Host-Pathogen Interactions Mediated by MDR Transporters in Fungi: As Pleiotropic as it Gets!



Abstract
:1. Introduction
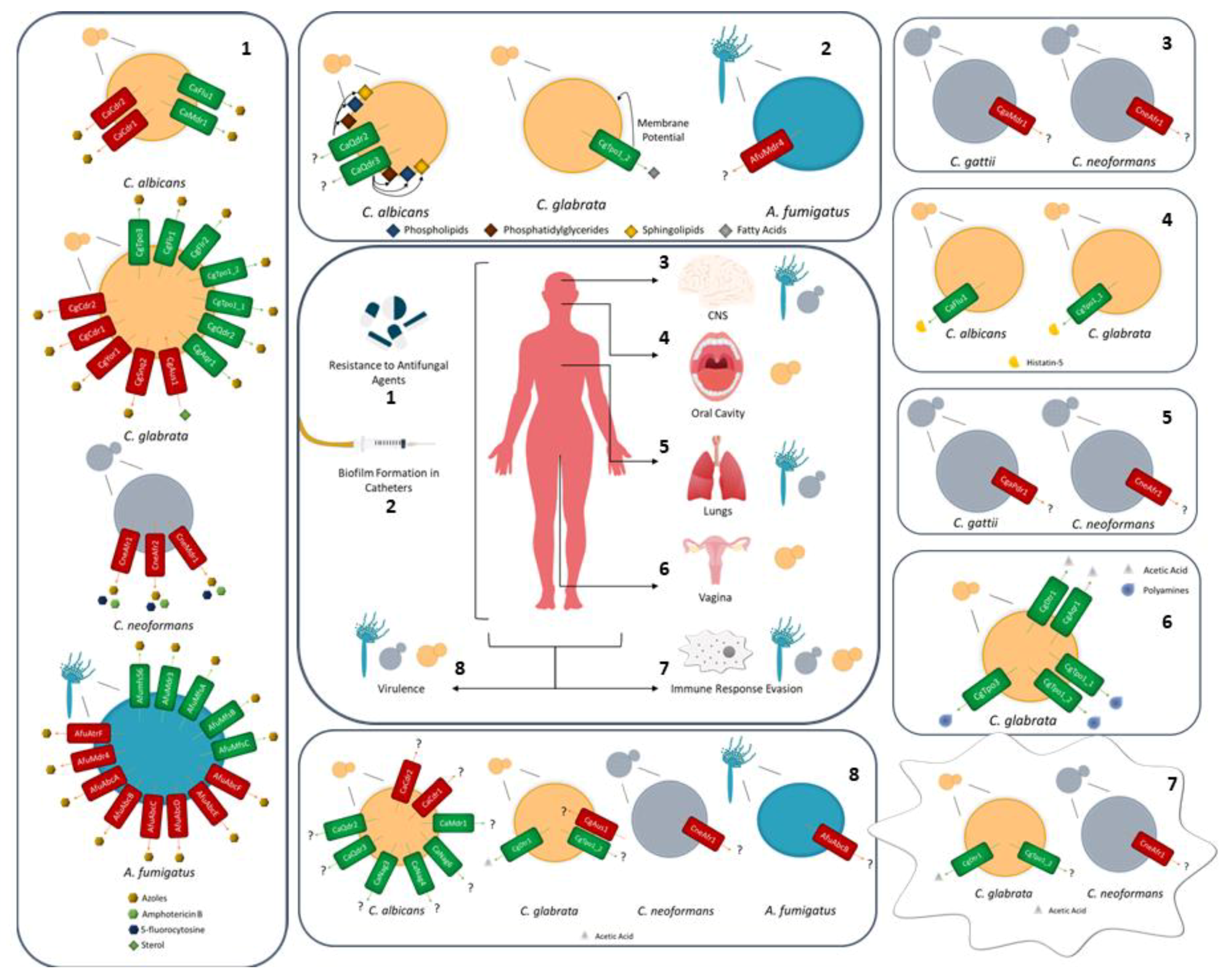
2. MDR Transporters in Fungal Pathogens: Mediators of Azole Drug Resistance
3. MDR Transporters in Fungal Pathogens: Mediating the Transport of Physiological Substrates as a Way to Adapt to Host Niches
