Genet. 2007, 80, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shi, T.; Cai, H.; Zhou, Y.; Lan, X.; Zhang, C.; Lei, C.; Qi, X.; Chen, H. Associations of MYH3 gene copy number variations with transcriptional expression and growth traits in Chinese cattle. Gene 2014, 535, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Uversky, V.N.; Gillespie, J.R.; Fink, A.L. Why are “natively unfolded” proteins unstructured under physiologic conditions? Proteins 2000, 41, 415–427. [Google Scholar] [CrossRef]
- Dyson, H.J.; Wright, P.E. Intrinsically unstructured proteins and their functions. Nat. Rev. Mol. Cell Biol. 2005, 6, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Forman-Kay, J.D.; Mittag, T. From sequence and forces to structure, function, and evolution of intrinsically disordered proteins. Structure 2013, 21, 1492–1499. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.P.; Dash, D. Intrinsic disorder in yeast transcriptional regulatory network. Proteins 2007, 68, 602–605. [Google Scholar] [CrossRef] [PubMed]
- Patil, A.; Nakamura, H. Disordered domains and high surface charge confer hubs with the ability to interact with multiple proteins in interaction networks. FEBS Lett. 2006, 580, 2041–2045. [Google Scholar] [CrossRef] [PubMed]
- Dunker, A.K.; Bondos, S.E.; Huang, F.; Oldfield, C.J. Intrinsically disordered proteins and multicellular organisms. Semin. Cell. Dev. Biol. 2015, 37, 44–55. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
