Did Lizards Follow Unique Pathways in Sex Chromosome Evolution?


Abstract
:1. Introduction
2. Sex Chromosome Evolution in Amniotes—The Classical Concept
2.1. Hitchhiking, Meiotic Drive and Imprinting
2.2. Multiple Origin, Rapid Transitions and Turnovers
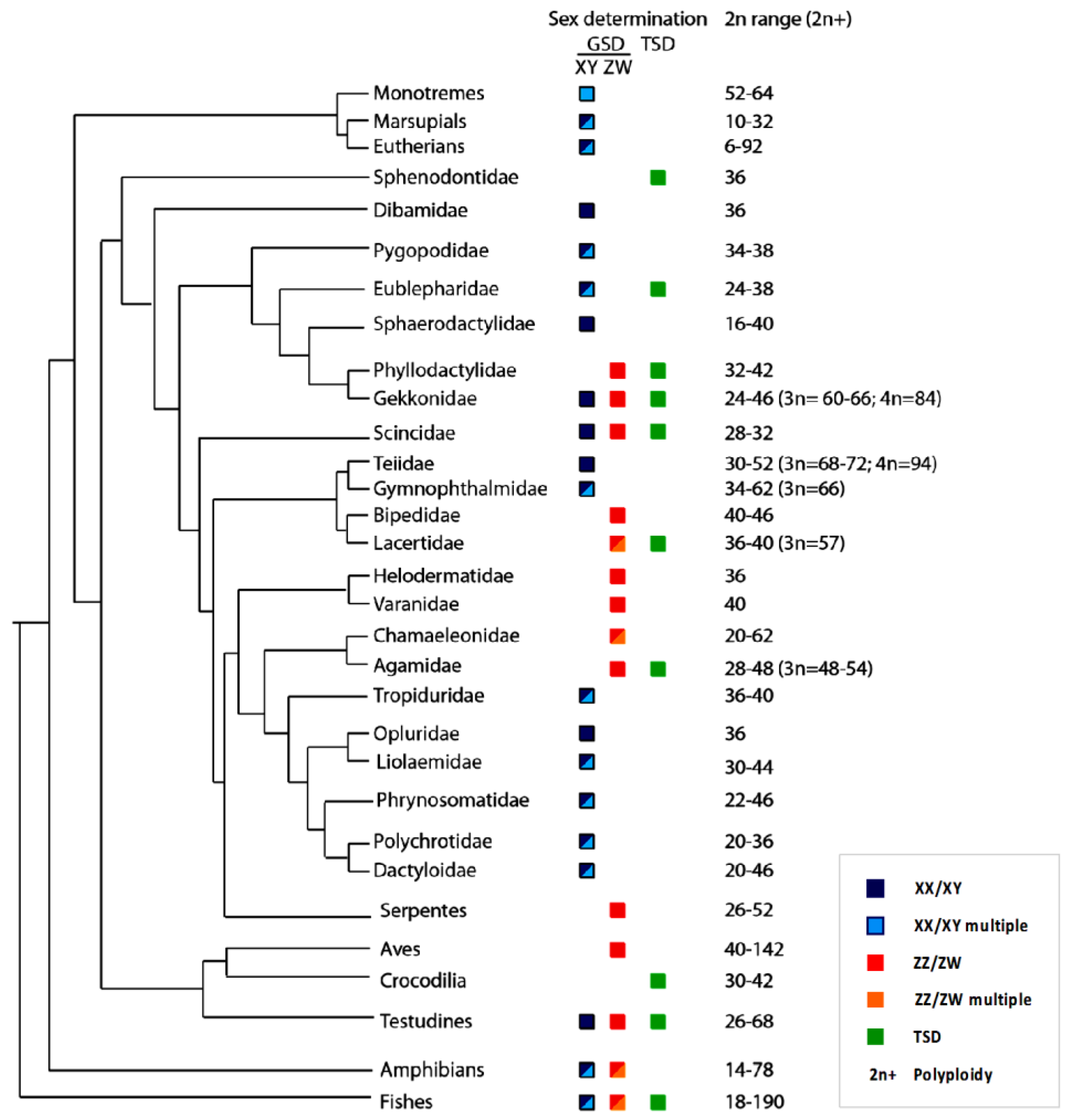
3. Overview of Current Understanding of Sex Determination and Sex Chromosomes in Lizards
3.1. Temperature-Dependent Sex Determination in Lizards
3.2. Karyotypes, Genotypic Sex Determination and Sex Chromosomes in Lizards
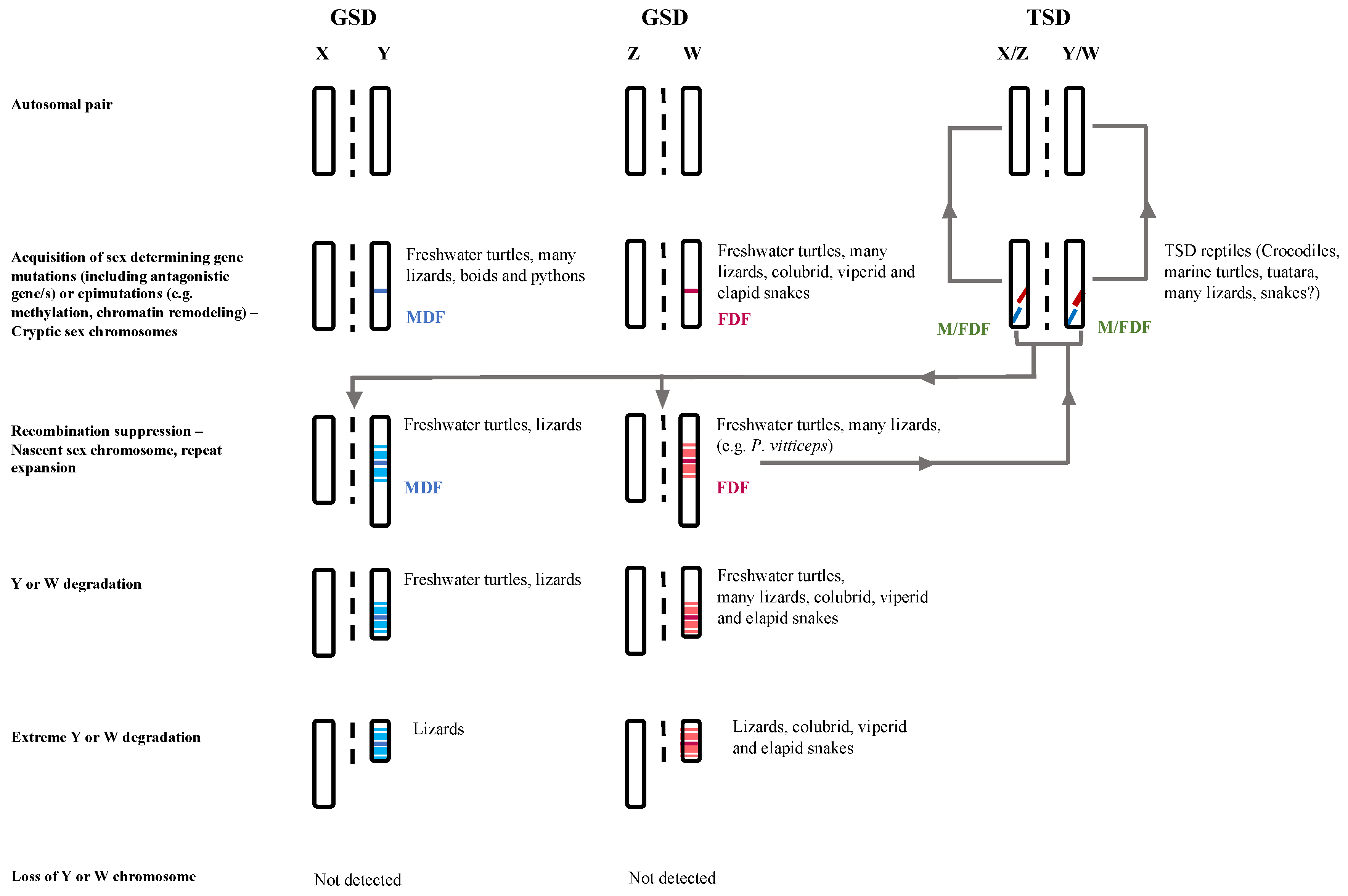
3.3. Sex Chromosome Differentiation in Lizards
4. Unique Pathway of Sex Chromosome Evolution in Lizards—A Different Pathway from the Classical Model (as That Proposed for Birds and Mammals)?
4.1. Temperature Dependent Sex Determination and Sex Chromosome Evolution
4.2. Multiple Sex Chromosomes in Lizards
4.3. Sex Chromosome Evolution in Lizards May Involve yet Undescribed Gene Regulatory Mechanisms
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Muller, H.J. A gene for the fourth chromosome of Drosophila. J. Exp. Zool. Part A Ecol. Genet. Physiol. 1914, 17, 325–336. [Google Scholar] [CrossRef]
- Ohno, S. Sex Chromosomes and Sex-Linked Genes; Springer: New York, NY, USA, 1967. [Google Scholar]
- Charlesworth, B. The evolution of sex chromosomes. Science 1991, 251, 1030–1033. [Google Scholar] [CrossRef] [PubMed]
- Bull, J. Sex determination in reptiles. Q. Rev. Biol. 1980, 55, 3–21. [Google Scholar] [CrossRef]
- Valenzuela, N. Evolution and maintenance of temperature-dependent sex determination. Temp.-Depend. Sex Determ. Vertebr. 2004, 14, 131–147. [Google Scholar]
- Ezaz, T.; Sarre, S.D.; O’Meally, D.; Graves, J.A.; Georges, A. Sex chromosome evolution in lizards: Independent origins and rapid transitions. Cytogenet. Genome Res. 2009, 127, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Deakin, J.E. Repetitive sequence and sex chromosome evolution in vertebrates. Adv. Evol. Biol. 2014, 2014, 104683. [Google Scholar] [CrossRef]
