Strengthening the One Health Agenda: The Role of Molecular Epidemiology in Aspergillus Threat Management



{kind=link}
{kind=link}
Abstract
:1. The Genus Aspergillus
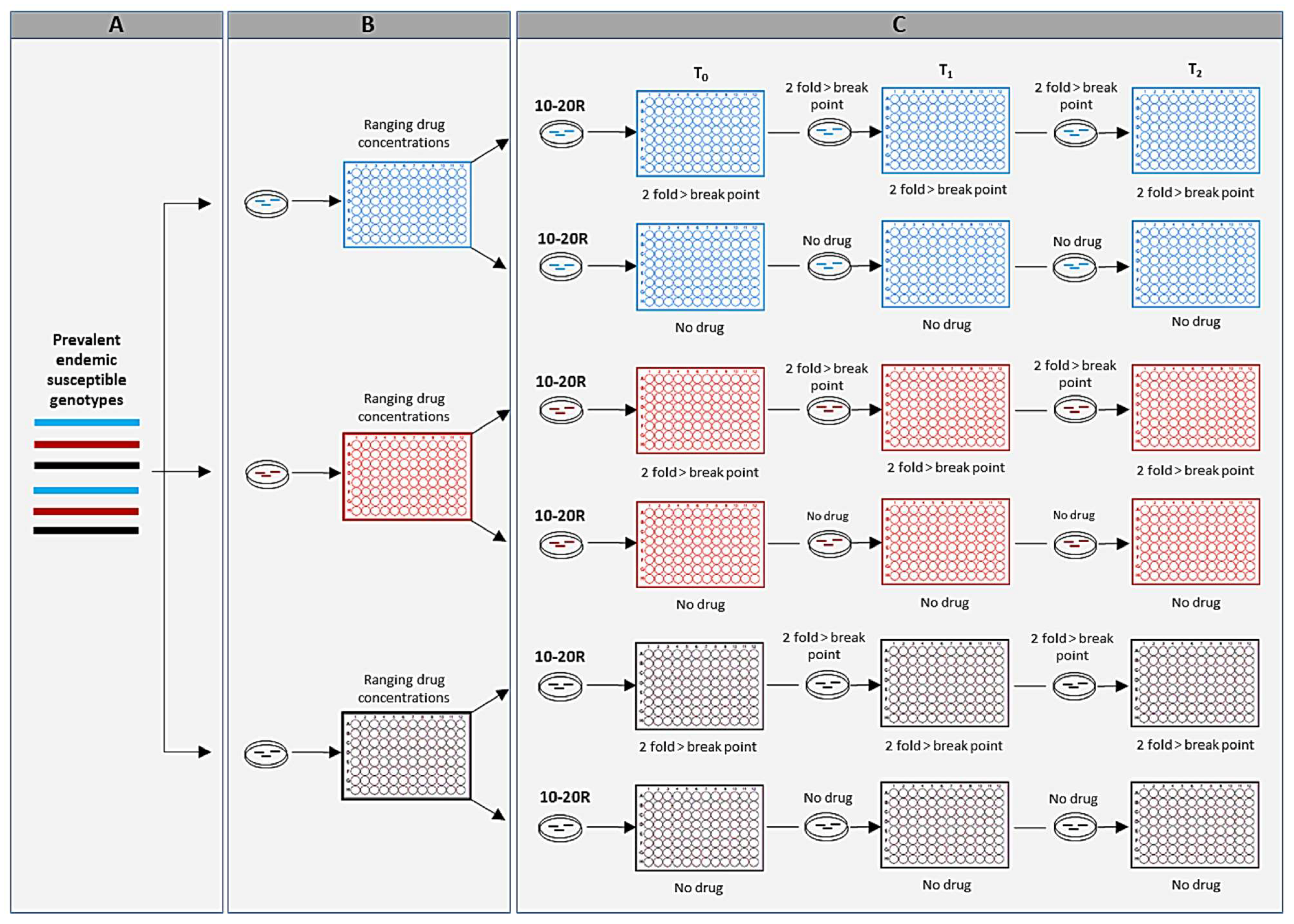
2. Molecular Epidemiology in Aspergillus Threat Management
2.1. The Usefulness of Molecular Epidemiology in Aspergillus Surveillance
2.2. The Usefulness of Molecular Epidemiology in Threat Preparedness
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and multi-national prevalence of fungal diseases—Estimate precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Vallabhaneni, S.; Mody, R.K.; Walker, T.; Chiller, T. The global burden of fungal diseases. Infect. Dis. Clin. 2016, 30, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pegorie, M.; Denning, D.W.; Welfare, W. Estimating the burden of invasive and serious fungal disease in the United Kingdom. J. Infect. 2017, 74, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Gamaletsou, M.N.; Drogari-Apiranthitou, M.; Denning, D.W.; Sipsas, N.V. An estimate of the burden of serious fungal diseases in Greece. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Gangneux, J.-P.; Bougnoux, M.-E.; Hennequin, C.; Godet, C.; Chandenier, J.; Denning, D.W.; Dupont, B. LIFE program, the Société française de mycologie médicale SFMM-study group. An estimation of burden of serious fungal infections in France. J. Mycol. Med. 2016, 26, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Oladele, R.O.; Denning, D.W. Burden of serious fungal infection in Nigeria. West Afr. J. Med. 2014, 33, 107–114. [Google Scholar] [PubMed]
- Ben, R.; Denning, D.W. Estimating the burden of fungal diseases in Israel. Isr. Med. Assoc. J. IMAJ 2015, 17, 374–379. [Google Scholar] [PubMed]
- Taj-Aldeen, S.J.; Chandra, P.; Denning, D.W. Burden of fungal infections in Qatar. Mycoses 2015, 58 (Suppl. 5), 51–57. [Google Scholar] [CrossRef] [PubMed]
- Lagrou, K.; Maertens, J.; Van Even, E.; Denning, D.W. Burden of serious fungal infections in Belgium. Mycoses 2015, 58 (Suppl. 5), 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osmanov, A.; Denning, D.W. Burden of serious fungal infections in Ukraine. Mycoses 2015, 58 (Suppl. 5), 94–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortensen, K.L.; Denning, D.W.; Arendrup, M.C. The burden of fungal disease in Denmark. Mycoses 2015, 58 (Suppl. 5), 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khwakhali, U.S.; Denning, D.W. Burden of serious fungal infections in Nepal. Mycoses 2015, 58 (Suppl. 5), 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badiane, A.S.; Ndiaye, D.; Denning, D.W. Burden of fungal infections in Senegal. Mycoses 2015, 58 (Suppl. 5), 63–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimko, N.; Kozlova, Y.; Khostelidi, S.; Shadrivova, O.; Borzova, Y.; Burygina, E.; Vasilieva, N.; Denning, D.W. The burden of serious fungal diseases in Russia. Mycoses 2015, 58 (Suppl. 