Nitrogen Supply and Leaf Age Affect the Expression of TaGS1 or TaGS2 Driven by a Constitutive Promoter in Transgenic Tobacco

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Construction
2.2. Plant Transformation and Growth Conditions
2.3. RNA Isolation and RT-qPCR Analysis
2.4. GS Activity Assay and Western Blotting
2.5. Determination of Free NH4+, NO3−, Total Amino Acid, and Total Chlorophyll Levels
2.6. Statistics
3. Results
3.1. Identification of Transgenic Tobacco
3.2. Characteristics of GS Expression in GS1-TR and GS2-TR
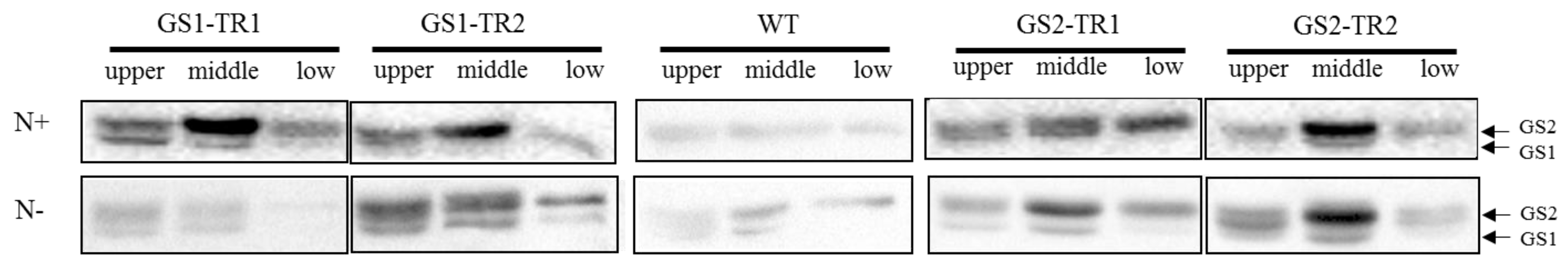
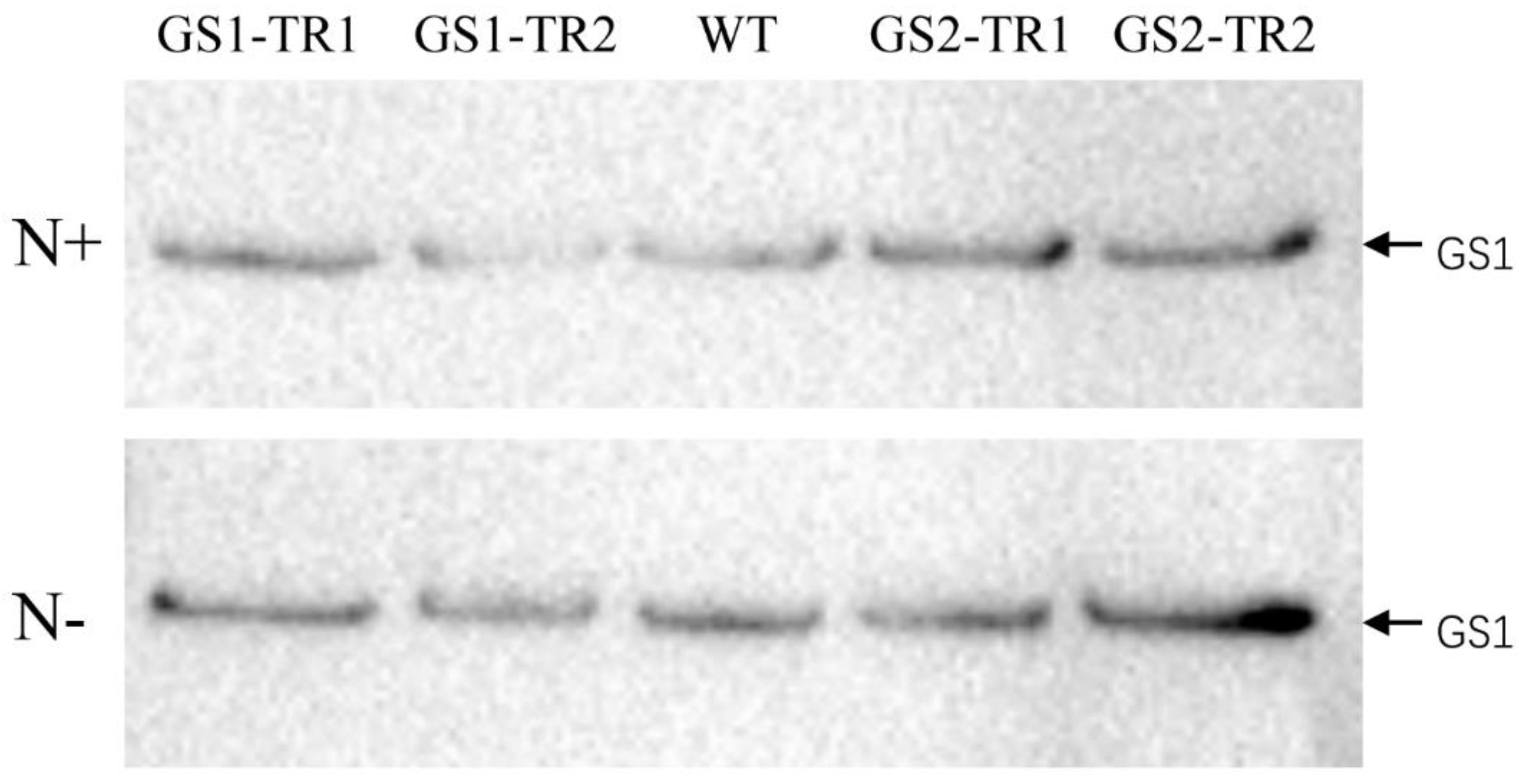
3.3. Characteristics of GS Subunit Expression in GS1-TR and GS2-TR
3.4. Characteristics of GS Activity in GS1-TR and GS2-TR
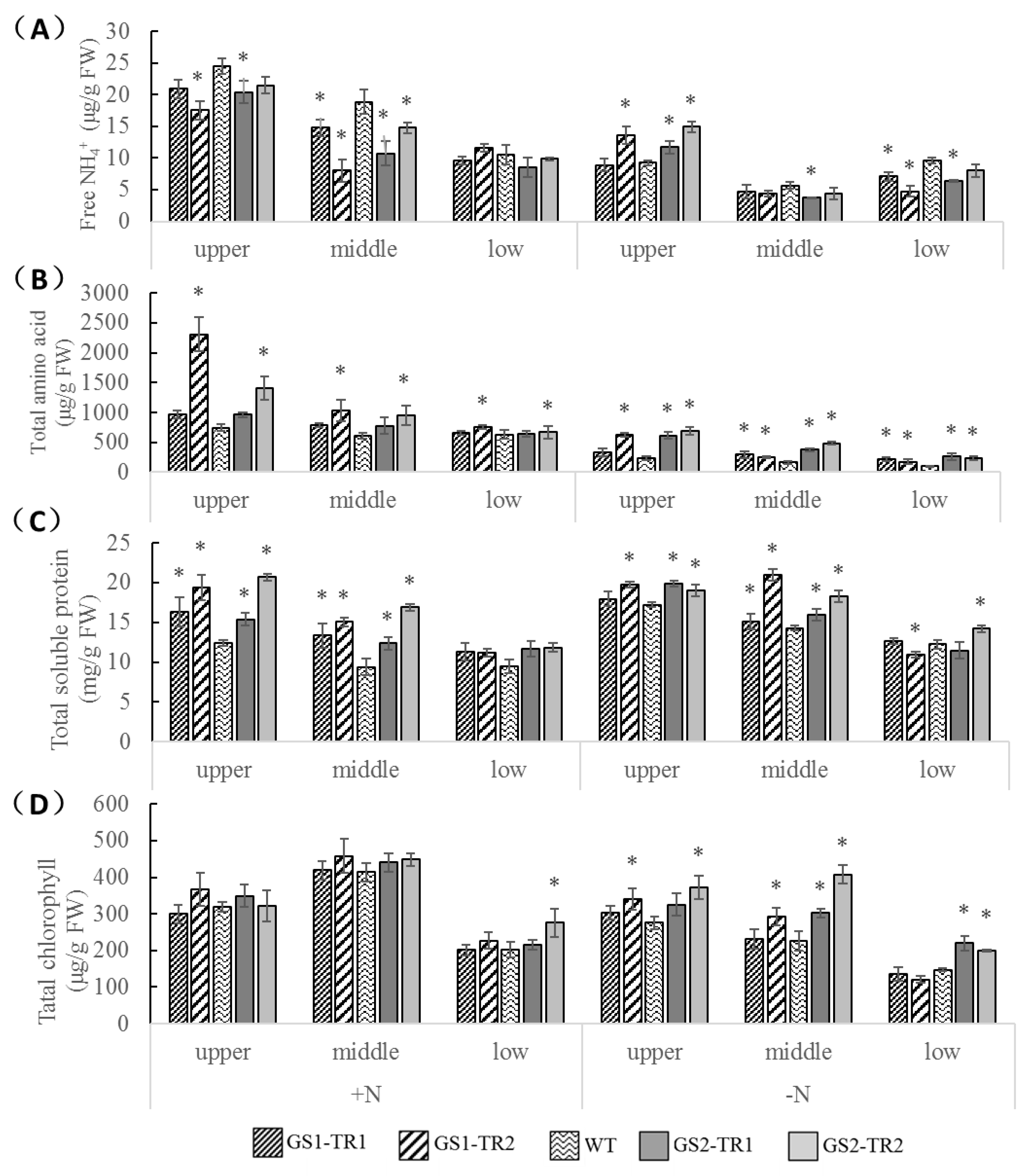
3.5. Characteristics of Nitrogen Metabolic Status in GS1-TR and GS2-TR
3.6. Phenotypes of GS1-TR and GS2-TR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kaur, G.; Asthir, B.; Bains, N.S.; Farooq, M. Nitrogen nutrition, its assimilation and remobilization in diverse wheat genotypes. Int. J. Agric. Biol. 2015, 17, 531–538. [Google Scholar] [CrossRef]
- Kichey, T.; Heumez, E.; Pocholle, D.; Pageau, K.; Vanacker, H.; Dubois, F.; Le Gouis, J.; Hirel, B. Combined agronomic and physiological aspects of nitrogen management in wheat highlight a central role for glutamine synthetase. New Phytol. 2006, 169, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.P.; Vitousek, P.M. Nitrogen in agriculture: Balancing the cost of an essential resource. Annu. Rev. Environ. Resour. 2009, 34, 97–125. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D.G. Technical report: Human alteration of the global nitrogen cycle: Sources and consequences. Ecol. Appl. 1997, 7, 737–750. [Google Scholar] [CrossRef]
- Thomsen, H.C.; Eriksson, D.; Møller, I.S.; Schjoerring, J.K. Cytosolic glutamine synthetase: A target for improvement of crop nitrogen use efficiency? Trends Plant Sci. 2014, 19, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Miflin, B.J.; Lea, P.J. Amino acid metabolism. Annu. Rev. Plant Physiol. 1977, 28, 299–329. [Google Scholar] [CrossRef]
- Edwards, J.W.; Walker, E.L.; Coruzzi, G.M. Cell-specific expression in transgenic plants reveals nonoverlapping roles for chloroplast and cytosolic glutamine synthetase. Proc. Natl. Acad. Sci. USA 1990, 87, 3459–3463. [Google Scholar] [CrossRef] [PubMed]
