The Transcription Factor ZafA Regulates the Homeostatic and Adaptive Response to Zinc Starvation in Aspergillus fumigatus

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Media
2.2. Culture of Aspergillus fumigatus for RNA Isolation
2.3. Microarray Experiments
2.4. RT-qPCR
2.5. Expression and Purification of the Recombinant ZafA Protein
2.6. Construction of Plasmids to Obtain Probes for Electrophoretic Mobility Shift Assays and DNase I Footprinting Experiments
2.7. Preparation of Probes for Electrophoretic Mobility Shift Assays
2.8. Preparation of Probes for DNase I Footprinting Assays
2.9. Electrophoretic Mobility Shift Assays and DNase I Footprinting Assays Reactions and Analyses
2.10. Construction of Plasmids Used for Aspergillus fumigatus Transformation
2.11. Generation of Protoplasts and Transformation of Aspergillus fumigatus
2.12. Detection of SOD Activity on PAGE Gels
3. Results
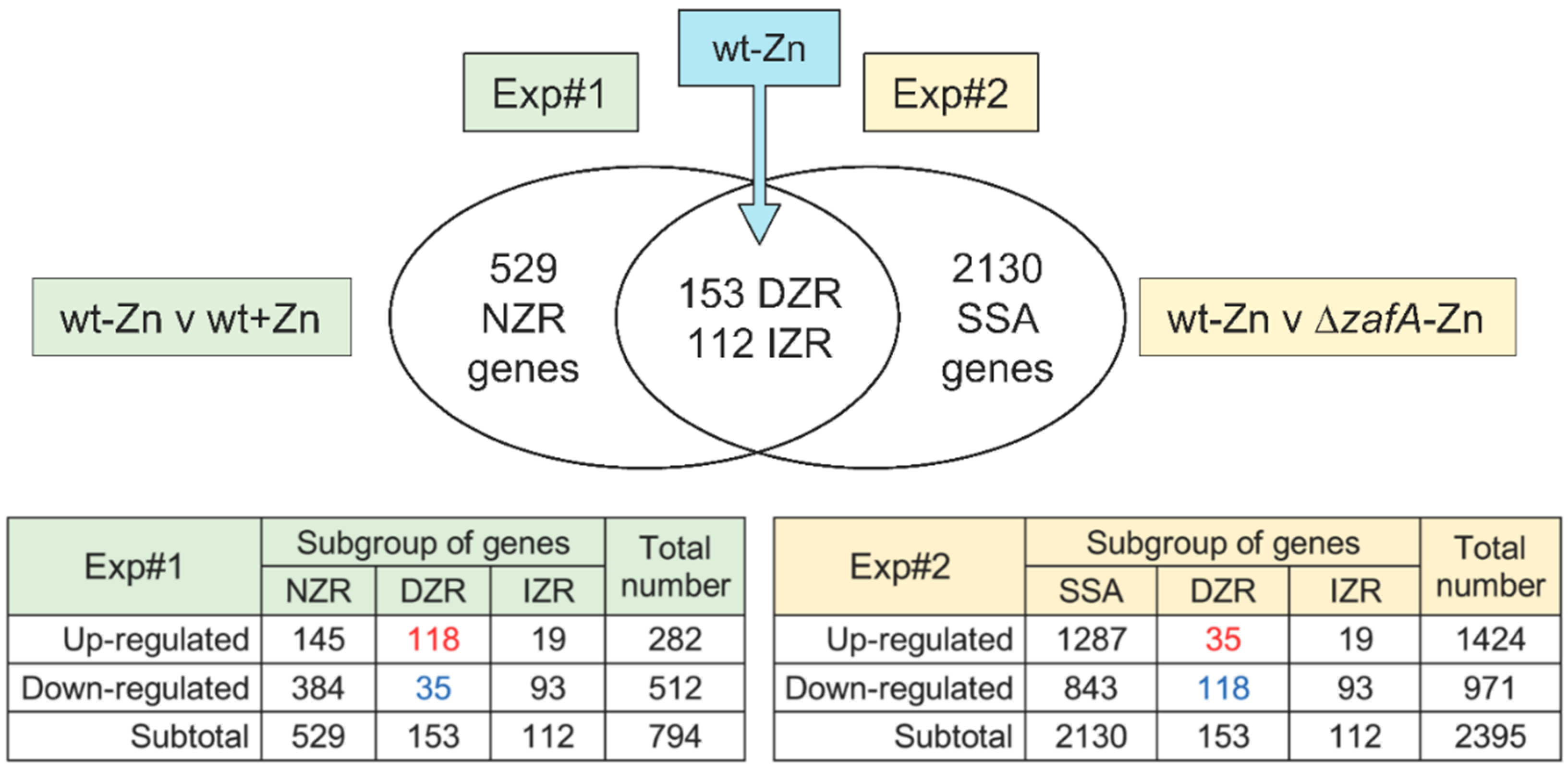
3.1. Under Zinc-Limiting Conditions ZafA Influences the Expression of Genes Related to Many Different Metabolic and Biosynthetic Processes in Addition to Those Required to Maintain Zinc Homeostasis
3.2. Zinc Starvation Influences the Expression of Genes Related to Many Different Biological Processes
3.3. The ZafA Transcription Factor Binds to Promoter Regions of Homeostatic Genes Whose Transcription Is Induced under Zinc-Limiting Conditions
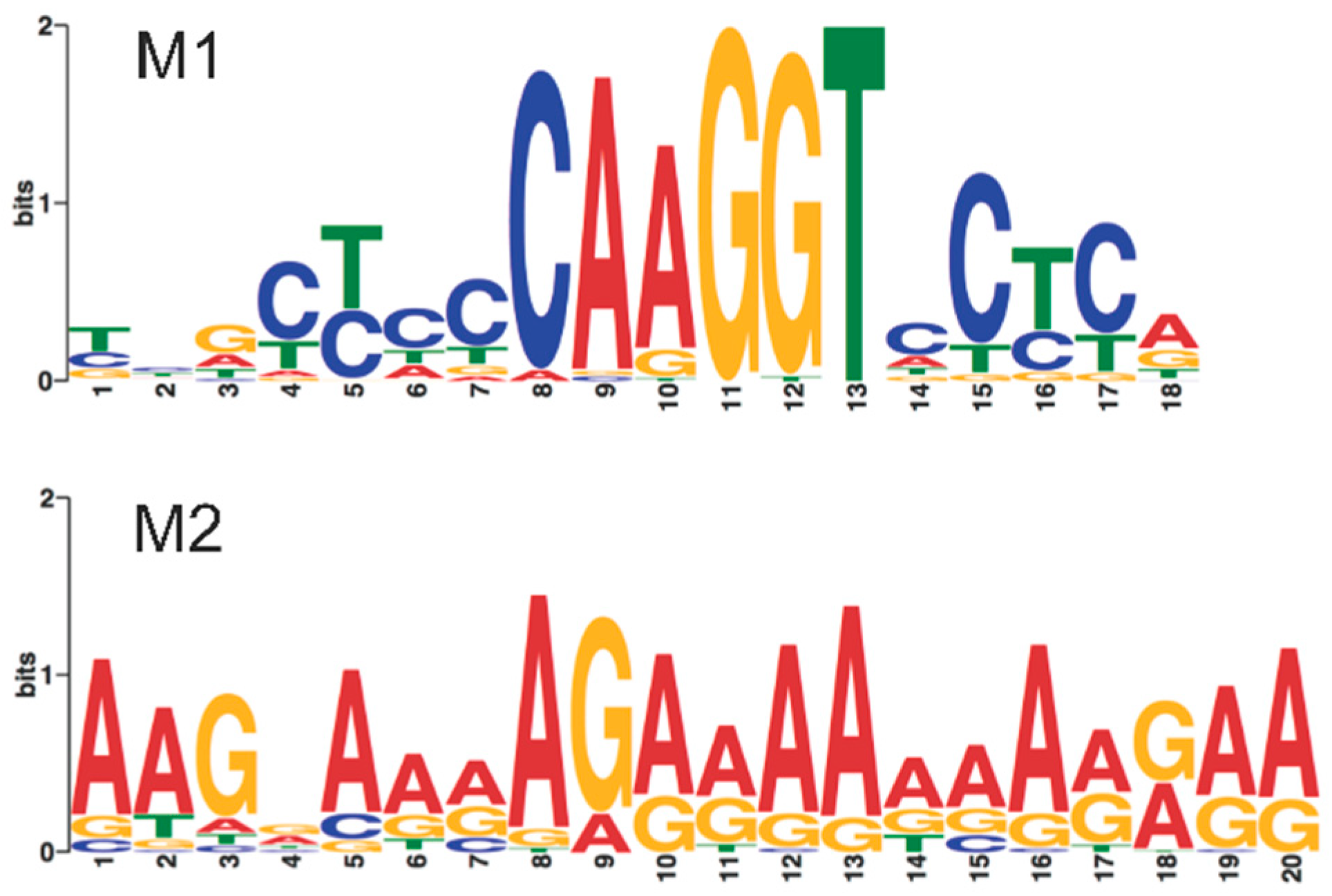
3.4. The ZafA Transcription Factor Binds Specifically to a 15-bp Zinc-Response Consensus Sequence
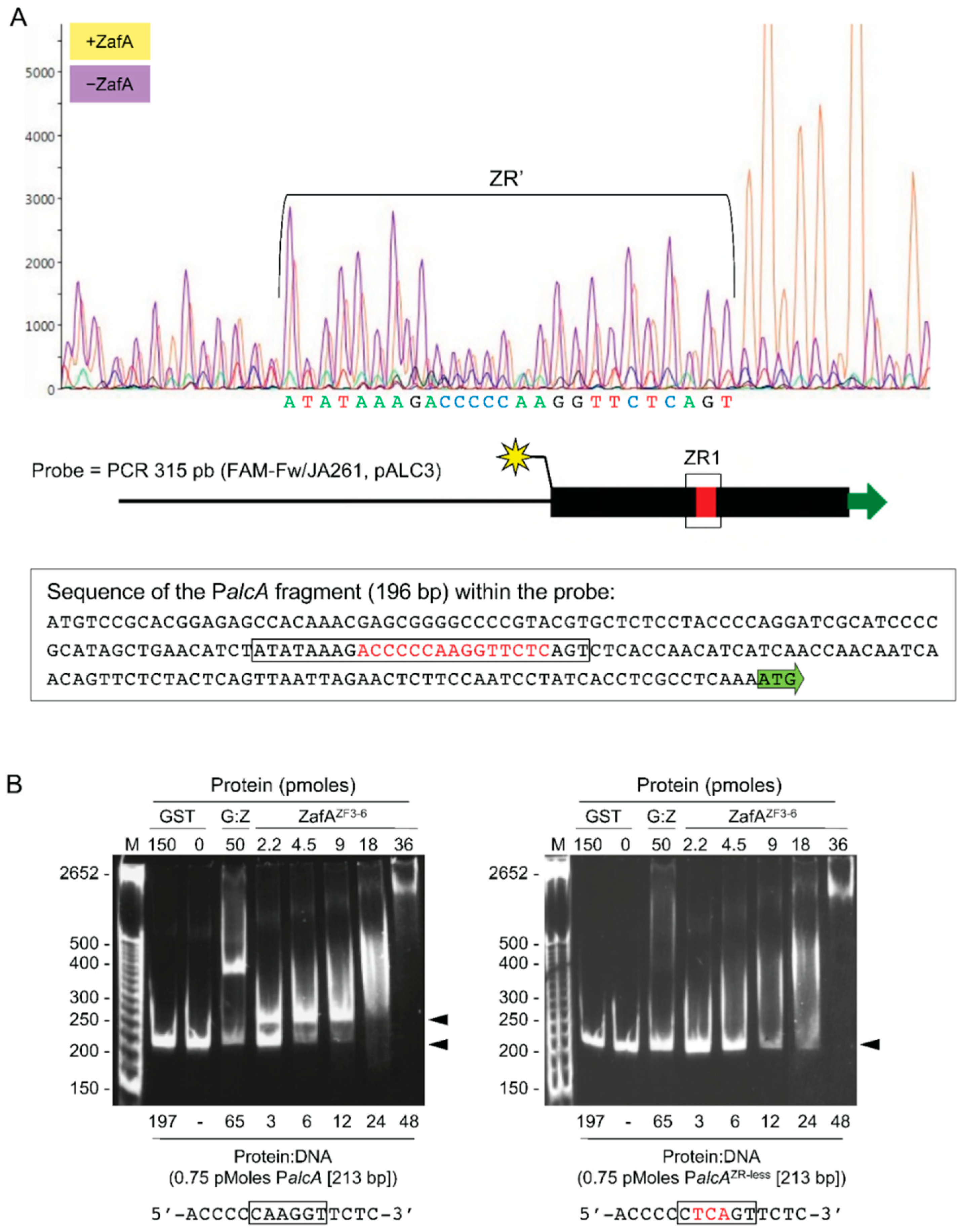
3.5. The Highly Conserved 5′-CARGGT-3′ Core of the ZR Motif Is Essential for ZafA Binding
3.6. ZafA Induces Gene Expression in vivo through Binding to the ZR Motifs
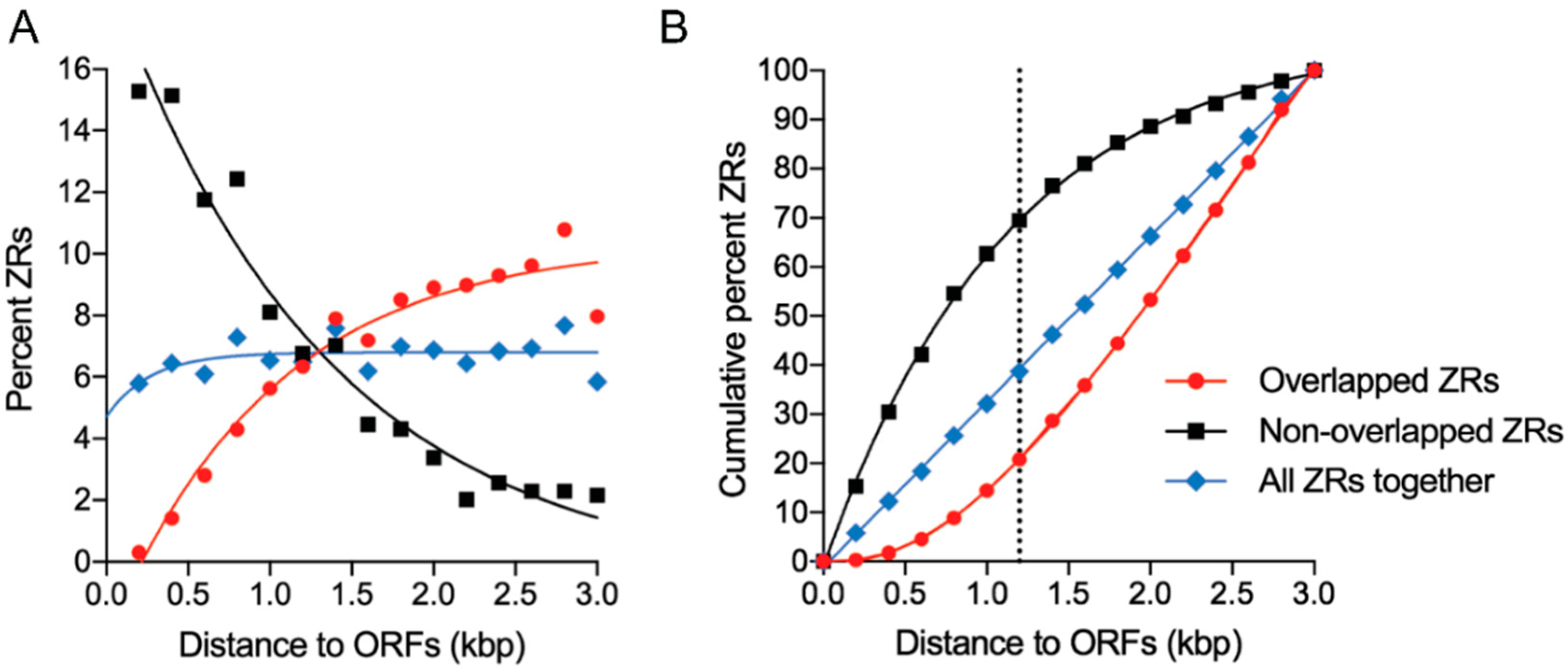
3.7. The ZR Motifs Appear to Function as Transcriptional Regulatory Sequences When Located within 1.2 Kb Upstream from the Predicted Translation Start Codon of the ORFs Whose Expression Is Influenced by Zinc
3.8. Putative Role of ZafA as a Repressor under Zinc-Limiting Conditions and Identification of the Most Likely Direct Target Genes of ZafA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Fundings
Acknowledgments
Conflicts of Interest
References
- Andreini, C.; Bertini, I.; Cavallaro, G.; Holliday, G.L.; Thornton, J.M. Metal ions in biological catalysis: From enzyme databases to general principles. J. Biol. Inorg. Chem. 2008, 13, 1205–1218. [Google Scholar] [CrossRef] [PubMed]
- Auld, D.S. The ins and outs of biological zinc sites. Biometals 2009, 22, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Zinc and sulfur: a critical biological partnership. Biochemistry 2004, 43, 3301–3309. [Google Scholar] [CrossRef] [PubMed]
- Eide, D.J. Zinc transporters and the cellular trafficking of zinc. Biochim. Biophys. Acta 2006, 1763, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D.; Findley, S.D. Cloning and functional characterization of a mammalian zinc transporter that confers resistance to zinc. EMBO J. 1995, 14, 639–649. [Google Scholar] [PubMed]
- Maret, W. Zinc biochemistry: From a single zinc enzyme to a key element of life. Adv. Nutr. 2013, 4, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Simons, T.J. Intracellular free zinc and zinc buffering in human red blood cells. J. Membr. Biol. 1991, 123, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Bessman, J.D.; Johnson, R.K. Erythrocyte volume distribution in normal and abnormal subjects. Blood 1975, 46, 369–379. [Google Scholar] [PubMed]
- Karthaus, M.; Buchheidt, D. Invasive aspergillosis: New insights into disease, diagnostic and treatment. Curr. Pharm. Des. 2013, 19, 3569–3594. [Google Scholar] [CrossRef] [PubMed]
- Amich, J.; Calera, J.A. Zinc acquisition: A key aspect in Aspergillus fumigatus virulence. Mycopathologia 2014, 178, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Amich, J.; Vicentefranqueira, R.; Leal, F.; Calera, J.A. Aspergillus fumigatus survival in alkaline and extreme zinc-limiting environments relies on the induction of a zinc homeostasis system encoded by the zrfC and aspf2 genes. Eukaryot. Cell 2010, 9, 424–437. [Google Scholar] [CrossRef] [PubMed]