4. MDR Transporters in Fungal Pathogens: Playing a Role in Virulence, Biofilm Formation, and Phagocytosis Evasion
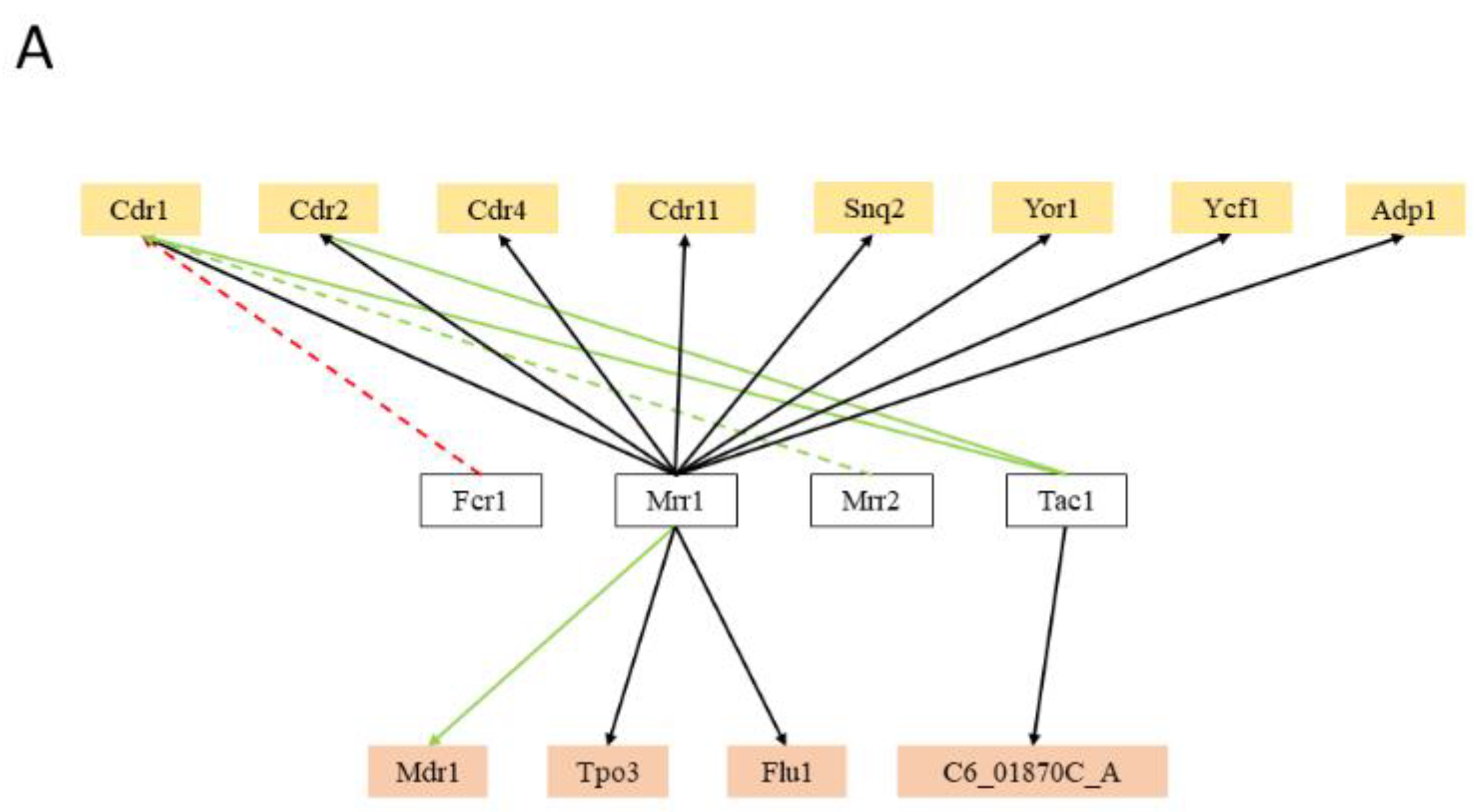
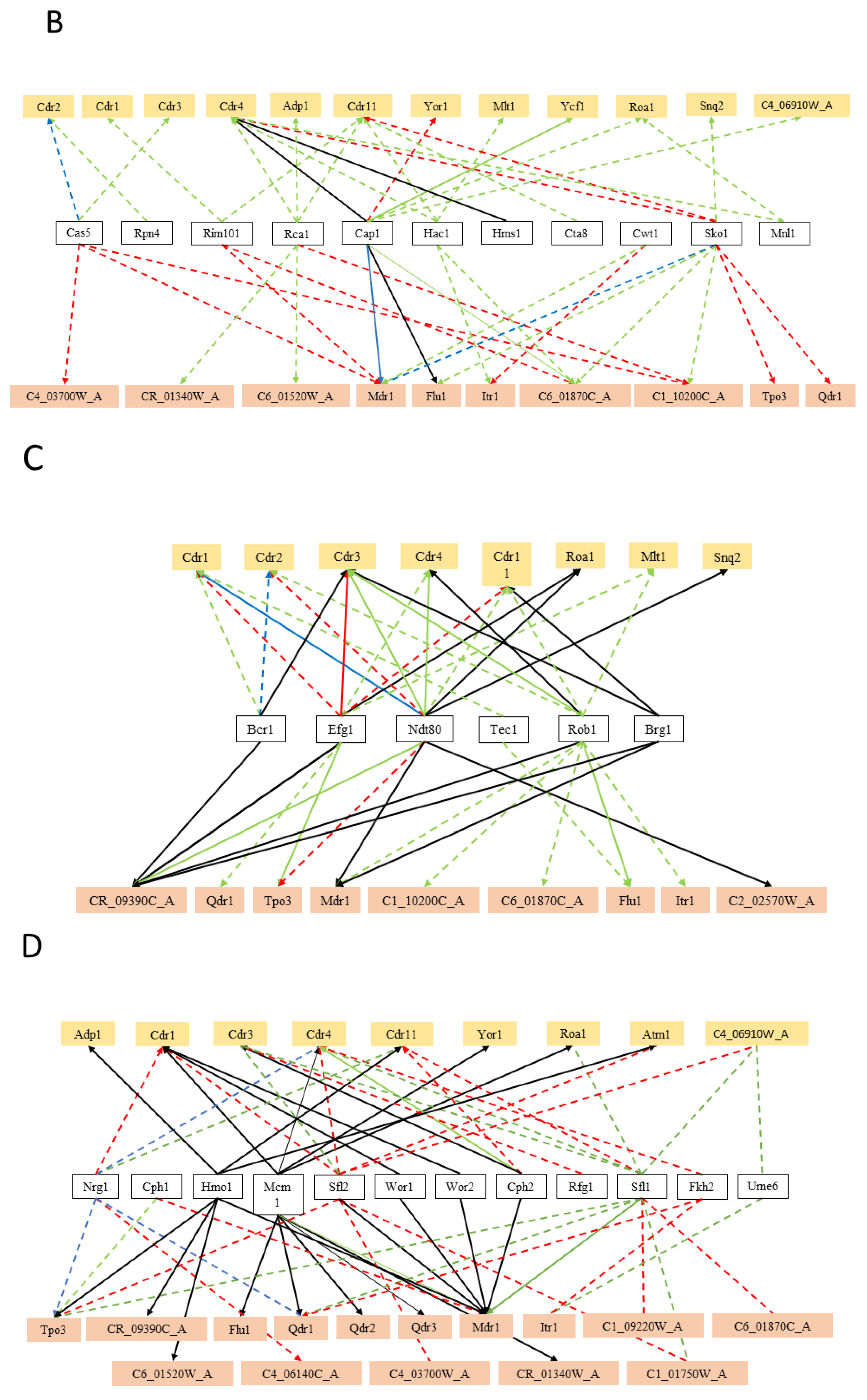
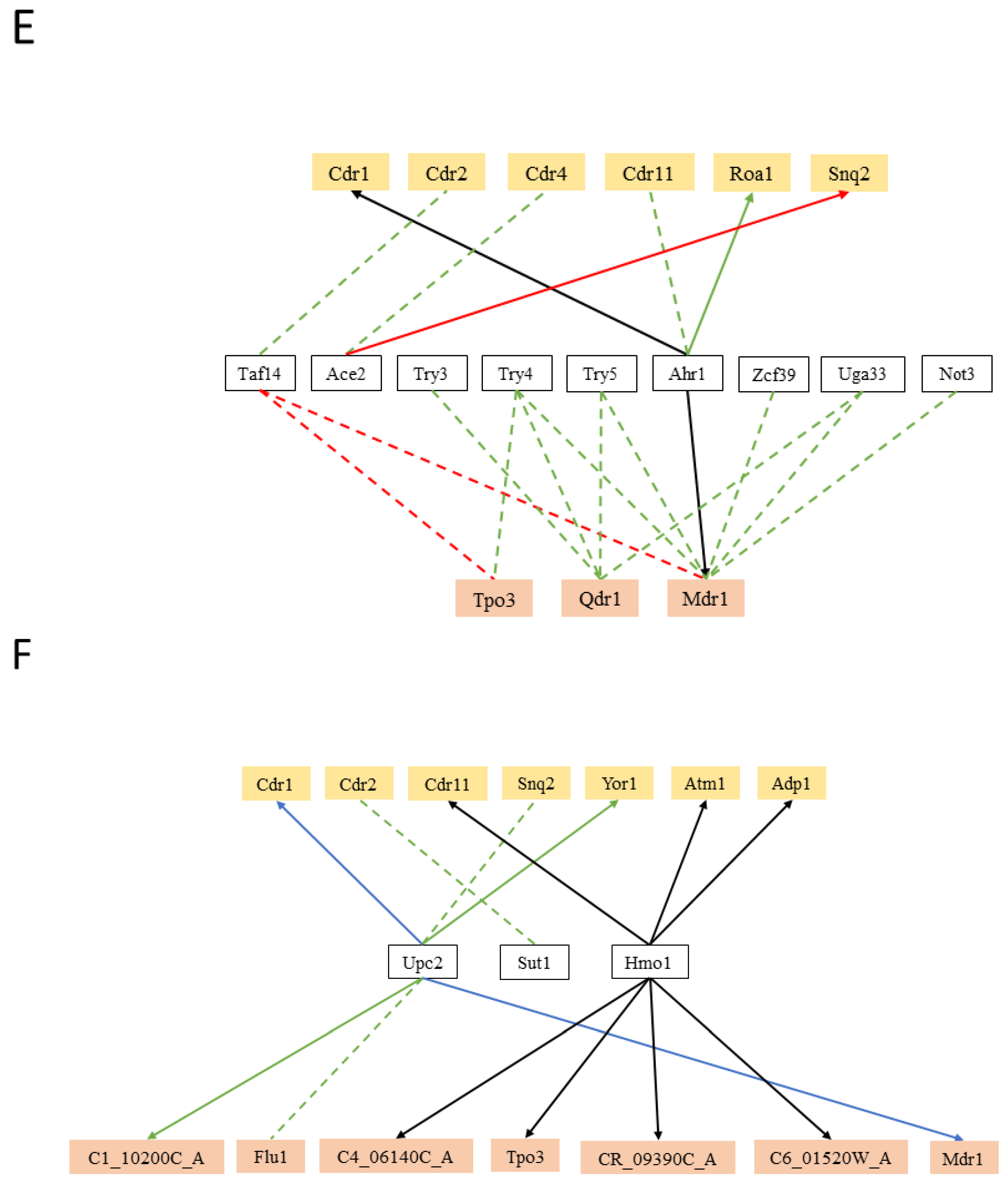
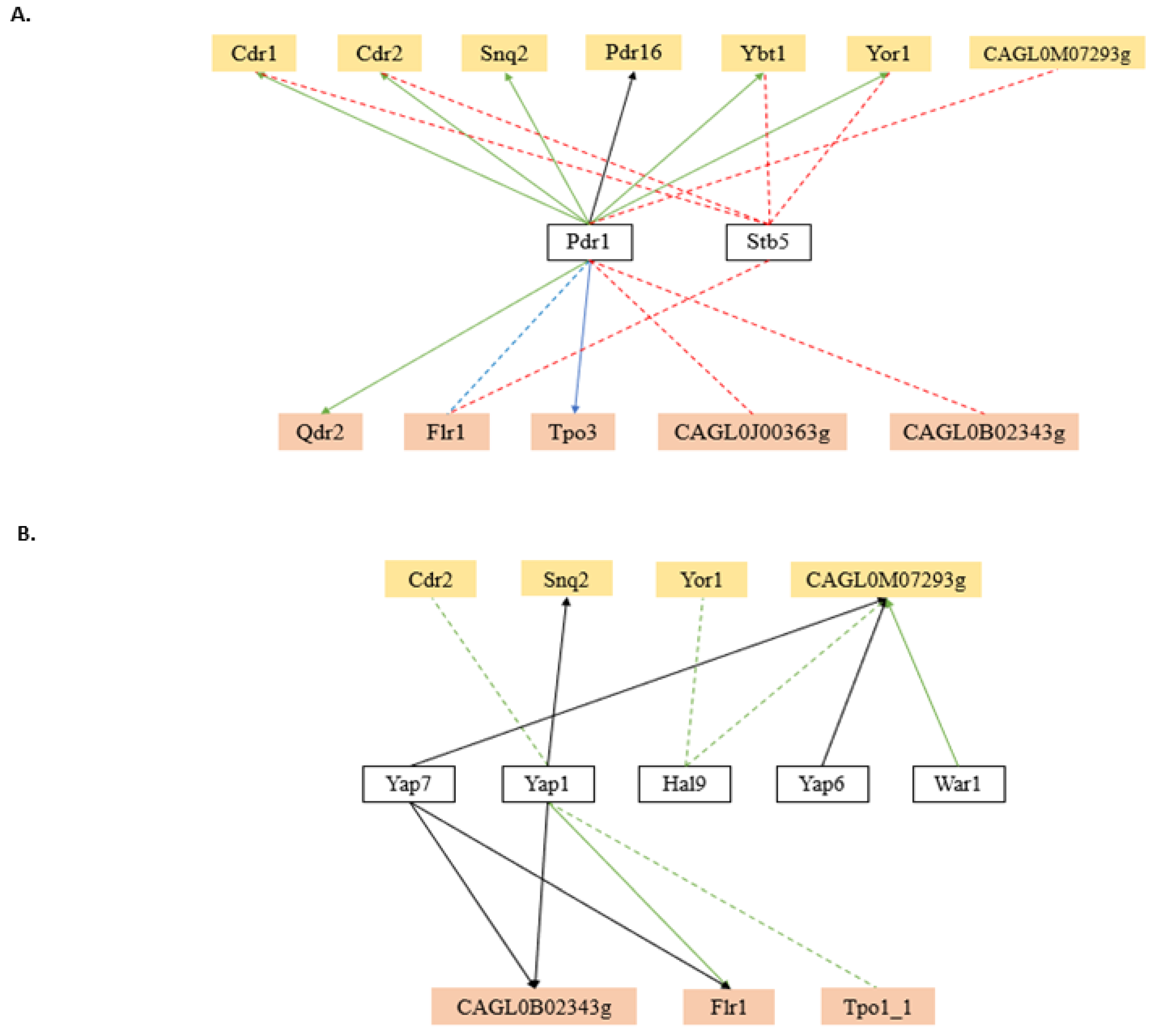
5. Hints on the Function of MDR Transporters Based on Transcription Regulation
6. Conclusions and Perspectives.
Funding
Conflicts of Interest
References
- Sims, C.R.; Ostrosky-Zeichner, L.; Rex, J.H. Invasive candidiasis in immunocompromised hospitalized patients. Arch. Med. Res. 2005, 36, 660–671. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.; Latge, J.-P.; Calderone, R. Signalling and oxidant adaptation in Candida albicans and Aspergillus fumigatus. Nat. Rev. Microbiol. 2006, 4, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Schmalzle, S.A.; Buchwald, U.K.; Gilliam, B.L.; Riedel, D.J. Cryptococcus neoformans infection in malignancy. Mycoses Diagn. Ther. Prophyl. Fungal Dis. 2016, 59, 542–552. [Google Scholar] [CrossRef]
- Wisplinghoff, H.; Bischoff, T.; Tallent, S.M.; Seifert, H.; Wenzel, R.P.; Edmond, M.B. Nosocomial bloodstream infections in US hospitals: Analysis of 24,179 cases from a prospective nationwide surveillance study. Clin. Infect. Dis. 2004, 39, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Fidel, P.L.; Vazquez, J.A.; Sobel, J.D. Candida glabrata: Review of epidemiology, pathogenesis, and clinical disease with comparison to C albicans. Clin. Microbiol. Rev. 1999, 12, 80–96. [Google Scholar] [PubMed]
- Mccullough, M.J.; Ross, B.C.; Reade, P.C. Candida albicans: A review of its history, taxonomy, epidemiology, virulence attributes, and methods of strain differentiation. Oral Maxillofac. Surg. 1996, 25, 136–144. [Google Scholar] [CrossRef]
- Da Silva Ferreira, M.E.S.; Colombo, A.L.; Paulsen, I.; Ren, Q.; Wortman, J.; Huang, J.; Goldman, M.H.S.; Goldman, G.H. The ergosterol biosynthesis pathway, transporter genes, and azole resistance in Aspergillus fumigatus. Med. Mycol. 2005, 43, S313–S319. [Google Scholar] [CrossRef]
- Dou, H.-T.; Xu, Y.-C.; Wang, H.-Z.; Li, T.-S. Molecular epidemiology of Cryptococcus neoformans and Cryptococcus gattii in China between 2007 and 2013 using multilocus sequence typing and the DiversiLab system. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, D.; Kuchler, K.; Ischer, F.; Pagani, J.L.; Monod, M.; Bille, J. Mechanisms of resistance to azole antifungal agents in Candida albicans isolates from AIDS patients involve specific multidrug transporters. Antimicrob. Agents Chemother. 1995, 39, 2378–2386. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.; Ribeiro, J.; Miranda, I.M.; Silva-dias, A.I.; Cavalheiro, M.; Costa-de-oliveira, S.; Rodrigues, A.G.; Teixeira, M.C. Clotrimazole drug resistance in Candida glabrata clinical isolates correlates with increased expression of the drug: H+ antiporters CgAqr1, CgTpo1_1, CgTpo3 and CgQdr2. Front. Microbiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Khandelwal, N.K.; Banerjee, A. Yeast ABC transporters in lipid trafficking. Fungal Genet. Biol. 2016, 93, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Cannon, R.D.; Holmes, A.R. Learning the ABC of oral fungal drug resistance. Mol. Oral Microbiol. 2015, 30, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Del Sorbo, G.; Schoonbeek, H.J.; De Waard, M.A. Fungal transporters involved in efflux of natural toxic compounds and fungicides. Fungal Genet. Biol. 2000, 30, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sá-Correia, I.; dos Santos, S.C.; Teixeira, M.C.; Cabrito, T.R.; Mira, N.P. Drug: H+ antiporters in chemical stress response in yeast. Trends Microbiol. 2008, 17, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Banerjee, A.; Khandelwal, N.K.; Dhamgaye, S. The ABCs of Candida albicans multidrug transporter Cdr1. Eukaryot. Cell 2015, 14, 1154–1164. [Google Scholar] [CrossRef] [PubMed]
- Pao, S.S.; Paulsen, I.T.; Saier, M.H. Major facilitator superfamily. Microbiol. Mol. Biol. Rev. 1998, 62, 1–34. [Google Scholar] [PubMed]
- Decottignies, A.; Grant, A.M.; Nichols, J.W.; de Wet, H.; McIntosh, D.B.; Goffeau, A. ATPase and multidrug transport activities of the overexpressed yeast ABC protein Yor1p. J. Biol. Chem. 1998, 273, 12612–12622. [Google Scholar] [CrossRef] [PubMed]
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.W.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.D.; Jacq, C.; Johnston, M.; et al. Life with 6000 genes. Science 1996, 274, 546–567. [Google Scholar] [CrossRef] [PubMed]
- Bauer, B.E.; Wolfger, H.; Kuchler, K. Inventory and function of yeast ABC proteins: About sex, stress, pleiotropic drug and heavy metal resistance. Biochim. Biophys. Acta 1999, 1461, 217–236. [Google Scholar] [CrossRef]
- Costa, C.; Dias, P.J.; Sá-Correia, I.; Teixeira, M.C. MFS multidrug transporters in pathogenic fungi: Do they have real clinical impact? Front. Physiol. 2014, 5, 197. [Google Scholar] [CrossRef] [PubMed]
- Franz, R.; Michel, S.; Morschhäuser, J. A fourth gene from the Candida albicans CDR family of ABC transporters. Gene 1998, 220, 91–98. [Google Scholar] [CrossRef]
- Balan, I.; Alarco, A.M.; Raymond, M. The Candida albicans CDR3 gene codes for an opaque-phase ABC transporter. J. Bacteriol. 1997, 179, 7210–7218. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, D.; Ischer, F.; Monod, M.; Bille, J. Cloning of Candida albicans genes conferring resistance to azole antifungal agents: Characterization of CDR2, a new multidrug ABC transporter gene. Microbiology 1997, 143, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.; Nunes, J.; Henriques, A.; Mira, N.P.; Nakayama, H.; Chibana, H.; Teixeira, M.C. Candida glabrata drug: H+ antiporter CgTpo3 (ORF CAGL0I10384g): Role in azole drug resistance and polyamine homeostasis. J. Antimicrob. Chemother. 2014, 69, 1767–1776. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.; Pires, C.; Cabrito, T.R.; Renaudin, A.; Ohno, M.; Chibana, H.; Sá-Correia, I.; Teixeira, M.C. Candida glabrata drug: H+ antiporter CgQdr2 confers imidazole drug resistance, being activated by transcription factor CgPdr1. Antimicrob. Agents Chemother. 2013, 57, 3159–3167. [Google Scholar] [CrossRef] [PubMed]
- Torelli, R.; Posteraro, B.; Ferrari, S.; La Sorda, M.; Fadda, G.; Sanglard, D.; Sanguinetti, M. The ATP-binding cassette transporter—Encoding gene CgSNQ2 is contributing to the CgPDR1-dependent azole resistance of Candida glabrata. Mol. Microbiol. 2008, 68, 186–201. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.; Henriques, A.; Pires, C.; Nunes, J.; Ohno, M.; Chibana, H.; Sá-Correia, I.; Teixeira, M.C. The dual role of Candida glabrata drug: H+ antiporter CgAqr1 (ORF CAGL0J09944g) in antifungal drug and acetic acid resistance. Front. Microbiol. 2013, 4, 170. [Google Scholar] [CrossRef] [PubMed]
- Tobin, M.B.; Peery, R.B.; Skatrud, P.L. Genes encoding multiple drug resistance-like proteins in Aspergillus fumigatus and Aspergillus flavus. Gene 1997, 200, 11–23. [Google Scholar] [CrossRef]
- Slaven, J.W.; Anderson, M.J.; Sanglard, D.; Dixon, G.K.; Bille, J.; Roberts, I.S.; Denning, D.W. Increased expression of a novel Aspergillus fumigatus ABC transporter gene, atrF, in the presence of itraconazole in an itraconazole resistant clinical isolate. Fungal Genet. Biol. 2002, 36, 199–206. [Google Scholar] [CrossRef]
- Thornewell, S.J.; Peery, R.B.; Skatrud, P.L. Cloning and characterization of CneMDR1: A Cryptococcus neoformans gene encoding a protein related to multidrug resistance proteins. Gene 1997, 201, 21–29. [Google Scholar] [CrossRef]