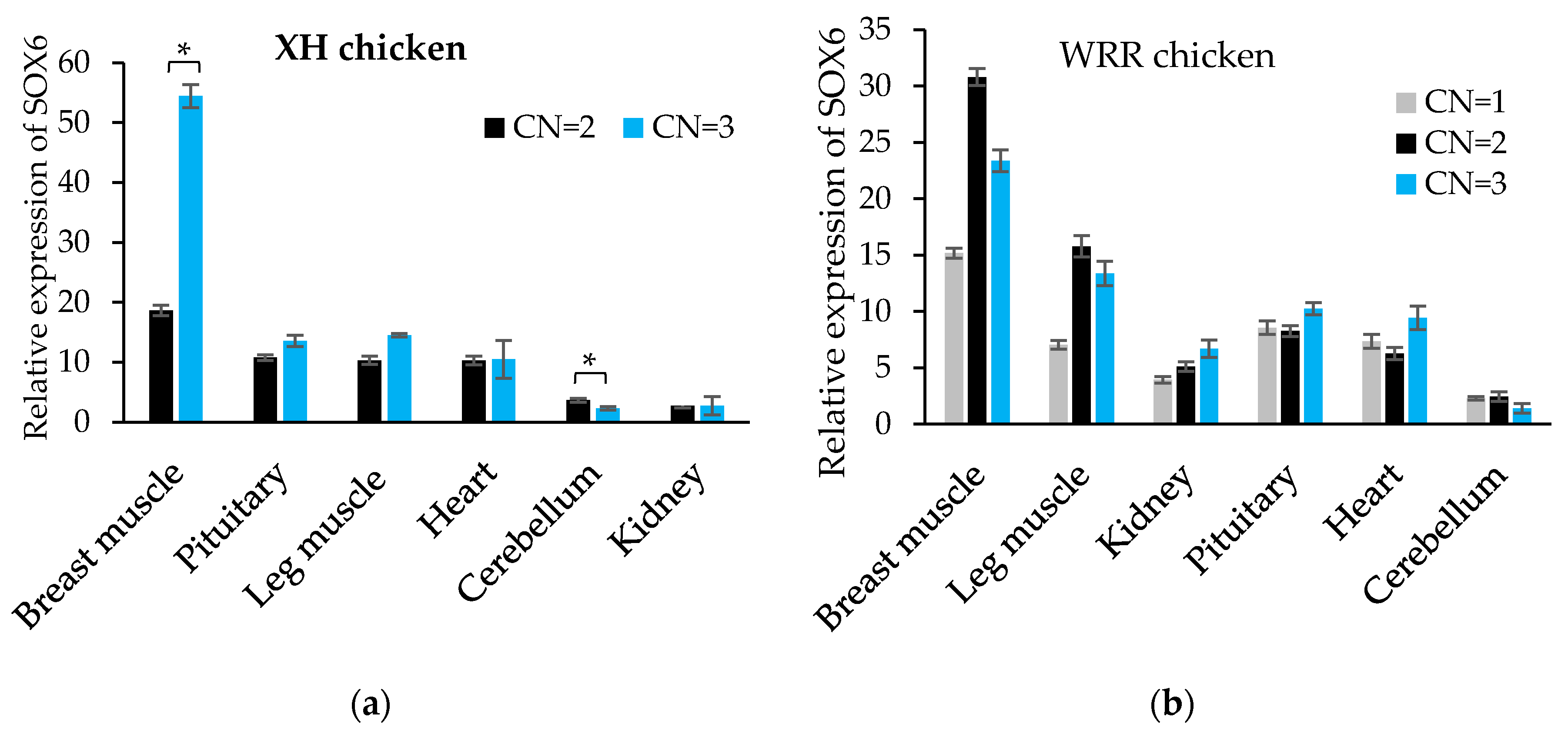{kind=link}
{kind=link}
{kind=link}
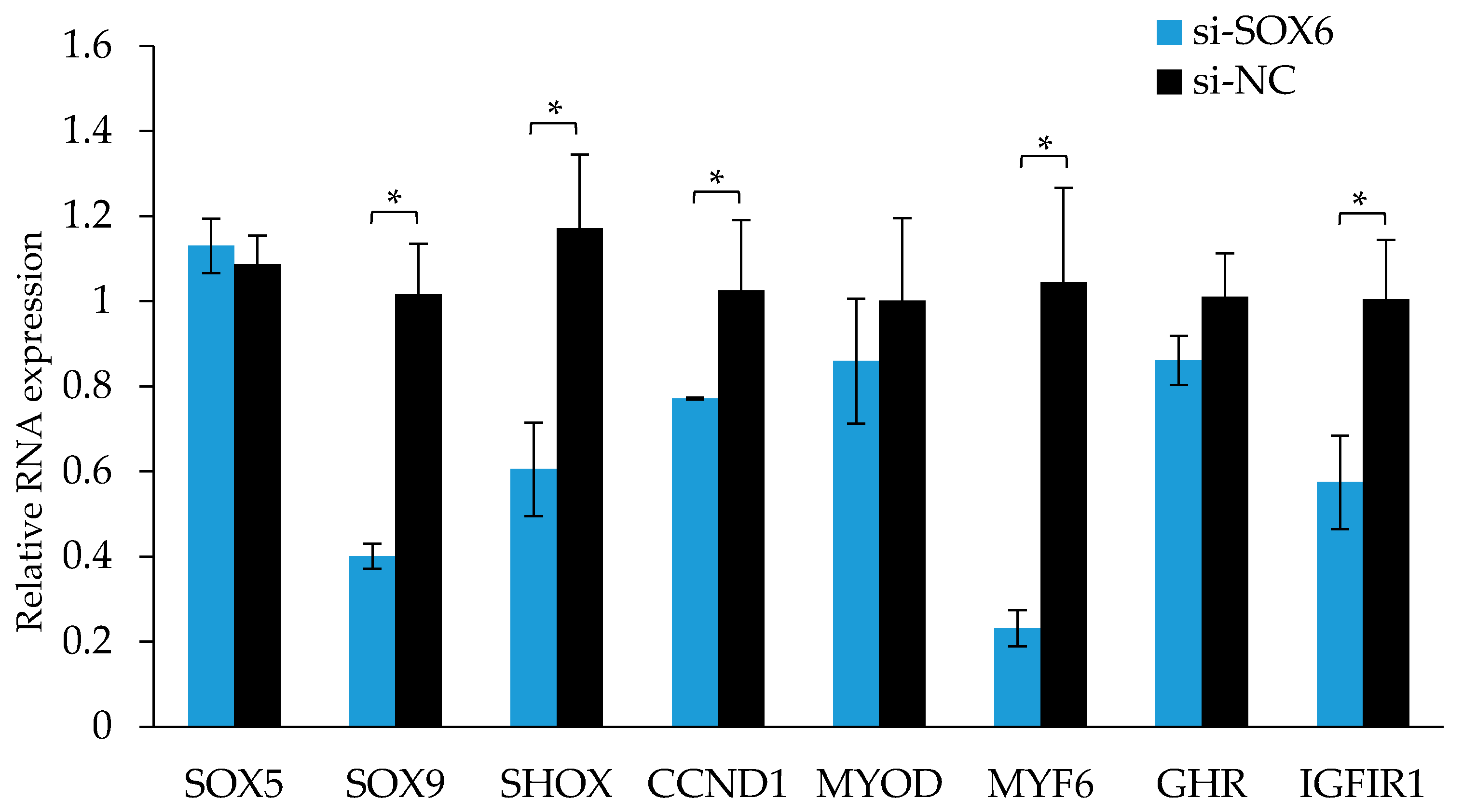{kind=link}
| CNP | Chr. | Start | End | CN | Gene | Frequency (%) |
|---|---|---|---|---|---|---|
| CNP1 | 1 | 114669300 | 115092768 | 0, 1 | EFHC2, MAOA | 3.83 |
| CNP2 | 2 | 24780291 | 24949275 | 1 | GLCCI1 | 5.66 |
| CNP3 | 4 | 88930573 | 89076844 | 3 | CD8A | 3.65 |
| CNP4 | 10 | 9320467 | 10038259 | 1 | ARPP19, KIAA1370, MYO5A | 3.65 |
| CNP5 | 16 | 220950 | 299166 | 0, 3 | B-G, LOC396417, YFV | 9.49 |
| CNP6 | 18 | 3342968 | 3377149 | 1 | RAB40B, WDR45B | 4.01 |
| CNP7 | 20 | 105179 | 224625 | 1 | C20orf4 | 4.01 |
| CNP8 | 1 | 19027876 | 19083051 | 3 | NOT_FOUND | 3.83 |
| CNP9 | 1 | 66088686 | 66170851 | 1, 0 | NOT_FOUND | 6.02 |
| CNP10 | 1 | 144183468 | 144308761 | 3 | NOT_FOUND | 8.76 |
| CNP11 | 1 | 167313565 | 167378938 | 1 | NOT_FOUND | 4.74 |
| CNP12 | 2 | 132873944 | 132955032 | 0, 1 | NOT_FOUND | 6.57 |
| CNP13 | 5 | 12088601 | 12242199 | 1 | SOX6 | 6.75 |
| CNP14 | 8 | 9713469 | 9770642 | 1 | NOT_FOUND | 4.20 |
| CNP15 | 2 | 27005716 | 27140786 | 1 | NOT_FOUND | 3.28 |
| CNP16 | 2 | 137594135 | 137647003 | 1 | NOT_FOUND | 3.10 |
| CNP17 | 8 | 29199309 | 29221258 | 1 | NOT_FOUND | 3.10 |
| CNP18 | 13 | 716593 | 769986 | 3 | PCDHA | 3.19 |
| CNP19 | Z | 12889235 | 12952470 | 0 | GHR | 3.19 |
| CNV Frequency (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Illumine SNP Array | AccuCopy® CNVplex® | De-novo 1 Sequencing | |||||||
