Novel Sequence Features of DNA Repair Genes/Proteins from Deinococcus Species Implicated in Protection from Oxidatively Generated Damage


Abstract
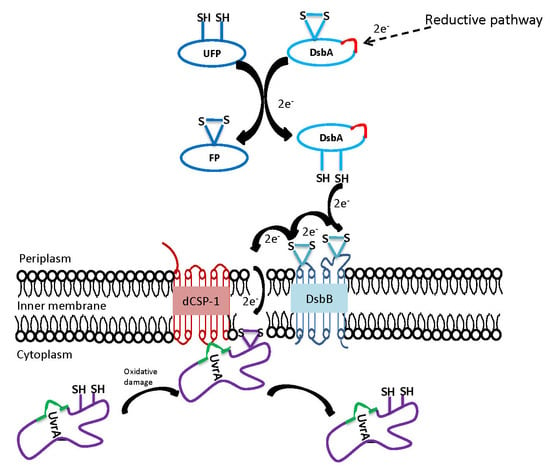
:
1. Introduction
2. Materials and Methods
2.1. Identification of CSIs in DNA Repair Proteins
2.2. Protein-Protein Interactions (PPIs) and Genetic Neighborhood Analyses
2.3. Homology Modeling of the Uvra1 Protein and Other Proteins from Deinococcus Species
3. Results
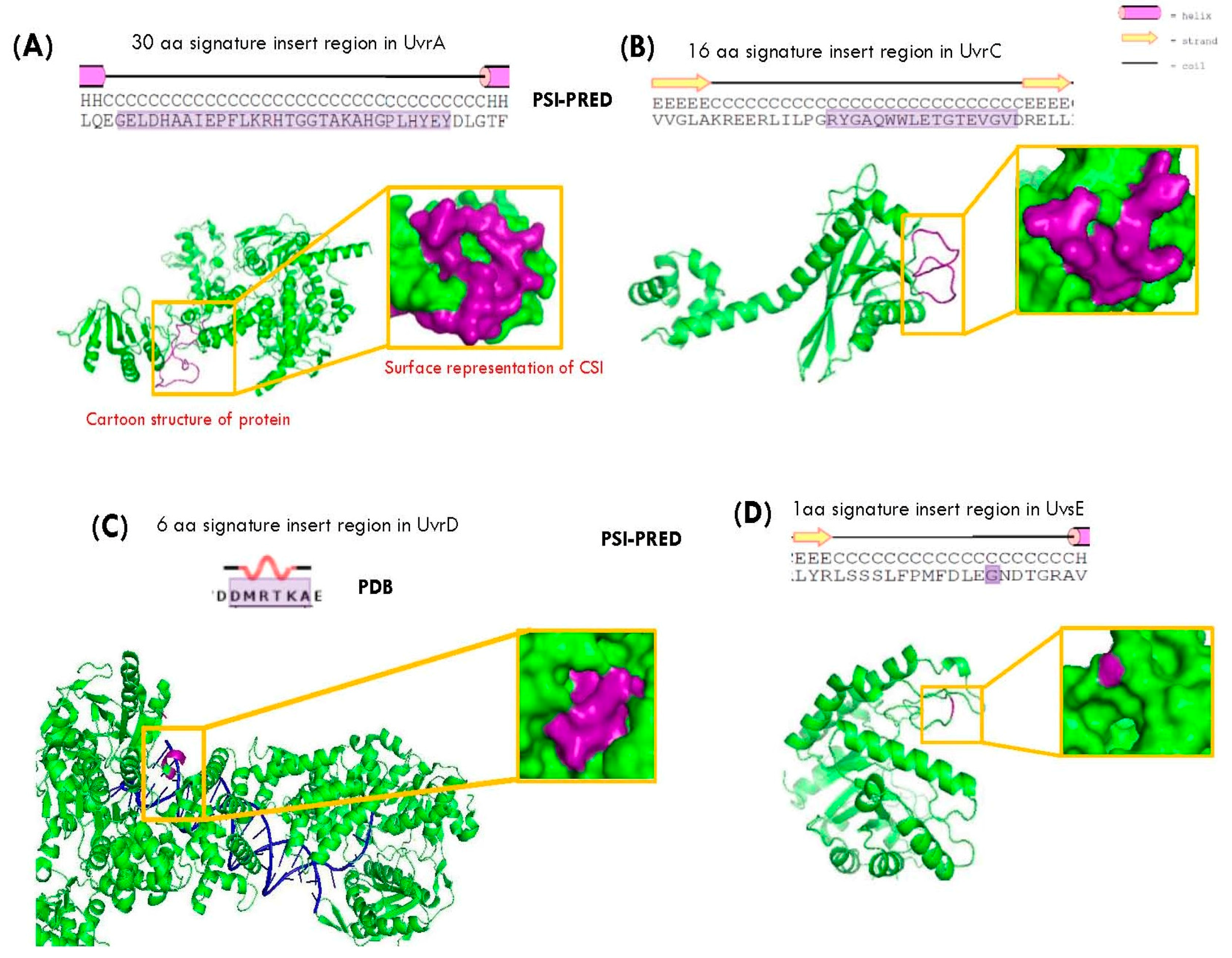
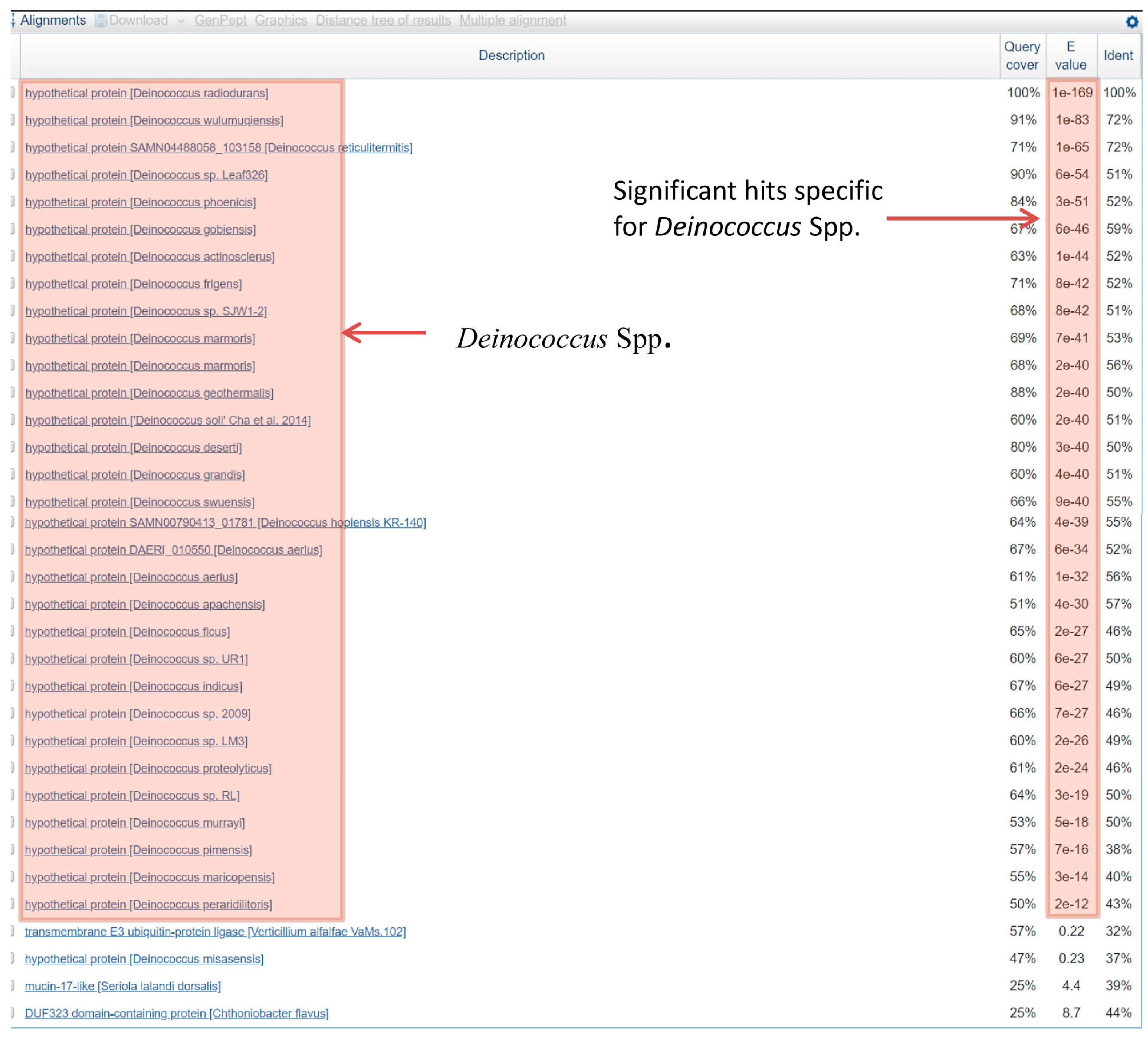
3.1. Conserved Signature Indels in DNA Repair Proteins Specific for the Deinococcus Species
3.2. Locations of the Conserved Indels (CSIs) in the Structures of the Proteins
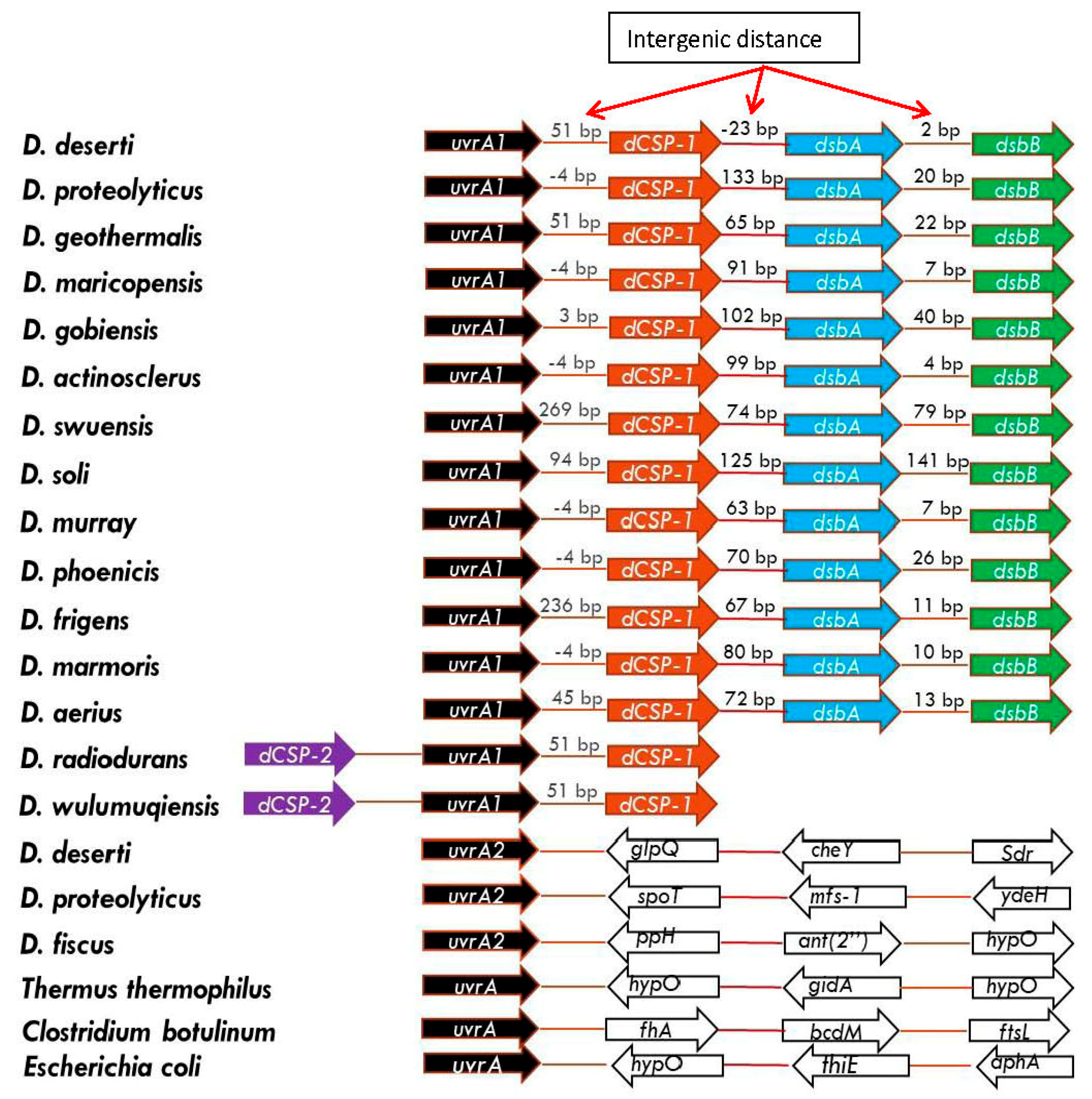
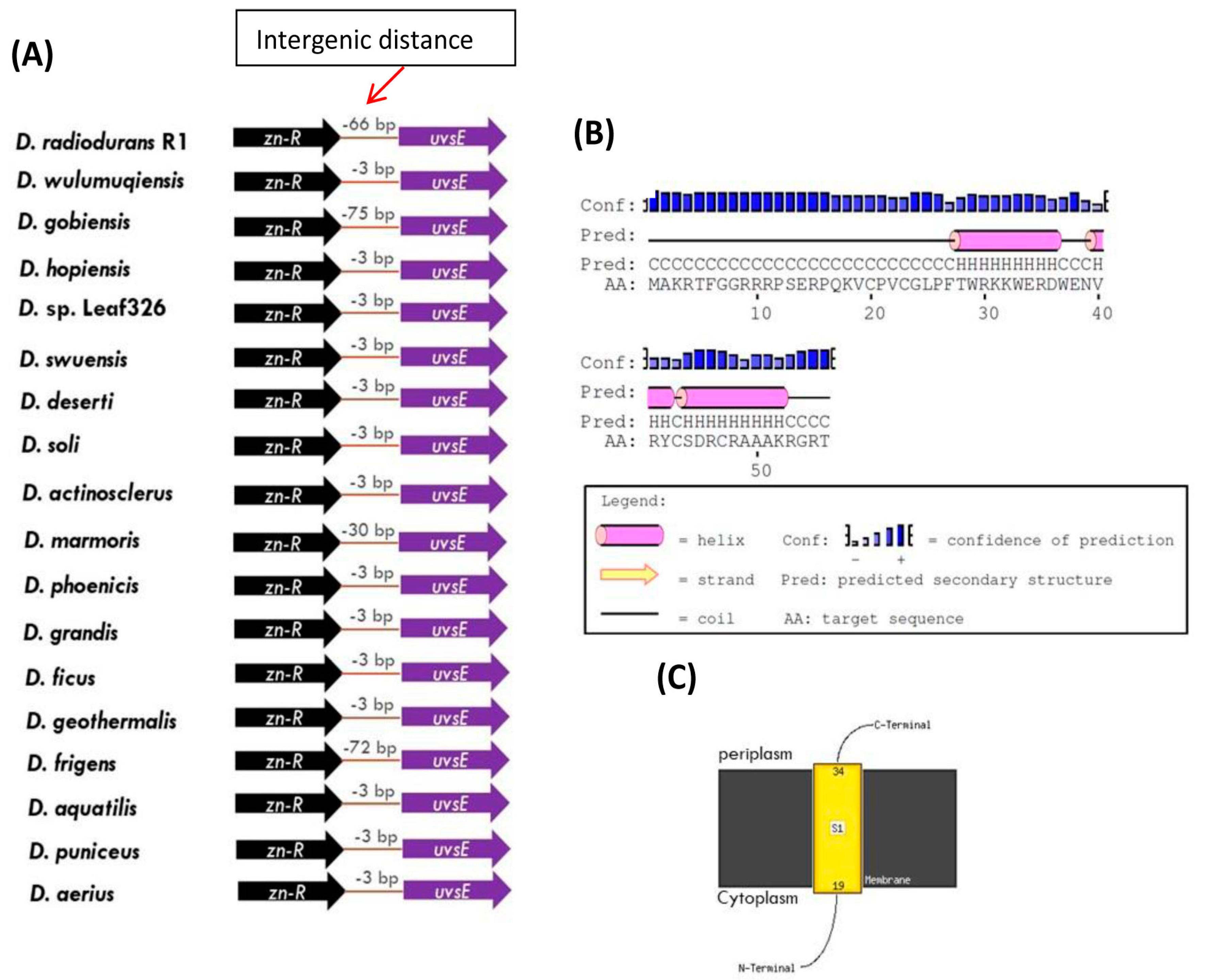
3.3. Novel Genomic Organization-Linkage of the Genes for UvrA1 and UvsE Proteins in Deinococcus Species
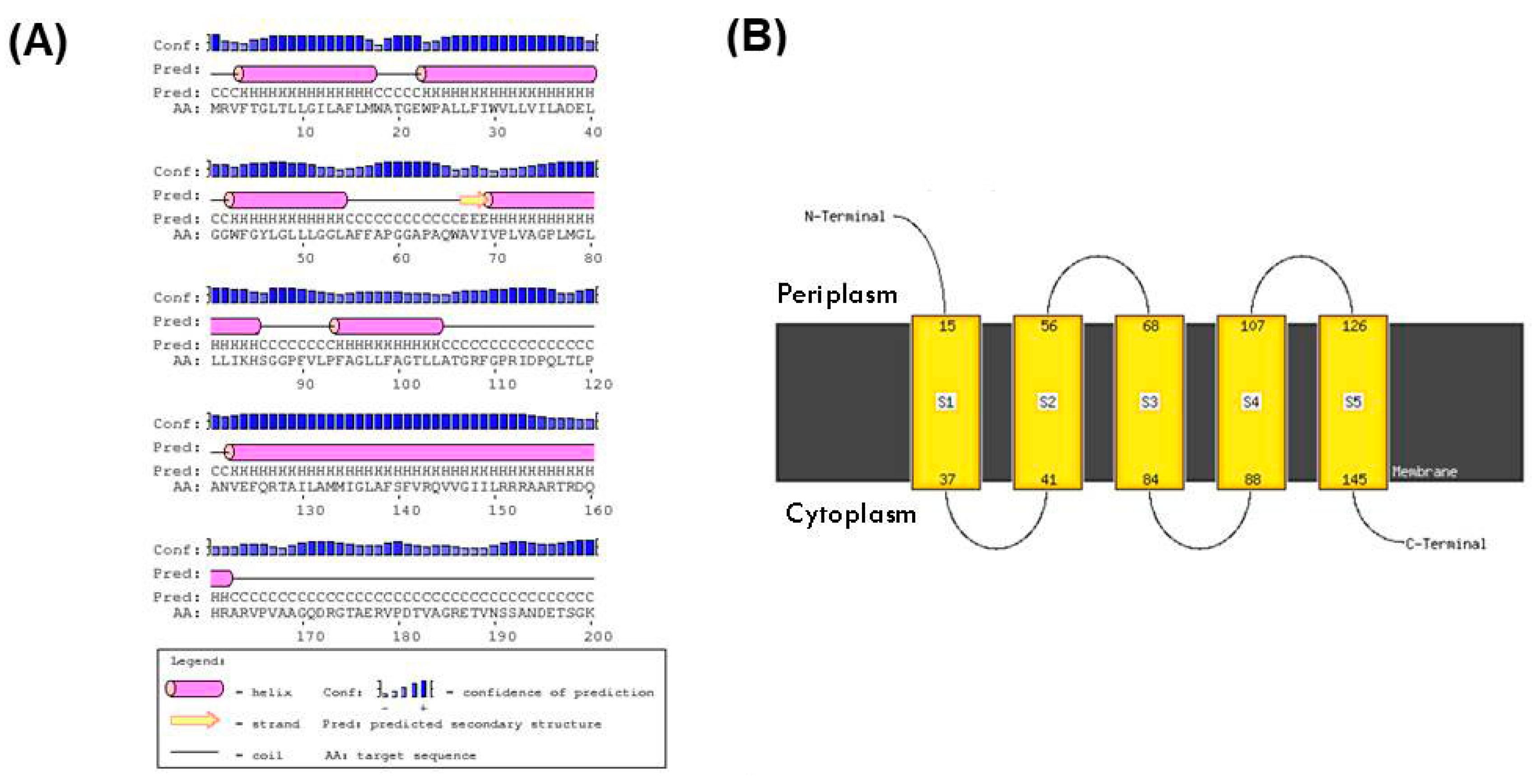
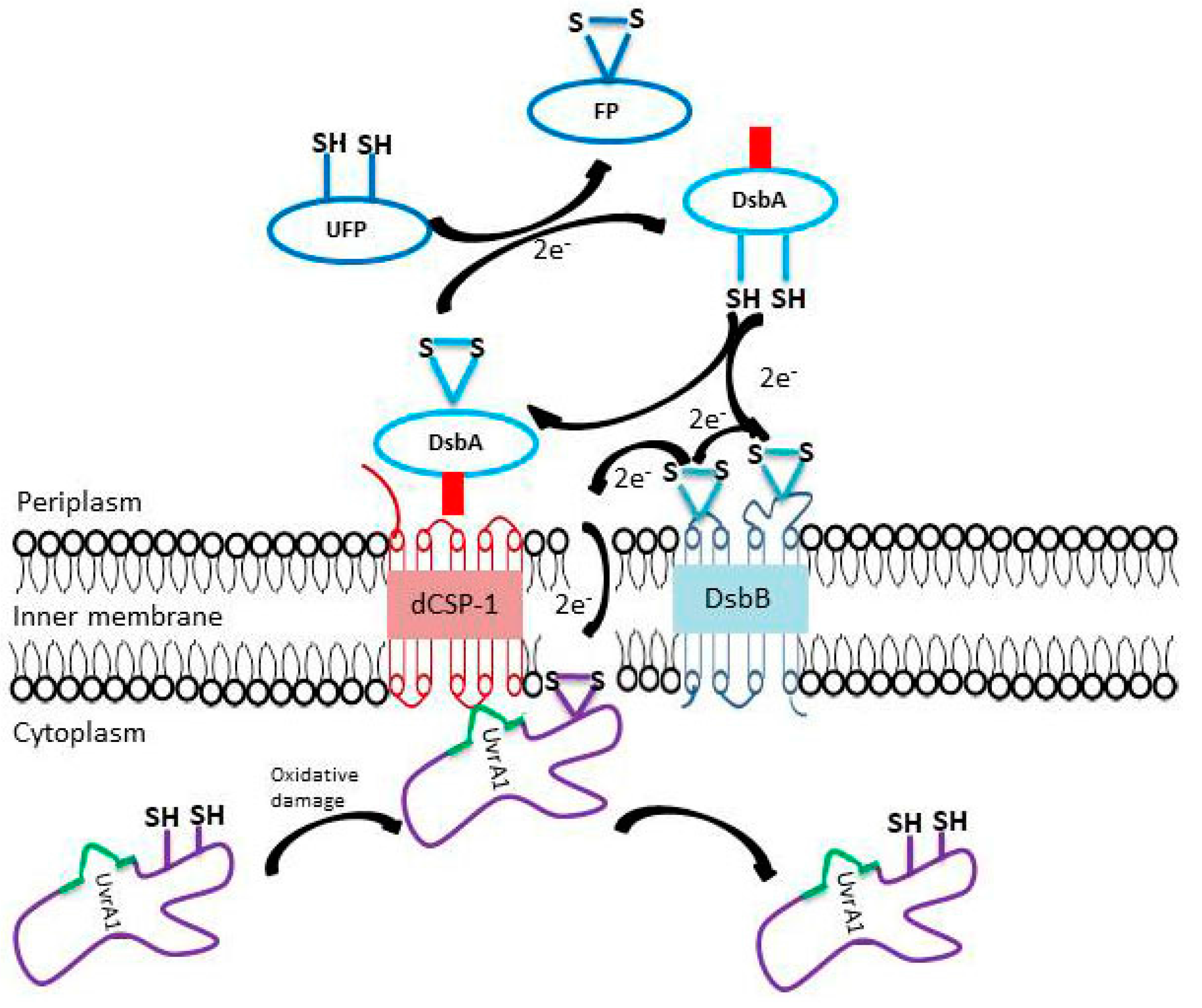
3.4. Structural and Biochemical Characteristics of the Proteins Linked to the UvrA1 Protein
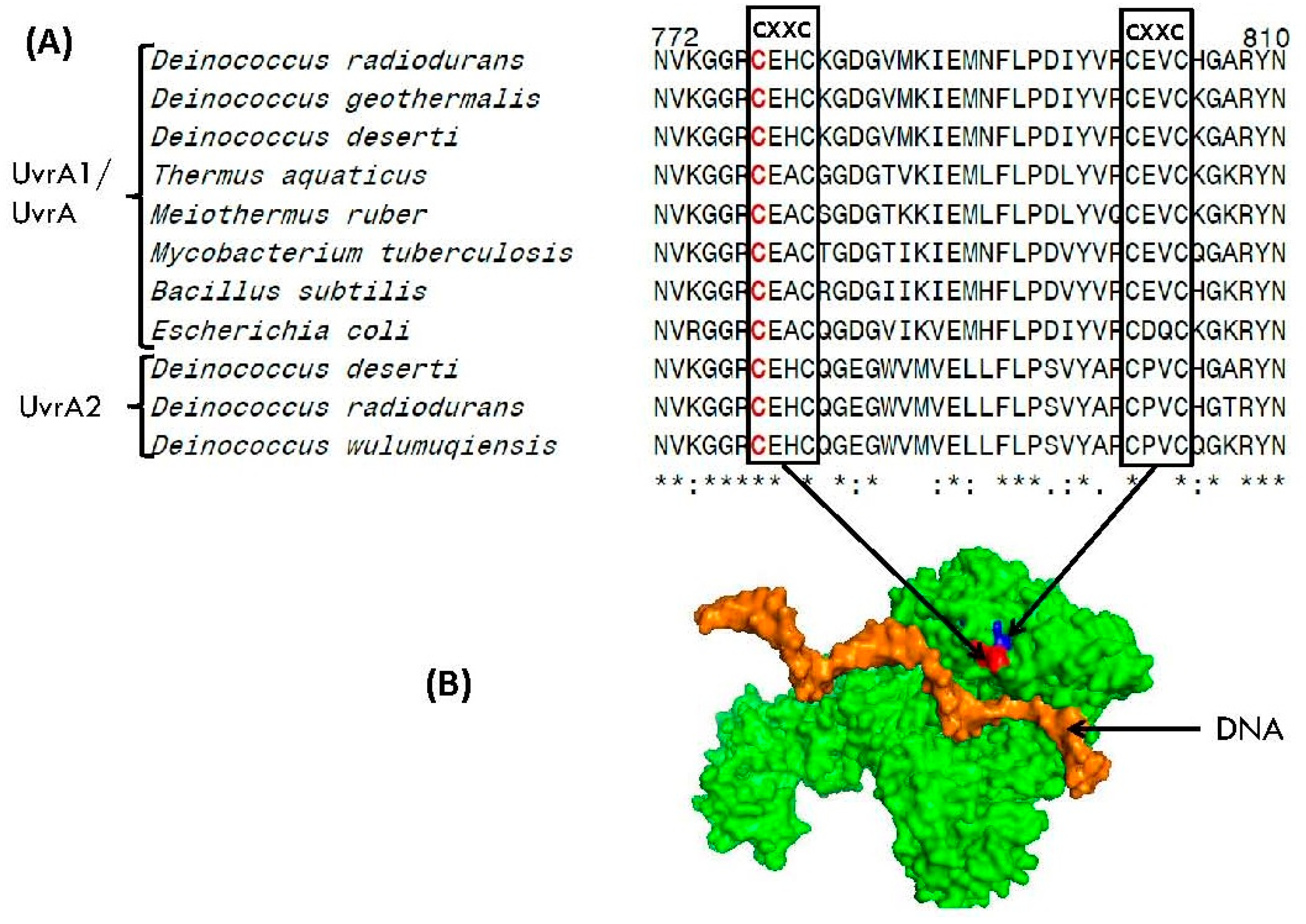
3.5. Presence of CXXC Motifs in DNA Repair Proteins
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Protein ID | # Of Cys Residues | CXXC Motifs | Figure Number |
|---|---|---|---|---|
| UvrA1 | DR1771 | 11 | 5 | Figure S17A |
| AlkA | DR2074 | 3 | - | - |
| Endonuclease V (Nfi) | DR2162 | 1 | - | - |
| MutM | DR0493 | 5 | 2 | Figure S17B |
| 8-Oxoguanine DNA glycosylase (MutY) | DR2285 | 6 | 1 | Figure S17C |
| Endonuclease III (Nth) | DR0928 | 6 | 1 | Figure S17D |
| Exonuclease III (Xth) | DR0354 | 2 | - | |
| UvrB | DR2275 | 6 | - | - |
| UvrC | DR1354 | 3 | - | - |
| UvsE | DR1819 | 4 | - | - |
| Rad25 | DRA0131 | 10 | 2 | Figure S17E |
| RecA | DR2340 | 1 | - | - |
| RecO | DR0819 | 4 | 2 | Figure S17F |
| RecR | DR0198 | 5 | 2 | Figure S17G |
| RecJ | DR1126 | 2 | - | - |
| RecN | DR1477 | 1 | - | - |
| RecQ | DR1289 | 9 | 1 | Figure S17H |
| RecD | DR1902 | 1 | - | - |
| SbcC | DR1922 | 4 | 1 | Figure S17I |
| SbcD | DR1921 | 1 | - | - |
| RuvA | DR1274 | 1 | - | - |
| RuvC | DR0440 | 2 | - | - |
| RecG | DR1916 | 4 | - | - |
| RadA | DR1105 | 5 | 2 | Figure S17J |
| Rad54 DNA helicase | DR1259 | 3 | - | - |
| DdrA | DR0423 | 2 | - | - |
| MutL | DR1696 | 4 | - | - |
| MutS | DR1039 | 4 | - | - |
| PolA | DR1707 | 5 | - | - |
| DNA polymerase III, α subunit (DnaE) | DR0507 | 14 | - | - |
| DNA polymerase III ε subunit (DnaQ) | DR0856 | 1 | - | - |
| DNA polymerase III subunit beta | DR0001 | 1 | - | - |
| DNA ligase (LigA) | DR2069 | 4 | 1 | Figure S17K |
| DNA polymerase III subunit gamma/tau | DR2410 | 7 | 1 | Figure S17L |
| UvrD | DR1775 | 1 | - | - |
| Gyrase A(GyrA) | DR1913 | 1 | - | - |
| Gyrase B(GyrB) | DR0906 | 2 | - | - |

References
- Battista, J.R.; Rainey, F.A.; Family, I. Deinococcaceae Brooks and Murray 1981, 356,VP emend. Rainey, Nobre, Schumann, Stackebrandt and da Costa 1997, 513. In Bergey’s Manual of Systematic Bacteriology Volume 1, The Archaea and the Deeply Branching and Phototrophic Bacteria; Boone, D.R., Castenholz, R.W., Eds.; Springer: New York, NY, USA, 2001; pp. 395–403. [Google Scholar]
- Battista, J.R.; Rainey, F.A.; Genus, I. Deinococcus Brooks and Murray 1981, 354,VP emend. Rainey, Nobre, Schumann, Stackebrandt and da Costa 1997, 513. In Bergey’s Manual of Systematic Bacteriology vol.1, The Archaea and the Deeply Branching and Phototrophic Bacteria; Boone, D.R., Castenholz, R.W., Eds.; Springer: New York, NY, USA, 2001; pp. 396–403. [Google Scholar]