5), 58–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faini, D.; Maokola, W.; Furrer, H.; Hatz, C.; Battegay, M.; Tanner, M.; Denning, D.W.; Letang, E. Burden of serious fungal infections in Tanzania. Mycoses 2015, 58 (Suppl. 5), 70–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinkó, J.; Sulyok, M.; Denning, D.W. Burden of serious fungal diseases in Hungary. Mycoses 2015, 58 (Suppl. 5), 29–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandengue, C.E.; Denning, D.W. The burden of serious fungal infections in Cameroon. J. Fungi 2018, 4, 44. [Google Scholar] [CrossRef] [PubMed]
- Guto, J.A.; Bii, C.C.; Denning, D.W. Estimated burden of fungal infections in Kenya. J. Infect. Dev. Ctries. 2016, 10, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Corzo-León, D.E.; Armstrong-James, D.; Denning, D.W. Burden of serious fungal infections in Mexico. Mycoses 2015, 58 (Suppl. 5), 34–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George Agrios. Plant Pathology, 5th ed.; Academic Press: New York, NY, USA, 2005; ISBN 978-0-12-044565-3. [Google Scholar]
- Bennett, J.W. An overview of the genus Aspergillus. In Aspergillus: Molecular Biology and Genomics; Katsuya, G., Masayuki, M., Eds.; Horizon Scientific Press: Norfolk, UK, 2010; pp. 1–17. [Google Scholar]
- Ashu, E.; Forsythe, A.; Vogan, A.; Xu, J. Filamentous Fungi in Fermented Foods. In Fermented Foods, Part I: Biochemistry and Biotechnology; Didier, M., Ramesh, R., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 60–90. ISBN 978-1-4987-4081-4. [Google Scholar]
- Dyer, P.S.; O’Gorman, C.M. A fungal sexual revolution: Aspergillus and Penicillium show the way. Curr. Opin. Microbiol. 2011, 14, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Geiser, D.M.; Klich, M.A.; Frisvad, J.C.; Peterson, S.W.; Varga, J.; Samson, R.A. The current status of species recognition and identification in Aspergillus. Stud. Mycol. 2007, 59, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sugui, J.A.; Kwon-Chung, K.J.; Juvvadi, P.R.; Latgé, J.-P.; Steinbach, W.J. Aspergillus fumigatus and related species. Cold Spring Harb. Perspect. Med. 2015, 5, a019786. [Google Scholar] [CrossRef] [PubMed]
- Guarro, J.; Xavier, M.O.; Severo, L.C. Differences and similarities amongst pathogenic Aspergillus species. In Aspergillosis: From Diagnosis to Prevention; Alessandro, C.P., Ed.; Springer: Dordrecht, The Netherlands, 2009; pp. 7–32. ISBN 978-90-481-2407-7. [Google Scholar]
- Varga, J.; Houbraken, J.; Van Der Lee, H.A.L.; Verweij, P.E.; Samson, R.A. Aspergillus calidoustus sp. nov., causative agent of human infections previously assigned to Aspergillus ustus. Eukaryot. Cell 2008, 7, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Balajee, S.A.; Gribskov, J.L.; Hanley, E.; Nickle, D.; Marr, K.A. Aspergillus lentulus sp. nov., a new sibling species of A. fumigatus. Eukaryot. Cell 2005, 4, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Gautier, M.; Normand, A.-C.; Ranque, S. Previously unknown species of Aspergillus. Clin. Microbiol. Infect. 