Transcriptome Profiling Reveals the EanI/R Quorum Sensing Regulon in Pantoea Ananatis LMG 2665T

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Identification of RNA Sampling Points
2.3. Total RNA Isolation
2.4. Library Construction and Sequencing
2.5. RNA Sequencing Data Analysis
2.6. Complementary DNA Synthesis and Quantitative Reverse Transcription Polymerase Chain Reaction Validation of RNA-Sequencing Data
2.7. Accession Number(s)
3. Results
3.1. RNA Sampling Points
3.2. Illumina Sequencing and Read Mapping
3.3. Quantitative Reverse Transcription Polymerase Chain Reaction Validation of RNA-Sequencing Data
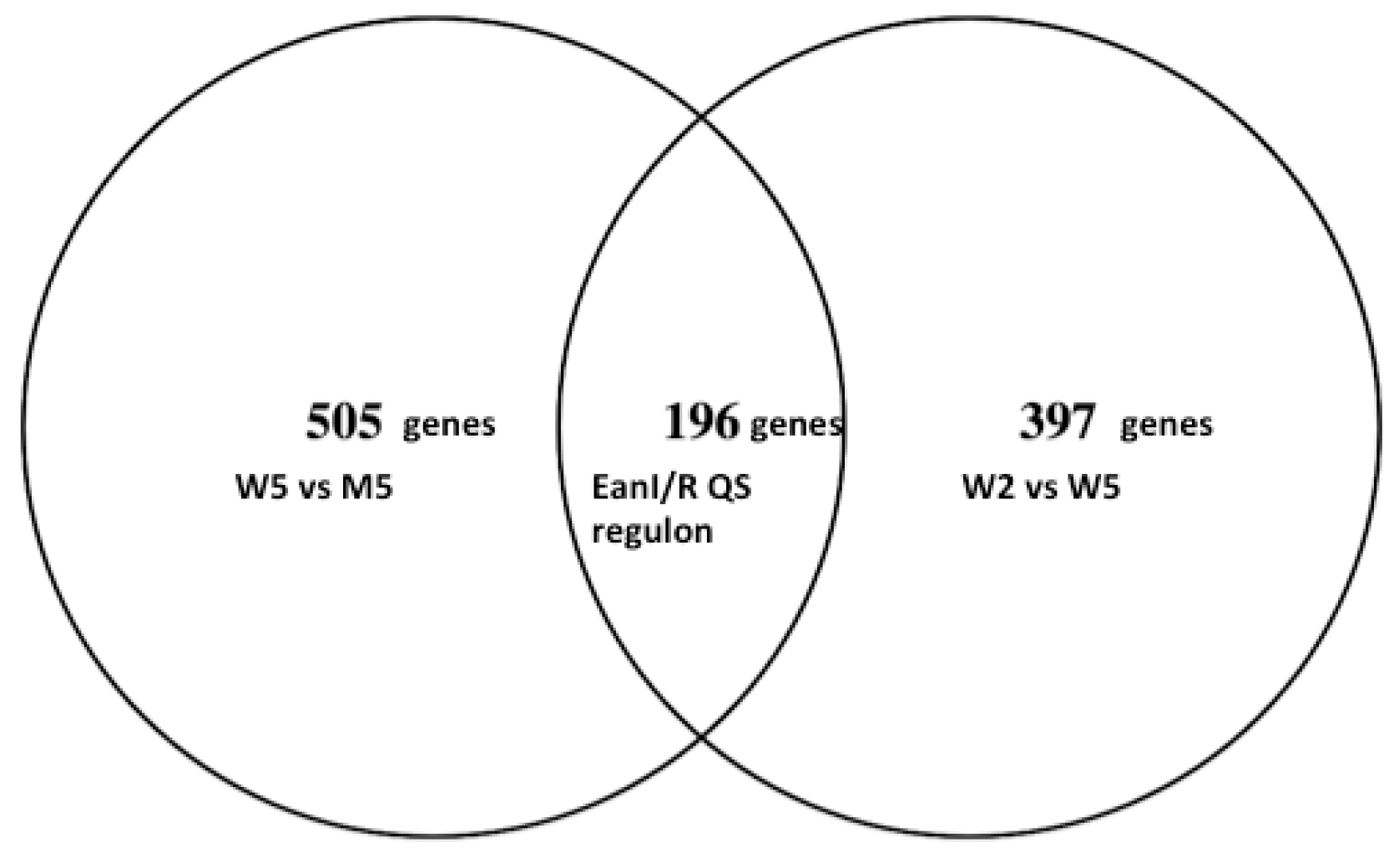
3.4. Identification of the EanI/R Quorum Sensing Regulon of Pantoea Ananatis LMG 2665T
3.5. Functional Annotation of the EanI/R Regulon in Pantoea ananatis
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Coutinho, T.A.; Venter, S.N. Pantoea ananatis: An unconventional plant pathogen. Mol. Plant Pathol. 2009, 10, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, T.; Preisig, O.; Mergaert, J.; Cnockaert, M.; Riedel, K.-H.; Swings, J.; Wingfield, M. Bacterial blight and dieback of Eucalyptus species, hybrids, and clones in South Africa. Plant Dis. 2002, 86, 20–25. [Google Scholar] [CrossRef]
- Cota, L.; Costa, R.; Silva, D.; Parreira, D.; Lana, U.; Casela, C. First report of pathogenicity of Pantoea ananatis in sorghum (Sorghum bicolor) in Brazil. Australas. Plant Dis. Notes 2010, 5, 120–122. [Google Scholar] [CrossRef]
- Morohoshi, T.; Nakamura, Y.; Yamazaki, G.; Ishida, A.; Kato, N.; Ikeda, T. The plant pathogen Pantoea ananatis produces N-acylhomoserine lactone and causes center rot disease of onion by quorum sensing. J. Bacteriol. 2007, 189, 8333–8338. [Google Scholar] [CrossRef] [PubMed]
- Sibanda, S.; Theron, J.; Shyntum, D.Y.; Moleleki, L.N.; Coutinho, T.A. Characterization of two LuxI/R homologs in Pantoea ananatis LMG 2665T. Can. J. Microbiol. 2016, 62, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Shyntum, D.Y.; Theron, J.; Venter, S.N.; Moleleki, L.N.; Toth, I.K.; Coutinho, T.A. Pantoea ananatis utilizes a type VI secretion system for pathogenesis and bacterial competition. Mol. Plant Microbe Interact. 2015, 28, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Shyntum, D.Y.; Venter, S.N.; Moleleki, L.N.; Toth, I.; Coutinho, T.A. Comparative genomics of type VI secretion systems in strains of Pantoea ananatis from different environments. BMC Genom. 2014, 15, 163. [Google Scholar] [CrossRef] [PubMed]