- Rosenberg, E. The family Deinococcaceae. In The Prokaryotes—Other Major Lineages of Bacteria and the Archaea; Rosenberg, E., DeLong, E., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: New York, NY, USA, 2014; pp. 613–615. [Google Scholar]
- Albuquerque, L.; Costa, M.S. The family Thermaceae. In The Prokaryotes—Other Major Lineages of Bacteria and the Archaea; Rosenberg, E., DeLong, E., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: New York, NY, USA, 2014; pp. 955–987. [Google Scholar]
- Ho, J.; Adeolu, M.; Khadka, B.; Gupta, R.S. Identification of distinctive molecular traits that are characteristic of the phylum “Deinococcus-Thermus” and distinguish its main constituent groups. Syst. Appl. Microbiol. 2016, 39, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Weisburg, W.G.; Giovannoni, S.J.; Woese, C.R. The Deinococcus-Thermus phylum and the effect of rRNA composition on phylogenetic tree construction. Syst. Appl. Microbiol. 1989, 11, 128–134. [Google Scholar] [CrossRef]
- Cox, M.M.; Battista, J.R. Deinococcus radiodurans—The consummate survivor. Nat. Rev. Microbiol. 2005, 3, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Battista, J.R. Against all odds: The survival strategies of Deinococcus radiodurans. Annu. Rev. Microbiol. 1997, 51, 203–224. [Google Scholar] [CrossRef] [PubMed]
- Krisko, A.; Radman, M. Biology of extreme radiation resistance: The way of Deinococcus radiodurans. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Mattimore, V.; Battista, J.R. Radioresistance of Deinococcus radiodurans: Functions necessary to survive ionizing radiation are also necessary to survive prolonged desiccation. J. Bacteriol. 1996, 178, 633–637. [Google Scholar] [CrossRef] [PubMed]
- Timmins, J.; Moe, E. A Decade of Biochemical and Structural Studies of the DNA Repair Machinery of Deinococcus radiodurans: Major Findings, Functional and Mechanistic Insight and Challenges. Comput. Struct. Biotechnol. J. 2016, 14, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Slade, D.; Radman, M. Oxidative stress resistance in Deinococcus radiodurans. Microbiol. Mol. Biol. Rev. 2011, 75, 133–191. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.M.; Keck, J.L.; Battista, J.R. Rising from the Ashes: DNA Repair in Deinococcus radiodurans. PLoS Genet. 2010, 6, e1000815. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.J. A new perspective on radiation resistance based on Deinococcus radiodurans. Nat. Rev. Microbiol. 2009, 7, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Blasius, M.; Sommer, S.; Hubscher, U. Deinococcus radiodurans: What belongs to the survival kit? Crit. Rev. Biochem. Mol. Biol. 2008, 43, 221–238. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Aravind, L.; Wolf, Y.I.; Tatusov, R.L.; Minton, K.W.; Koonin, E.V.; Daly, M.J. Genome of the extremely radiation-resistant bacterium Deinococcus radiodurans viewed from the perspective of comparative genomics. Microbiol. Mol. Biol. Rev. 2001, 65, 44–79. [Google Scholar] [CrossRef] [PubMed]
- Battista, J.R.; Earl, A.M.; Park, M.J. Why is Deinococcus radiodurans so resistant to ionizing radiation? Trends Microbiol. 1999, 7, 362–365. [Google Scholar] [CrossRef]