- Tobin, A.; Ridley, S.; Stewart, G. Changes in the activities of chloroplast and cytosolic isoenzymes of glutamine synthetase during normal leaf growth and plastid development in wheat. Planta 1985, 163, 544–548. [Google Scholar] [CrossRef] [PubMed]
- Hirel, B.; Gadal, P. Glutamine synthetase in rice a comparative study of the enzymes from roots and leaves. Plant Physiol. 1980, 66, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Bernard, S.M.; Møller, A.L.B.; Dionisio, G.; Kichey, T.; Jahn, T.P.; Dubois, F.; Baudo, M.; Lopes, M.S.; Tercé-Laforgue, T.; Foyer, C.H. Gene expression, cellular localisation and function of glutamine synthetase isozymes in wheat (Triticum aestivum L.). Plant Mol. Biol. 2008, 67, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Wallsgrove, R.M.; Turner, J.C.; Hall, N.P.; Kendall, A.C.; Bright, S.W. Barley mutants lacking chloroplast glutamine synthetase—biochemical and genetic analysis. Plant Physiol. 1987, 83, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Avila-Ospina, L.; Moison, M.; Yoshimoto, K.; Masclaux-Daubresse, C. Autophagy, plant senescence, and nutrient recycling. J. Exp. Bot. 2014, 65, 3799–3811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, M.; Møller, I.; Schjoerring, J. Two cytosolic glutamine synthetase isoforms play specific roles for seed germination and seed yield structure in Arabidopsis. J. Exp. Bot. 2015, 66, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Brugiere, N.; Dubois, F.; Limami, A.M.; Lelandais, M.; Roux, Y.; Sangwan, R.S.; Hirel, B. Glutamine synthetase in the phloem plays a major role in controlling proline production. Plant Cell 1999, 11, 1995–2012. [Google Scholar] [CrossRef] [PubMed]
- Kamachi, K.; Yamaya, T.; Mae, T.; Ojima, K. A role for glutamine synthetase in the remobilization of leaf nitrogen during natural senescence in rice leaves. Plant Physiol. 1991, 96, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Oaks, A.; Hirel, B. Nitrogen metabolism in roots. Annu. Rev. Plant Physiol. 1985, 36, 345–365. [Google Scholar] [CrossRef]
- Bernard, S.M.; Habash, D.Z. The importance of cytosolic glutamine synthetase in nitrogen assimilation and recycling. New Phytol. 2009, 182, 608–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funayama, K.; Kojima, S.; Tabuchi-Kobayashi, M.; Sawa, Y.; Nakayama, Y.; Hayakawa, T.; Yamaya, T. Cytosolic glutamine synthetase1;2 is responsible for the primary assimilation of ammonium in rice roots. Plant Cell Physiol. 