| CNP | CN 2 (Genotype) | XH | WRR | XH × WRR F1 | XH × WRR F2 | XH | WRR | XH | WRR |
| CNP2 | 1 (1/0) | - 3 | - | - | 5.66 | - | - | - | - |
| CNP4 | 1 (1/0) | 7.14 | 14.29 | - | 3.40 | - | - | - | - |
| CNP6 | 1 (1/0) | - | 7.14 | 3.57 | 4.00 | - | - | - | 10.00 |
| CNP10 | 3 (1/2) | - | 14.29 | 17.86 | 8.20 | 6.00 | - | 100.00 | 100.00 |
| CNP13 | 1 (1/0) | 14.29 | 7.14 | - | 6.80 | - | 1.00 | - | - |
| 3 (1/2) | - | - | - | - | 2.00 | 4.00 | 4.00 | 10.00 | |
| CNP ID | Trait | p Value 1 | Contribution Rate (%) | Mean ± SD 2 | |
|---|---|---|---|---|---|
| CNP14 | breast muscle conductivity | 0.04675 | 26.81 | 8.19 ± 0.67 a,3 (CN = 1) | 6.32 ± 0.18 b (CN = 2) |
| breast muscle dry matter content (%) | 0.0119 | 25.93 | 25.23 ± 0.34 B (CN = 1) | 26.19 ± 0.21 A (CN = 2) | |
| CNP17 | 63 d tibia length (mm) | 0.0029 | 52.50 | 90.67 ± 3.49 A (CN = 1) | 76.1 ± 0.69 B (CN = 2) |
| CNP ID | CN | Gene | Up or Down-Regulate Growth Traits |
|---|---|---|---|
| CNP2 | 1 | GLCCI1 | ↑ |
| CNP3 | 3 | CD8A | ↓ |
| CNP4 | 1 | ARPP19, KIAA1370, MYO5A | ↓ |
| CNP6 | 1 | WDR45B, RAB40B | ↑ |
| CNP10 | 3 | NOT FOUND | ↑ |
| CNP13 | 1 | SOX6 | ↓ |
| CNP ID | CNP Region | Type | Validation | Positive Verification Rate (%) |
|---|---|---|---|---|
| CNP2 | chr2: 24780291–24949275 | LOSS | Yes | 80 |
| CNP3 | chr4: 88930573–89076844 | GAIN | Yes | 100 |
| CNP4 | chr10: 9320467–10038259 | LOSS | Yes | 80 |
| CNP6 | chr18: 3342968–3377149 | LOSS | Yes | 80 |
| CNP10 | chr1: 144183468–144308761 | GAIN | Yes | 100 |