- Weller-Stuart, T.; Toth, I.; de Maayer, P.; Coutinho, T. Swimming and twitching motility are essential for attachment and virulence of Pantoea ananatis in onion seedlings. Mol. Plant Pathol. 2017, 18, 734–745. [Google Scholar] [CrossRef] [PubMed]
- De Maayer, P.; Chan, W.Y.; Venter, S.N.; Toth, I.K.; Birch, P.R.; Joubert, F.; Coutinho, T.A. Genome sequence of Pantoea ananatis LMG20103, the causative agent of Eucalyptus blight and dieback. J. Bacteriol. 2010, 192, 2936–2937. [Google Scholar] [CrossRef] [PubMed]
- De Kievit, T.R.; Iglewski, B.H. Bacterial quorum sensing in pathogenic relationships. Infect. Immun. 2000, 68, 4839–4849. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, C.; Parsek, M.R.; Greenberg, E.P. Regulation of gene expression by cell-to-cell communication: Acyl-homoserine lactone quorum sensing. Annu. Rev. Genet. 2001, 35, 439–468. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, R.; Burke, A.K.; Cormier, G.; Jensen, R.V.; Stevens, A.M. Transcriptome-based analysis of the Pantoea stewartii quorum-sensing regulon and identification of EsaR direct targets. Appl. Environ. Microbiol. 2014, 80, 5790–5800. [Google Scholar] [CrossRef] [PubMed]
- Adam, Z.; Tambong, J.T.; Lewis, C.T.; Lévesque, C.A.; Chen, W.; Bromfield, E.S.; Khan, I.U.; Xu, R. Draft genome sequence of Pantoea ananatis strain LMG 2665T, a bacterial pathogen of pineapple fruitlets. Genome Announc. 2014, 2, e00489-14. [Google Scholar] [CrossRef] [PubMed]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef] [PubMed]
- Minogue, T.D.; Trebra, M.W.V.; Bernhard, F.; Bodman, S.B.V. The autoregulatory role of EsaR, a quorum-sensing regulator in Pantoea stewartii ssp. stewartii: Evidence for a repressor function. Mol. Microbiol. 2002, 44, 1625–1635. [Google Scholar] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- FastQC, version 0.11.5; Simon Andrews: Babraham, UK, 2017; Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 30 January 2018).