- Vicentefranqueira, R.; Moreno, M.A.; Leal, F.; Calera, J.A. The zrfA and zrfB genes of Aspergillus fumigatus encode the zinc transporter proteins of a zinc uptake system induced in an acid, zinc-depleted environment. Eukaryot. Cell 2005, 4, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Amich, J.; Vicentefranqueira, R.; Mellado, E.; Ruiz-Carmuega, A.; Leal, F.; Calera, J.A. The ZrfC alkaline zinc transporter is required for Aspergillus fumigatus virulence and its growth in the presence of the Zn/Mn-chelating protein calprotectin. Cell Microbiol. 2014, 16, 548–564. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.A.; Ibrahim-Granet, O.; Vicentefranqueira, R.; Amich, J.; Ave, P.; Leal, F.; Latge, J.P.; Calera, J.A. The regulation of zinc homeostasis by the ZafA transcriptional activator is essential for Aspergillus fumigatus virulence. Mol. Microbiol. 2007, 64, 1182–1197. [Google Scholar] [CrossRef] [PubMed]
- Eide, D.J. Homeostatic and adaptive responses to zinc deficiency in Saccharomyces cerevisiae. J. Biol. Chem. 2009, 284, 18565–18569. [Google Scholar] [CrossRef] [PubMed]
- Lyons, T.J.; Gasch, A.P.; Gaither, L.A.; Botstein, D.; Brown, P.O.; Eide, D.J. Genome-wide characterization of the Zap1p zinc-responsive regulon in yeast. Proc. Natl. Acad. Sci. USA 2000, 97, 7957–7962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- North, M.; Steffen, J.; Loguinov, A.V.; Zimmerman, G.R.; Vulpe, C.D.; Eide, D.J. Genome-wide functional profiling identifies genes and processes important for zinc-limited growth of Saccharomyces cerevisiae. PLoS Genet. 2012, 8, e1002699. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Bird, A.J.; Chung, L.M.; Newton, M.A.; Winge, D.R.; Eide, D.J. Differential control of Zap1-regulated genes in response to zinc deficiency in Saccharomyces cerevisiae. BMC Genom. 2008, 9, 370. [Google Scholar] [CrossRef] [PubMed]
- Nobile, C.J.; Nett, J.E.; Hernday, A.D.; Homann, O.R.; Deneault, J.S.; Nantel, A.; Andes, D.R.; Johnson, A.D.; Mitchell, A.P. Biofilm matrix regulation by Candida albicans Zap1. PLoS Biol. 2009, 7, e1000133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, R.O.; Fogaca, N.S.; Kmetzsch, L.; Schrank, A.; Vainstein, M.H.; Staats, C.C. Zap1 regulates zinc homeostasis and modulates virulence in Cryptococcus gattii. PLoS ONE 2012, 7, e43773. [Google Scholar] [CrossRef] [PubMed]
- Hatayama, M.; Aruga, J. Gli protein nuclear localization signal. Vitam. Horm. 2012, 88, 73–89. [Google Scholar] [PubMed]
- Espeso, E.A.; Tilburn, J.; Sanchez-Pulido, L.; Brown, C.V.; Valencia, A.; Arst, H.N., Jr.; Peñalva, M.A. Specific DNA recognition by the Aspergillus nidulans three zinc finger transcription factor PacC. J. Mol. Biol. 1997, 274, 466–480. [Google Scholar] [CrossRef] [PubMed]
- Evans-Galea, M.V.; Blankman, E.; Myszka, D.G.; Bird, A.J.; Eide, D.J.; Winge, D.R. Two of the five zinc fingers in the Zap1 transcription factor DNA binding domain dominate site-specific DNA binding. Biochemistry 2003, 42, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Pavletich, N.P.; Pabo, C.O. Crystal structure of a five-finger GLI-DNA complex: New perspectives on zinc fingers. Science 1993, 261, 1701–1707. [Google Scholar] [CrossRef] [PubMed]
- Calera, J.A.; Ovejero, M.C.; López-Medrano, R.; Segurado, M.; Puente, P.; Leal, F. Characterization of the Aspergillus nidulans aspnd1 gene demonstrates that the ASPND1 antigen, which it encodes, and several Aspergillus fumigatus immunodominant antigens belong to the same family. Infect. Immun. 1997, 65, 1335–1344. [Google Scholar] [PubMed]