2013, 54, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Hirel, B.; Le, G.J.; Ney, B.; Gallais, A. The challenge of improving nitrogen use efficiency in crop plants: Towards a more central role for genetic variability and quantitative genetics within integrated approaches. J. Exp. Bot. 2007, 58, 2369–2387. [Google Scholar] [CrossRef] [PubMed]
- Kichey, T.; Hirel, B.; Heumez, E.; Dubois, F.; Le Gouis, J. In winter wheat (Triticum aestivum L.), post-anthesis nitrogen uptake and remobilisation to the grain correlates with agronomic traits and nitrogen physiological markers. Field Crops Res. 2007, 102, 22–32. [Google Scholar] [CrossRef]
- Martin, A.; Lee, J.; Kichey, T.; Gerentes, D.; Zivy, M.; Tatout, C.; Dubois, F.; Balliau, T.; Valot, B.; Davanture, M. Two cytosolic glutamine synthetase isoforms of maize are specifically involved in the control of grain production. Plant Cell 2006, 18, 3252–3274. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiong, S.; Wei, Y.; Meng, X.; Wang, X.; Ma, X. The role of glutamine synthetase isozymes in enhancing nitrogen use efficiency of n-efficient winter wheat. Sci. Rep. 2017, 7, 1000. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Xing, G.; Niu, X.; Dou, C.; Han, Y. Progress and application prospects of glutamine synthase in plants. Chin. J. Biotechnol. 2015, 31, 1301–1312. [Google Scholar]
- Habash, D.Z.; Massiah, A.J.; Rong, H.L.; Wallsgrove, R.M.; Leigh, R.A. The role of cytosolic glutamine synthetase in wheat. Ann. Appl. Biol. 2001, 138, 83–89. [Google Scholar] [CrossRef]
- Fuentes, S.I.; Allen, D.J.; Ortiz-Lopez, A.; Hernández, G. Over-expression of cytosolic glutamine synthetase increases photosynthesis and growth at low nitrogen concentrations. J. Exp. Bot. 2001, 52, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, F.; Fu, J.; Cantón, F.R.; García-Gutiérrez, A.; Cánovas, F.M.; Kirby, E.G. Expression of a conifer glutamine synthetase gene in transgenic poplar. Planta 1999, 210, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, I.C.; Brears, T.; Knight, T.J.; Clark, A.; Coruzzi, G.M. Overexpression of cytosolic glutamine synthetase. Relation to nitrogen, light, and photorespiration. Plant Physiol. 2002, 129, 1170–1180. [Google Scholar] [CrossRef] [PubMed]