- Ezaz, T.; Srikulnath, K.; Graves, J.A.M. Origin of amniote sex chromosomes: An ancestral super-sex chromosome, or common requirements? J. Hered. 2017, 108, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Livernois, A.M.; Graves, J.A.; Waters, P.D. The origin and evolution of vertebrate sex chromosomes and dosage compensation. Heredity 2012, 108, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Vicoso, B.; Emerson, J.J.; Zektser, Y.; Mahajan, S.; Bachtrog, D. Comparative sex chromosome genomics in snakes: Differentiation, evolutionary strata, and lack of global dosage compensation. PLoS Biol. 2013, 11, e1001643. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.A.M. Evolution of vertebrate sex chromosomes and dosage compensation. Nat. Rev. Genet. 2016, 17, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Stiglec, R.; Veyrunes, F.; Marshall Graves, J.A. Relationships between vertebrate ZW and XY sex chromosome systems. Curr. Biol. 2006, 16, R736–R743. [Google Scholar] [CrossRef] [PubMed]
- Janzen, F.J.; Phillips, P.C. Exploring the evolution of environmental sex determination, especially in reptiles. J. Evol. Biol. 2006, 19, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
- Pokorna, M.; Kratochvíl, L. Phylogeny of sex-determining mechanisms in squamate reptiles: Are sex chromosomes an evolutionary trap? Zool. J. Linn. Soc. 2009, 156, 168–183. [Google Scholar] [CrossRef]
- Gamble, T.; Coryell, J.; Ezaz, T.; Lynch, J.; Scantlebury, D.P.; Zarkower, D. Restriction site-associated DNA sequencing (RAD-seq) reveals an extraordinary number of transitions among gecko sex-determining systems. Mol. Biol. Evol. 2015, 32, 1296–1309. [Google Scholar] [CrossRef] [PubMed]
- Quinn, A.E.; Radder, R.S.; Sarre, S.D.; Georges, A.; Ezaz, T.; Shine, R. Isolation and development of a molecular sex marker for Bassiana duperreyi, a lizard with XX/XY sex chromosomes and temperature-induced sex reversal. Mol. Genet. Genom. 2009, 281, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Holleley, C.E.; O’Meally, D.; Sarre, S.D.; Graves, J.A.M.; Ezaz, T.; Matsubara, K.; Azad, B.; Zhang, X.W.; Georges, A. Sex reversal triggers the rapid transition from genetic to temperature-dependent sex. Nature 2015, 523, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Shine, R.; Warner, D.A.; Radder, R. Windows of embryonic sexual lability in two lizard species with environmental sex determination. Ecology 2007, 88, 1781–1788. [Google Scholar] [CrossRef] [PubMed]