- Wang, L.; Wang, S.; Li, W. RSeQC: Quality control of RNA-seq experiments. Bioinformatics 2012, 28, 2184–2185. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Takle, G.W.; Toth, I.K.; Brurberg, M.B. Evaluation of reference genes for real-time RT-PCR expression studies in the plant pathogen Pectobacterium atrosepticum. BMC Plant Biol. 2007, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Moleleki, L.N.; Pretorius, R.G.; Tanui, C.K.; Mosina, G.; Theron, J. A quorum sensing-defective mutant of Pectobacterium carotovorum ssp. brasiliense 1692 is attenuated in virulence and unable to occlude xylem tissue of susceptible potato plant stems. Mol. Plant Pathol. 2017, 18, 32–44. [Google Scholar] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Winans, S.C. The quorum-sensing transcriptional regulator TraR requires its cognate signaling ligand for protein folding, protease resistance, and dimerization. Proc. Natl. Acad. Sci. USA 2001, 98, 1507–1512. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Coulthurst, S.J.; Pritchard, L.; Hedley, P.E.; Ravensdale, M.; Humphris, S.; Burr, T.; Takle, G.; Brurberg, M.-B.; Birch, P.R. Quorum sensing coordinates brute force and stealth modes of infection in the plant pathogen Pectobacterium atrosepticum. PLoS Pathog. 2008, 4, e1000093. [Google Scholar] [CrossRef] [PubMed]
- De Maayer, P.; Chan, W.Y.; Rubagotti, E.; Venter, S.N.; Toth, I.K.; Birch, P.R.; Coutinho, T.A. Analysis of the Pantoea ananatis pan-genome reveals factors underlying its ability to colonize and interact with plant, insect and vertebrate hosts. BMC Genom. 2014, 15, 404. [Google Scholar] [CrossRef] [PubMed]
- Antúnez-Lamas, M.; Cabrera-Ordonez, E.; Lopez-Solanilla, E.; Raposo, R.; Trelles-Salazar, O.; Rodríguez-Moreno, A.; Rodríguez-Palenzuela, P. Role of motility and chemotaxis in the pathogenesis of Dickeya dadantii 3937 (ex Erwinia chrysanthemi 3937). Microbiology 2009, 155, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Allen, C. The plant pathogen Ralstonia solanacearum needs aerotaxis for normal biofilm formation and interactions with its tomato host. J. Bacteriol. 2007, 189, 6415–6424. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.M.; Shibata, S.; Aizawa, S.-I.; Tsuyumu, S. Motility is an important determinant for pathogenesis of Erwinia carotovora subsp. carotovora. Physiol. Mol. Plant Pathol. 2005, 66, 134–143. [Google Scholar] [CrossRef]
- Jang, M.S.; Goo, E.; An, J.H.; Kim, J.; Hwang, I. Quorum sensing controls flagellar morphogenesis in Burkholderia glumae. PLoS ONE 2014, 9, e84831. [Google Scholar] [CrossRef] [PubMed]
- Fatima, U.; Senthil-Kumar, M. Plant and pathogen nutrient acquisition strategies. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-W.; Jeong, K.-S.; Han, S.-W.; Lee, S.-E.; Phee, B.-K.; Hahn, T.-R.; Ronald, P. The Xanthomonas oryzae pv. oryzae PhoPQ two-component system is required for AvrXA21 activity, HRPG expression, and virulence. J. Bacteriol. 2008, 190, 2183–2197. [Google Scholar] [PubMed]