- Brauer, E.K.; Rochon, A.; Bi, Y.M.; Bozzo, G.G.; Rothstein, S.J.; Shelp, B.J. Reappraisal of nitrogen use efficiency in rice overexpressing glutamine synthetase1. Physiol. Plant. 2011, 141, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Zhou, Y.; Xiao, J.; Li, X.; Zhang, Q.; Lian, X. Overexpressed glutamine synthetase gene modifies nitrogen metabolism and abiotic stress responses in rice. Plant Cell Rep. 2009, 28, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Huang, M.; Su, J. Highly effective expression of glutamine synthetase genes GS1 and GS2 in transgenic rice plants increases nitrogen-deficiency tolerance. J. Plant Physiol. Mol. Biol. 2005, 31, 492–498. [Google Scholar]
- Migge, A.; Carrayol, E.; Hirel, B.; Becker, T.W. Leaf-specific overexpression of plastidic glutamine synthetase stimulates the growth of transgenic tobacco seedlings. Planta 2000, 210, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Harper, C.J.; Hayward, D.; Kidd, M.; Wiid, I.; van Helden, P. Glutamate dehydrogenase and glutamine synthetase are regulated in response to nitrogen availability in Mycobacterium smegmatis. BMC Microbiol. 2010, 10, 138. [Google Scholar] [CrossRef] [PubMed]
- Caputo, C.; Criado, M.V.; Roberts, I.N.; Gelso, M.A.; Barneix, A.J. Regulation of glutamine synthetase 1 and amino acids transport in the phloem of young wheat plants. Plant Physiol. Biochem. 2009, 47, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Fu, J.; Drijber, R.A.; Gao, Y. Expression patterns of five genes involved in nitrogen metabolism in two winter wheat (Triticum aestivum L.) genotypes with high and low nitrogen utilization efficiencies. J. Cereal Sci. 2015, 61, 48–54. [Google Scholar] [CrossRef]
- Avila-Ospina, L.; Marmagne, A.; Talbotec, J.; Krupinska, K.; Masclaux-Daubresse, C. The identification of new cytosolic glutamine synthetase and asparagine synthetase genes in barley (Hordeum vulgare L.), and their expression during leaf senescence. J. Exp. Bot. 2015, 66, 2013–2026. [Google Scholar] [CrossRef] [PubMed]
- Brugière, N.; Dubois, F.; Masclaux, C.; Sangwan, R.S.; Hirel, B. Immunolocalization of glutamine synthetase in senescing tobacco (Nicotiana tabacum L.) leaves suggests that ammonia assimilation is progressively shifted to the mesophyll cytosol. Planta 2000, 211, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Ortega, J.L.; Temple, S.J.; Sengupta-Gopalan, C. Constitutive overexpression of cytosolic glutamine synthetase (GS1) gene in transgenic alfalfa demonstrates that GS1 may be regulated at the level of RNA stability and protein turnover. Plant Physiol. 2001, 126, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Lan-Ying, L.; Kononov, M.E.; Burgund, B.; Frame, B.R.; Kan, W.; Gelvin, S.B. Novel plant transformation vectors containing the superpromoter. Plant Physiol. 2007, 145, 1294–1300. [Google Scholar]