- Posteraro, B.; Sanguinetti, M.; Sanglard, D.; La Sorda, M.; Boccia, S.; Romano, L.; Morace, G.; Fadda, G.; Cassette, B. Identification and characterization of a Cryptococcus neoformans ATP binding cassette (ABC) transporter-encoding gene, CnAFR1, involved in the resistance to fluconazole. Mol. Microbiol. 2003, 47, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Gaur, M.; Choudhury, D.; Prasad, R. Complete inventory of ABC proteins in human pathogenic yeast, Candida albicans. J. Mol. Microbiol. Biotechnol. 2005, 9, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Dujon, B.; Sherman, D.; Fischer, G.; Durrens, P.; Casaregola, S.; Lafontaine, I.; de Montigny, J.; Marck, C.; Neuvéglise, C.; Talla, E.; et al. Genome evolution in yeasts. Nature 2004, 430, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Nierman, W.C.; Pain, A.; Anderson, M.J.; Wortman, J.R.; Kim, H.S.; Arroyo, J.; Berriman, M.; Abe, K.; Archer, D.B.; Bermejo, C.; et al. Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature 2005, 438, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Loftus, B.J.; Fung, E.; Roncaglia, P.; Rowley, D.; Amedeo, P.; Vamathevan, J.; Miranda, M.; Anderson, I.J.; Fraser, J.A.; Allen, J.E.; et al. The genome of the Basidiomycetous yeast and human pathogen Cryptococcus neoformans. Science 2005, 307, 1321–1324. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.R.; Cardno, T.S.; Strouse, J.J.; Ivnitski-Steele, I.; Keniya, M.V.; Lackovic, K.; Monk, B.C.; Sklar, L.A.; Cannon, R.D. Targeting efflux pumps to overcome antifungal drug resistance. Future Med. Chem. 2016, 8, 1485–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- K Redhu, A.; Shah, A.H.; Prasad, R. MFS transporters of Candida species and their role in clinical drug resistance. FEMS Yeast Res. 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Lupetti, A.; Danesi, R.; Campa, M.; Del Tacca, M.; Kelly, S. Molecular basis of resistance to azole antifungals. Trends Mol. Med. 2002, 8, 76–81. [Google Scholar] [CrossRef]
- Sanguinetti, M.; Posteraro, B.; La Sorda, M.; Torelli, R.; Fiori, B.; Santangelo, R.; Delogu, G.; Fadda, G. Role of AFR1, an ABC transporter-encoding gene, in the in vivo response to fluconazole and virulence of Cryptococcus neoformans. Infect. Immun. 2006, 74, 1352–1359. [Google Scholar] [CrossRef] [PubMed]
- Cannon, R.D.; Lamping, E.; Holmes, A.R.; Niimi, K.; Baret, P.V.; Keniya, M.V.; Tanabe, K.; Niimi, M.; Goffeau, A.; Monk, B.C. Efflux-mediated antifungal drug resistance. Clin. Microbiol. Rev. 2009, 22, 291–321. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; De Wergifosse, P.; Goffeau, A.; Balzi, E. Molecular cloning and characterization of a novel gene of Candida albicans, CDR1, conferring multiple resistance to drugs and antifungals. Curr. Genet. 1995, 27, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Siikala, E.; Rautemaa, R.; Richardson, M.; Saxen, H.; Bowyer, P.; Sanglard, D. Persistent Candida albicans colonization and molecular mechanisms of azole resistance in autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy (APECED) patients. J. Antimicrob. Chemother. 2010, 65, 2505–2513. [Google Scholar] [CrossRef] [PubMed]
- Moran, G.P.; Sanglard, D.; Donnelly, S.M.; Shanley, D.B.; Sullivan, D.J.; Coleman, D.C. Identification and expression of multidrug transporters responsible for fluconazole resistance in Candida dubliniensis. Antimicrob. Agents Chemother. 1998, 42, 1819–1830. [Google Scholar] [PubMed]
- Berkow, E.L.; Manigaba, K.; Parker, J.E.; Barker, K.S.; Kelly, S.L.; Rogers, P.D. Multidrug transporters and alterations in sterol biosynthesis contribute to azole antifungal resistance in Candida parapsilosis. Antimicrob. Agents Chemother. 2015, 59, 5942–5950. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.C.R.; Fuchs, B.B.; Pinhati, H.M.S.; Siqueira, R.A.; Hagen, F.; Meis, J.F.; Mylonakis, E.; Colombo, A.L. Candida parapsilosis resistance to fluconazole: Molecular mechanisms and in vivo impact in infected Galleria mellonella larvae. Antimicrob. Agents Chemother. 2015, 59, 6581–6587. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.J.; Won, E.J.; Shin, J.H.; Kim, S.H.; Lee, W.G.; Kim, M.N.; Lee, K.; Shin, M.G.; Suh, S.P.; Ryang, D.W.; et al. Resistance mechanisms and clinical features of fluconazole-nonsusceptible Candida tropicalis isolates compared with fluconazole-less-susceptible isolates. Antimicrob. Agents Chemother. 2016, 60, 3653–3661. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, D.; Ischer, F.; Bille, J. Role of ATP-binding-cassette transporter genes in high-frequency acquisition of resistance to azole antifungals in Candida glabrata. Antimicrob. Agents Chemother. 2001, 45, 1174–1183. [Google Scholar] [CrossRef] [PubMed]
- Vermitsky, J.-P.; Earhart, K.D.; Smith, W.L.; Homayouni, R.; Edlind, T.D.; Rogers, P.D. Pdr1 regulates multidrug resistance in Candida glabrata: Gene disruption and genome-wide expression studies. Mol. Microbiol. 2006, 61, 704–722. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K.; Edlind, T.D. Identification and expression of multidrug resistance-related ABC transporter genes in Candida krusei. Med. Mycol. 2001, 39, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Lamping, E.; Ranchod, A.; Nakamura, K.; Tyndall, J.D.A.; Niimi, K.; Holmes, A.R.; Niimi, M.; Cannon, R.D. Abc1p is a multidrug efflux transporter that tips the balance in favor of innate azole resistance in Candida krusei. Antimicrob. Agents Chemother. 2009, 53, 354–369. [Google Scholar] [CrossRef] [PubMed]
- Lamping, E.; Monk, B.C.; Niimi, K.; Holmes, A.R.; Tsao, S.; Tanabe, K.; Niimi, M.; Uehara, Y.; Cannon, R.D. Characterization of three classes of membrane proteins involved in fungal azole resistance by functional hyperexpression in Saccharomyces cerevisiae. Eukaryot. Cell 2007, 6, 1150–1165. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zhao, M.; Chen, J.; Wu, R.; Zhang, J.; Cui, R.; Jiang, Y.; Chen, J.; Cao, X.; Xing, Y.; et al. Overexpression of both ERG11 and ABC2 genes might be responsible for itraconazole resistance in clinical isolates of Candida krusei. PLoS ONE 2015, 10, e0136185. [Google Scholar] [CrossRef] [PubMed]
- Meneau, I.; Coste, A.T.; Sanglard, D. Identification of Aspergillus fumigatus multidrug transporter genes and their potential involvement in antifungal resistance. Med. Mycol. 2016, 54, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Abad, A.; Victoria Fernández-Molina, J.; Bikandi, J.; Ramírez, A.; Margareto, J.; Sendino, J.; Luis Hernando, F.; Pontón, J.; Garaizar, J.; Rementeria, A. What makes Aspergillus fumigatus a successful pathogen? Genes and molecules involved in invasive aspergillosis. Rev. Iberoam. Micol. 2010, 27, 155–182. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, A.M.; Goldman, G.H.; Park, S.; Marras, S.A.E.; Delmas, G.; Oza, U.; Lolans, K.; Dudley, M.N.; Mann, P.A.; Perlin, D.S. Multiple resistance mechanisms among Aspergillus fumigatus mutants with high-level resistance to itraconazole. Antimicrob. Agents Chemother. 2003, 47, 1719–1726. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Ferreira, M.E.; Malavazi, I.; Savoldi, M.; Brakhage, A.A.; Goldman, M.H.S.; Kim, H.S.; Nierman, W.C.; Goldman, G.H. Transcriptome analysis of Aspergillus fumigatus exposed to voriconazole. Curr. Genet. 2006, 50, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Sionov, E.; Khanal Lamichhane, A.; Kwon-Chung, K.J.; Chang, Y.C. Roles of three Cryptococcus neoformans and Cryptococcus gattii efflux pump-coding genes in response to drug treatment. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Basso, L.R.; Gast, C.E.; Bruzual, I.; Wong, B. Identification and properties of plasma membrane azole efflux pumps from the pathogenic fungi Cryptococcus gattii and Cryptococcus neoformans. J. Antimicrob. Chemother. 2015, 70, 1396–1407. [Google Scholar] [CrossRef] [PubMed]
- Wirsching, S.; Michel, S.; Morschhäuser, J. Targeted gene disruption in Candida albicans wild-type strains: The role of the MDR1 gene in fluconazole resistance of clinical Candida albicans isolates. Mol. Microbiol. 2000, 36, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Wirsching, S.; Moran, G.P.; Sullivan, D.J.; Coleman, D.C.; Morschhäuser, J. MDR1-mediated drug resistance in Candida dubliniensis. Antimicrob. Agents Chemother. 2001, 45, 3416–3421. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.P.; Miranda, I.M.; Guida, A.; Synnott, J.; Rocha, R.; Silva, R.; Amorim, A.; Pina-Vaz, C.; Butler, G.; Rodrigues, A.G. Transcriptional profiling of azole-resistant Candida parapsilosis strains. Antimicrob. Agents Chemother. 2011, 55, 3546–3556. [Google Scholar] [CrossRef] [PubMed]