- Nakka, S.; Qi, M.; Zhao, Y. The Erwinia amylovora PhoPQ system is involved in resistance to antimicrobial peptide and suppresses gene expression of two novel type III secretion systems. Microbiolo. Res. 2010, 165, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Mellgren, E.M.; Kloek, A.P.; Kunkel, B.N. Mqo, a tricarboxylic acid cycle enzyme, is required for virulence of Pseudomonas syringae pv. tomato strain DC3000 on Arabidopsis thaliana. J. Bacteriol. 2009, 191, 3132–3141. [Google Scholar] [PubMed]
- Morohoshi, T.; Oseki, K.; Ikeda, T. Exopolysaccharide production is influenced by sugars, N-acylhomoserine lactone, and transcriptional regulators RcsA and RcsB, but does not affect pathogenicity in the plant pathogen Pantoea ananatis. Biosci. Biotechnol. Biochem. 2011, 75, 997–999. [Google Scholar] [CrossRef] [PubMed]
- Burbank, L.; Mohammadi, M.; Roper, M.C. Siderophore-mediated iron acquisition influences motility and is required for full virulence of the xylem-dwelling bacterial phytopathogen Pantoea stewartii subsp. stewartii. Appl. Environ. Microbiol. 2015, 81, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Tanui, C.K.; Shyntum, D.Y.; Priem, S.L.; Theron, J.; Moleleki, L.N. Influence of the ferric uptake regulator (Fur) protein on pathogenicity in Pectobacterium carotovorum subsp. brasiliense. PLoS ONE 2017, 12, e0177647. [Google Scholar] [CrossRef] [PubMed]
- Expert, D. Withholding and exchanging iron: Interactions between Erwinia spp. and their plant hosts. Annu. Rev. Phytopathol. 1999, 37, 307–334. [Google Scholar] [CrossRef] [PubMed]
- Rossez, Y.; Wolfson, E.B.; Holmes, A.; Gally, D.L.; Holden, N.J. Bacterial flagella: Twist and stick, or dodge across the kingdoms. PLoS Pathog. 2015, 11, e1004483. [Google Scholar] [CrossRef] [PubMed]
- De Orué Lucana, D.O. Redox Sensing: Novel Avenues and Paradigms; Mary Ann Liebert, Inc.: New Rochelle, NY, USA, 2012. [Google Scholar]
- Hérault, E.; Reverchon, S.; Nasser, W. Role of the LysR-type transcriptional regulator PecT and DNA supercoiling in the thermoregulation of PEL genes, the major virulence factors in Dickeya dadantii. Environ. Microbiol. 2014, 16, 734–745. [Google Scholar] [CrossRef] [PubMed]
- Thomson, N.R.; Nasser, W.; McGowan, S.; Sebaihia, M.; Salmond, G.P. Erwinia carotovora has two KdgR-like proteins belonging to the IciR family of transcriptional regulators: Identification and characterization of the RexZ activator and the KdgR repressor of pathogenesis. Microbiology 1999, 145, 1531–1545. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.M.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.; Soares, M.; Facincani, A.; Ferreira, C.; Ferreira, R.; Ferro, M.; Gozzo, F.; Felestrino, É.; Assis, R.; Garcia, C. Proteomics-based identification of differentially abundant proteins reveals adaptation mechanisms of Xanthomonas citri subsp. citri during Citrus sinensis infection. BMC Microbiol. 2017, 17, 155. [Google Scholar] [PubMed]