2016, 22, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Alastruey-Izquierdo, A.; Alcazar-Fuoli, L.; Cuenca-Estrella, M. Antifungal susceptibility profile of cryptic species of Aspergillus. Mycopathologia 2014, 178, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Balajee, S.A.; Kano, R.; Baddley, J.W.; Moser, S.A.; Marr, K.A.; Alexander, B.D.; Andes, D.; Kontoyiannis, D.P.; Perrone, G.; Peterson, S.; et al. Molecular identification of Aspergillus species collected for the transplant-associated infection surveillance network. J. Clin. Microbiol. 2009, 47, 3138–3141. [Google Scholar] [CrossRef] [PubMed]
- Alastruey-Izquierdo, A.; Mellado, E.; Peláez, T.; Pemán, J.; Zapico, S.; Alvarez, M.; Rodríguez-Tudela, J.L.; Cuenca-Estrella, M.; Group, F.S. Population-based survey of filamentous fungi and antifungal resistance in Spain (FILPOP Study). Antimicrob. Agents Chemother. 2013, 57, 3380–3387. [Google Scholar] [CrossRef] [PubMed]
- Negri, C.E.; Gonçalves, S.S.; Xafranski, H.; Bergamasco, M.D.; Aquino, V.R.; Castro, P.T.O.; Colombo, A.L. Cryptic and rare Aspergillus species in Brazil: Prevalence in clinical samples and in vitro susceptibility to triazoles. J. Clin. Microbiol. 2014, 52, 3633–3640. [Google Scholar] [CrossRef] [PubMed]
- Maturu, V.N.; Agarwal, R. Itraconazole in chronic pulmonary aspergillosis: In whom, for how long, and at what dose? Lung India Off. Organ Indian Chest Soc. 2015, 32, 309–312. [Google Scholar] [CrossRef]
- Agarwal, R. Allergic bronchopulmonary aspergillosis. Chest 2009, 135, 805–826. [Google Scholar] [CrossRef] [PubMed]
- Dagenais, T.R.T.; Keller, N.P. Pathogenesis of Aspergillus fumigatus in invasive aspergillosis. Clin. Microbiol. Rev. 2009, 22, 447–465. [Google Scholar] [CrossRef] [PubMed]
- Vonberg, R.-P.; Gastmeier, P. Nosocomial aspergillosis in outbreak settings. J. Hosp. Infect. 2006, 63, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.J.; Peppercorn, A.; Miller, M.B.; Sickbert-Benett, E.; Rutala, W.A. Preventing healthcare-associated Aspergillus infections: Review of recent CDC/HICPAC recommendations. Med. Mycol. 2009, 47, S199–S209. [Google Scholar] [CrossRef] [PubMed]
- Balajee, S.A.; Tay, S.T.; Lasker, B.A.; Hurst, S.F.; Rooney, A.P. Characterization of a novel gene for strain typing reveals substructuring of Aspergillus fumigatus across North America. Eukaryot. Cell 2007, 6, 1392–1399. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Cheng, A.C.; Devitt, B.; Hughes, A.J.; Campbell, P.; Styles, K.; Low, J.; Athan, E. Successful control of an outbreak of invasive aspergillosis in a regional haematology unit during hospital construction works. J. Hosp. Infect. 2008, 69, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Guinea, J.; García de Viedma, D.; Peláez, T.; Escribano, P.; Muñoz, P.; Meis, J.F.; Klaassen, C.H.W.; Bouza, E. Molecular epidemiology of Aspergillus fumigatus: An in-depth genotypic analysis of isolates involved in an outbreak of invasive aspergillosis. J. Clin. Microbiol. 2011, 49, 3498–3503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peláez, T.; Muñoz, P.; Guinea, J.; Valerio, M.; Giannella, M.; Klaassen, C.H.W.; Bouza, E. Outbreak of invasive aspergillosis after major heart surgery caused by spores in the air of the intensive care unit. Clin. Infect. Dis. 2012, 54, e24–e31. [Google Scholar] [CrossRef] [PubMed]