- Bizerra, F.C.; Nakamura, C.V.; De Poersch, C.; Estivalet Svidzinski, T.I.; Borsato Quesada, R.M.; Goldenberg, S.; Krieger, M.A.; Yamada-Ogatta, S.F. Characteristics of biofilm formation by Candida tropicalis and antifungal resistance. FEMS Yeast Res. 2008, 8, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, D.; Bille, J.; Sanglard, D. A novel multidrug efflux transporter gene of the major facilitator superfamily from Candida albicans (FLU1) conferring resistance to fluconazole. Microbiology 2000, 146, 2743–2754. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Perlin, D.S. Establishing surrogate markers for fluconazole resistance in Candida albicans. Microb. Drug Resist. 2005, 11, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Pais, P.; Costa, C.; Pires, C.; Shimizu, K.; Chibana, H.; Teixeira, M.C. Membrane proteome-wide response to the antifungal drug clotrimazole in Candida glabrata: role of the transcription factor CgPdr1 and the drug: H+ antiporters CgTpo1_1 and CgTpo1_2. Mol. Cell. Proteom. 2016, 15, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Pais, P.; Pires, C.; Costa, C.; Okamoto, M.; Chibana, H.; Teixeira, M.C. Membrane proteomics analysis of the Candida glabrata response to 5-flucytosine: Unveiling the role and regulation of the drug efflux transporters CgFlr1 and CgFlr2. Front. Microbiol. 2016, 7, 2045. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.-F.; Krol, A.A.; Sarti, K.E.; Bennett, J.E. Candida glabrata PDR1, a transcriptional regulator of a pleiotropic drug resistance network, mediates azole resistance in clinical isolates and petite mutants. Antimicrob. Agents Chemother. 2006, 50, 1384–1392. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Schmidt, J.A.; Moye-Rowley, W.S. Regulation of the CgPdr1 transcription factor from the pathogen Candida glabrata. Eukaryot. Cell 2011, 10, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Coste, A.; Turner, V.; Ischer, F.; Morschhäuser, J.; Forche, A.; Selmecki, A.; Berman, J.; Bille, J.; Sanglard, D. A mutation in Tac1p, a transcription factor regulating CDR1 and CDR2, is coupled with loss of heterozygosity at chromosome 5 to mediate antifungal resistance in Candida albicans. Genetics 2006, 172, 2139–2156. [Google Scholar] [CrossRef] [PubMed]
- Coste, A.; Selmecki, A.; Forche, A.; Diogo, D.; Bougnoux, M.E.; D’Enfert, C.; Berman, J.; Sanglard, D. Genotypic evolution of azole resistance mechanisms in sequential Candida albicans isolates. Eukaryot. Cell 2007, 6, 1889–1904. [Google Scholar] [CrossRef] [PubMed]
- Morschhäuser, J.; Barker, K.S.; Liu, T.T.; BlaB-Warmuth, J.; Homayouni, R.; Rogers, P.D. The transcription factor Mrr1p controls expression of the MDR1 efflux pump and mediates multidrug resistance in Candida albicans. PLoS Pathog. 2007, 3, e164. [Google Scholar] [CrossRef] [PubMed]
- Dunkel, N.; Blaß, J.; Rogers, P.D.; Morschhäuser, J. Mutations in the multi-drug resistance regulator MRR1, followed by loss of heterozygosity, are the main cause of MDR1 overexpression in fluconazole-resistant Candida albicans strains. Mol. Microbiol. 2008, 69, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.T.; Znaidi, S.; Barker, K.S.; Xu, L.; Homayouni, R.; Saidane, S.; Morschhäuser, J.; Nantel, A.; Raymond, M.; Rogers, P.D. Genome-wide expression and location analyses of the Candida albicans Tac1p regulon. Eukaryot. Cell 2007, 6, 2122–2138. [Google Scholar] [CrossRef] [PubMed]
- Znaidi, S.; De Deken, X.; Weber, S.; Rigby, T.; Nantel, A.; Raymond, M. The zinc cluster transcription factor Tac1p regulates PDR16 expression in Candida albicans. Mol. Microbiol. 2007, 66, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Schubert, S.; Barker, K.S.; Znaidi, S.; Schneider, S.; Dierolf, F.; Dunkel, N.; Aïd, M.; Boucher, G.; Rogers, P.D.; Raymond, M.; et al. Regulation of efflux pump expression and drug resistance by the transcription factors Mrr1, Upc2, and Cap1 in Candida albicans. Antimicrob. Agents Chemother. 2011, 55, 2212–2223. [Google Scholar] [CrossRef] [PubMed]
- Coste, A.T.; Karababa, M.; Ischer, F.; Bille, J.; Sanglard, D. TAC1, transcriptional activator of CDR genes, is a new transcription factor involved in the regulation of Candida albicans ABC transporters CDR1 and CDR2. Eukaryot. Cell 2004, 3, 1639–1652. [Google Scholar] [CrossRef] [PubMed]
- Caudle, K.E.; Barker, K.S.; Wiederhold, N.P.; Xu, L.; Homayouni, R.; Rogers, P.D. Genomewide expression profile analysis of the Candida glabrata Pdr1 regulon. Eukaryot. Cell 2011, 10, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.-F.; Sammons, L.R.; Zhang, X.; Suffis, S.D.; Su, Q.; Myers, T.G.; Marr, K.A.; Bennett, J.E. Microarray and molecular analyses of the azole resistance mechanism in Candida glabrata oropharyngeal isolates. Antimicrob. Agents Chemother. 2010, 54, 3308–3317. [Google Scholar] [CrossRef] [PubMed]
- Selmecki, A.; Forche, A.; Berman, J. Genomic plasticity of the human fungal pathogen Candida albicans. Eukaryot. Cell 2010, 9, 991–1008. [Google Scholar] [CrossRef] [PubMed]
- Poláková, S.; Blume, C.; Zárate, J.A.; Mentel, M.; Jørck-Ramberg, D.; Stenderup, J.; Piskur, J. Formation of new chromosomes as a virulence mechanism in yeast Candida glabrata. Proc. Natl. Acad. Sci. USA 2009, 106, 2688–2693. [Google Scholar] [CrossRef] [PubMed]
- Sionov, E.; Lee, H.; Chang, Y.C.; Kwon-Chung, K.J. Cryptococcus neoformans overcomes stress of azole drugs by formation of disomy in specific multiple chromosomes. PLoS Pathog. 2010, 6, e1000848. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D.; Miura, D.; Shimizu, K.; Paul, S.; Ohba, A.; Gonoi, T.; Watanabe, A.; Kamei, K.; Shintani, T.; Moye-Rowley, W.S.; et al. A novel Zn2-Cys6 transcription factor AtrR plays a key role in an azole resistance mechanism of Aspergillus fumigatus by Co-regulating cyp51A and cdr1B expressions. PLoS Pathog. 2017, 13. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Doering, T.L.; Moye-Rowley, W.S. Cryptococcus neoformans Yap1 is required for normal fluconazole and oxidative stress resistance. Fungal Genet. Biol. 2015, 74, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lev, S.; Desmarini, D.; Chayakulkeeree, M.; Sorrell, T.C.; Djordjevic, J.T. The Crz1/Sp1 transcription factor of Cryptococcus neoformans is activated by calcineurin and regulates cell wall integrity. PLoS ONE 2012, 7, e51403. [Google Scholar] [CrossRef] [PubMed]
- Noble, S.M.; Gianetti, B.A.; Witchley, J.N. Candida albicans cell-type switching and functional plasticity in the mammalian host. Nat. Rev. Microbiol. 2017, 15, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.P.; Brown, G.D.; Netea, M.G.; Gow, N.A.R. Metabolism impacts upon Candida immunogenicity and pathogenicity at multiple levels. Trends Microbiol. 2014, 22, 614–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levitz, S.M. The Ecology of Cryptococcus neoformans and the epidemiology of cryptococcosis. Rev. Infect. Dis. 1991, 13, 1163–1169. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.C.-A.; Meyer, W.; Sorrell, T.C. Cryptococcus gattii Infections. Clin. Microbiol. Rev. 2014, 27, 980–1024. [Google Scholar] [CrossRef] [PubMed]
- Latgé, J.-P. Aspergillus fumigatus and Aspergillosis. Clin. Microbiol. Rev. 1999, 12, 310–350. [Google Scholar] [CrossRef] [PubMed]
- Polke, M.; Hube, B.; Jacobsen, I.D. Candida survival strategies. In Advances in Applied Microbiology; Elsevier Ltd.: New York, NY, USA, 2015; Volume 91, pp. 139–235. ISBN 9780128022504. [Google Scholar]
- Villar, C.C.; Dongari-Bagtzoglou, A. Immune defence mechanisms and immunoenhancement strategies in oropharyngeal candidiasis. Expert Rev. Mol. Med. 2008, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathews, M.; Jia, H.P.; Guthmiller, J.M.; Losh, G.; Graham, S.; Johnson, G.K.; Tack, B.F.; McCray, P.B. Production of β-defensin antimicrobial peptides by the oral mucosa and salivary glands. Infect. Immun. 1999, 67, 2740–2745. [Google Scholar] [PubMed]
- Wunder, D.; Dong, J.; Baev, D.; Edgerton, M. Human Salivary Histatin 5 Fungicidal action does not induce programmed cell death pathways in Candida albicans. Antimicrob. Agents Chemother. 2004, 48, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Smith, L.M. Cryptococcus—Epithelial interactions. J. Fungi 2017, 3, 53. [Google Scholar] [CrossRef] [PubMed]
- Heung, L.J. Innate immune responses to Cryptococcus. J. Fungi 2017, 3, 35. [Google Scholar] [CrossRef] [PubMed]
- Leopold Wager, C.M.; Hole, C.R.; Wozniak, K.L.; Wormley, F.L. Cryptococcus and phagocytes: Complex interactions that influence disease outcome. Front. Microbiol. 2016, 7, 105. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Gomez, D.; Dominguez-Soto, A.; Ancochea, J.; Jimenez-Heffernan, J.A.; Leal, J.A.; Corbi, A.L. Dendritic cell-specific intercellular adhesion molecule 3-grabbing nonintegrin mediates binding and internalization of Aspergillus fumigatus conidia by dendritic cells and macrophages. J. Immunol. 2018, 173, 5635–5643. [Google Scholar] [CrossRef]