- Minton, K.W. DNA repair in the extremely radioresistant bacterium Deinococcus radiodurans. Mol. Microbiol. 1994, 13, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Krisko, A.; Radman, M. Protein damage and death by radiation in Escherichia coli and Deinococcus radiodurans. Proc. Natl. Acad. Sci. USA 2010, 107, 14373–14377. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.J.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Leapman, R.D.; Lai, B.; Ravel, B.; Li, S.M.; Kemner, K.M.; et al. Protein oxidation implicated as the primary determinant of bacterial radioresistance. PLoS Biol. 2007, 5, e92. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Omelchenko, M.V.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Lapidus, A.; Copeland, A.; Kim, E.; Land, M.; et al. Deinococcus geothermalis: The pool of extreme radiation resistance genes shrinks. PLoS ONE 2007, 2, e955. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, E.; Gupta, R.S. Identification of signature proteins that are distinctive of the Deinococcus-Thermus phylum. Int. Microbiol. 2007, 10, 201–208. [Google Scholar] [PubMed]
- De Groot, A.; Dulermo, R.; Ortet, P.; Blanchard, L.; Guerin, P.; Fernandez, B.; Vacherie, B.; Dossat, C.; Jolivet, E.; Siguier, P.; et al. Alliance of proteomics and genomics to unravel the specificities of Sahara bacterium Deinococcus deserti. PLoS Genet. 2009, 5, e1000434. [Google Scholar] [CrossRef] [PubMed]
- Norais, C.A.; Chitteni-Pattu, S.; Wood, E.A.; Inman, R.B.; Cox, M.M. DdrB protein, an alternative Deinococcus radiodurans SSB induced by ionizing radiation. J. Biol. Chem. 2009, 284, 21402–21411. [Google Scholar] [CrossRef] [PubMed]
- Sugiman-Marangos, S.N.; Weiss, Y.M.; Junop, M.S. Mechanism for accurate, protein-assisted DNA annealing by Deinococcus radiodurans DdrB. Proc. Natl. Acad. Sci. USA 2016, 113, 4308–4313. [Google Scholar] [CrossRef] [PubMed]
- Kota, S.; Charaka, V.K.; Misra, H.S. PprA, a pleiotropic protein for radioresistance, works through DNA gyrase and shows cellular dynamics during postirradiation recovery in Deinococcus radiodurans. J. Genet. 2014, 93, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Selvam, K.; Duncan, J.R.; Tanaka, M.; Battista, J.R. DdrA, DdrD and PprA: Components of UV and mitomycin C resistance in Deinococcus radiodurans R1. PLoS ONE 2013, 8, e69007. [Google Scholar] [CrossRef] [PubMed]
- Ghosal, D.; Omelchenko, M.V.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Venkateswaran, A.; Zhai, M.; Kostandarithes, H.M.; Brim, H.; Makarova, K.S.; et al. How radiation kills cells: Survival of Deinococcus radiodurans and Shewanella oneidensis under oxidative stress. FEMS Microbiol. Rev. 2005, 29, 361–375. [Google Scholar] [PubMed]
- Du, J.; Gebicki, J.M. Proteins are major initial cell targets of hydroxyl free radicals. Int. J. Biochem. Cell Biol. 2004, 36, 2334–2343. [Google Scholar] [CrossRef] [PubMed]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A.; Linn, S. DNA damage and oxygen radical toxicity. Science 1988, 240, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Gaidamakova, E.K.; Grichenko, O.; Matrosova, V.Y.; Hoeke, V.; Klimenkova, P.; Conze, I.H.; Volpe, R.P.; Tkavc, R.; Gostinčar, C.; et al. Across the tree of life, radiation resistance is governed by antioxidant Mn(2+), gauged by paramagnetic resonance. Proc. Natl. Acad. Sci. USA 2017, 114, E9253–E9260. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.J.; Gaidamakova, E.K.; Matrosova, V.Y.; Kiang, J.G.; Fukumoto, R.; Lee, D.Y.; Wehr, N.B.; Viteri, G.A.; Berlett, B.S.; Levine, R.L. Small-molecule antioxidant proteome-shields in Deinococcus radiodurans. PLoS ONE 2010, 5, e12570. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.J.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Venkateswaran, A.; Wehr, N.B.; Viteri, G.A.; Berlett, B.S.; Levine, R.L. Accumulation of Mn(II) in Deinococcus radiodurans facilitates gamma-radiation resistance. Science 2004, 306, 1025–1028. [Google Scholar] [CrossRef] [PubMed]