- Quinn, A.E.; Sarre, S.D.; Ezaz, T.; Marshall Graves, J.A.; Georges, A. Evolutionary transitions between mechanisms of sex determination in vertebrates. Biol. Lett. 2011, 7, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Raynaud, A.; Pieau, C. Embryonic development of the genital system. In Biology of the Reptilia; Gans, C., Billet, F., Eds.; Wiley: New York, NY, USA, 1985; Volume 15, pp. M9–M30. [Google Scholar]
- Deeming, D.; Ferguson, M.; Mittwoch, U.; Wolf, U.; Dorizzi, M.; Zaborski, P.; Sharma, H. Environmental regulation of sex determination in reptiles. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1988, 322, 19–39. [Google Scholar] [CrossRef] [PubMed]
- Ewert, M.A.; Nelson, C.E. Sex determination in turtles: Diverse patterns and some possible adaptive values. Copeia 1991, 1991, 50–69. [Google Scholar] [CrossRef]
- Moritz, C.; Adams, M.; Donnellan, S.; Baverstock, P. The origin and evolution of parthenogenesis in Heteronotia binoei (Gekkonidae): Genetic diversity among bisexual populations. Copeia 1990, 1990, 333–348. [Google Scholar] [CrossRef]
- Moritz, C.; Uzzell, T.; Spolsky, C.; Hotz, H.; Darevsky, I.; Kupriyanova, L.; Danielyan, F. The material ancestry and approximate age of parthenogenetic species of Caucasian rock lizards (Lacerta: Lacertidae). Genetica 1992, 87, 53–62. [Google Scholar] [CrossRef]
- Pyron, R.A.; Burbrink, F.T. Early origin of viviparity and multiple reversions to oviparity in squamate reptiles. Ecol. Lett. 2014, 17, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Sites, J.W.; Bickham, J.W.; Haiduk, M.W. Derived X chromosome in the turtle genus Staurotypus. Science 1979, 206, 1410–1412. [Google Scholar] [CrossRef] [PubMed]
- Uetz, P.; Hošek, J.; Hallermann, J. The Reptile Database. 2014. Available online: http://www.reptile-database.org/ (accessed on 30 March 2018).