- Priebe, S.; Kreisel, C.; Horn, F.; Guthke, R.; Linde, J. FungiFun2: A comprehensive online resource for systematic analysis of gene lists from fungal species. Bioinformatics 2015, 31, 445–446. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Medina-Rivera, A.; Defrance, M.; Sand, O.; Herrmann, C.; Castro-Mondragon, J.A.; Delerce, J.; Jaeger, S.; Blanchet, C.; Vincens, P.; Caron, C.; et al. RSAT 2015: Regulatory Sequence Analysis Tools. Nucleic Acids Res. 2015, 43, W50–W56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zianni, M.; Tessanne, K.; Merighi, M.; Laguna, R.; Tabita, F.R. Identification of the DNA bases of a DNase I footprint by the use of dye primer sequencing on an automated capillary DNA analysis instrument. J. Biomol. Tech. 2006, 17, 103–113. [Google Scholar] [PubMed]
- Nierman, W.C.; Pain, A.; Anderson, M.J.; Wortman, J.R.; Kim, H.S.; Arroyo, J.; Berriman, M.; Abe, K.; Archer, D.B.; Bermejo, C.; et al. Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature 2005, 438, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Eide, D.J. Zap1p, a metalloregulatory protein involved in zinc-responsive transcriptional regulation in Saccharomyces cerevisiae. Mol. Cell Biol. 1997, 17, 5044–5052. [Google Scholar] [CrossRef] [PubMed]
- Do, E.; Hu, G.; Caza, M.; Kronstad, J.W.; Jung, W.H. The ZIP family zinc transporters support the virulence of Cryptococcus neoformans. Med. Mycol. 2016, 54, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Schneider Rde, O.; Diehl, C.; dos Santos, F.M.; Piffer, A.C.; Garcia, A.W.; Kulmann, M.I.; Schrank, A.; Kmetzsch, L.; Vainstein, M.H.; Staats, C.C. Effects of zinc transporters on Cryptococcus gattii virulence. Sci. Rep. 2015, 5, 10104. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Solis, N.V.; Ehrlich, R.L.; Woolford, C.A.; Filler, S.G.; Mitchell, A.P. Activation and alliance of regulatory pathways in C. albicans during mammalian infection. PLoS Biol. 2015, 13, e1002076. [Google Scholar] [CrossRef] [PubMed]
- Bottcher, B.; Palige, K.; Jacobsen, I.D.; Hube, B.; Brunke, S. Csr1/Zap1 maintains zinc homeostasis and influences virulence in Candida dubliniensis but is not coupled to morphogenesis. Eukaryot. Cell 2015, 14, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Kujoth, G.C.; Sullivan, T.D.; Merkhofer, R.; Lee, T.J.; Wang, H.; Brandhorst, T.; Wuthrich, M.; Klein, B.S. CRISPR/Cas9-mediated gene disruption reveals the importance of zinc metabolism for fitness of the dimorphic fungal pathogen Blastomyces dermatitidis. MBio 2018, 9, e00412-18. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Bird, A.J.; Winge, D.R.; Eide, D.J. Regulation of the yeast TSA1 peroxiredoxin by ZAP1 is an adaptive response to the oxidative stress of zinc deficiency. J. Biol. Chem. 2007, 282, 2184–2195. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Roje, S.; Sandoval, F.J.; Bird, A.J.; Winge, D.R.; Eide, D.J. Repression of sulfate assimilation is an adaptive response of yeast to the oxidative stress of zinc deficiency. J. Biol. Chem. 2009, 284, 27544–27556. [Google Scholar] [CrossRef] [PubMed]
- Amich, J.; Schafferer, L.; Haas, H.; Krappmann, S. Regulation of sulphur assimilation is essential for virulence and affects iron homeostasis of the human-pathogenic mould Aspergillus fumigatus. PLoS Pathog. 2013, 9, e1003573. [Google Scholar] [CrossRef] [PubMed]
- Haas, H. Fungal siderophore metabolism with a focus on Aspergillus fumigatus. Nat. Prod. Rep. 2014, 31, 1266–1276. [Google Scholar] [CrossRef] [PubMed]
- Dix, D.R.; Bridgham, J.T.; Broderius, M.A.; Byersdorfer, C.A.; Eide, D.J. The FET4 gene encodes the low affinity Fe(II) transport protein of Saccharomyces cerevisiae. J. Biol. Chem. 1994, 269, 26092–26099. [Google Scholar] [PubMed]