- Shashidhar, R.; Kumar, S.A.; Misra, H.S.; Bandekar, J.R. Evaluation of the role of enzymatic and nonenzymatic antioxidant systems in the radiation resistance of Deinococcus. Can. J. Microbiol. 2010, 56, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, E.; Gupta, R.S. Distinctive protein signatures provide molecular markers and evidence for the monophyletic nature of the Deinococcus-Thermus phylum. J. Bacteriol. 2004, 186, 3097–3107. [Google Scholar] [CrossRef] [PubMed]
- Akiva, E.; Itzhaki, Z.; Margalit, H. Built-in loops allow versatility in domain-domain interactions: Lessons from self-interacting domains. Proc. Natl. Acad. Sci. USA 2008, 105, 13292–13297. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Gupta, R.S. Conserved inserts in the Hsp60 (GroEL) and Hsp70 (DnaK) proteins are essential for cellular growth. Mol. Genet. Genom. 2009, 281, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S.; Nanda, A.; Khadka, B. Novel molecular, structural and evolutionary characteristics of the phosphoketolases from bifidobacteria and Coriobacteriales. PLoS ONE 2017, 12, e0172176. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Panchenko, A.R. Mechanisms of protein oligomerization, the critical role of insertions and deletions in maintaining different oligomeric states. Proc. Natl. Acad. Sci. USA 2010, 107, 20352–20357. [Google Scholar] [CrossRef] [PubMed]
- Chandra, G.; Chater, K.F. Developmental biology of Streptomyces from the perspective of 100 actinobacterial genome sequences. FEMS Microbiol. Rev. 2014, 38, 345–379. [Google Scholar] [CrossRef] [PubMed]
- Ricci, D.P.; Melfi, M.D.; Lasker, K.; Dill, D.L.; McAdams, H.H.; Shapiro, L. Cell cycle progression in Caulobacter requires a nucleoid-associated protein with high AT sequence recognition. Proc. Natl. Acad. Sci. USA 2016, 113, E5952–E5961. [Google Scholar] [CrossRef] [PubMed]
- Lorenzini, E.; Singer, A.; Singh, B.; Lam, R.; Skarina, T.; Chirgadze, N.Y.; Savchenko, A.; Gupta, R.S. Structure and protein-protein interaction studies on Chlamydia trachomatis protein CT670 (YscO Homolog). J. Bacteriol. 2010, 192, 2746–2756. [Google Scholar] [CrossRef] [PubMed]
- Sugiman-Marangos, S.N.; Peel, J.K.; Weiss, Y.M.; Ghirlando, R.; Junop, M.S. Crystal structure of the DdrB/ssDNA complex from Deinococcus radiodurans reveals a DNA binding surface involving higher-order oligomeric states. Nucleic Acids Res. 2013, 41, 9934–9944. [Google Scholar] [CrossRef] [PubMed]
- Harris, D.R.; Tanaka, M.; Saveliev, S.V.; Jolivet, E.; Earl, A.M.; Cox, M.M.; Battista, J.R. Preserving genome integrity: The DdrA protein of Deinococcus radiodurans R1. PLoS Biol. 2004, 2, e304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kota, S.; Misra, H.S. PprA: A protein implicated in radioresistance of Deinococcus radiodurans stimulates catalase activity in Escherichia coli. Appl. Microbiol. Biotechnol. 2006, 72, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Truglio, J.J.; Croteau, D.L.; Van Houten, B.; Kisker, C. Prokaryotic nucleotide excision repair: The UvrABC system. Chem. Rev. 2006, 106, 233–252. [Google Scholar] [CrossRef] [PubMed]
- Petit, C.; Sancar, A. Nucleotide excision repair: From E. coli to man. Biochimie 1999, 81, 15–25. [Google Scholar] [CrossRef]
- Evans, D.M.; Moseley, B.E. Roles of the uvsC, uvsD, uvsE, and mtcA genes in the two pyrimidine dimer excision repair pathways of Deinococcus radiodurans. J. Bacteriol. 1983, 156, 576–583. [Google Scholar] [PubMed]
- Inaba, K.; Ito, K. Structure and mechanisms of the DsbB-DsbA disulfide bond generation machine. Biochim. Biophys. Acta 2008, 1783, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Shouldice, S.R.; Heras, B.; Walden, P.M.; Totsika, M.; Schembri, M.A.; Martin, J.L. Structure and function of DsbA, a key bacterial oxidative folding catalyst. Antioxid. Redox Signal. 2011, 14, 1729–1760. [Google Scholar] [CrossRef] [PubMed]