- Ni, M.; Cui, D.; Einstein, J.; Narasimhulu, S.; Vergara, C.E.; Gelvin, S.B. Strength and tissue specificity of chimeric promoters derived from the octopine and mannopine synthase genes. Plant J. 1995, 7, 661–676. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Chen, Z.-Z.; Zhou, X.-F.; Yin, H.-B.; Li, X.; Xin, X.-F.; Hong, X.-H.; Zhu, J.-K.; Gong, Z. Overexpression of SOS (salt overly sensitive) genes increases salt tolerance in transgenic Arabidopsis. Mol. Plant 2009, 2, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.Y.; Duan, L.S.; Zhang, J.C.; Zhang, Z.X.; Mi, G.Q.; Ren, H.Z. Cucumber (Cucumis sativus L.) over-expressing cold-induced transcriptome regulator ICE1 exhibits changed morphological characters and enhances chilling tolerance. Sci. Hortic. 2010, 124, 29–33. [Google Scholar] [CrossRef]
- Li, Y.J.; Zhang, J.C.; Zhang, J.; Hao, L.; Hua, J.P.; Duan, L.S.; Zhang, M.C.; Li, Z.H. Expression of an Arabidopsis molybdenum cofactor sulphurase gene in soybean enhances drought tolerance and increases yield under field conditions. Plant Biotechnol. J. 2013, 11, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, T.; Li, G.; Cheng, Z.; Liu, N.; Ma, X. Cloning of glutamine synthetases in wheat and analysis of their expression characteristics. J. Henan Agric. Univ. 2012, 46, 487–492. [Google Scholar]
- Horsch, R.B.; Fraley, R.T. A simple and general method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar]
- Ma, X.; Li, L.; Zhao, P.; Xiong, S.; Guo, F. Effect of water control on activities of nitrogen assimilation enzymes and grain quality in winter wheat. Acta Phytoecol. Sin. 2005, 29, 48–53. [Google Scholar]
- Gordon, S.A.; Fleck, A.; Bell, J. Optimal conditions for the estimation of ammonium by the Berthelot reaction. Ann. Clin. Biochem. 1978, 15, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Xie, H. Determination of nitrogen content in nitrate by salicylic acid colorimetry in water. Guizhou Agric. Sci. 1999, 27, 40–41. [Google Scholar]
- Graan, T.; Ort, D.R. Quantitation of the rapid electron donors to p700, the functional plastoquinone pool, and the ratio of the photosystems in spinach chloroplasts. J. Biol. Chem. 1984, 259, 14003–14010. [Google Scholar] [PubMed]