- Wibbels, T.; Bull, J.J.; Crews, D. Chronology and morphology of temperature-dependent sex determination. J. Exp. Zool. 1991, 260, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.J. Phylogeny of the major tetrapod groups: Morphological data and divergence dates. J. Mol. Evol. 1990, 30, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.J.; Donoghue, P.C. Paleontological evidence to date the tree of life. Mol. Biol. Evol. 2006, 24, 26–53. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.A.M. Sex chromosome specialization and degeneration in mammals. Cell 2006, 124, 901–914. [Google Scholar] [CrossRef] [PubMed]
- Rens, W.; Grützner, F.; O’Brien, P.C.; Fairclough, H.; Graves, J.A.; Ferguson-Smith, M.A. Resolution and evolution of the duck-billed platypus karyotype with an X1Y1X2Y2X3Y3X4Y4X5Y5 male sex chromosome constitution. Proc. Nat. Acad. Sci. USA 2004, 101, 16257–16261. [Google Scholar] [CrossRef] [PubMed]
- Cortez, D.; Marin, R.; Toledo-Flores, D.; Froidevaux, L.; Liechti, A.; Waters, P.D.; Grützner, F.; Kaessmann, H. Origins and functional evolution of Y chromosomes across mammals. Nature 2014, 508, 488. [Google Scholar] [CrossRef] [PubMed]
- White, M. Animal Cytology and Evolution; Cambridge University: London, UK, 1973. [Google Scholar]
- Charlesworth, D.; Charlesworth, B. Sex differences in fitness and selection for centric fusions between sex-chromosomes and autosomes. Genet. Res. 1980, 35, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.J. Evolution of Sex Determining Mechanisms; The Benjamin/Cummings Publishing Company, Inc.: San Francisco, CA, USA, 1983. [Google Scholar]
- Rice, W.R. Genetic hitchhiking and the evolution of reduced genetic activity of the Y sex chromosome. Genetics 1987, 116, 161–167. [Google Scholar] [PubMed]
- Marshall Graves, J.A. Weird animal genomes and the evolution of vertebrate sex and sex chromosomes. Annu. Rev. Genet. 2008, 42, 565–586. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.E.; Dean, R.; Zimmer, F.; Mank, J.E. How to make a sex chromosome. Nat. Commun. 2016, 7, 12087. [Google Scholar] [CrossRef] [PubMed]
- Lahn, B.T.; Page, D.C. Four evolutionary strata on the human X chromosome. Science 1999, 286, 964–967. [Google Scholar] [CrossRef] [PubMed]
- Moghadam, H.K.; Pointer, M.A.; Wright, A.E.; Berlin, S.; Mank, J.E. W chromosome expression responds to female-specific selection. Proc. Nat. Acad. Sci. USA 2012, 109, 8207–8211. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.E.; Harrison, P.W.; Montgomery, S.H.; Pointer, M.A.; Mank, J.E. Independent stratum formation on the avian sex chromosomes reveals inter-chromosomal gene conversion and predominance of purifying selection on the W chromosome. Evolution 2014, 68, 3281–3295. [Google Scholar] [CrossRef] [PubMed]
- Bachtrog, D. Y-chromosome evolution: Emerging insights into processes of Y-chromosome degeneration. Nat. Rev. Genet. 2013, 14, 113. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the evolution of heteromorphic sex chromosomes. Heredity 2005, 95, 118. [Google Scholar] [CrossRef] [PubMed]
- Modi, W.S.; Crews, D. Sex chromosomes and sex determination in reptiles. Curr. Opin. Genet. Dev. 2005, 15, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex determination: Why so many ways of doing it? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, H.J. The relation of recombination to mutational advance. Mutat. Res. 1964, 1, 2–9. [Google Scholar] [CrossRef]
- Muller, H.J. Some genetic aspects of sex. Am. Nat. 1932, 66, 118–138. [Google Scholar] [CrossRef]
- Pennell, M.W.; Kirkpatrick, M.; Otto, S.P.; Vamosi, J.C.; Peichel, C.L.; Valenzuela, N.; Kitano, J. Y fuse? Sex chromosome fusions in fishes and reptiles. PLoS Genet. 2015, 11, e1005237. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, B.; Coyne, J.; Barton, N. The relative rates of evolution of sex chromosomes and autosomes. Am. Nat. 1987, 130, 113–146. [Google Scholar] [CrossRef]
- Bachtrog, D.; Kirkpatrick, M.; Mank, J.E.; McDaniel, S.F.; Pires, J.C.; Rice, W.; Valenzuela, N. Are all sex chromosomes created equal? Trends Genet. 2011, 27, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Ellegren, H. Sex-chromosome evolution: Recent progress and the influence of male and female heterogamety. Nat. Rev. Genet. 2011, 12, 157. [Google Scholar] [CrossRef] [PubMed]
- Gorelick, R. Evolution of dioecy and sex chromosomes via methylation driving Muller’s ratchet. Biol. J. Linn. Soc. 2003, 80, 353–368. [Google Scholar] [CrossRef]
- Charlesworth, B. Model for evolution of Y chromosomes and dosage compensation. Proc. Nat. Acad. Sci. USA 1978, 75, 5618–5622. [Google Scholar] [CrossRef] [PubMed]
- De Villena F, P.M.; Sapienza, C. Female meiosis drives karyotypic evolution in mammals. Genetics 2001, 159, 1179–1189. [Google Scholar]
- Jaenike, J. Sex chromosome meiotic drive. Annu. Rev. Ecol. Syst. 2001, 32, 25–49. [Google Scholar] [CrossRef]
- Helleu, Q.; Gérard, P.R.; Montchamp-Moreau, C. Sex chromosome drive. Cold Spring Harb. Perspect. Biol. 2015, 7, a017616. [Google Scholar] [CrossRef] [PubMed]