- Dutton, R.J.; Boyd, D.; Berkmen, M.; Beckwith, J. Bacterial species exhibit diversity in their mechanisms and capacity for protein disulfide bond formation. Proc. Natl. Acad. Sci. USA 2008, 105, 11933–11938. [Google Scholar] [CrossRef] [PubMed]
- Marino, S.M.; Gladyshev, V.N. Analysis and functional prediction of reactive cysteine residues. J. Biol. Chem. 2012, 287, 4419–4425. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Mueller, K.L.; Grossman, L. A mutational study of the C-terminal zinc-finger motif of the Escherichia coli UvrA protein. J. Biol. Chem. 1994, 269, 10771–10775. [Google Scholar] [PubMed]
- Pakotiprapha, D.; Inuzuka, Y.; Bowman, B.R.; Moolenaar, G.F.; Goosen, N.; Jeruzalmi, D.; Verdine, G.L. Crystal structure of Bacillus stearothermophilus UvrA provides insight into ATP-modulated dimerization, UvrB interaction, and DNA binding. Mol. Cell 2008, 29, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Croteau, D.L.; DellaVecchia, M.J.; Wang, H.; Bienstock, R.J.; Melton, M.A.; Van Houten, B. The C-terminal zinc finger of UvrA does not bind DNA directly but regulates damage-specific DNA binding. J. Biol. Chem. 2006, 281, 26370–26381. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S. Identification of Conserved Indels that are Useful for Classification and Evolutionary Studies. In Bacterial Taxonomy, Methods in Microbiology Volume 41; Goodfellow, M., Sutcliffe, I.C., Chun, J., Eds.; Elsevier: London, UK, 2014; pp. 153–182. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- White, O.; Eisen, J.A.; Heidelberg, J.F.; Hickey, E.K.; Peterson, J.D.; Dodson, R.J.; Haft, D.H.; Gwinn, M.L.; Nelson, W.C.; Richardson, D.L.; et al. Genome sequence of the radioresistant bacterium Deinococcus radiodurans R1. Science 1999, 286, 1571–1577. [Google Scholar] [CrossRef] [PubMed]
- Jeanmougin, F.; Thompson, J.D.; Gouy, M.; Higgins, D.G.; Gibson, T.J. Multiple sequence alignment with Clustal X. Trends Biochem. Sci. 1998, 23, 403–405. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Von Mering, C.; Jensen, L.J.; Snel, B.; Hooper, S.D.; Krupp, M.; Foglierini, M.; Jouffre, N.; Huynen, M.A.; Bork, P. STRING: Known and predicted protein-protein associations, integrated and transferred across organisms. Nucleic Acids Res. 2005, 33, D433–D437. [Google Scholar] [CrossRef] [PubMed]
- Sali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef] [PubMed]
- Buchan, D.W.; Minneci, F.; Nugent, T.C.; Bryson, K.; Jones, D.T. Scalable web services for the PSIPRED Protein Analysis Workbench. Nucleic Acids Res. 2013, 41, W349–W357. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.W.; Prlic, A.; Bi, C.; Bluhm, W.F.; Christie, C.H.; Dutta, S.; Green, R.K.; Goodsell, D.S.; Westbrook, J.D.; Woo, J.; et al. The RCSB Protein Data Bank: Views of structural biology for basic and applied research and education. Nucleic Acids Res. 2015, 43, D345–D356. [Google Scholar] [CrossRef] [PubMed]
- Pakotiprapha, D.; Samuels, M.; Shen, K.; Hu, J.H.; Jeruzalmi, D. Structure and mechanism of the UvrA-UvrB DNA damage sensor. Nat. Struct. Mol. Biol. 2012, 19, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Karakas, E.; Truglio, J.J.; Croteau, D.; Rhau, B.; Wang, L.; Van Houten, B.; Kisker, C. Structure of the C-terminal half of UvrC reveals an RNase H endonuclease domain with an Argonaute-like catalytic triad. EMBO J. 2007, 26, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Meulenbroek, E.M.; Peron, C.C.; Jala, I.; Iwai, S.; Moolenaar, G.F.; Goosen, N.; Pannu, N.S. UV damage endonuclease employs a novel dual-dinucleotide flipping mechanism to recognize different DNA lesions. Nucleic Acids Res. 2013, 41, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.Y.; Sali, A. Statistical potential for assessment and prediction of protein structures. Protein Sci. 2006, 15, 2507–2524. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Zhang, Y. Improving the physical realism and structural accuracy of protein models by a two-step atomic-level energy minimization. Biophys. J. 2011, 101, 2525–2534. [Google Scholar] [CrossRef] [PubMed]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef] [PubMed]
- Sippl, M.J. Recognition of errors in three-dimensional structures of proteins. Proteins 1993, 17, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Lovell, S.C.; Davis, I.W.; Arendall, W.B., III; de Bakker, P.I.; Word, J.M.; Prisant, M.G.; Richardson, J.S.; Richardson, D.C. Structure validation by Calpha geometry: phi, psi and Cbeta deviation. Proteins 2003, 50, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Colovos, C.; Yeates, T.O. Verification of protein structures: Patterns of nonbonded atomic interactions. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, D.; Luthy, R.; Bowie, J.U. VERIFY3D: Assessment of protein models with three-dimensional profiles. Methods Enzymol. 1997, 277, 396–404. [Google Scholar] [PubMed]
- Benkert, P.; Tosatto, S.C.; Schomburg, D. QMEAN: A comprehensive scoring function for model quality assessment. Proteins 2008, 71, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Moseley, B.E.; Evans, D.M. Isolation and properties of strains of Micrococcus (Deinococcus) radiodurans unable to excise ultraviolet light-induced pyrimidine dimers from DNA: Evidence for two excision pathways. J. Gen. Microbiol. 1983, 129, 2437–2445. [Google Scholar] [CrossRef] [PubMed]
- Stelter, M.; Acajjaoui, S.; McSweeney, S.; Timmins, J. Structural and mechanistic insight into DNA unwinding by Deinococcus radiodurans UvrD. PLoS ONE 2013, 8, e77364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmins, J.; Gordon, E.; Caria, S.; Leonard, G.; Acajjaoui, S.; Kuo, M.S.; Monchois, V.; McSweeney, S. Structural and mutational analyses of Deinococcus radiodurans UvrA2 provide insight into DNA binding and damage recognition by UvrAs. Structure 2009, 17, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Narumi, I.; Funayama, T.; Kikuchi, M.; Watanabe, H.; Matsunaga, T.; Nikaido, O.; Yamamoto, K. Characterization of pathways dependent on the uvsE, uvrA1, or uvrA2 gene product for UV resistance in Deinococcus radiodurans. J. Bacteriol. 2005, 187, 3693–3697. [Google Scholar] [CrossRef] [PubMed]
- Dam, P.; Olman, V.; Harris, K.; Su, Z.; Xu, Y. Operon prediction using both genome-specific and general genomic information. Nucleic Acids Res. 2007, 35, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Hagelsieb, G. The power of operon rearrangements for predicting functional associations. Comput. Struct. Biotechnol. J. 2015, 13, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Ma, Q.; Zhou, C.; Chen, X.; Zhang, H.; Yang, J.; Mao, F.; Lai, W.; Xu, Y. DOOR 2.0: Presenting operons and their functions through dynamic and integrated views. Nucleic Acids Res. 2014, 42, D654–D659. [Google Scholar] [CrossRef] [PubMed]
- Dandekar, T.; Snel, B.; Huynen, M.; Bork, P. Conservation of gene order: A fingerprint of proteins that physically interact. Trends Biochem. Sci. 1998, 23, 324–328. [Google Scholar] [CrossRef]
- Overbeek, R.; Fonstein, M.; D’Souza, M.; Pusch, G.D.; Maltsev, N. The use of gene clusters to infer functional coupling. Proc. Natl. Acad. Sci. USA 1999, 96, 2896–2901. [Google Scholar] [CrossRef] [PubMed]