- Mhedbi-Hajri, N.; Yahiaoui, N.; Mondy, S.; Hue, N.; Pélissier, F.; Faure, D.; Dessaux, Y. Transcriptome analysis revealed that a quorum sensing system regulates the transfer of the pAt megaplasmid in Agrobacterium tumefaciens. BMC Genom. 2016, 17, 661. [Google Scholar] [CrossRef] [PubMed]
- Dalsing, B.L.; Truchon, A.N.; Gonzalez-Orta, E.T.; Milling, A.S.; Allen, C. Ralstonia solanacearum uses inorganic nitrogen metabolism for virulence, ATP production, and detoxification in the oxygen-limited host xylem environment. mBio 2015, 6, e02471-14. [Google Scholar] [CrossRef] [PubMed]
- Morohoshi, T.; Ogata, Y.; Ikeda, T. Cell aggregation is negatively regulated by N-acylhomoserine lactone-mediated quorum sensing in Pantoea ananatis SK-1. J. Biosci. Bioeng. 2011, 112, 566–569. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Mapped Reads (%) | Uniquely Mapped Reads (%) | RNA Integrity Number (RIN) |
|---|---|---|---|
| P. ananatis 2665TeanΔI/R samples | |||
| M5-1 | 19,318,790 | 16,835,822 | 9.5 |
| M5-2 | 20,712,376 | 18,833,927 | 9.2 |
| M5-3 | 20,108,558 | 17,421,192 | 9.5 |
| P. ananatis 2665T samples | |||
| W2-1 | 19,364,330 | 17,023,603 | 9.5 |
| W2-2 | 19,483,482 | 17,520,238 | 9.6 |
| W5-1 | 19,953,814 | 15,768,738 | 8.8 |
| W5-3 | 20,417,384 | 18,445,793 | 9.1 |
| Gene Name | Protein ID | Primer Name and Sequence | Source |
|---|---|---|---|
| Methyl-accepting chemotaxis protein I (tsr) | WP_014605659.1 | TsRF CATGAATGAGATCGTCAGTGCG | This study |
| TsRR GTTGTGTTACGCGATCCATCTC | |||
| Flagellar hook-associated protein 2 (fliD) | WP_014593941.1 | FliDF CAAATGATGGCAGTCTGTCGC | This study |
| FliDR GTGATCGACACGCCGTTAATC | |||
| Carbamoyl-phosphate synthase (carB) | WP_014592889.1 | CarBF GATCCGAAAGTCCACCTTG | This study |
| CarBR GATTGAATACGCCGTCCAC | |||
| Hypothetical protein | WP_028715941.1 | HypF CAACTGGCGGACTACCAAC | This study |
| HypR GCCCTGACCAGTAATTGTCAG | |||
| Shikimate 5-dehydrogenase (aroE) | WP_014606596.1 | ShikiF CGACAGCGTTATTCTGACC | This study |
| ShikiR AATAAGCTCAGGACGCAGG | |||
| Glycoside hydrolase | WP_028715822.1 | GlyF GCGTTGCTACCGCAAATCAAG | This study |
| GlyR GTAACACCTTGCGTGTGACC |
| Functional Category | Gene or Protein Encoded | Log2 Ratio | |
|---|---|---|---|
| W2 vs. W5 | M5 vs. W5 | ||
| Genes up regulated between the wild-type strain LMG 2665T in the presence of AHLs (at OD600 = 0.5) versus the same strain in the absence if AHLs (OD600 = 0.2) | |||
| Redox sensing | 3-oxoacyl-ACP reductase | 1.833 | 1.638 |
| Oxidoreductase | 2.115 | 1.93 | |
| Metabolism | pyruvate oxidase | 1.156 | 1.217 |
| malate:quinone oxidoreductase | 1.42 | 0.561 | |
| transketolase/WP_014593932.1 | 1.401 | 1.588 | |
| lysine 6-monooxygenase | 1.351 | 0.966 | |
| ketol-acid reductoisomerase | 1.187 | 0.888 | |
| alcA | 1.532 | 0.869 | |
| IucA | 2.106 | 1.444 | |
| siderophore-interacting protein (WP_014606889.1) | 1.686 | 1.014 | |
| pyridine nucleotide-disulphide oxidoreductase | 0.845 | −0.738 | |
| Wza | 0.971 | −0.984 | |
| Flagella formation and flagella mediated Motility | fliA | 2.837 | 0.848 |