- Peichel, C.L. Convergence and divergence in sex-chromosome evolution. Nat. Genet. 2017, 49, 321. [Google Scholar] [CrossRef] [PubMed]
- Lemos, B.; Branco, A.T.; Jiang, P.-P.; Hartl, D.L.; Meiklejohn, C.D. Genome-wide gene expression effects of sex chromosome imprinting in Drosophila. G3 (Bethesda). 2014, 4, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sha, K. A mechanistic view of genomic imprinting. Annu. Rev. Genom. Hum. Genet. 2008, 9, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Ferguson-Smith, A.C. Genomic imprinting: The emergence of an epigenetic paradigm. Nat. Rev. Genet. 2011, 12, 565. [Google Scholar] [CrossRef] [PubMed]
- Sarre, S.D.; Ezaz, T.; Georges, A. Transitions between sex-determining systems in reptiles and amphibians. Annu. Rev. Genom. Hum. Genet. 2011, 12, 391–406. [Google Scholar] [CrossRef] [PubMed]
- Koopman, P.; Munsterberg, A.; Capel, B.; Vivian, N.; Lovellbadge, R. Expression of a candidate sex-determining gene during mouse testis differentiation. Nature 1990, 348, 450–452. [Google Scholar] [CrossRef] [PubMed]
- Vaiman, D.; Pailhoux, E. Mammalian sex reversal and intersexuality: Deciphering the sex-determination cascade. Trends Genet. 2000, 16, 488–494. [Google Scholar] [CrossRef]
- Sinclair, A.H.; Berta, P.; Palmer, M.S.; Hawkins, J.R.; Griffiths, B.L.; Smith, M.J. A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature 1990, 346, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.A.M. Mammals that break the rules—Genetics of marsupials and monotremes. Annu. Rev. Genet. 1996, 30, 233–260. [Google Scholar] [CrossRef] [PubMed]
- Just, W.; Rau, W.; Vogul, W.; Akhverdian, M.; Fredga, K.; Graves, J.A.M. Absence of Sry in species of the vole Ellobius. Nat. Genet. 1995, 11, 117–118. [Google Scholar] [CrossRef] [PubMed]
- Soullier, S.; Hanni, C.; Catzeflis, F.; Berta, P.; Laudet, V. Male sex determination in the spiny rat Tokudaia osimensis (Rodentia: Muridae) is not Sry dependent. Mamm. Genome 1998, 9, 590–592. [Google Scholar] [CrossRef] [PubMed]
- Sutou, S.; Mitsui, Y.; Tsuchiya, K. Sex determination without the Y Chromosome in two Japanese rodents Tokudaia osimensis osimensis and Tokudaia osimensis spp. Mamm. Genome 2001, 12, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Katz, M.; Sinclair, A.H. DMRT1 is upregulated in the gonads during female-to-male sex reversal in ZW chicken embryos. Biol. Reprod. 2003, 68, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Koopman, P.; Gubbay, J.; Vivian, N.; Goodfellow, P.; Lovell-Badge, R. Male development of chromosomally female mice transgenic for Sry. Nature 1991, 351, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Arlt, D.; Bensch, S.; Hansson, B.; Hasselquist, D.; Westerdahl, H. Observation of a ZZW female in a natural population: Implications for avian sex determination. Proc. R. Soc. Lond. B Biol. Sci. 2004, 271, S249–S251. [Google Scholar] [CrossRef] [PubMed]
- Solari, A.J. Sex Chromosomes and Sex Determination in Vertebrates; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Olmo, E. Rate of chromosome changes and speciation in reptiles. Genetica 2005, 125, 185–203. [Google Scholar] [CrossRef] [PubMed]
- Cree, A.; Thompson, M.B.; Daugherty, C.H. Tuatara sex determination. Nature 1995, 375, 543. [Google Scholar] [CrossRef]
- Ewert, M.A.; Jackson, D.R.; Nelson, C.E. Patterns of temperature-dependent sex determination in turtles. J. Exp. Zool. 1994, 270, 3–15. [Google Scholar] [CrossRef]
- Ferguson, M.W.J.; Joanen, T. Temperature of egg incubation determines sex in Alligator mississippiensis. Nature 1982, 296, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Harlow, P.S. Temperature-dependent sex determination in lizards. In Temperature-Dependent Sex Determination in Vertebrates; Valenzuela, N., Lance, V., Eds.; Smithsonian Institute Press: Washington, DC, USA, 2004; pp. 42–52. [Google Scholar]
- Radder, R.S.; Quinn, A.E.; Georges, A.; Sarre, S.D.; Shine, R. Genetic evidence for co-occurrence of chromosomal and thermal sex-determining systems in a lizard. Biol. Lett. 2008, 4, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Quinn, A.E.; Georges, A.; Sarre, S.D.; Guarino, F.; Ezaz, T.; Graves, J.A.M. Temperature sex reversal implies sex gene dosage in a reptile. Science 2007, 316, 411. [Google Scholar] [CrossRef] [PubMed]
- Holleley, C.E.; Sarre, S.D.; O’Meally, D.; Georges, A. Sex reversal in reptiles: Reproductive oddity or powerful driver of evolutionary change? Sex. Dev. 2016, 10, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Pen, I.; Uller, T.; Feldmeyer, B.; Harts, A.; While, G.M.; Wapstra, E. Climate-driven population divergence in sex-determining systems. Nature 2010, 468, 436–438. [Google Scholar] [CrossRef] [PubMed]