- Carvalho, H.; Pereira, S.; Sunkel, C.; Salema, R. Detection of a cytosolic glutamine synthetase in leaves of Nicotiana tabacum L. by immunocytochemical methods. Plant Physiol. 1992, 100, 1591–1594. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.J.; Smith, S.J. Cytosolic nitrate ion homeostasis: Could it have a role in sensing nitrogen status? Ann. Bot. 2008, 101, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Hirel, B.; Marsolier, M.C.; Hoarau, A.; Hoarau, J.; Brangeon, J.; Schafer, R.; Verma, D.P. Forcing expression of a soybean root glutamine synthetase gene in tobacco leaves induces a native gene encoding cytosolic enzyme. Plant Mol. Biol. 1992, 20, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.J.; Beatty, P.H.; Good, A.G.; Muench, D.G. Manipulation of microRNA expression to improve nitrogen use efficiency. Plant Sci. 2013, 210, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Jones-Rhoades, M.W.; Bartel, D.P.; Bartel, B. MicroRNAs and their regulatory roles in plants. Annu. Rev. Plant Biol. 2006, 57, 19–53. [Google Scholar] [CrossRef] [PubMed]
- Sunkar, R.; Chinnusamy, V.; Zhu, J.H.; Zhu, J.K. Small RNAs as big players in plant abiotic stress responses and nutrient deprivation. Trends Plant Sci. 2007, 12, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhong, S.; Li, X.; Li, W.; Rothstein, S.J.; Zhang, S.; Bi, Y.; Xie, C. Genome-wide identification of microRNAs in response to low nitrate availability in maize leaves and roots. PLoS ONE 2011, 6, e28009. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, S.; Nonis, A.; Begheldo, M.; Manoli, A.; Palme, K.; Caporale, G.; Ruperti, B.; Quaggiotti, S. Expression and tissue-specific localization of nitrate-responsive miRNAs in roots of maize seedlings. Plant Cell Environ. 2012, 35, 1137–1155. [Google Scholar] [CrossRef] [PubMed]
- Nischal, L.; Mohsin, M.; Khan, I.; Kardam, H.; Wadhwa, A.; Abrol, Y.P.; Iqbal, M.; Ahmad, A. Identification and comparative analysis of microRNAs associated with low-N tolerance in rice genotypes. PLoS ONE 2012, 7, e50261. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Ding, H.; Zhu, J.-K.; Zhang, F.; Li, W.-X. Involvement of miR169 in the nitrogen-starvation responses in Arabidopsis. New Phytol. 2011, 190, 906–915. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; He, H.; Yu, D. Identification of nitrogen starvation-responsive microRNAs in Arabidopsis thaliana. PLoS ONE 2012, 7, e48951. [Google Scholar] [CrossRef] [PubMed]