- Pettit, A.C.; Kropski, J.A.; Castilho, J.L.; Schmitz, J.E.; Rauch, C.A.; Mobley, B.C.; Wang, X.J.; Spires, S.S.; Pugh, M.E. The index case for the fungal meningitis outbreak in the United States. N. Engl. J. Med. 2012, 367, 2119–2125. [Google Scholar] [CrossRef] [PubMed]
- Vena, A.; Muñoz, P.; Pelaez, T.; Guinea, J.; Valerio, M.; Bouza, E. Non-construction related Aspergillus outbreak in non-hematological patients related to high concentrations of airborne spores in non-HEPA filtered areas. Open Forum Infect. Dis. 2015, 2, 352. [Google Scholar] [CrossRef]
- Kabbani, D.; Goldraich, L.; Ross, H.; Rotstein, C.; Husain, S. Outbreak of invasive aspergillosis in heart transplant recipients: The role of screening computed tomography scans in asymptomatic patients and universal antifungal prophylaxis. Transpl. Infect. Dis. 2017, 20, e12808. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Menendez, O.; Alastruey-Izquierdo, A.; Mellado, E.; Cuenca-Estrella, M. Triazole resistance in Aspergillus spp.: A worldwide problem? J. Fungi 2016, 2, 21. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rubio, R.; Cuenca-Estrella, M.; Mellado, E. Triazole resistance in Aspergillus species: An emerging problem. Drugs 2017, 77, 599–613. [Google Scholar] [CrossRef] [PubMed]
- Reichert-Lima, F.; Lyra, L.; Pontes, L.; Moretti, M.L.; Pham, C.D.; Lockhart, S.R.; Schreiber, A.Z. Surveillance for azoles resistance in Aspergillus spp. highlights a high number of amphotericin B-resistant isolates. Mycoses 2018, 61. [Google Scholar] [CrossRef] [PubMed]
- Ashu, E.; Korfanty, G.; Samarasinghe, H.; Pum, N.; Man, Y.; Yamamura, D.; Xu, J. Widespread presence of amphotericin B resistant Aspergillus fumigatus in Hamilton, Canada. Infect. Drug Resist. 2018. under review. [Google Scholar]
- Paul, R.A.; Rudramurthy, S.M.; Meis, J.F.; Mouton, J.W.; Chakrabarti, A. A novel Y319H substitution in CYP51C associated with azole resistance in Aspergillus flavus. Antimicrob. Agents Chemother. 2015, 59, 6615–6619. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Sun, Y.; Chen, W.; Liu, W.; Wan, Z.; Bu, D.; Li, R. The T788G mutation in the CYP51C gene confers voriconazole resistance in Aspergillus flavus causing aspergillosis. Antimicrob. Agents Chemother. 2012, 56, 2598–2603. [Google Scholar] [CrossRef] [PubMed]
- Sharma, C.; Kumar, R.; Kumar, N.; Masih, A.; Gupta, D.; Chowdhary, A. Investigation of multiple resistance mechanisms in voriconazole-resistant Aspergillus flavus clinical isolates from a chest hospital surveillance in Delhi, India. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Nami, S.; Baradaran, B.; Mansoori, B.; Kordbacheh, P.; Rezaie, S.; Falahati, M.; Mohamed Khosroshahi, L.; Safara, M.; Zaini, F. The utilization of RNA silencing technology to mitigate the voriconazole resistance of Aspergillus Flavus; lipofectamine-based delivery. Adv. Pharm. Bull. 2017, 7, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Candida auris: A rapidly emerging cause of hospital-acquired multidrug-resistant fungal infections globally. PLoS Pathog. 2017, 13, e1006290. [Google Scholar] [CrossRef] [PubMed]