- Schrettl, M.; Kim, H.S.; Eisendle, M.; Kragl, C.; Nierman, W.C.; Heinekamp, T.; Werner, E.R.; Jacobsen, I.; Illmer, P.; Yi, H.; et al. SreA-mediated iron regulation in Aspergillus fumigatus. Mol. Microbiol. 2008, 70, 27–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasmin, S.; Abt, B.; Schrettl, M.; Moussa, T.A.; Werner, E.R.; Haas, H. The interplay between iron and zinc metabolism in Aspergillus fumigatus. Fungal. Genet. Biol. 2009, 46, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Schrettl, M.; Beckmann, N.; Varga, J.; Heinekamp, T.; Jacobsen, I.D.; Jochl, C.; Moussa, T.A.; Wang, S.; Gsaller, F.; Blatzer, M.; et al. HapX-mediated adaption to iron starvation is crucial for virulence of Aspergillus fumigatus. PLoS Pathog. 2010, 6, e1001124. [Google Scholar] [CrossRef] [PubMed]
- Gsaller, F.; Hortschansky, P.; Beattie, S.R.; Klammer, V.; Tuppatsch, K.; Lechner, B.E.; Rietzschel, N.; Werner, E.R.; Vogan, A.A.; Chung, D.; et al. The Janus transcription factor HapX controls fungal adaptation to both iron starvation and iron excess. EMBO J. 2014, 33, 2261–2276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDiarmid, C.W.; Milanick, M.A.; Eide, D.J. Induction of the ZRC1 metal tolerance gene in zinc-limited yeast confers resistance to zinc shock. J. Biol. Chem. 2003, 278, 15065–15072. [Google Scholar] [CrossRef] [PubMed]
- Miyabe, S.; Izawa, S.; Inoue, Y. Expression of ZRC1 coding for suppressor of zinc toxicity is induced by zinc-starvation stress in Zap1-dependent fashion in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2000, 276, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Blatzer, M.; Barker, B.M.; Willger, S.D.; Beckmann, N.; Blosser, S.J.; Cornish, E.J.; Mazurie, A.; Grahl, N.; Haas, H.; Cramer, R.A. SREBP coordinates iron and ergosterol homeostasis to mediate triazole drug and hypoxia responses in the human fungal pathogen Aspergillus fumigatus. PLoS Genet. 2011, 7, e1002374. [Google Scholar] [CrossRef] [PubMed]
- Chung, D.; Barker, B.M.; Carey, C.C.; Merriman, B.; Werner, E.R.; Lechner, B.E.; Dhingra, S.; Cheng, C.; Xu, W.; Blosser, S.J.; et al. ChIP-seq and in vivo transcriptome analyses of the Aspergillus fumigatus SREBP SrbA reveals a new regulator of the fungal hypoxia response and virulence. PLoS Pathog. 2014, 10, e1004487. [Google Scholar] [CrossRef] [PubMed]
- Vödisch, M.; Scherlach, K.; Winkler, R.; Hertweck, C.; Braun, H.P.; Roth, M.; Haas, H.; Werner, E.R.; Brakhage, A.A.; Kniemeyer, O. Analysis of the Aspergillus fumigatus proteome reveals metabolic changes and the activation of the pseurotin A biosynthesis gene cluster in response to hypoxia. J. Proteome Res. 2011, 10, 2508–2524. [Google Scholar] [CrossRef] [PubMed]
- Maguire, S.L.; Wang, C.; Holland, L.M.; Brunel, F.; Neuveglise, C.; Nicaud, J.M.; Zavrel, M.; White, T.C.; Wolfe, K.H.; Butler, G. Zinc finger transcription factors displaced SREBP proteins as the major Sterol regulators during Saccharomycotina evolution. PLoS Genet. 2014, 10, e1004076. [Google Scholar] [CrossRef] [PubMed]
- Baumann, K.; Dato, L.; Graf, A.B.; Frascotti, G.; Dragosits, M.; Porro, D.; Mattanovich, D.; Ferrer, P.; Branduardi, P. The impact of oxygen on the transcriptome of recombinant S. cerevisiae and P. pastoris—A comparative analysis. BMC Genom. 2012, 12, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Synnott, J.M.; Guida, A.; Mulhern-Haughey, S.; Higgins, D.G.; Butler, G. Regulation of the hypoxic response in Candida albicans. Eukaryot. Cell 2010, 9, 1734–1746. [Google Scholar] [CrossRef] [PubMed]
- Kroll, K.; Pahtz, V.; Hillmann, F.; Vaknin, Y.; Schmidt-Heck, W.; Roth, M.; Jacobsen, I.D.; Osherov, N.; Brakhage, A.A.; Kniemeyer, O. Identification of hypoxia-inducible target genes of Aspergillus fumigatus by transcriptome analysis reveals cellular respiration as an important contributor to hypoxic survival. Eukaryot. Cell 2014, 13, 1241–1253. [Google Scholar] [CrossRef] [PubMed]
- Camarasa, C.; Faucet, V.; Dequin, S. Role in anaerobiosis of the isoenzymes for Saccharomyces cerevisiae fumarate reductase encoded by OSM1 and FRDS1. Yeast 2007, 24, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Holmstrom, K.M.; Finkel, T. Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Messner, K.R.; Imlay, J.A. Mechanism of superoxide and hydrogen peroxide formation by fumarate reductase, succinate dehydrogenase, and aspartate oxidase. J. Biol. Chem. 2002, 277, 42563–42571. [Google Scholar] [CrossRef] [PubMed]