- Georges, A.; Ezaz, T.; Quinn, A.E.; Sarre, S.D. Are reptiles predisposed to temperature-dependent sex determination? Sex. Dev. 2010, 4, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Quinn, A.E.; Sarre, S.D.; O’Meally, D.; Georges, A.; Graves, J.A. Molecular marker suggests rapid changes of sex-determining mechanisms in Australian dragon lizards. Chromosome Res. 2009, 17, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Gamble, T. A review of sex determining mechanisms in Geckos (Gekkota: Squamata). Sex. Dev. 2010, 4, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Organ, C.L.; Janes, D.E. Evolution of sex chromosomes in Sauropsida. Integr. Comp. Biol. 2008, 48, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Osgood, D. Sex ratio and incubation temperature in a watersnake. Q. Rev. Biol. 1980, 55, 21. [Google Scholar]
- Dunlap, K.D.; Lang, J.W. Offspring sex ratio varies with maternal size in the common garter snake, Thamnophis sirtalis. Copeia 1990, 1990, 568–570. [Google Scholar] [CrossRef]
- Hammack, S.H. Reproduction of the Colombian milk snake Laniprupellis Iriangulitm andesiana at the Dallas Zoo. Int. Zoo Yearb. 1989, 28, 172–177. [Google Scholar] [CrossRef]
- Burger, J.; Zappalorti, R. Effects of incubation temperature on sex ratios in pine snakes: Differential vulnerability of males and females. Am. Nat. 1988, 132, 492–505. [Google Scholar] [CrossRef]
- Du, W.; Ji, X. The effects of incubation temperature on hatching success, embryonic use of energy and hatchling morphology in the stripe-tailed ratsnake Elaphe taeniura. Asiat. Herpetol. Res. 2008, 11, 24–30. [Google Scholar]
- Robert, K.A.; Thompson, M.B. Sex determination: Viviparous lizard selects sex of embryos. Nature 2001, 412, 698–699. [Google Scholar] [CrossRef] [PubMed]
- Wapstra, E.; Olsson, M.; Shine, R.; Edwards, A.; Swain, R.; Joss, J.M. Maternal basking behaviour determines offspring sex in a viviparous reptile. Proc. R. Soc. Lond. B Biol. Sci. 2004, 271, S230–S232. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Lin, L.-H.; Luo, L.-G.; Lu, H.-L.; Gao, J.-F.; Han, J. Gestation temperature affects sexual phenotype, morphology, locomotor performance, and growth of neonatal brown forest skinks, Sphenomorphus indicus. Biol. J. Linn. Soc. 2006, 88, 453–463. [Google Scholar] [CrossRef]
- Le Galliard, J.F.; Fitze, P.; Cote, J.; Massot, M.; Clobert, J. Female common lizards (Lacerta vivipara) do not adjust their sex-biased investment in relation to the adult sex ratio. J. Evol. Biol. 2005, 18, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
- Georges, A. Female turtles from hot nests: Is it duration of incubation or proportion of development at high temperatures that matters? Oecologia 1989, 81, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Crews, D.; Bergeron, J.M.; Bull, J.J.; Flores, D.; Tousignant, A.; Skipper, J.K.; Wibbels, T. Temperature-dependent sex determination in reptiles: Proximate mechanisms, ultimate outcomes, and practical applications. Genesis 1994, 15, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Pieau, C. Temperature variation and sex determination in reptiles. BioEssays 1996, 18, 19–26. [Google Scholar] [CrossRef]
- Ciofi, C.; Swingland, I.R. Environmental sex determination in reptiles. Appl. Anim. Behav. Sci. 1997, 51, 251–265. [Google Scholar] [CrossRef]
- Shine, R. Why is sex determined by nest temperature in many reptiles? Trends Ecol. Evol. 1999, 14, 186–189. [Google Scholar] [CrossRef]
- Harlow, P.S. The Ecology of Sex-Determining Mechanisms in Australian Agamid Lizards; Macquarie University: Sydney, Australia, 2001. [Google Scholar]
- Ewert, M.A.; Etchberger, C.R.; Nelson, C.E. Turtle sex-determining modes and TSD patterns, and some TSD pattern correlates. In Temperature-Dependent Sex Determination in Vertebrates; Smithsonian Books: Washington, DC, USA, 2004; pp. 21–32. [Google Scholar]
- O’Meally, D.; Miller, H.; Patel, H.; Graves, J.M.; Ezaz, T. The first cytogenetic map of the tuatara, Sphenodon punctatus. Cytogenet. Genome Res. 2009, 127, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Pokorná, M.; Giovannotti, M.; Kratochvíl, L.; Kasai, F.; Trifonov, V.A.; O’Brien, P.C.; Caputo, V.; Olmo, E.; Ferguson-Smith, M.A.; Rens, W. Strong conservation of the bird Z chromosome in reptilian genomes is revealed by comparative painting despite 275 million years divergence. Chromosoma 2011, 120, 455. [Google Scholar] [CrossRef] [PubMed]
- Janes, D.E.; Elsey, R.M.; Langan, E.M.; Valenzuela, N.; Edwards, S.V. Sex-biased expression of sex-differentiating genes FOXL2 and FGF9 in American alligators, Alligator mississippiensis. Sex. Dev. 2013, 7, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Pezaro, N.; Doody, J.S.; Thompson, M.B. The ecology and evolution of temperature-dependent reaction norms for sex determination in reptiles: A mechanistic conceptual model. Biol. Rev. 2017, 92, 1348–1364. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.G.; Parham, J.F.; Sellas, A.B.; Faircloth, B.C.; Glenn, T.C.; Papenfuss, T.J.; Henderson, J.B.; Hansen, M.H.; Simison, W.B. A phylogenomic analysis of turtles. Mol. Phylogenet. Evol. 2015, 83, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Olmo, E.; Signorino, G. Chromorep: A Reptile Chromosomes Database. 2005. Available online: http://chromorep.univpm.it/ (accessed on 30 March 2018).