| CNP13 | chr5: 12088601–12242199 | LOSS | Yes | 100 |
| CNP | Region 1 | CNV Type | ||
|---|---|---|---|---|
| XH | WRR | |||
| CNP2 | GLCCI1E04 | CN = 2 | CN = 2 | |
| CNP4 | ARPP19E02 | CN = 2 | CN = 2 | |
| CNP6 | RAB40BE03 | CN = 2 | CN = 2 | |
| FAM214AE05 | CN = 2 | CN = 2 | ||
| CNP13 | SOX6E09 | CN = 2 | CN = 2 | CN = 1 (1%) 1 |
| Breast Muscle | Pituitary | Leg Muscle | Heart | Cerebellum | Kidney | |
|---|---|---|---|---|---|---|
| XH | 0.0000323436 | 0.53629558 | 0.830763012 | 0.958416629 | 0.026859996 | 0.177897854 |
| WRR | 0.001143708 | 0.147867296 | 0.190177707 | 0.400274735 | 0.175778229 | 0.255664008 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.; Lin, X.; Zhang, Z.; Jiang, M.; Rao, Y.; Nie, Q.; Zhang, X. Copy Number Variation in SOX6 Contributes to Chicken Muscle Development. Genes 2018, 9, 42. https://doi.org/10.3390/genes9010042
Lin S, Lin X, Zhang Z, Jiang M, Rao Y, Nie Q, Zhang X. Copy Number Variation in SOX6 Contributes to Chicken Muscle Development. Genes. 2018; 9(1):42. https://doi.org/10.3390/genes9010042
Chicago/Turabian StyleLin, Shudai, Xiran Lin, Zihao Zhang, Mingya Jiang, Yousheng Rao, Qinghua Nie, and Xiquan Zhang. 2018. "Copy Number Variation in SOX6 Contributes to Chicken Muscle Development" Genes 9, no. 1: 42. https://doi.org/10.3390/genes9010042
APA StyleLin, S., Lin, X., Zhang, Z., Jiang, M., Rao, Y., Nie, Q., & Zhang, X. (2018). Copy Number Variation in SOX6 Contributes to Chicken Muscle Development. Genes, 9(1), 42. https://doi.org/10.3390/genes9010042