- Ellett, F.; Jorgensen, J.; Frydman, G.H.; Jones, C.N.; Irimia, D. Neutrophil interactions stimulate evasive hyphal branching by Aspergillus fumigatus. PLoS Pathog. 2017, 13, e1006154. [Google Scholar] [CrossRef] [PubMed]
- O’Hanlon, D.E.; Moench, T.R.; Cone, R.A. Vaginal pH and microbicidal lactic acid when lactobacilli dominate the microbiota. PLoS ONE 2013, 8, e80074. [Google Scholar] [CrossRef] [PubMed]
- Chaudry, A.N.; Travers, P.J.; Yuenger, J.; Colletta, L.; Evans, P.; Zenilman, J.M.; Tummon, A. Analysis of vaginal acetic acid in patients undergoing treatment for bacterial vaginosis. J. Clin. Microbiol. 2004, 42, 5170–5175. [Google Scholar] [CrossRef] [PubMed]
- Fidel, P.L., Jr. Immunity in vaginal candidiasis. Curr. Opin. Infect. Dis. 2005, 18, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.X.; Li, T.; Zhang, X.; Wang, S.X.; Liu, Z.H. Lactobacillus crispatus modulates vaginal epithelial cell innate response to Candida albicans. Chin. Med. J. 2017, 130, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Sipos, G.; Kuchler, K. Fungal ATP-binding cassette (ABC) transporters in drug resistance & detoxification. Curr. Drug Targets 2006, 7, 471–481. [Google Scholar] [PubMed]
- Krishnamurthy, S.; Gupta, V.; Snehlata, P.; Prasad, R. Characterisation of human steroid hormone transport mediated by Cdr1p, a multidrug transporter of Candida albicans, belonging to the ATP binding cassette super family. FEMS Microbiol. Lett. 1998, 158, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, S.; Chatterjee, U.; Gupta, V.; Prasad, R.; Das, P.; Snehlata, P.; Hasnain, S.E.; Prasad, R. Deletion of transmembrane domain 12 of CDR1, a multidrug transporter from Candida albicans, leads to altered drug specificity: Expression of a yeast multidrug transporter in baculovirus expression system. Yeast 1998, 14, 535–550. [Google Scholar] [CrossRef]
- Bauters, T.G.M.; Dhont, M.A.; Temmerman, M.I.L.; Nelis, H.J. Prevalence of vulvovaginal candidiasis and susceptibility to fluconazole in women. Am. J. Obstet. Gynecol. 2002, 187, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Spinillo, A.; Bernuzzi, A.M.; Cevini, C.; Gulminetti, R.; Luzi, S.; De Santolo, A. The relationship of bacterial vaginosis, Candida and Trichomonas infection to symptomatic vaginitis in postmenopausal women attending a vaginitis clinic. Maturitas 1997, 27, 253–260. [Google Scholar] [CrossRef]
- Spinillo, A.; Capuzzo, E.; Nicola, S.; Baltaro, F.; Ferrari, A.; Monaco, A. The impact of oral contraception on vulvovaginal candidiasis. Contraception 1995, 51, 293–297. [Google Scholar] [CrossRef]
- Fidel, P.L.; Cutright, J.; Steele, C. Effects of reproductive hormones on experimental vaginal candidiasis. Infect. Immun. 2000, 68, 651–657. [Google Scholar] [CrossRef] [PubMed]
- White, S.; Larsen, B. Candida albicans morphogenesis is influenced by estogen. Cell. Mol. Life Sci. 1997, 53, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Yeater, K.M.; Hoyer, L.L. Cellular and molecular biology of Candida albicans estrogen response. Eukaryot. Cell 2006, 5, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Prinz, W.A. ATP-binding Cassette (ABC) transporters mediate nonvesicular, raft-modulated sterol movement from the plasma membrane to the endoplasmic reticulum. J. Biol. Chem. 2004, 279, 45226–45234. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, L.J.; Balderes, D.A.; Wharton, B.; Tinkelenberg, A.H.; Rao, G.; Sturley, S.L. Transcriptional profiling identifies two members of the ATP-binding cassette transporter superfamily required for sterol uptake in yeast. J. Biol. Chem. 2002, 277, 32466–32472. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.L.; Uhrig, J.; Vu, K.; Singapuri, A.; Dennis, M.; Gelli, A.; Thompson, G.R. Fluconazole susceptibility in Cryptococcus gattii is dependent on the ABC transporter Pdr11. Antimicrob. Agents Chemother. 2016, 60, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Nagi, M.; Tanabe, K.; Ueno, K.; Nakayama, H.; Aoyama, T.; Chibana, H.; Yamagoe, S.; Umeyama, T.; Oura, T.; Ohno, H.; et al. The Candida glabrata sterol scavenging mechanism, mediated by the ATP-binding cassette transporter Aus1p, is regulated by iron limitation. Mol. Microbiol. 2013, 88, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Bard, M.; Sturm, A.M.; Pierson, C.A.; Brown, S.; Rogers, K.M.; Nabinger, S.; Eckstein, J.; Barbuch, R.; Lees, N.D.; Howell, S.A.; et al. Sterol uptake in Candida glabrata: Rescue of sterol auxotrophic strains. Diagn. Microbiol. Infect. Dis. 2005, 52, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Hazen, K.C.; Stei, J.; Darracott, C.; Breathnach, A.; May, J.; Howell, S.A. Isolation of cholesterol-dependent Candida glabrata from clinical specimens. Diagn. Microbiol. Infect. Dis. 2005, 52, 35–37. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Q.; Tsai, H.F.; Mandal, A.; Walker, B.A.; Noble, J.A.; Fukuda, Y.; Bennett, J.E. Sterol uptake and sterol biosynthesis act coordinately to mediate antifungal resistance in Candida glabrata under azole and hypoxic stress. Mol. Med. Rep. 2018, 17, 6585–6597. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.H.; Singh, A.; Dhamgaye, S.; Chauhan, N.; Vandeputte, P.; Suneetha, K.J.; Kaur, R.; Mukherjee, P.K.; Chandra, J.; Ghannoum, M.A.; et al. Novel role of a family of major facilitator transporters in biofilm development and virulence of Candida albicans. Biochem. J. 2014, 460, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Nunes, P.A.; Tenreiro, S.; Sá-Correia, I. Resistance and adaptation to quinidine in Saccharomyces cerevisiae: role of QDR1 (YIL120w), encoding a plasma membrane transporter of the major facilitator superfamily required for multidrug resistance. Antimicrob. Agents Chemother. 2001, 45, 1528–1534. [Google Scholar] [CrossRef] [PubMed]
- Vargas, R.C.; Tenreiro, S.; Teixeira, M.C.; Fernandes, A.R.; Sá-correia, I. Saccharomyces cerevisiae multidrug transporter Qdr2p (Yil121wp): Localization and function as a quinidine resistance determinant. Antimicrob. Agents Chemother. 2004, 48, 2531–2537. [Google Scholar] [CrossRef] [PubMed]
- Tenreiro, S.; Vargas, R.C.; Teixeira, M.C.; Magnani, C.; Sá-Correia, I. The yeast multidrug transporter Qdr3 (Ybr043c): Localization and role as a determinant of resistance to quinidine, barban, cisplatin, and bleomycin. Biochem. Biophys. Res. Commun. 2005, 327, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; Costa, C.; Mil-Homens, D.; Romão, D.; de Carvalho, C.C.C.R.; Pais, P.; Mira, N.P.; Fialho, A.M.; Teixeira, M.C. The multidrug resistance transporters CgTpo1_1 and CgTpo1_2 play a role in virulence and biofilm formation in the human pathogen Candida glabrata. Cell. Microbiol. 2017, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Qi, Y.; Yan, D.; Liu, H.; Chen, X.; Liu, L. CgMED3 changes membrane sterol composition to help Candida glabrata tolerate. Appl. Environ. Microbiol. 2017, 83, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Kumar, R.; Tati, S.; Puri, S.; Edgerton, M. Candida albicans Flu1-Mediated efflux of salivary histatin 5 reduces its cytosolic concentration and fungicidal activity. Antimicrob. Agents Chemother. 2013, 57, 1832–1839. [Google Scholar] [CrossRef] [PubMed]
- Kuchler, K.; Sterne, R.E.; Thorner, J. Saccharomyces cerevisiae STE6 gene product: A novel pathway for protein export in eukaryotic cells. EMBO J. 1989, 8, 3973–3984. [Google Scholar] [CrossRef] [PubMed]