- Sarma, S.; Upadhyay, S. Current perspective on emergence, diagnosis and drug resistance in Candida auris. Infect. Drug Resist. 2017, 10, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Dettori, M.; Piana, A.; Deriu, M.G.; Lo Curto, P.; Cossu, A.; Musumeci, R.; Cocuzza, C.; Astone, V.; Contu, M.A.; Sotgiu, G. Outbreak of multidrug-resistant Acinetobacter baumannii in an intensive care unit. New Microbiol. 2014, 37, 185–191. [Google Scholar] [PubMed]
- Ghaith, D.M.; Zafer, M.M.; Al-Agamy, M.H.; Alyamani, E.J.; Booq, R.Y.; Almoazzamy, O. The emergence of a novel sequence type of MDR Acinetobacter baumannii from the intensive care unit of an Egyptian tertiary care hospital. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 34. [Google Scholar] [CrossRef] [PubMed]
- Zarrilli, R.; Casillo, R.; Di Popolo, A.; Tripodi, M.-F.; Bagattini, M.; Cuccurullo, S.; Crivaro, V.; Ragone, E.; Mattei, A.; Galdieri, N.; et al. Molecular epidemiology of a clonal outbreak of multidrug-resistant Acinetobacter baumannii in a university hospital in Italy. Clin. Microbiol. Infect. 2007, 13, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Eybpoosh, S.; Haghdoost, A.A.; Mostafavi, E.; Bahrampour, A.; Azadmanesh, K.; Zolala, F. Molecular epidemiology of infectious diseases. Electron. Phys. 2017, 9, 5149–5158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villari, P.; Iacuzio, L.; Torre, I.; Scarcella, A. Molecular epidemiology as an effective tool in the surveillance of infections in the neonatal intensive care unit. J. Infect. 1998, 37, 274–281. [Google Scholar] [CrossRef]
- De Valk, H.A.; Klaassen, C.H.W.; Meis, J.F.G.M. Molecular typing of Aspergillus species. Mycoses 2008, 51, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Varga, J. Molecular typing of Aspergilli: Recent developments and outcomes. Med. Mycol. 2006, 44, 149–161. [Google Scholar] [CrossRef]
- Ashu, E.E.; Xu, J. The roles of sexual and asexual reproduction in the origin and dissemination of strains causing fungal infectious disease outbreaks. Infect. Genet. Evol. 2015, 36, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Lavergne, R.-A.; Chouaki, T.; Hagen, F.; Toublanc, B.; Dupont, H.; Jounieaux, V.; Meis, J.F.; Morio, F.; Le Pape, P. Home environment as a source of life-threatening azole-resistant Aspergillus fumigatus in immunocompromised patients. Clin. Infect. Dis. 2017, 64, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Ashu, E.E.; Hagen, F.; Chowdhary, A.; Meis, J.F.; Xu, J. Global population genetic analysis of Aspergillus fumigatus. MSphere 2017, 2, e00019-17. [Google Scholar] [CrossRef] [PubMed]