| flgK | 2.447 | 0.795 | |
| flagellin/WP_014593940.1 | 3.053 | 0.952 | |
| fliZ | 2.607 | 1.012 | |
| fliL | 2.336 | 0.797 | |
| fligL | 2.2 | 0.881 | |
| fliM | 2.032 | 0.699 | |
| flhA | 2.086 | 1.488 | |
| flgG | 2.39 | 1.855 | |
| flagellar basal body P-ring protein | 2.104 | 1.542 | |
| fliS | 2.681 | 1.161 | |
| fliE | 2.293 | 1.921 | |
| flgN | 1.325 | 0.468 | |
| flgA | 2.279 | 1.357 | |
| flagellin/WP_013027859.1 | 1.6 | 4.051 | |
| flagellar basal body rod modification protein | 2.779 | 1.991 | |
| motB | 1.728 | 0.595 | |
| flit | 2.475 | 1.313 | |
| flgF | 2.508 | 1.697 | |
| flgJ | 1.934 | 1.267 | |
| flhB | 2.199 | 1.559 | |
| flgE | 2.392 | 1.892 | |
| flgC | 2.743 | 2.05 | |
| flgL | 2.201 | 0.881 | |
| flagellar basal body L-ring protein | 2.487 | 1.699 | |
| flgB | 2.875 | 1.96 | |
| flip | 1.89 | 0.595 | |
| Cell adhesion and biofilm formation | fliD/WP_014593941.1 | 2.93 | 1.104 |
| bcsO | 1.136 | 0.699 | |
| yhjQ | 0.942 | 0.717 | |
| Gene regulators | luxR | 0.816 | 1.276 |
| Fis family transcriptional regulator | 0.874 | 0.961 | |
| transcriptional regulator/WP_028714798.1 | 1.669 | 0.877 | |
| transcriptional regulator/WP_013027687.1 | 0.589 | 0.639 | |
| proQ | 2.497 | 0.534 | |
| luxI | 0.734 | 6.21 | |
| dgdR | 1.747 | −0.55 | |
| Methyl accepting proteins and Chemotaxis | WP_019105711.1 | 2.45 | 1.955 |
| WP_028714945.1 | 0.994 | 0.683 | |
| WP_050442519.1 | 1.888 | 0.734 | |
| WP_014605659.1 | 3.141 | 0.869 | |
| exbD | 2.259 | 1.512 | |
| chemotaxis protein-glutamate O-methyltransferase | 2.259 | 0.761 | |
| WP_014605660.1 | 2.716 | 0.882 | |
| chemotaxis response regulator (cheZ) | 2.194 | 0.717 | |
| Regulator of chemotaxis (cheA) | 1.888 | 0.743 | |
| chemotaxis response regulator (cheY) | 2.497 | 0.777 | |
| Outer membrane proteins | ligand-gated channel protein | 1.061 | 1.621 |
| lpoA | 1.102 | 0.431 | |
| Outer Membrane protein/ WP_026020991.1 | 1.806 | −0.929 | |
| Cell wall synthesis | endopeptidase | 2.145 | 0.907 |
| D-alanyl-D-alanine carboxypeptidase | 1.618 | 0.953 | |
| Stress response | hslV | 1.741 | 1.539 |
| carbamoyl phosphate synthase large subunit | 1.053 | 1.514 | |
| universal stress protein B | 1.457 | −0.751 | |
| Transport | magnesium-translocating P-type ATPase | 2.884 | 1.793 |
| MFS transporter/WP_028715176.1 | 2.149 | 1.176 | |
| pyridine nucleotide-disulphide oxidoreductase | 2.32 | 1.387 | |
| anion permease | 2.394 | 1.498 | |
| ligand-gated channel protein/WP_014595119.1 | 1.491 | 0.954 | |
| MFS transporter/WP_028715950.1 | 1.363 | 0.958 | |
| NCS2 family permease | 0.687 | 0.836 | |
| pyridine nucleotide-disulphide oxidoreductase | 2.32 | 1.387 | |
| glucose dehydrogenase | 0.732 | 0.544 | |
| WP_014593931.1/Transketolase | 1.267 | 1.129 | |
| sugar ABC transporter substrate-binding protein | 1.686 | 0.847 | |
| ferrous iron transporter A | 0.928 | −2.909 | |
| Hypothetical proteins | WP_013026151.1 | 2.51 | 2.214 |
| WP_019106310.1 | 1.013 | 0.601 | |
| WP_014593929.1 | 1.724 | 1.689 | |