- Chaiyen, P.; Fraaije, M.W.; Mattevi, A. The enigmatic reaction of flavins with oxygen. Trends Biochem. Sci. 2012, 37, 373–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korshunov, S.; Imlay, J.A. Two sources of endogenous hydrogen peroxide in Escherichia coli. Mol. Microbiol. 2010, 75, 1389–1401. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Steffen, J.; Eide, D.J. Cytosolic superoxide dismutase (SOD1) is critical for tolerating the oxidative stress of zinc deficiency in yeast. PLoS ONE 2009, 4, e7061. [Google Scholar] [CrossRef] [PubMed]
- MacDiarmid, C.W.; Taggart, J.; Kerdsomboon, K.; Kubisiak, M.; Panascharoen, S.; Schelble, K.; Eide, D.J. Peroxiredoxin chaperone activity is critical for protein homeostasis in zinc-deficient yeast. J. Biol. Chem. 2013, 288, 31313–31327. [Google Scholar] [CrossRef] [PubMed]
- Bignell, E.; Cairns, T.C.; Throckmorton, K.; Nierman, W.C.; Keller, N.P. Secondary metabolite arsenal of an opportunistic pathogenic fungus. Philos. Trans. R Soc. Lond. B Biol. Sci. 2016, 371, 20160023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scharf, D.H.; Remme, N.; Heinekamp, T.; Hortschansky, P.; Brakhage, A.A.; Hertweck, C. Transannular disulfide formation in gliotoxin biosynthesis and its role in self-resistance of the human pathogen Aspergillus fumigatus. J. Am. Chem. Soc. 2010, 132, 10136–10141. [Google Scholar] [CrossRef] [PubMed]
- Schrettl, M.; Carberry, S.; Kavanagh, K.; Haas, H.; Jones, G.W.; O’Brien, J.; Nolan, A.; Stephens, J.; Fenelon, O.; Doyle, S. Self-protection against gliotoxin—A component of the gliotoxin biosynthetic cluster, GliT, completely protects Aspergillus fumigatus against exogenous gliotoxin. PLoS Pathog. 2010, 6, e1000952. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.N.; Toyotome, T.; Muraosa, Y.; Watanabe, A.; Wuren, T.; Bunsupa, S.; Aoyagi, K.; Yamazaki, M.; Takino, M.; Kamei, K. GliA in Aspergillus fumigatus is required for its tolerance to gliotoxin and affects the amount of extracellular and intracellular gliotoxin. Med. Mycol. 2014, 52, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Dolan, S.K.; Owens, R.A.; O’Keeffe, G.; Hammel, S.; Fitzpatrick, D.A.; Jones, G.W.; Doyle, S. Regulation of nonribosomal peptide synthesis: Bis-thiomethylation attenuates gliotoxin biosynthesis in Aspergillus fumigatus. Chem. Biol. 2014, 21, 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Owens, R.A.; O’Keeffe, G.; Smith, E.B.; Dolan, S.K.; Hammel, S.; Sheridan, K.J.; Fitzpatrick, D.A.; Keane, T.M.; Jones, G.W.; Doyle, S. Interplay between gliotoxin resistance, secretion, and the methyl/methionine cycle in Aspergillus fumigatus. Eukaryot. Cell 2015, 14, 941–957. [Google Scholar] [CrossRef] [PubMed]
- Owens, R.A.; Hammel, S.; Sheridan, K.J.; Jones, G.W.; Doyle, S. A proteomic approach to investigating gene cluster expression and secondary metabolite functionality in Aspergillus fumigatus. PLoS ONE 2014, 9, e106942. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, L.; Owens, R.A.; Dolan, S.K.; O’Keeffe, G.; Schrettl, M.; Kavanagh, K.; Jones, G.W.; Doyle, S. The Aspergillus fumigatus protein GliK protects against oxidative stress and is essential for gliotoxin biosynthesis. Eukaryot. Cell 2012, 11, 1226–1238. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.S.; Shim, J.S.; Kim, J.A.; Kang, S.W.; Kwon, H.J. Discovery of gliotoxin as a new small molecule targeting thioredoxin redox system. Biochem. Biophys. Res. Commun. 2007, 359, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Yin, X.; Wang, X.; Zhou, P.; Guo, F.B. Re-annotation of protein-coding genes in the genome of Saccharomyces cerevisiae based on support vector machines. PLoS ONE 2013, 8, e64477. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Butler, E.; Rodgers, J.; Spizzo, T.; Duesterhoeft, S.; Eide, D. Regulation of zinc homeostasis in yeast by binding of the ZAP1 transcriptional activator to zinc-responsive promoter elements. J. Biol. Chem. 1998, 273, 28713–28720. [Google Scholar] [CrossRef] [PubMed]