- Ashu, E.E.; Korfanty, G.A.; Xu, J. Evidence of unique genetic diversity in Aspergillus fumigatus isolates from Cameroon. Mycoses 2017, 60, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Perez, S.; Blanco, J.L.; Alba, P.; Garcia, M.E. Mating type and invasiveness are significantly associated in Aspergillus fumigatus. Med. Mycol. 2010, 48, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Cheema, M.S.; Christians, J.K. Virulence in an insect model differs between mating types in Aspergillus fumigatus. Med. Mycol. 2011, 49, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.C.; Garcia-Rubio, R.; Alcazar-Fuoli, L.; Peláez, T.; Mellado, E. Could the determination of Aspergillus fumigatus mating type have prognostic value in invasive aspergillosis? Mycoses 2018, 61, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Bain, J.M.; Tavanti, A.; Davidson, A.D.; Jacobsen, M.D.; Shaw, D.; Gow, N.A.; Odds, F.C. Multilocus sequence typing of the pathogenic fungus Aspergillus fumigatus. J. Clin. Microbiol. 2007, 45, 1469–1477. [Google Scholar] [CrossRef] [PubMed]
- De Valk, H.A.; Meis, J.F.G.M.; Curfs, I.M.; Muehlethaler, K.; Mouton, J.W.; Klaassen, C.H.W. Use of a novel panel of nine short tandem repeats for exact and high-resolution fingerprinting of Aspergillus fumigatus isolates. J. Clin. Microbiol. 2005, 43, 4112–4120. [Google Scholar] [CrossRef] [PubMed]
- García-Lerma, J.G.; Nidtha, S.; Blumoff, K.; Weinstock, H.; Heneine, W. Increased ability for selection of zidovudine resistance in a distinct class of wild-type HIV-1 from drug-naive persons. Proc. Natl. Acad. Sci. USA 2001, 98, 13907–13912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdolrasouli, A.; Rhodes, J.; Beale, M.A.; Hagen, F.; Rogers, T.R.; Chowdhary, A.; Meis, J.F.; Armstrong-James, D.; Fisher, M.C. Genomic context of azole resistance mutations in Aspergillus fumigatus determined using whole-genome sequencing. MBio 2015, 6, e00536-15. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Ashu, E.; Sharma, C.; Kathuria, S.; Chowdhary, A.; Xu, J. Diversity and origins of Indian multi-triazole resistant strains of Aspergillus fumigatus. Mycoses 2016, 59, 450–466. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 2nd ed.; Approved Standard, CLSI M38-A2; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- Dunne, K.; Hagen, F.; Pomeroy, N.; Meis, J.F.; Rogers, T.R. Intercountry transfer of triazole-resistant Aspergillus fumigatus on Plant Bulbs. Clin. Infect. Dis. 2017, 65, 147–149. [Google Scholar] [CrossRef] [PubMed]
- Lamoth, F. Aspergillus fumigatus-related species in clinical practice. Front. Microbiol. 2016, 7, 683. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Tian, S.; Yu, N.; Zhang, X.; Jia, X.; Zhai, H.; Sun, Q.; Han, L. Development and evaluation of a loop-mediated isothermal amplification method for rapid detection of Aspergillus fumigatus. J. Clin. Microbiol. 2016, 54, 950–955. [Google Scholar] [CrossRef] [PubMed]
- Powers-Fletcher, M.V.; Hanson, K.E. Molecular diagnostic testing for Aspergillus. J. Clin. Microbiol. 2016, 54, 2655–2660. [Google Scholar] [CrossRef] [PubMed]