- Pyron, R.A.; Burbrink, F.T.; John, J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 2013, 13, 93. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wiens, J.J. Combining phylogenomic and supermatrix approaches, and a time-calibrated phylogeny for squamate reptiles (lizards and snakes) based on 52 genes and 4162 species. Mol. Phylogenet. Evol. 2016, 94, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Capel, B. Vertebrate sex determination: Evolutionary plasticity of a fundamental switch. Nat. Rev. Genet. 2017, 18, 675. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Altmanova, M.; Johnson Pokorna, M.; Velensky, P.; Sanchez Baca, A.; Kratochvil, L. Evolution of karyotypes in chameleons. Genes 2017, 8, 382. [Google Scholar] [CrossRef] [PubMed]
- Patawang, I.; Chuaynkern, Y.; Supanuam, P.; Maneechot, N.; Pinthong, K.; Tanomtong, A. Cytogenetics of the skinks (Reptilia, Scincidae) from Thailand; IV: Newly investigated karyotypic features of Lygosoma quadrupes and Scincella melanosticta. Caryologia 2017, 71, 29–34. [Google Scholar] [CrossRef]
- Wrigley, J.; Graves, J.M. Sex chromosome homology and incomplete, tissue-specific X-inactivation suggest that monotremes represent an intermediate stage of mammalian sex chromosome evolution. J. Hered. 1988, 79, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Rens, W.; O’Brien, P.; Fairclough, H.; Harman, L.; Graves, J.; Ferguson-Smith, M. Reversal and convergence in marsupial chromosome evolution. Cytogenet. Genome Res. 2003, 102, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Altmanová, M.; Rovatsos, M.; Kratochvíl, L.; Johnson Pokorná, M. Minute Y chromosomes and karyotype evolution in Madagascan iguanas (Squamata: Iguania: Opluridae). Biol. J. Linn. Soc. 2016, 118, 618–633. [Google Scholar] [CrossRef]
- Pokorná, M.; Rábová, M.; Ráb, P.; Ferguson-Smith, M.A.; Rens, W.; Kratochvíl, L. Differentiation of sex chromosomes and karyotypic evolution in the eye-lid geckos (Squamata: Gekkota: Eublepharidae), a group with different modes of sex determination. Chromosome Res. 2010, 18, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Spangenberg, V.; Arakelyan, M.; Galoyan, E.; Matveevsky, S.; Petrosyan, R.; Bogdanov, Y.; Danielyan, F.; Kolomiets, O. Reticulate evolution of the rock lizards: Meiotic chromosome dynamics and spermatogenesis in diploid and triploid males of the genus Darevskia. Genes 2017, 8, 149. [Google Scholar] [CrossRef] [PubMed]
- Gregory, T.R. Animal Genome Size Database. 2018. Available online: http://www.genomesize.com/ (accessed on 30 March 2018).
- Organ, C.L.; Moreno, R.G.; Edwards, S.V. Three tiers of genome evolution in reptiles. Integr. Comp. Biol. 2008, 48, 494–504. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Quinn, A.E.; Miura, I.; Sarre, S.D.; Georges, A.; Marshall Graves, J.A. The dragon lizard Pogona vitticeps has ZZ/ZW micro-sex chromosomes. Chromosome Res. 2005, 13, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Olmo, E. Reptilia; Gebruder Borntraeger: Stuttgart, Germany, 1986; Volume 4. [Google Scholar]
- Janzen, F.J.; Paukstis, G.L. Environmental sex determination in reptiles: Ecology, evolution, and experimental design. Q. Rev. Biol. 1991, 66, 149–179. [Google Scholar] [CrossRef] [PubMed]
- Shine, R.; Elphick, M.; Donnellan, S. Co-occurrence of multiple, supposedly incompatible modes of sex determination in a lizard population. Ecol. Lett. 2002, 5, 486–489. [Google Scholar] [CrossRef]
- Fridolfsson, A.-K.; Cheng, H.; Copeland, N.G.; Jenkins, N.A.; Liu, H.-C.; Raudsepp, T.; Woodage, T.; Chowdhary, B.; Halverson, J.; Ellegren, H. Evolution of the avian sex chromosomes from an ancestral pair of autosomes. Proc. Nat. Acad. Sci. USA 1998, 95, 8147–8152. [Google Scholar] [CrossRef] [PubMed]
- Takehana, Y.; Hamaguchi, S.; Sakaizumi, M. Different origins of ZZ/ZW sex chromosomes in closely related medaka fishes, Oryzias javanicus and O. hubbsi. Chromosome Res. 2008, 16, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Schubert, I.; Lysak, M.A. Interpretation of karyotype evolution should consider chromosome structural constraints. Trends Genet. 2011, 27, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, R.F.; Kirkpatrick, M. Local adaptation and the evolution of chromosome fusions. Evolution 2014, 68, 2747–2756. [Google Scholar] [CrossRef] [PubMed]
- Lande, R. Effective deme sizes during long-term evolution estimated from rates of chromosomal rearrangement. Evolution 1979, 33, 234–251. [Google Scholar] [CrossRef] [PubMed]