| WP_014333251.1 | 1.197 | 0.612 | |
| WP_014604614.1 | 1.65 | 1.154 | |
| WP_028715174.1 | 1.039 | 0.632 | |
| WP_013027988.1 | 0.902 | 0.654 | |
| WP_013026097.1 | 1.84 | 1.293 | |
| WP_013024239.1 | 0.998 | 0.882 | |
| WP_014593930.1 | 0.902 | 0.847 | |
| WP_028715108.1 | 0.703 | 0.814 | |
| WP_050442523.1 | 0.955 | 1.876 | |
| WP_013025497.1 | 0.671 | 0.711 | |
| WP_014332946.1(putative motility protein) | 3.038 | 0.542 | |
| WP_013025421.1 | 3.227 | 0.923 | |
| WP_033765947.1 | 2.576 | 1.051 | |
| WP_028715248.1 | 1.979 | 1.442 | |
| WP_028715109.1 | 1.95 | 1.442 | |
| WP_014593989.1 | 1.376 | 0.594 | |
| WP_013024991.1 | 1.996 | −0.63 | |
| WP_014604946.1 | 0.597 | −0.777 | |
| WP_028715989.1 | 0.77 | −0.76 | |
| WP_050442541.1 | 1.443 | −2.11 | |
| WP_013025383.1 | 1.579 | −0.98 | |
| WP_028716027.1 | 1.033 | −0.715 | |
| Other | srfB (virulence factor) | 1.681 | 1.21 |
| Maa | 2.816 | 2.347 | |
| N6-hydroxylysine O-acetyltransferase | 1.74 | 0.936 | |
| Aminotransferase | 1.281 | 1.153 | |
| acyl—CoA ligase | 1.049 | 1.111 | |
| dTDP-4-dehydrorhamnose 3,5-epimerase | 1.467 | 1.13 | |
| traF | 1.88 | 0.524 | |
| peptidase C39 | 0.831 | 1.103 | |
| dusB | 0.983 | 0.713 | |
| cytochrome ubiquinol oxidase subunit II | 0.698 | 1.17 | |
| 2,5-didehydrogluconate reductase A | 1.438 | −0.598 | |
| aspartyl β-hydroxylase | 1.016 | −1.295 | |
| DNA polymerase III subunit ε | 1.574 | −0.854 | |
| Genes down regulated between the wild-type strain LMG 2665T in the presence of AHLs (at OD600 = 0.5) versus the same strain in the absence of AHLs (OD600 = 0.2). | |||
| Metabolism | erythrose-4-phosphate dehydrogenase | −0.977 | −0.762 |
| apbE | −0.62 | −1.113 | |
| nicotinate-nucleotide diphosphorylase | −1.559 | −0.78 | |
| dGTPase | −0.498 | −0.44 | |
| suhB | −0.649 | −0.472 | |
| L-aspartate oxidase | −1.706 | −0.601 | |
| Protease | −1.64 | −0.711 | |
| cystathionine β-lyase | −0.789 | −0.575 | |
| peptidylprolyl isomerase | −0.722 | 0.624 | |
| betaine-aldehyde dehydrogenase | −2.498 | 0.571 | |
| choline dehydrogenase | −2.154 | 0.674 | |
| 5-oxopent-3-ene-1,2,5-tricarboxylate decarboxylase | −0.837 | 2.096 | |
| Outer membrane proteins | ompC/WP_013026835.1 | −1.236 | −2.393 |
| Stress response | FAD/NAD(P) binding domain-containing protein | −0.93 | −1.039 |
| GNAT family N-acetyltransferase | −0.751 | −0.596 | |
| class C β-lactamase | −0.953 | −0.743 | |
| exodeoxyribonuclease I | −0.533 | −0.487 | |
| NUDIX hydrolase | −0.751 | −0.759 | |
| htpG | −1.09 | 0.877 | |
| groEL | −0.921 | 0.814 | |
| dnaJ | −0.84 | 0.742 | |
| sulfurtransferase | −0.733 | 0.971 | |
| hslU | −0.619 | 0.473 | |
| Transport | MFS transporter/WP_033765526.1 | −2.103 | −0.716 |
| sulfate transporter subunit | −1.613 | −0.567 | |
| C4-dicarboxylate ABC transporter | −1.445 | −0.993 | |
| MATE family efflux transporter | −1.266 | −0.806 | |
| tcyP | −0.908 | −1.025 | |
| MFS transporter/WP_028714804.1 | −0.957 | −0.716 | |
| ABC transporter permease | −0.712 | −0.522 | |
| flavocytochrome c | −0.712 | −0.934 | |