- Etienne, K.A.; Kano, R.; Balajee, S.A. Development and validation of a microsphere-based luminex assay for rapid identification of clinically relevant Aspergilli. J. Clin. Microbiol. 2009, 47, 1096–1100. [Google Scholar] [CrossRef] [PubMed]
- Bhimji, A.; Bhaskaran, A.; Singer, L.G.; Kumar, D.; Humar, A.; Pavan, R.; Lipton, J.; Kuruvilla, J.; Schuh, A.; Yee, K.; et al. Aspergillus galactomannan detection in exhaled breath condensate compared to bronchoalveolar lavage fluid for the diagnosis of invasive aspergillosis in immunocompromised patients. Clin. Microbiol. Infect. 2018, 24, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Mello, E.; Posteraro, B.; Vella, A.; De Carolis, E.; Torelli, R.; D’Inzeo, T.; Verweij, P.E.; Sanguinetti, M. Susceptibility testing of common and uncommon Aspergillus species against posaconazole and other mold-active antifungal azoles using the Sensititre method. Antimicrob. Agents Chemother. 2017, 61, e00168-17. [Google Scholar] [CrossRef] [PubMed]
- Hageskal, G.; Kristensen, R.; Fristad, R.F.; Skaar, I. Emerging pathogen Aspergillus calidoustus colonizes water distribution systems. Med. Mycol. 2011, 49, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Egli, A.; Fuller, J.; Humar, A.; Lien, D.; Weinkauf, J.; Nador, R.; Kapasi, A.; Kumar, D. Emergence of Aspergillus calidoustus infection in the era of post-transplantation azole prophylaxis. Transplantation 2012, 94, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Springer, J.; White, P.L.; Hamilton, S.; Michel, D.; Barnes, R.A.; Einsele, H.; Löffler, J. Comparison of performance characteristics of Aspergillus PCR in testing a range of blood-based samples in accordance with international methodological recommendations. J. Clin. Microbiol. 2016, 54, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Wang, K.; Gao, W.; Su, X.; Qian, Q.; Lu, X.; Song, Y.; Guo, Y.; Shi, Y. Evaluation of PCR on bronchoalveolar lavage fluid for diagnosis of invasive aspergillosis: A bivariate meta-analysis and systematic review. PLoS ONE 2011, 6, e28467. [Google Scholar] [CrossRef] [PubMed]
- Reinwald, M.; Buchheidt, D.; Hummel, M.; Duerken, M.; Bertz, H.; Schwerdtfeger, R.; Reuter, S.; Kiehl, M.G.; Barreto-Miranda, M.; Hofmann, W.-K.; et al. Diagnostic performance of an Aspergillus-specific nested PCR assay in cerebrospinal fluid samples of immunocompromised patients for detection of central nervous system aspergillosis. PLoS ONE 2013, 8, e56706. [Google Scholar] [CrossRef] [PubMed]
- White, P.L.; Bretagne, S.; Klingspor, L.; Melchers, W.J.G.; McCulloch, E.; Schulz, B.; Finnstrom, N.; Mengoli, C.; Barnes, R.A.; Donnelly, J.P.; et al. European Aspergillus PCR initiative Aspergillus PCR: One step closer to standardization. J. Clin. Microbiol. 2010, 48, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- White, P.L.; Perry, M.D.; Loeffler, J.; Melchers, W.; Klingspor, L.; Bretagne, S.; McCulloch, E.; Cuenca-Estrella, M.; Finnstrom, N.; Donnelly, J.P.; et al. European Aspergillus PCR initiative critical stages of extracting DNA from Aspergillus fumigatus in whole-blood specimens. J. Clin. Microbiol. 2010, 48, 3753–3755. [Google Scholar] [CrossRef] [PubMed]