- Pavletich, N.P.; Pabo, C.O. Zinc finger-DNA recognition: Crystal structure of a Zif268-DNA complex at 2.1 A. Science 1991, 252, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Nagaich, A.K.; Appella, E.; Harrington, R.E. DNA bending is essential for the site-specific recognition of DNA response elements by the DNA binding domain of the tumor suppressor protein p53. J. Biol. Chem. 1997, 272, 14842–14849. [Google Scholar] [CrossRef] [PubMed]
- Bird, A.J.; Blankman, E.; Stillman, D.J.; Eide, D.J.; Winge, D.R. The Zap1 transcriptional activator also acts as a repressor by binding downstream of the TATA box in ZRT2. EMBO J. 2004, 23, 1123–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bird, A.J.; Gordon, M.; Eide, D.J.; Winge, D.R. Repression of ADH1 and ADH3 during zinc deficiency by Zap1-induced intergenic RNA transcripts. EMBO J. 2006, 25, 5726–5734. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Detailed Genotype | Reference |
|---|---|---|
| CEA17 | pyrG1 (auxotrophic PyrG–) | [11] |
| AF14 | Wild-type (isogenic to CEA17) | [12] |
| AF171 | ΔzafA::hisG (isogenic to CEA17) | [14] |
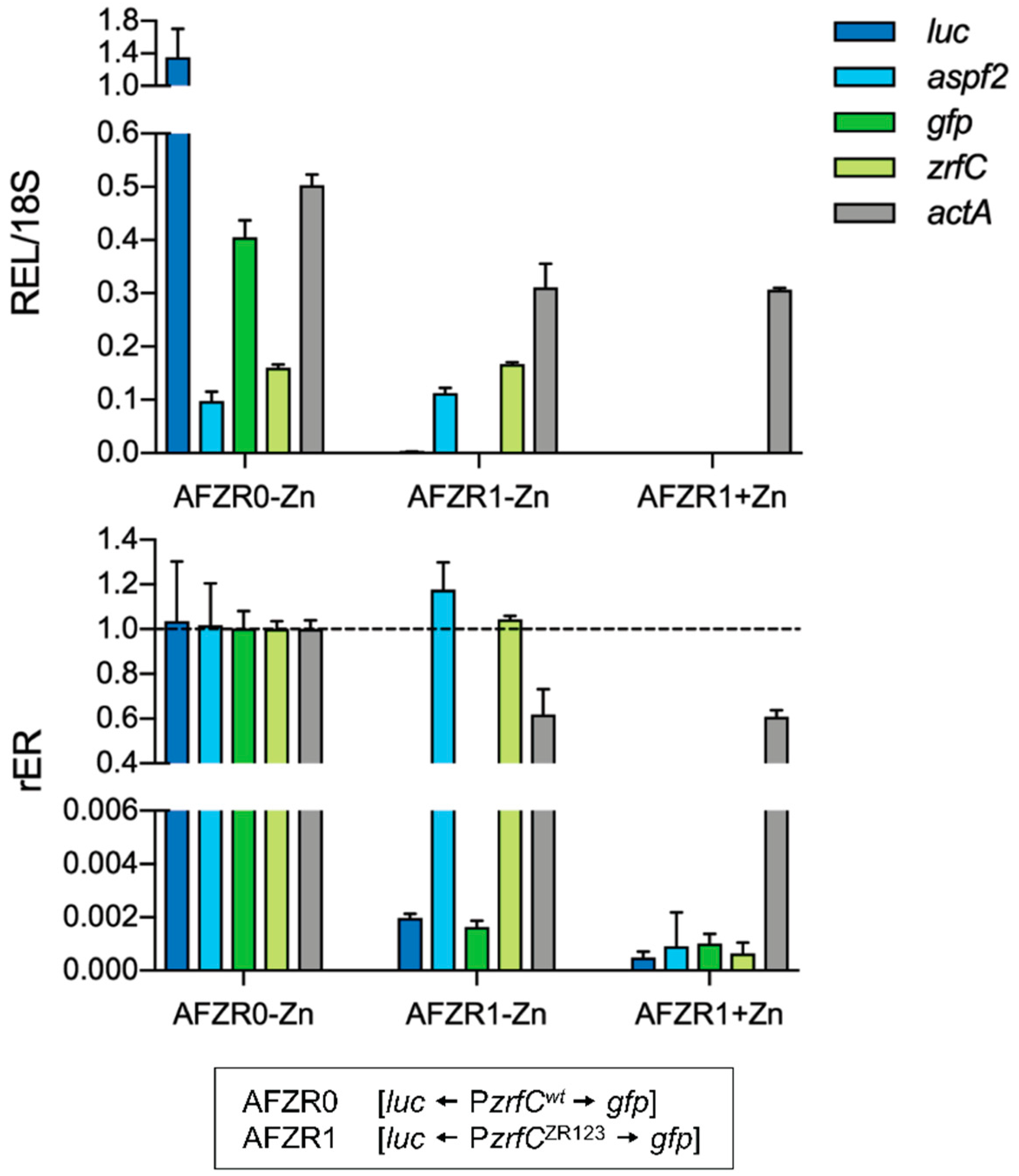
| AFZR0 | Wild-type [luc ← PzrfCwt → gfp] (isogenic to CEA17) | This study |
| AFZR1 | Wild-type [luc ← PzrfCZR123 → gfp] (isogenic to CEA17) | This study |
| Gene ID | Group of Genes | Gene Name | wt − Zn/wt + Zn rER ± SD | ∆zafA − Zn/wt − Zn rER ± SD | Predicted Effect of ZafA on Gene Expression | Reallocating Subgroup of Genes * |
|---|---|---|---|---|---|---|
| AFUA_1G01550 | DZR | zrfA | 231.3 ± 27.8 | −207.7 ± 18.9 | Induction | |
| AFUA_1G02150 | Control | gfdA | −1.25 ± 0.15 | 1.17 ± 0.11 | ||
| AFUA_1G03150 | DZR | erg24A | −49.9 ± 7.0 | 2.0 ± 0.2 | Repression | |
| AFUA_1G04620 | IZR | - | −4.9 ± 0.4 | −1.9 ± 0.2 | ||
| AFUA_1G07480 | NZR | hem13 | −6.0 ± 0.8 | −1.6 ± 0.1 | IZR | |
| AFUA_1G10060 | NZR | - | 3.1 ± 0.2 | −1.2 ± 0.1 | ||
| AFUA_1G10080 | DZR | zafA | 31.5 ± 3.0 | −21582 ± 4316 | Induction | |
| AFUA_1G10130 | NZR | sahA | 1.9 ± 0.1 | 1.1 ± 0.1 | ||
| AFUA_1G12830 | IZR | niaD | −1.6 ± 0.1 | −5.2 ± 0.7 | ||
| AFUA_1G13510 | NZR | facB | 4.6 ± 0.6 | 1.0 ± 0.1 | ||
| AFUA_1G14550 | DZR | sodC/sod3 | 1698.7 ± 198.2 | −1176.5 ± 142.4 | Induction | |
| AFUA_1G15590 | NZR | cybS/sdh4 | 2.9 ± 0.4 | 1.1 ± 0.1 | ||
| AFUA_1G17190 | NZR | sidI | −92.5 ± 8.7 | 1.1 ± 0.1 | ||
| AFUA_2G00320 | NZR | erg3B | −271.7 ± 31.7 | −1.6 ± 0.2 | IZR | |
| AFUA_2G01260 | DZR | srbA | −15.8 ± 2.1 | 2.1 ± 0.2 | Repression | |
| AFUA_2G03010 | NZR | - | −1.7 ± 0.2 | −1.0 ± 0.1 | ||
| AFUA_2G03700 | NZR | hmg1 | −2.6 ± 0.2 | −1.1 ± 0.1 | ||
| AFUA_2G03860 | DZR | zrfB | 33.0 ± 2.7 | −79.3 ± 7.7 | Induction | |
| AFUA_2G07680 | IZR | sidA | −27.3 ± 3.3 | −6.9 ± 0.6 | ||
| AFUA_2G08740 | DZR | zrfF | 24.8 ± 1.8 | −16.0 ± 2.0 | Induction | |
| AFUA_2G11120 | DZR | gtmA | 5.7 ± 0.7 | −20.3 ± 1.8 | Induction | |
| AFUA_2G14570 | NZR | zrcC | −2.1 ± 0.2 | −1.1 ± 0.1 | ||
| AFUA_2G15010 | NZR | - | −1.5 ± 0.2 | −1.2 ± 0.1 | ||
| AFUA_2G15290 | DZR | - | −26.0 ± 3.6 | 3.2 ± 0.3 | Repression | |
| AFUA_2G17550 | IZR | ayg1 | −2.7 ± 0.4 | −103.5 ± 10.8 | ||
| AFUA_3G02270 | DZR | cat1 | 18.9 ± 1.8 | −2.0± 0.2 | Induction | |
| AFUA_3G03640 | IZR | mirB | −3225.8 ± 304.5 | −10.0 ± 1.2 | ||
| AFUA_3G14440 | NZR | - | −3.5 ± 0.5 | −1.3 ± 0.1 | ||
| AFUA_4G03410 | NZR | fhpA | −3.1 ± 0.3 | −1.4 ± 0.2 | ||
| AFUA_4G03460 | IZR | srbB | −50.4 ± 5.9 | −1.1 ± 0.1 | NZR | |
| AFUA_4G03930 | DZR | cysX | 12786.9 ± 1492.2 | −7751.9 ± 938.0 | Induction | |
| AFUA_4G06530 | NZR | metR | −1.6 ± 0.2 | 1.2 ± 0.1 | ||
| AFUA_4G09560 | DZR | zrfC | 1270.1 ± 108.0 | −5377.1 ± 424.8 | Induction | |
| AFUA_4G10730 | Control | rvb1 | 1.1 ± 0.2 | −1.2 ± 0.1 | ||
| AFUA_4G12840 | IZR | - | −1.6 ± 0.2 | −1.7 ± 0.1 | ||