| microcin B17 transporter | −0.954 | 1.872 | |
| MFS transporter/WP_014598266.1 | −1.048 | 1.327 | |
| MFS transporter/WP_013024876.1 | −0.701 | 1.31 | |
| nickel transporter | −0.58 | 1.433 | |
| Gene regulators | DNA-binding response regulator | −0.6 | −0.603 |
| lysR/WP_028714815.1 | −1.675 | −0.718 | |
| draR | −0.974 | −1.295 | |
| lclR | −0.862 | −1.004 | |
| pbsX | −0.863 | −0.758 | |
| DNA-binding response regulator | −0.734 | 1.974 | |
| yqeI | −0.747 | 1.789 | |
| cdaR | −1.262 | 0.891 | |
| ompR | −0.734 | 1.974 | |
| Hypothetical proteins | WP_028715941.1 | −1.129 | −1.493 |
| WP_014593863.1 | −1.269 | −0.683 | |
| WP_028715707.1 | −1.481 | −1.226 | |
| WP_026021031.1 | −1.69 | −1.957 | |
| WP_028715464.1 | −0.864 | −1.032 | |
| WP_014594750.1 | −0.711 | −0.535 | |
| WP_028714922.1 | −1.067 | −1.138 | |
| WP_014605434.1 | −1.964 | −1.308 | |
| WP_014606741.1 | −1.173 | −0.939 | |
| WP_028715967.1 | −0.788 | −0.779 | |
| WP_028715704.1 | −0.939 | −0.922 | |
| WP_013024364.1 | −0.83 | −0.595 | |
| WP_028715521.1 | −0.742 | −0.578 | |
| WP_028715342.1 | −0.874 | −1.013 | |
| WP_050442548.1 | −1.663 | −1.265 | |
| WP_026021018.1 | −1.426 | −0.732 | |
| WP_014593149.1 | −1.931 | −1.472 | |
| WP_013026749.1 | −0.722 | 1.177 | |
| WP_022622675.1 | −0.63 | 1.053 | |
| WP_013024276.1 | −1.146 | 0.744 | |
| Cell wall synthesis | penicillin-binding protein 2 | −0.598 | 1.418 |
| ftsI | −0.726 | 1.406 | |
| murJ | −0.562 | 0.549 | |
| D-alanyl-D-alanine carboxypeptidase/WP_013026332.1 | −1.335 | 3.127 | |
| N-acetylmuramoyl-L-alanine amidase | −1.08 | 1.914 | |
| murein transglycosylase B | −1.155 | 0.608 | |
| Other | FUSC family protein | −0.918 | −0.704 |
| quinolinate synthetase | −1.122 | −0.863 | |
| dGTPase | −0.498 | −0.44 | |
| diaA | −0.658 | 0.527 | |
| U32 family peptidase | −0.641 | 0.884 | |
| peptidylprolyl isomerase | −0.722 | 0.624 | |
| K+/H+ antiporter | −0.727 | 0.879 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sibanda, S.; Kwenda, S.; Tanui, C.K.; Shyntum, D.Y.; Coutinho, T.A.; Moleleki, L.N. Transcriptome Profiling Reveals the EanI/R Quorum Sensing Regulon in Pantoea Ananatis LMG 2665T. Genes 2018, 9, 148. https://doi.org/10.3390/genes9030148
Sibanda S, Kwenda S, Tanui CK, Shyntum DY, Coutinho TA, Moleleki LN. Transcriptome Profiling Reveals the EanI/R Quorum Sensing Regulon in Pantoea Ananatis LMG 2665T. Genes. 2018; 9(3):148. https://doi.org/10.3390/genes9030148
Chicago/Turabian StyleSibanda, Siphathele, Stanford Kwenda, Collins K. Tanui, Divine Y. Shyntum, Teresa A. Coutinho, and Lucy N. Moleleki. 2018. "Transcriptome Profiling Reveals the EanI/R Quorum Sensing Regulon in Pantoea Ananatis LMG 2665T" Genes 9, no. 3: 148. https://doi.org/10.3390/genes9030148
APA StyleSibanda, S., Kwenda, S., Tanui, C. K., Shyntum, D. Y., Coutinho, T. A., & Moleleki, L. N. (2018). Transcriptome Profiling Reveals the EanI/R Quorum Sensing Regulon in Pantoea Ananatis LMG 2665T. Genes, 9(3), 148. https://doi.org/10.3390/genes9030148