- Alanio, A.; Bretagne, S. Challenges in microbiological diagnosis of invasive Aspergillus infections. F1000Research 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- White, P.L.; Parr, C.; Thornton, C.; Barnes, R.A. Evaluation of real-time PCR, galactomannan enzyme-linked immunosorbent assay (ELISA), and a novel lateral-flow device for diagnosis of invasive aspergillosis. J. Clin. Microbiol. 2013, 51, 1510–1516. [Google Scholar] [CrossRef] [PubMed]
- Aguado, J.M.; Vázquez, L.; Fernández-Ruiz, M.; Villaescusa, T.; Ruiz-Camps, I.; Barba, P.; Silva, J.T.; Batlle, M.; Solano, C.; Gallardo, D.; et al. PCRAGA Study Group; Spanish Stem Cell Transplantation Group; Study Group of Medical Mycology of the Spanish Society of Clinical Microbiology and Infectious Diseases; Spanish Network for Research in Infectious Diseases. Serum galactomannan versus a combination of galactomannan and polymerase chain reaction-based Aspergillus DNA detection for early therapy of invasive aspergillosis in high-risk hematological patients: A randomized controlled trial. Clin. Infect. Dis. 2015, 60, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Hebart, H.; Klingspor, L.; Klingebiel, T.; Loeffler, J.; Tollemar, J.; Ljungman, P.; Wandt, H.; Schaefer-Eckart, K.; Dornbusch, H.J.; Meisner, C.; et al. A prospective randomized controlled trial comparing PCR-based and empirical treatment with liposomal amphotericin B in patients after allo-SCT. Bone Marrow Transplant. 2009, 43, 553–561. [Google Scholar] [CrossRef] [PubMed]
- De Pauw, B.; Walsh, T.J.; Donnelly, J.P.; Stevens, D.A.; Edwards, J.E.; Calandra, T.; Pappas, P.G.; Maertens, J.; Lortholary, O.; Kauffman, C.A.; et al. Revised definitions of invasive fungal disease from the European Organization for Research and Treatment of Cancer/Invasive Fungal Infections Cooperative Group and the National Institute of Allergy and Infectious Diseases Mycoses Study Group (EORTC/MSG) consensus group. Clin. Infect. Dis. 2008, 46, 1813–1821. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashu, E.E.; Xu, J. Strengthening the One Health Agenda: The Role of Molecular Epidemiology in Aspergillus Threat Management. Genes 2018, 9, 359. https://doi.org/10.3390/genes9070359
Ashu EE, Xu J. Strengthening the One Health Agenda: The Role of Molecular Epidemiology in Aspergillus Threat Management. Genes. 2018; 9(7):359. https://doi.org/10.3390/genes9070359
Chicago/Turabian StyleAshu, Eta E., and Jianping Xu. 2018. "Strengthening the One Health Agenda: The Role of Molecular Epidemiology in Aspergillus Threat Management" Genes 9, no. 7: 359. https://doi.org/10.3390/genes9070359
APA StyleAshu, E. E., & Xu, J. (2018). Strengthening the One Health Agenda: The Role of Molecular Epidemiology in Aspergillus Threat Management. Genes, 9(7), 359. https://doi.org/10.3390/genes9070359