- Raymond, M.; Dignard, D.; Alarco, A.M.; Mainville, N.; Magee, B.B.; Thomas, D.Y. A Ste6p/P-glycoprotein homologue from the asexual yeast Candida albicans transports the a-factor mating pheromone in Saccharomyces cerevisiae. Mol. Microbiol. 1998, 27, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, Y.P.; Shen, W.C. A homolog of Ste6, the a-factor transporter in Saccharomyces cerevisiae, is required for mating but not for monokaryotic fruiting in Cryptococcus neoformans. Eukaryot. Cell 2005, 4, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, J.G.; Beauvais, A.; Beau, R.; McGary, K.L.; Latgé, J.P.; Rokas, A. Global transcriptome changes underlying colony growth in the opportunistic human pathogen Aspergillus fumigatus. Eukaryot. Cell 2012, 11, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Kragl, C.; Schrettl, M.; Abt, B.; Sarg, B.; Lindner, H.H.; Haas, H. EstB-mediated hydrolysis of the siderophore triacetylfusarinine C optimizes iron uptake of Aspergillus fumigatus. Eukaryot. Cell 2007, 6, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Schrettl, M.; Kim, H.S.; Eisendle, M.; Kragl, C.; Nierman, W.C.; Heinekamp, T.; Werner, E.R.; Jacobsen, I.; Illmer, P.; Yi, H.; et al. SreA-mediated iron regulation in Aspergillus fumigatus. Mol. Microbiol. 2008, 70, 27–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, G. Iron and immunity: A double-edged sword. Eur. J. Clin. Investig. 2002, 32, 70–78. [Google Scholar] [CrossRef]
- Albertsen, M.; Bellahn, I.; Krämer, R.; Waffenschmidt, S. Localization and function of the yeast multidrug transporter Tpo1p. J. Biol. Chem. 2003, 278, 12820–12825. [Google Scholar] [CrossRef] [PubMed]
- Tomitori, H.; Kashiwagi, K.; Asakawa, T.; Kakinuma, Y.; Michael, A.J.; Igarashi, K. Multiple polyamine transport systems on the vacuolar membrane in yeast. Biochem. J. 2001, 353, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.C.; Cabrito, R.; Hanif, Z.M.; Vargas, R.C.; Tenreiro, S.; Sá-Correia, I. Yeast response and tolerance to polyamine toxicity involving the drug: H+ antiporter Qdr3 and the transcription factors Yap1 and Gcn4. Microbiology 2011, 157, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Tyms, A. Polyamines and the growth of bacteria and viruses. In The Physiology of Polyamines; Bachrach, U., Heime, Y., Eds.; CRC Press: Boca Raton, FL, USA, 1989. [Google Scholar]
- Tenreiro, S.; Nunes, P.A.; Viegas, C.A.; Neves, M.S.; Teixeira, M.C.; Cabral, G.; Sá-Correia, I. AQR1 gene (ORF YNL065w) encodes a plasma membrane transporter of the major facilitator superfamily that confers resistance to short-chain monocarboxylic acids and quinidine in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2002, 292, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Velasco, I.; Tenreiro, S.; Calderon, I.L.; André, B. Saccharomyces cerevisiae Aqr1 is an internal-membrane transporter involved in excretion of amino acids. Eukaryot. Cell 2004, 3, 1492–1503. [Google Scholar] [CrossRef] [PubMed]
- Romão, D.; Cavalheiro, M.; Mil-Homens, D.; Santos, R.; Pais, P.; Costa, C.; Takahashi-Nakaguchi, A.; Fialho, A.M.; Chibana, H.; Teixeira, M.C. A New determinant of Candida glabrata virulence: The acetate exporter CgDtr1. Front. Cell. Infect. Microbiol. 2017, 7, 473. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, R.T.; Cunha, D.V.; Wang, C.; Pereira, L.; Silva, S.; Salazar, S.B.; Schröder, M.S.; Okamoto, M.; Takahashi-Nakaguchi, A.; Chibana, H.; et al. The CgHaa1-Regulon Mediates Response and Tolerance to Acetic Acid Stress in the Human Pathogen Candida glabrata. G3: Genes Genomes Genet. 2017, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Smriti; Krishnamurthy, S.; Dixit, B.L.; Gupta, C.M.; Milewski, S.; Prasad, R. ABC transporters Cdr1p, Cdr2p and Cdr3p of a human pathogen Candida albicans are general phospholipid translocators. Yeast 2002, 19, 303–318. [Google Scholar] [CrossRef]
- Fox, L.M.; Cox, D.G.; Lockridge, J.L.; Wang, X.; Chen, X.; Scharf, L.; Trott, D.L.; Ndonye, R.M.; Veerapen, N.; Besra, G.S.; et al. Recognition of lyso-phospholipids by human natural killer T lymphocytes. PLoS Biol. 2009, 7, e1000228. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.; Fox, L.; Tian, R.; Bardet, W.; Skaley, M.; Mojsilovic, D.; Gumperz, J.; Hildebrand, W. Determination of cellular lipids bound to human CD1d molecules. PLoS ONE 2009, 4, e5325. [Google Scholar] [CrossRef] [PubMed]
- De Libero, G.; Mori, L. Recognition of lipid antigens by T cells. Nat. Rev. Immunol. 2005, 5, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Joosten, L.A.B.; van der Meer, J.W.M.; Kullberg, B.-J.; van de Veerdonk, F.L. Immune defence against Candida fungal infections. Nat. Rev. Immunol. 2015, 15, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D. Innate antifungal immunity: The key role of phagocytes. Annu. Rev. Immunol. 2011, 29, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Miramón, P.; Kasper, L.; Hube, B. Thriving within the host: Candida spp. interactions with phagocytic cells. Med. Microbiol. Immunol. 2013, 202, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Seider, K.; Brunke, S.; Schild, L.; Jablonowski, N.; Wilson, D.; Majer, O.; Barz, D.; Haas, A.; Kuchler, K.; Schaller, M.; et al. the facultative intracellular pathogen Candida glabrata subverts macrophage cytokine production and phagolysosome maturation. J. Immunol. 2011, 187, 3072–3086. [Google Scholar] [CrossRef] [PubMed]
- Orsi, C.F.; Colombari, B.; Ardizzoni, A.; Peppoloni, S.; Neglia, R.; Posteraro, B.; Morace, G.; Fadda, G.; Blasi, E. The ABC transporter-encoding gene AFR1 affects the resistance of Cryptococcus neoformans to microglia-mediated antifungal activity by delaying phagosomal maturation. FEMS Yeast Res. 2009, 9, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Goulart, L.; Silva, L.K.R.E.; Chiapello, L.; Silveira, C.; Crestani, J.; Masih, D.; Vainstein, M.H. Cryptococcus neoformans and Cryptococcus gattii genes preferentially expressed during rat macrophage infection. Med. Mycol. 2010, 48, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Fontes, A.C.L.; Oliveira, D.B.; Santos, J.R.A.; Carneiro, H.C.S.; De Queiroz Ribeiro, N.; De Oliveira, L.V.N.; Barcellos, V.A.; Paixão, T.A.; Abrahão, J.S.; Resende-Stoianoff, M.A.; et al. A subdose of fluconazole alters the virulence of Cryptococcus gattii during murine cryptococcosis and modulates type I interferon expression. Med. Mycol. 2017, 55, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Diekema, D.; Moye-Rowley, W.S. Contributions of Aspergillus fumigatus ATP-binding cassette transporter proteins to drug resistance and virulence. Eukaryot. Cell 2013, 12, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- White, T.C. Increased mRNA Levels of ERG16, CDR, and MDR1 correlate with increases in azole resistance in Candida albicans isolates from a patient infected with human immunodeficiency virus. Antimicrob. Agents Chemother. 1997, 41, 1482–1487. [Google Scholar] [PubMed]
- Hiller, D.; Sanglard, D.; Morschhauser, J. Overexpression of the MDR1 gene is sufficient to confer increased resistance to toxic compounds in Candida albicans. Antimicrob. Agents Chemother. 2006, 50, 1365–1371. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.M.; Henry, L.K.; Jiang, W.; Koltin, Y. Reduced virulence of Candida albicans mutants affected in multidrug resistance. Infect. Immun. 1995, 63, 4515–4518. [Google Scholar] [PubMed]
- Yamada-Okabe, T.; Yamada-Okabe, H. Characterization of the CaNAG3, CaNAG4, and CaNAG6 genes of the pathogenic fungus Candida albicans: Possible involvement of these genes in the susceptibilities of cytotoxic agents. FEMS Microbiol. Lett. 2002, 212, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, M.; Teixeira, M.C. Candida biofilms: Threats, challenges, and promising strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, R.; Mowat, E.; McCulloch, E.; Lappin, D.F.; Jones, B.; Lang, S.; Majithiya, J.B.; Warn, P.; Williams, C.; Ramage, G. Azole resistance of Aspergillus fumigatus biofilms is partly associated with efflux pump activity. Antimicrob. Agents Chemother. 2011, 55, 2092–2097. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Sanguinetti, M.; Torelli, R.; Posteraro, B.; Sanglard, D. Contribution of CgPDR1-regulated genes in enhanced virulence of azole-resistant Candida glabrata. PLoS ONE 2011, 6, e17589. [Google Scholar] [CrossRef] [PubMed]