| AFUA_5G01030 | DZR | gpdB | −1.2 ± 0.1 | 77.7 ± 7.5 | SSA | |
| AFUA_5G01970 | NZR | gpdA | 1.8 ± 0.2 | 1.0 ± 0.1 | ||
| AFUA_5G02180 | NZR | cysB | −2.6 ± 0.2 | 1.1 ± 0.1 | ||
| AFUA_5G03800 | NZR | ftrA | −31.0 ± 2.2 | 1.4 ± 0.2 | ||
| AFUA_5G03920 | NZR | hapX | −12.2 ± 1.5 | −1.2 ± 0.1 | ||
| AFUA_5G04130 | Control | phoA | 1.1 ± 0.1 | −1.3 ± 0.2 | ||
| AFUA_5G06240 | DZR | alcC | −26.8 ± 1.9 | 3.8 ± 0.5 | Repression | |
| AFUA_5G06270 | NZR | hemA | 1.8 ± 0.2 | −1.0 ± 0.1 | ||
| AFUA_5G08090 | DZR | pyroA | 2.7 ± 0.3 | −1.8 ± 0.2 | Induction | |
| AFUA_5G09240 | DZR | sodA/sod1 | −4.7 ± 0.7 | 7.3 ± 0.8 | Repression | |
| AFUA_5G09360 | Control | calA | 1.1 ± 0.1 | 1.1 ± 0.1 | ||
| AFUA_5G09680 | NZR | carC/sdh3 | 3.9 ± 0.4 | −1.0 ± 0.1 | ||
| AFUA_5G10560 | Control | cox5A | −1.2 ± 0.2 | 1.0 ± 0.1 | ||
| AFUA_6G00690 | DZR | - | −2.8 ± 0.3 | 5.2 ± 0.6 | Repression | |
| AFUA_6G04430 | NZR | - | 2.0 ± 0.2 | 1.3 ± 0.2 | ||
| AFUA_6G04740 | Control | actA | −1.0 ± 0.1 | 1.2 ± 0.1 | ||
| AFUA_6G05160 | DZR | azf1 | 1.9 ± 0.2 | −23.9 ± 2.9 | Induction | |
| AFUA_6G07720 | IZR | acuF | −2.2 ± 0.2 | −1.7 ± 0.2 | ||
| AFUA_6G09630 | DZR | gliZ | 45.9 ± 6.0 | −67.2 ± 5.3 | Induction | |
| AFUA_6G09710 | DZR | gliA | 3548.5 ± 87.4 | −3039.5 ± 294.8 | Induction | |
| AFUA_6G09740 | DZR | gliT | 53.6 ± 3.9 | −477.3 ± 59.7 | Induction | |
| AFUA_7G00250 | IZR | tubB2 | −7.0 ± 0.5 | −10.0 ± 1.2 | ||
| AFUA_7G02560 | Control | dld1 | −1.1 ± 0.1 | 1.0 ± 0.1 | ||
| AFUA_7G04730 | IZR | enb1 | −48.3 ± 5.8 | −2.4 ± 0.2 | ||
| AFUA_7G05070 | NZR | frdA | −56.5 ± 7.4 | 1.9 ± 0.1 | Repression | DZR |
| AFUA_7G06570 | DZR | zrcA | −5.9 ± 0.7 | 16.6 ± 1.5 | Repression | |
| AFUA_7G06790 | DZR | yct1 | 6039.9 ± 362.4 | −5555.6 ± 577.8 | Induction | |
| AFUA_8G02450 | DZR | gzbA | 84.8 ± 8.0 | −275.5 ± 32.2 | Induction | |
| AFUA_8G02620 | DZR | mchC | 1155.6 ± 134.9 | −524.3 ± 63.4 | Induction |
| Gene ID | Gene Name | Strand | Start | End | Sequence ZR Motif | Type of ZR Motif |
|---|---|---|---|---|---|---|
| AFUA_1G01550 | zrfA | D | −75 | −61 | GACCTCAAGGTACCC | ZR2 |
| R | −105 | −91 | ACTGCCAAGGTCCTT | ZR0 | ||
| D | −157 | −143 | GCTCACAAGGTGTGT | ZR0* | ||
| AFUA_1G05900 | - | D | −393 | −379 | TTCGTCAAGGTTGTT | ZR0* |
| AFUA_1G09810 | - | R | −168 | −154 | ACTCCCAGGGTACTT | ZR0 |
| AFUA_1G10080 | zafA | D | −361 | −347 | GCTCCCAAGGTCCTC | ZR0 |
| D | −448 | −434 | CCCCTCAGGGTATTA | ZR0* | ||
| D | −624 | −610 | GCTCGCAGGGTCCTC | ZR5 | ||
| D | −657 | −643 | TCCCCCAGGGTCCTC | ZR0 | ||
| AFUA_1G12170 | - | R | −364 | −350 | ATTACCAAGGTGATA | ZR0* |
| AFUA_1G12850 | crnA | R | −500 | −486 | TGTCCCAGGGTGTAC | ZR0* |
| AFUA_1G14550 | sodC/sod3 | R | −123 | −109 | AGCATCAAGGTCCTC | ZR2 |
| D | −160 | −146 | GATCCCAAGGTCCCC | ZR2 | ||
| AFUA_1G14560 | msdS | D | −664 | −650 | TCTACCAGGGTATAT | ZR8 |
| D | −928 | −914 | ATCGCCAGGGTCTTA | ZR9 | ||
| AFUA_1G14700 | - | R | −181 | −167 | GCACACAAGGTACTC | ZR0* |
| AFUA_2G01260 | srbA | D | −886 | −872 | TTGGTCAAGGTCCTT | ZR3 |
| AFUA_2G01610 | - | D | −841 | −827 | GTCAGCAGGGTCATC | ZR0* |
| AFUA_2G02950 | - | D | −160 | −146 | GTCTCCAAGGTCTCC | ZR4 |
| AFUA_2G03860 | zrfB | R | −359 | −345 | ATCCTCAAGGTCCCT | ZR0 |
| D | −496 | −482 | GCCACCAAGGTCCTT | ZR0 | ||
| D | −794 | −780 | TCTCCCAAGGTCCCC | ZR0 | ||
| D | −893 | −879 | ACTCCCAAGGTCCTC | ZR0 | ||
| AFUA_2G06140 | - | R | −461 | −447 | GTTCCCAAGGTCTCC | ZR0 |
| AFUA_2G07810 | - | D | −756 | −742 | GTAGGCAAGGTGCCT | ZR0* |
| AFUA_2G08280 | maeA | R | −910 | −896 | TTCACCAGGGTAGAC | ZR0* |
| AFUA_2G08740 | zrfF | R | −329 | −315 | TCTCCCAAGGTCCGC | ZR8 |
| R | −616 | −602 | ATAGCCAAGGTAGCT | ZR0* | ||
| AFUA_2G15290 | - | R | −68 | −54 | GTCAACAGGGTGCTC | ZR5 |
| AFUA_3G02270 | cat1 | D | −1052 | −1038 | TTGACCAGGGTCTCT | ZR3 |
| AFUA_3G10680 | - | R | −179 | −165 | GATCCCAAGGTCCTT | ZR2 |
| D | −233 | −219 | TCCCCCAGGGTCCCC | ZR0 | ||
| AFUA_3G13100 | - | D | −581 | −567 | TTCGTCAAGGTTTCA | ZR0* |