- Inaba, K.; Murakami, S.; Suzuki, M.; Nakagawa, A.; Yamashita, E.; Okada, K.; Ito, K. Crystal structure of the DsbB-DsbA complex reveals a mechanism of disulfide bond generation. Cell 2006, 127, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Jaciuk, M.; Nowak, E.; Skowronek, K.; Tanska, A.; Nowotny, M. Structure of UvrA nucleotide excision repair protein in complex with modified DNA. Nat. Struct. Mol. Biol. 2011, 18, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Wagner, K.; Moolenaar, G.F.; Goosen, N. Role of the insertion domain and the zinc-finger motif of Escherichia coli UvrA in damage recognition and ATP hydrolysis. DNA Repair 2011, 10, 483–496. [Google Scholar] [CrossRef] [PubMed]
- Radman, M. Protein damage, radiation sensitivity and aging. DNA Repair 2016, 44, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Schoeffler, A.J.; May, A.P.; Berger, J.M. A domain insertion in Escherichia coli GyrB adopts a novel fold that plays a critical role in gyrase function. Nucleic Acids Res. 2010, 38, 7830–7844. [Google Scholar] [CrossRef] [PubMed]
- Cumming, R.C.; Andon, N.L.; Haynes, P.A.; Park, M.; Fischer, W.H.; Schubert, D. Protein disulfide bond formation in the cytoplasm during oxidative stress. J. Biol. Chem. 2004, 279, 21749–21758. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. How oxygen damages microbes: Oxygen tolerance and obligate anaerobiosis. Adv. Microb. Physiol. 2002, 46, 111–153. [Google Scholar] [PubMed]
- Dean, C.J.; Alexander, P. Sensitization of radio-resistant bacteria to X-rays by iodoacetamide. Nature 1962, 196, 1324–1326. [Google Scholar] [CrossRef]
- Dardalhon-Samsonoff, M.; Rebeyrotte, N. Role of DNA-membrane attachment in repair of radiation damage in Micrococcus radiodurans. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1975, 27, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Vukovic-Nagy, B.; Fox, B.W.; Fox, M. The release of a deoxyribonucleic acid fragment after x-irradiation of Micrococcus radiodurans. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1974, 25, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Duggan, D.E.; Anderson, A.W.; Elliker, P.R. Inactivation of the radiation-resistant spoilage bacterium Micrococcus radiodurans. II. Radiation inactivation rates as influenced by menstruum temperature, preirradiation heat treatment, and certain reducing agents. Appl. Microbiol. 1963, 11, 413–417. [Google Scholar] [PubMed]
- Joe, M.H.; Jung, S.W.; Im, S.H.; Lim, S.Y.; Song, H.P.; Kwon, O.; Kim, D.H. Genome-wide response of Deinococcus radiodurans on cadmium toxicity. J. Microbiol. Biotechnol. 2011, 21, 438–447. [Google Scholar] [PubMed]
- Burrell, A.D.; Feldschreiber, P.; Dean, C.J. DNA-membrane association and the repair of double breaks in x-irradiated Micrococcus radiodurans. Biochim. Biophys. Acta 1971, 247, 38–53. [Google Scholar] [CrossRef]
- Driedger, A.A. Are there multiple attachments between bacterial DNA and the cell membrane? Can. J. Microbiol. 1970, 16, 881–882. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.G.; Kovalsky, O.; Grossman, L. DNA damage-dependent recruitment of nucleotide excision repair and transcription proteins to Escherichia coli inner membranes. Nucleic Acids Res. 1997, 25, 3151–3158. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Inaba, K. The disulfide bond formation (Dsb) system. Curr. Opin. Struct. Biol. 2008, 18, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Bardwell, J.C.; McGovern, K.; Beckwith, J. Identification of a protein required for disulfide bond formation in vivo. Cell 1991, 67, 581–589. [Google Scholar] [CrossRef]