- Vale-Silva, L.A.; Moeckli, B.; Torelli, R.; Posteraro, B.; Sanglard, D. Upregulation of the adhesin gene EPA1 mediated by PDR1 in Candida glabrata leads to enhanced host colonization. mSphere 2016, 1, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, P.T.; Pais, P.; Costa, C.; Manna, S.; Sa-Correia, I.; Teixeira, M.C. The PathoYeastract database: An information system for the analysis of gene and genomic transcription regulation in pathogenic yeasts. Nucleic Acids Res. 2017, 45, D597–D603. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Monge, R.; Román, E.; Arana, D.M.; Prieto, D.; Urrialde, V.; Nombela, C.; Pla, J. The Sko1 protein represses the yeast-to-hypha transition and regulates the oxidative stress response in Candida albicans. Fungal Genet. Biol. 2010, 47, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; De Micheli, M.; Coleman, S.T.; Sanglard, D.; Moye-Rowley, W.S. Analysis of the oxidative stress regulation of the Candida albicans transcription factor, Cap1p. Mol. Microbiol. 2000, 36, 618–629. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.; Wilson, R.B.; Mitchell, A.P. RIM101-dependent and-independent pathways govern pH responses in Candida albicans. Mol. Cell. Biol. 2000, 20, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.; Edwars John, J.; Mitchell, A.P.; Ibrahim, A.S. Candida albicans RIM101 pH response pathway is required for host-pathogen interactions. Infect. Immun. 2000, 68, 5953–5959. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, S.; Leach, M.D.; Priest, C.L.; Brown, A.J.P. Role of the heat shock transcription factor, Hsf1, in a major fungal pathogen that is obligately associated with warm-blooded animals. Mol. Microbiol. 2009, 74, 844–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leach, M.D.; Cowen, L.E. to sense or die: mechanisms of temperature sensing in fungal pathogens. Curr. Fungal Infect. Rep. 2014, 8, 185–191. [Google Scholar] [CrossRef]
- Bruno, V.M.; Kalachikov, S.; Subaran, R.; Nobile, C.J.; Kyratsous, C.; Mitchell, A.P. Control of the C. albicans cell wall damage response by transcriptional regulator Cas5. PLoS Pathog. 2006, 2, e21. [Google Scholar] [CrossRef] [PubMed]
- Vasicek, E.M.; Berkow, E.L.; Bruno, V.M.; Mitchell, A.P.; Wiederhold, N.P.; Barker, K.S.; Rogers, P.D. Disruption of the transcriptional regulator Cas5 results in enhanced killing of Candida albicans by Fluconazole. Antimicrob. Agents Chemother. 2014, 58, 6807–6818. [Google Scholar] [CrossRef] [PubMed]
- Nobile, C.J.; Fox, E.P.; Nett, J.E.; Sorrells, T.R.; Mitrovich, Q.M.; Hernday, A.D.; Tuch, B.B.; Andes, D.R.; Johnson, A.D. A Recently evolved transcriptional network controls biofilm development in Candida albicans. Cell 2013, 148, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Chandra, J.; Kuhn, D.M.; Ghannoum, M.A. Mechanism of fluconazole resistance in Candida albicans biofilms: Phase-specific role of efflux pumps and membrane sterols. Infect. Immun. 2003, 71, 4333–4340. [Google Scholar] [CrossRef] [PubMed]
- Lepak, A.; Nett, J.; Lincoln, L.; Marchillo, K.; Andes, D. Time course of microbiologic outcome and gene expression in Candida albicans during and following in vitro and in vivo exposure to fluconazole. Antimicrob. Agents Chemother. 2006, 50, 1311–1319. [Google Scholar] [CrossRef] [PubMed]
- Fox, E.P.; Bui, C.K.; Nett, J.E.; Hartooni, N.; Mui, M.C.; Andes, D.R.; Nobile, C.J.; Johnson, A.D. An expanded regulatory network temporally controls Candida albicans biofilm formation. Mol. Microbiol. 2015, 96, 1226–1239. [Google Scholar] [CrossRef] [PubMed]
- Nobile, C.J.; Mitchell, A.P. Genetics and genomics of Candida albicans biofilm formation. Cell. Microbiol. 2006, 8, 1382–1391. [Google Scholar] [CrossRef] [PubMed]
- Lane, S.; Birse, C.; Zhou, S.; Matson, R.; Liu, H. DNA array studies demonstrate convergent regulation of virulence factors by Cph1, Cph2, and Efg1 in Candida albicans. J. Biol. Chem. 2001, 276, 48988–48996. [Google Scholar] [CrossRef] [PubMed]
- Ha, K.C.; White, T.C. Effects of azole antifungal drugs on the transition from yeast cells to hyphae in susceptible and resistant isolates of the pathogenic yeast Candida albicans. Antimicrob. Agents Chemother. 1999, 43, 763–768. [Google Scholar] [PubMed]
- Mogavero, S.; Tavanti, A.; Senesi, S.; Rogers, P.D.; Morschhäuser, J. Differential requirement of the transcription factor Mcm1 for activation of the Candida albicans multidrug efflux pump MDR1 by its regulators Mrr1 and Cap1. Antimicrob. Agents Chemother. 2011, 55, 2061–2066. [Google Scholar] [CrossRef] [PubMed]
- Znaidi, S.; Nesseir, A.; Chauvel, M.; Rossignol, T.; D’Enfert, C. A comprehensive functional portrait of two heat shock factor-type transcriptional regulators involved in Candida albicans morphogenesis and virulence. PLoS Pathog. 2013, 9, e1003519. [Google Scholar] [CrossRef] [PubMed]
- Silver, P.M.; Oliver, B.G.; White, T.C. Role of Candida albicans Transcription factor Upc2p in Drug resistance and sterol metabolism. Eukaryot. Cell 2004, 3, 1391–1397. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, H.; Hogues, H.; Mallick, J.; Sellam, A.; Nantel, A.; Whiteway, M. Evolutionary tinkering with conserved components of a transcriptional regulatory network. PLoS Biol. 2010, 8, e1000329. [Google Scholar] [CrossRef] [PubMed]
- Dogra, S.; Krishnamurthy, S.; Gupta, V.; Dixit, B.L.; Gupta, C.M.; Sanglard, D.; Prasad, R. Asymmetric distribution of phosphatidylethanolamine in C. albicans: Possible mediation by CDR1, a multidrug transporter belonging to ATP binding cassette (ABC) superfamily. Yeast 1999, 15, 111–121. [Google Scholar] [CrossRef]
- Noble, J.A.; Tsai, H.-F.; Suffis, S.D.; Su, Q.; Myers, T.G.; Bennett, J.E. STB5 is a negative regulator of azole resistance in Candida glabrata. Antimicrob. Agents Chemother. 2013, 57, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Roetzer, A.; Klopf, E.; Gratz, N.; Marcet-Houben, M.; Hiller, E.; Rupp, S.; Gabaldón, T.; Kovarik, P.; Schüller, C. Regulation of Candida glabrata oxidative stress resistance is adapted to host environment. FEBS Lett. 2011, 585, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.H.; Miyazaki, T.; Tsai, H.F.; Bennett, J.E. The bZip transcription factor Cgap1p is involved in multidrug resistance and required for activation of multidrug transporter gene CgFLR1 in Candida glabrata. Gene 2007, 386, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Merhej, J.; Thiebaut, A.; Blugeon, C.; Pouch, J.; Ali Chaouche, M.E.A.; Camadro, J.-M.; Le Crom, S.; Lelandais, G.; Devaux, F. A network of paralogous stress response transcription factors in the human pathogen Candida glabrata. Front. Microbiol. 2016, 7, 645. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Total # | Characterised # | Characterised ORFs | Role in MDR | Physiological Role | Pathogenicity and Virulence Features | |
|---|---|---|---|---|---|---|---|
| ABC Proteins | Candida albicans | 28 | 4 | orf19.6000/CDR1 | Multidrug transporter of ABC superfamily | Transport of phospholipids (in-to-out direction), steroids | Induced by β-estradiol, progesterone, corticosteroid, or cholesterol; Spider biofilm induced |
| orf19.5958/CDR2 | Multidrug transporter of ABC superfamily, overexpressed in azole-resistant isolates | Transports phospholipids (in-to-out direction) | Repressed in young biofilms | ||||
| orf19.1313/CDR3 | - | Transporter of the Pdr/Cdr family of the ATP-binding cassette superfamily; transports phospholipids (out-to-in direction); expressed in opaque-phase cells | Induced by macrophage interaction; Spider biofilm induced | ||||
| orf19.5079/CDR4 | - | - | Rat catheter and flow model biofilm induced | ||||
| Candida glabrata | 18 | 5 | CAGL0M01760g/CDR1 | Multidrug transporter of ABC superfamily, involved in resistance to azoles, expression regulated by Pdr1p, increased abundance in azole resistant strains | Expression increased by loss of the mitochondrial genome | - | |
| CAGL0F02717g/CDR2 (PDH1) | Multidrug transporter, predicted plasma membrane ATP-binding cassette (ABC) transporter; regulated by Pdr1p; involved in fluconazole resistance | - | - | ||||
| CAGL0I04862g /SNQ2 | Predicted plasma membrane ATP-binding cassette (ABC) transporter, putative transporter involved in multidrug resistance; involved in Pdr1p-mediated azole resistance | - | - | ||||
| CAGL0G00242g/YOR1 | Putative ABC transporter involved in multidrug efflux; gene is upregulated in azole-resistant strain | - | - | ||||
| CAGL0F01419g/AUS1 | - | ATP-binding cassette transporter involved in sterol uptake | Necessary for C. glabrata virulence in a mice model of disseminated infection | ||||
| Candida tropicalis | 22 | 1 | CDR1 | Induced in clinical azole resistant isolates | - | - | |
| Candida parapsilosis | 19 | 1 | CPAR2_405290/CDR1 | Induced in clinical azole resistant isolates | - | - | |
| Candida dubliniensis | 19 | 1 | Cd36_85210/CDR1 | Predicted multidrug transporter of ABC superfamily, involved in multidrug resistance, overexpressed in fluconazole-resistant derivatives obtained in vitro | - | - | |