| AFUA_3G13940 | - | D | −1073 | −1059 | TTTTCCAAGGTGTTG | ZR0 * |
| AFUA_4G03920 | - | R | −307 | −293 | CTCACCAAGGTCCCC | ZR1 |
| D | −438 | −424 | ACCTCCAAGGTCCTG | ZR0* | ||
| R | −576 | −562 | TCTATCAAGGTAATT | ZR7 | ||
| R | −605 | −591 | GCCTCCAAGGTAGTC | ZR0* | ||
| AFUA_4G03930 | cysX | D | −272 | −258 | GCCTCCAAGGTAGTC | ZR0* |
| D | −301 | −287 | TCTATCAAGGTAATT | ZR1 | ||
| R | −439 | −425 | ACCTCCAAGGTCCTG | ZR0* | ||
| D | −570 | −556 | CTCACCAAGGTCCCC | ZR7 | ||
| AFUA_4G09560 | zrfC | R | −196 | −182 | ACTCCCAAGGTATCT | ZR0 |
| R | −313 | −299 | GCTCTCAAGGTCCTC | ZR0 | ||
| R | −637 | −623 | GCTACCAAGGTGTCC | ZR0 | ||
| D | −749 | −735 | GCCATCAGGGTAGAC | ZR0* | ||
| AFUA_4G09580 | aspf2 | R | −149 | −135 | GCCATCAGGGTAGAC | ZR0* |
| D | −261 | −247 | GCTACCAAGGTGTCC | ZR0 | ||
| D | −585 | −571 | GCTCTCAAGGTCCTC | ZR0 | ||
| D | −702 | −688 | ACTCCCAAGGTATCT | ZR0 | ||
| AFUA_4G10460 | hcsA | R | −538 | −524 | ATCGGCAAGGTACAT | ZR0* |
| AFUA_5G02010 | - | D | −122 | −108 | TTCTCCAGGGTCTTA | ZR0* |
| AFUA_5G03060 | - | R | −807 | −793 | GCGCGCAAGGTACTT | ZR0* |
| AFUA_5G05710 | - | R | −100 | −86 | TTCACCAAGGTTTTG | ZR0* |
| AFUA_5G06240 | alcC | D | −595 | −581 | TCCCCCAGGGTACAT | ZR8 |
| AFUA_5G09240 | sodA/sod1 | R | −198 | −184 | ATTCACAGGGTATTA | ZR0* |
| AFUA_5G12780 | - | D | −99 | −85 | ACTCCCAAGGTACTC | ZR0 |
| AFUA_5G13940 | - | R | −231 | −217 | TCTGTCAGGGTCTGT | ZR8 |
| AFUA_6G00690 | - | D | −77 | −63 | GTCCACAAGGTCTTC | ZR5 |
| AFUA_6G08580 | fkbp4 | D | −134 | −120 | GCCCTCAAGGTTCCT | ZR6 |
| AFUA_6G09310 | - | D | −1068 | −1054 | AGTGACAAGGTATTC | ZR0* |
| AFUA_6G09630 | gliZ | D | −790 | −776 | TCTAACAAGGTCCTC | ZR5 |
| D | −836 | −822 | GCCCCCAAGGTGCCT | ZR0 | ||
| AFUA_6G09710 | gliA | R | −469 | −455 | GTTTGCAAGGTACTC | ZR0* |
| R | −522 | −508 | TCCCCCAAGGTCACA | ZR0* | ||
| AFUA_6G10260 | akr1 | D | −180 | −166 | GTCGTCAAGGTTCCC | ZR6 |
| AFUA_7G02360 | - | R | −323 | −309 | TCGATCAAGGTGCTT | ZR3 |
| AFUA_7G03970 | - | D | −396 | −382 | TGTACCAAGGTATGT | ZR0* |
| AFUA_7G06570 | zrcA | D | −915 | −901 | GTCCCCAAGGTACTC | ZR0 |
| AFUA_7G06790 | yct1 | D | −224 | −210 | GTTCACAAGGTTCTT | ZR0* |
| D | −305 | −291 | TCTACCAAGGTCCTT | ZR0 | ||
| AFUA_7G06810 | sarA | R | −187 | −173 | TCCAACAAGGTACCT | ZR5 |
| R | −276 | −262 | CCTCCCAGGGTTCTC | ZR0* | ||
| AFUA_8G01930 | - | R | −603 | −589 | GCCATCAAGGTCGAT | ZR0* |
| AFUA_8G02450 | gzbA | R | −165 | −151 | GTCTTCAAGGTTCTC | ZR0* |
| D | −401 | −387 | GTCCCCAAGGTTCTC | ZR6 | ||
| AFUA_8G02460 | - | R | −617 | −603 | GTCCCCAAGGTTCTC | ZR6 |
| D | −853 | −839 | GTCTTCAAGGTTCTC | ZR0* | ||
| AFUA_8G02620 | mchC | R | −65 | −51 | ACCCCCAAGGTTCGC | ZR0* |
| R | −172 | −158 | ACTCCCAAGGTACCT | ZR0 | ||
| R | −192 | −178 | GGGCTCAAGGTCCTC | ZR0* |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vicentefranqueira, R.; Amich, J.; Marín, L.; Sánchez, C.I.; Leal, F.; Calera, J.A. The Transcription Factor ZafA Regulates the Homeostatic and Adaptive Response to Zinc Starvation in Aspergillus fumigatus. Genes 2018, 9, 318. https://doi.org/10.3390/genes9070318
Vicentefranqueira R, Amich J, Marín L, Sánchez CI, Leal F, Calera JA. The Transcription Factor ZafA Regulates the Homeostatic and Adaptive Response to Zinc Starvation in Aspergillus fumigatus. Genes. 2018; 9(7):318. https://doi.org/10.3390/genes9070318
Chicago/Turabian StyleVicentefranqueira, Rocío, Jorge Amich, Laura Marín, Clara Inés Sánchez, Fernando Leal, and José Antonio Calera. 2018. "The Transcription Factor ZafA Regulates the Homeostatic and Adaptive Response to Zinc Starvation in Aspergillus fumigatus" Genes 9, no. 7: 318. https://doi.org/10.3390/genes9070318
APA StyleVicentefranqueira, R., Amich, J., Marín, L., Sánchez, C. I., Leal, F., & Calera, J. A. (2018). The Transcription Factor ZafA Regulates the Homeostatic and Adaptive Response to Zinc Starvation in Aspergillus fumigatus. Genes, 9(7), 318. https://doi.org/10.3390/genes9070318