- Kawagoshi, T.; Uno, Y.; Matsubara, K.; Matsuda, Y.; Nishida, C. The ZW micro-sex chromosomes of the Chinese soft-shelled turtle (Pelodiscus sinensis, Trionychidae, Testudines) have the same origin as chicken chromosome 15. Cytogenet. Genome Res. 2009, 125, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Badenhorst, D.; Stanyon, R.; Engstrom, T.; Valenzuela, N. A ZZ/ZW microchromosome system in the spiny softshell turtle, Apalone spinifera, reveals an intriguing sex chromosome conservation in Trionychidae. Chromosome Res. 2013, 21, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Gamble, T.; Geneva, A.J.; Glor, R.E.; Zarkower, D. Anolis sex chromosomes are derived from a single ancestral pair. Evolution 2014, 68, 1027–1041. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Pokorná, M.; Altmanová, M.; Kratochvíl, L. Cretaceous park of sex determination: Sex chromosomes are conserved across iguanas. Biol. Lett. 2014, 10, 20131093. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Vukić, J.; Altmanová, M.; Johnson Pokorná, M.; Moravec, J.; Kratochvíl, L. Conservation of sex chromosomes in lacertid lizards. Mol. Ecol. 2016, 25, 3120–3126. [Google Scholar] [CrossRef] [PubMed]
- Srikulnath, K.; Matsubara, K.; Uno, Y.; Nishida, C.; Olsson, M.; Matsuda, Y. Identification of the linkage group of the Z sex chromosomes of the sand lizard (Lacerta agilis, Lacertidae) and elucidation of karyotype evolution in lacertid lizards. Chromosoma 2014, 123, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Srikulnath, K.; Nishida, C.; Matsubara, K.; Uno, Y.; Thongpan, A.; Suputtitada, S.; Apisitwanich, S.; Matsuda, Y. Karyotypic evolution in squamate reptiles: Comparative gene mapping revealed highly conserved linkage homology between the butterfly lizard (Leiolepis reevesii rubritaeniata, Agamidae, Lacertilia) and the Japanese four-striped rat snake (Elaphe quadrivirgata, Colubridae, Serpentes). Chromosome Res. 2009, 17, 975–986. [Google Scholar] [CrossRef] [PubMed]
- Banno, K.; Kisu, I.; Yanokura, M.; Tsuji, K.; Masuda, K.; Ueki, A.; Kobayashi, Y.; Yamagami, W.; Nomura, H.; Tominaga, E. Epimutation and cancer: A new carcinogenic mechanism of Lynch syndrome. Int. J. Oncol. 2012, 41, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Mank, J.E.; Uller, T. The evolution of sex determination in animals. Adv. Evol. Dev. Biol. 2014, 2, 15–36. [Google Scholar]
- Pokorná, M.; Altmanová, M.; Kratochvíl, L. Multiple sex chromosomes in the light of female meiotic drive in amniote vertebrates. Chromosome Res. 2014, 22, 35–44. [Google Scholar] [CrossRef] [PubMed]
- O’Meally, D.; Ezaz, T.; Georges, A.; Sarre, S.D.; Graves, J.A.M. Are some chromosomes particularly good at sex? Insights from amniotes. Chromosome Res. 2012, 20, 7–19. [Google Scholar] [CrossRef] [PubMed]
- O’Meally, D.; Patel, H.R.; Stiglec, R.; Sarre, S.D.; Georges, A.; Graves, J.A.M.; Ezaz, T. Non-homologous sex chromosomes of birds and snakes share repetitive sequences. Chromosome Res. 2010, 18, 787–800. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, A.L.; Metzger, K.J.; Miller, A.; Rhen, T. A novel candidate gene for temperature-dependent sex determination in the common snapping turtle. Genetics 2016, 203, 557–571. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Taxon | Chromosome | Genome Size (Gb) [119] | |||
|---|---|---|---|---|---|
| 2n Range | Macro Range | Micro Range | Low | High | |
| Tuatara | 36 | 28 | 8 | 4.9 | |
| Lizards | 20–62 | 10–38 | 0–28 | 1.03 | 3.8 |
| Snakes | 26–50 | 10–38 | 0–36 | 1.3 | 3.7 |
| Crocodilians | 30–42 | 30–42 | 0 | 1.3 | 3.9 |
| Freshwater Turtles | 26–68 | 10–36 | 0–56 | 1.4 | 5.3 |
| Marine turtles | 56 | 24–32 | 24–32 | 2.6 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alam, S.M.I.; Sarre, S.D.; Gleeson, D.; Georges, A.; Ezaz, T. Did Lizards Follow Unique Pathways in Sex Chromosome Evolution? Genes 2018, 9, 239. https://doi.org/10.3390/genes9050239
Alam SMI, Sarre SD, Gleeson D, Georges A, Ezaz T. Did Lizards Follow Unique Pathways in Sex Chromosome Evolution? Genes. 2018; 9(5):239. https://doi.org/10.3390/genes9050239
Chicago/Turabian StyleAlam, Shayer Mahmood Ibney, Stephen D. Sarre, Dianne Gleeson, Arthur Georges, and Tariq Ezaz. 2018. "Did Lizards Follow Unique Pathways in Sex Chromosome Evolution?" Genes 9, no. 5: 239. https://doi.org/10.3390/genes9050239
APA StyleAlam, S. M. I., Sarre, S. D., Gleeson, D., Georges, A., & Ezaz, T. (2018). Did Lizards Follow Unique Pathways in Sex Chromosome Evolution? Genes, 9(5), 239. https://doi.org/10.3390/genes9050239