| Candida krusei | 9 | 2 | ABC1 | Upregulated during azole stress, involved in innate fluconazole resistance, confers fluconazole resistance through drug efflux upon hyperexpression in S. cerevisiae | - | - | |
| ABC2 | Upregulated during azole stress, expression correlated with itraconazole resistance | - | - | ||||
| Aspergillus fumigatus | 49 | 7 | Afu6g04360/ATRF | Putative ABC transporter; drug efflux pump; involved in itraconazole resistance | - | - | |
| Afu1g12690/MDR4 | ABC multidrug transporter; induced by voriconazole exposure in vitro and in mice; involved in itraconazole resistance | - | Biofilm growth regulated protein | ||||
| Afu1g17440/ABCA | ABC drug exporter; induced during voriconazole stress | - | Overexpression leads to an augmentation of virulence in the presence of voriconazole in the G. mellonella model of infection | ||||
| Afu1g10390/ABCB | Putative ABC multidrug transporter; transcript induced by voriconazole | - | Necessary for full virulence of A. fumigatus in the G. mellonella model of infection | ||||
| Afu1g14330/ABCC | Putative ABC transporter; induced during voriconazole stress; mutation causes increased itraconazole, voriconazole and posaconazole sensitivity | - | - | ||||
| Afu6g03470/ABCD | Putative MDR1 family ABC transporter; induced during voriconazole stress | - | Biofilm growth regulated protein | ||||
| Afu7g00480/ABCE | Putative ABC transporter; induced during voriconazole stress | - | Biofilm growth regulated protein | ||||
| Afu3g07300/ATRI | Putative ABC transporter; induced during voriconazole stressmutation causes increased itraconazole and voriconazole sensitivity | - | - | ||||
| Cryptococcus neoformans | 54 | 3 | AFR1 | Pump required for azole efflux and other xenobiotics, including cycloheximide, nocodazole, and trichostatin A; involved in clinical fluconazole resistance; role in susceptibility towards amphotericin B and 5-fluorocytosine: expression was positively regulated by CnCrz1 and CnYap1 in response to fluconazole | - | Necessary for full virulence of C. neoformans in intravenous and in inhalation mouse models; overexpression upon C. neoformans phagocytosis; and involved in the resistance against microglia | |
| AFR2 | Role in susceptibility towards amphotericin B and 5-fluorocytosine | - | - | ||||
| MDR1 | Confers itraconazole resistance upon hyperexpression in S. cerevisiae; role in susceptibility towards amphotericin B and 5-fluorocytosine | - | - | ||||
| Cryptococcus gattii | 23 | 2 | MDR1 | Confers fluconazole resistance in S. cerevisiae | - | Overexpressed in cells recovered from the brain of infected mice | |
| PDR11 | Necessary for fluconazole resistance in the VGII clinical strain | - | Overexpressed in cells recovered from the lungs of infected mice |
| Species | Total # | Characterised # | Characterised ORFs | Role in MDR | Physiological Role | Pathogenicity and Virulence Features | |
|---|---|---|---|---|---|---|---|
| MFS Proteins | Candida albicans | 26 | 8 | orf19.5604/MDR1 | Major mediator of azole resistance; methotrexate is preferred substrate; overexpression in drug-resistant clinical isolates confers fluconazole resistance; repressed in young biofilm | - | Necessary for full virulence in C. albicans in immunocompetent and immunocompromised mice models |
| orf19.6577/FLU1 | Involved in the resistance towards fluconazole, ketoconazole, and itraconazole; confers fluconazole resistance in S. cerevisiae | Involved in histatin-5 efflux | - | ||||
| orf19.2160/NAG4 | Required for wild-type cycloheximide resistance | - | Required for wild-type mouse virulence | ||||
| orf19.2158/NAG3 | Required for wild-type cycloheximide resistance | - | Required for wild-type mouse virulence; Spider biofilm repressed | ||||
| orf19.2151/NAG6 | Required for wild-type cycloheximide resistance | - | Required for wild-type mouse virulence | ||||
| orf19.508/QDR1 | - | Involved in lipid homeostasis | Involved in biofilm architecture and thickness and virulence | ||||
| orf19.6992/QDR2 | - | Involved in lipid homeostasis | Involved in biofilm architecture and thickness and virulence in a murine model of hematogenously disseminated candidiasis | ||||
| orf19.136/QDR3 | - | Involved in lipid homeostasis | Involved in biofilm architecture and thickness and virulence in a murine model of hematogenously disseminated candidiasis | ||||
| Candida glabrata | 15 | 8 | CAGL0J09944g/AQR1 | Involved in resistance to flucytosine and imidazoles | Involved in resistance to acetic acid | - | |
| CAGL0G03927g/TPO1_1 | Putative drug:H+ antiporter, involved in efflux of clotrimazole; required for resistance to clotrimazole and other drugs | Involved in the resistance to histatin-5; involved in spermine resistance | Involved in virulence | ||||
| CAGL0E03674g/TPO1_2 | Putative drug:H+ antiporter, involved in efflux of clotrimazole; required for resistance to clotrimazole and other drugs | Involved in fatty acid and sterol homeostasis upon biofilm formation; involved in spermine resistance | Involved in virulence in the G. mellonella model and biofilm formation | ||||
| CAGL0I10384g/TPO3 | Confers imidazole and triazole drug resistance; activated by CgPdr1 | Involved in polyamine homeostasis | - | ||||
| CAGL0G08624g/QDR2 | Confers imidazole drug resistance, involved in clotrimazole efflux; activated by CgPdr1; upregulated in azole-resistant strain | - | - | ||||
| CAGL0H06017g/FLR1 | Confers resistance to benomyl; gene is downregulated in azole-resistant strain | - | - | ||||
| CAGL0H06039g/FLR2 | Multidrug transporter of the major facilitator superfamily involved in 5-flucytosine resistance | - | - | ||||
| CAGL0M06281g/DTR1 | - | Acetate exporter in the plasma membrane | Required for virulence in G. mellonella model | ||||
| Candida tropicalis | 26 | 1 | MDR1 | Overexpression in resistant clinical isolates and upon biofilm formation | - | - | |
| Candida parapsilosis | 34 | 1 | CPAR2_301760/MDR1 | Member of the MDR family of major facilitator transporter superfamily; putative drug transporter; expression increased in fluconazole and voriconazole resistant strains | - | - | |
| Candida dubliniensis | 14 | 1 | Cd36_63890/MDR1 | Predicted multidrug transporter of ABC superfamily, involved in multidrug resistance | - | - | |
| Aspergillus fumigatus | 278 | 6 | Afu6g09710/GLIA | - | Predicted major facilitator type glioxin transporter, encoded in the putative gliotoxin biosynthetic gene cluster | ||
| Afu1g05010/MFS56 | Putative MFS transporter; mutation causes increased azole sensitivity | - | - | ||||
| Afu3g03500/MDR3 | Putative multidrug resistance protein; transcript upregulated in response to amphotericin B; displays itraconazole-increased expression in resistant mutants | - | - | ||||
| Afu8g05710/MFSA | Highly expressed during voriconazole stress | Putative major facilitator superfamily (MFS) sugar transporter | Calcium induced; transcript upregulated in conidia exposed to neutrophils | ||||
| Afu1g15490/MFSB | Putative major facilitator superfamily (MFS) transporter; highly expressed during voriconazole stress | - | - | ||||
| Afu1g03200/MFSC | Putative major facilitator superfamily (MFS) transporter; highly expressed during voriconazole stress | - | - | ||||
| Cryptococcus neoformans | 86 | 3 | CNA07070 | - | Dityrosine transporter | - | |
| CNC03290 | - | - | - | ||||
| CND00440/AFLT | - | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavalheiro, M.; Pais, P.; Galocha, M.; Teixeira, M.C. Host-Pathogen Interactions Mediated by MDR Transporters in Fungi: As Pleiotropic as it Gets! Genes 2018, 9, 332. https://doi.org/10.3390/genes9070332
Cavalheiro M, Pais P, Galocha M, Teixeira MC. Host-Pathogen Interactions Mediated by MDR Transporters in Fungi: As Pleiotropic as it Gets! Genes. 2018; 9(7):332. https://doi.org/10.3390/genes9070332
Chicago/Turabian StyleCavalheiro, Mafalda, Pedro Pais, Mónica Galocha, and Miguel C. Teixeira. 2018. "Host-Pathogen Interactions Mediated by MDR Transporters in Fungi: As Pleiotropic as it Gets!" Genes 9, no. 7: 332. https://doi.org/10.3390/genes9070332
APA StyleCavalheiro, M., Pais, P., Galocha, M., & Teixeira, M. C. (2018). Host-Pathogen Interactions Mediated by MDR Transporters in Fungi: As Pleiotropic as it Gets! Genes, 9(7), 332. https://doi.org/10.3390/genes9070332