- Kojer, K.; Riemer, J. Balancing oxidative protein folding: The influences of reducing pathways on disulfide bond formation. Biochim. Biophys. Acta 2014, 1844, 1383–1390. [Google Scholar] [CrossRef] [PubMed]
- Narumi, I.; Satoh, K.; Kikuchi, M.; Funayama, T.; Kitayama, S.; Yanagisawa, T.; Watanabe, H.; Yamamoto, K. Molecular analysis of the Deinococcus radiodurans recA locus and identification of a mutation site in a DNA repair-deficient mutant, rec30. Mutat. Res. 1999, 435, 233–243. [Google Scholar] [CrossRef]
- Masure, H.R.; Pearce, B.J.; Shio, H.; Spellerberg, B. Membrane targeting of RecA during genetic transformation. Mol. Microbiol. 1998, 27, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Agostini, H.J.; Carroll, J.D.; Minton, K.W. Identification and characterization of uvrA, a DNA repair gene of Deinococcus radiodurans. J. Bacteriol. 1996, 178, 6759–6765. [Google Scholar] [CrossRef] [PubMed]
- Lagkouvardos, I.; Jehl, M.A.; Rattei, T.; Horn, M. Signature protein of the PVC superphylum. Appl. Environ. Microbiol. 2014, 80, 440–445. [Google Scholar] [CrossRef] [PubMed]









| Protein Name | Repair Pathway | Protein ID a | Indel Size | Indel b Position | Figure |
|---|---|---|---|---|---|
| UvrA1 | NER | DR1771 | 30 aa ins | 421–514 | Figure 1A |
| UvrC | NER | DR1354 | 16 aa ins | 459–535 | Figure 1B |
| UvrD | MP | DR1775 | 6 aa ins | 516–576 | Figure 2A |
| UvsE | UVER | DR1819 | 1 aa ins | 48–125 | Figure 2B |
| MutY | BER | DR2285 | 4 aa ins | 24–82 | Figure S1 |
| MutM | BER | DR0493 | 2 aa del | 168–225 | Figure S2 |
| Endonuclease III (Nth) | BER | DR0928 | 2 aa ins | 144–228 | Figure S3 |
| RecA c | HR | DR2340 | 1 aa ins | 216–280 | Figure S4 |
| RecR c | HR | DR0198 | 2 aa del | 104–164 | Figure S5 |
| DNA helicase (RecQ) | HR | DR1289 | 2 aa del | 453–510 | Figure S6 |
| Helicase RecD protein | HR | DR1902 | 2 aa del | 376–427 | Figure S7A |
| Helicase RecD protein | HR | DR1902 | 2 aa del | 426–493 | Figure S7B |
| RuvC protein | HR | DR0440 | 2 aa del | 82–147 | Figure S8 |
| DNA helicase RecG protein | HR | DR1916 | 1 aa ins | 14–73 | Figure S9 |
| DNA Repair protein RadA | HR | DR1105 | 1 aa del; c | 175–195 | [5] |
| 2 aa del | 225–257 | ||||
| DNA polymerase I (PolA) | MP | DR1707 | 2 aa ins | 191–257 | Figure S10 |
| DNA polymerase III, α subunit (DnaE) | MP | DR0507 | 65 aa ins | 315–491 | Figure S11 |
| DNA polymerase III, α subunit (DnaE) | MP | DR0507 | 2 aa ins | 75–131 | Figure S12 |
| DNA ligase (LigA) | MP | DR2069 | 3 aa ins | 101–169 | Figure S13 |
| Gyrase A (GyrA) | MP | DR1913 | 1 aa ins | 265–341 | Figure S14 |
| Gyrase B (GyrB) | MP | DR0906 | 1 aa ins | 27–99 | Figure S15 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, F.M.N.; Gupta, R.S. Novel Sequence Features of DNA Repair Genes/Proteins from Deinococcus Species Implicated in Protection from Oxidatively Generated Damage. Genes 2018, 9, 149. https://doi.org/10.3390/genes9030149
Hassan FMN, Gupta RS. Novel Sequence Features of DNA Repair Genes/Proteins from Deinococcus Species Implicated in Protection from Oxidatively Generated Damage. Genes. 2018; 9(3):149. https://doi.org/10.3390/genes9030149
Chicago/Turabian StyleHassan, F. M. Nazmul, and Radhey S. Gupta. 2018. "Novel Sequence Features of DNA Repair Genes/Proteins from Deinococcus Species Implicated in Protection from Oxidatively Generated Damage" Genes 9, no. 3: 149. https://doi.org/10.3390/genes9030149
APA StyleHassan, F. M. N., & Gupta, R. S. (2018). Novel Sequence Features of DNA Repair Genes/Proteins from Deinococcus Species Implicated in Protection from Oxidatively Generated Damage. Genes, 9(3), 149. https://doi.org/10.3390/genes9030149