- El Omari, R.; Rueda-López, M.; Avila, C.; Crespillo, R.; Nhiri, M.; Cánovas, F.M. Ammonium tolerance and the regulation of two cytosolic glutamine synthetases in the roots of sorghum. Funct. Plant Biol. 2010, 37, 55–63. [Google Scholar] [CrossRef]
- Oliveira, I.C.; Coruzzi, G.M. Carbon and amino acids reciprocally modulate the expression of glutamine synthetase in Arabidopsis. Plant Physiol. 1999, 121, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.W.; Coruzzi, G.M. Photorespiration and light act in concert to regulate the expression of the nuclear gene for chloroplast glutamine synthetase. Plant Cell 1989, 1, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and signaling aspects underpinning the regulation of plant carbon nitrogen interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef] [PubMed]
- Seger, M.; Gebril, S.; Tabilona, J.; Peel, A.; Sengupta-Gopalan, C. Impact of concurrent overexpression of cytosolic glutamine synthetase (GS1) and sucrose phosphate synthase (SPS) on growth and development in transgenic tobacco. Planta 2015, 241, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.M. The complex regulation of senescence. CRC Crit. Rev. Plant Sci. 2012, 31, 124–147. [Google Scholar] [CrossRef]
- Tegeder, M. Transporters involved in source to sink partitioning of amino acids and ureides: Opportunities for crop improvement. J. Exp. Bot. 2014, 65, 1865–1878. [Google Scholar] [CrossRef] [PubMed]
- Masclaux-Daubresse, C.; Reisdorf-Cren, M.; Orsel, M. Leaf nitrogen remobilisation for plant development and grain filling. Plant Biol. 2008, 10, 23–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N+ | N− | |||||||
|---|---|---|---|---|---|---|---|---|
| Plant Height (cm) | Leaf Area (cm2) | Plant DW (g) | Root DW (g) | Plant Height (cm) | Leaf Area (cm2) | Plant DW (g) | Root DW (g) | |
| GS1-TR1 | 35.5 ± 1.1 * | 3732 ± 449 * | 5.2 ± 0.8 | 0.64 ± 0.09 * | 21.2 ± 2 | 1392 ± 66 * | 3.9 ± 0.4 * | 0.72 ± 0.03 * |
| GS1-TR2 | 33 ± 1.5 * | 3340 ± 323 * | 6 ± 1.2 | 0.65 ± 0.07 * | 25.5 ± 2.3 * | 1552 ± 75 * | 5.2 ± 0.5 * | 0.89 ± 0.04 * |
| WT | 26 ± 2.3 | 2331 ± 317 | 4.2 ± 0.2 | 0.51 ± 0.02 | 20.3 ± 1 | 1190 ± 53 | 3.1 ± 0.2 | 0.55 ± 0.04 |
| GS2-TR1 | 27.8 ± 1.9 | 2885 ± 241 | 5.3 ± 1.2 | 0.55 ± 0.07 | 19 ± 1.1 | 1442 ± 126 * | 3.5 ± 0.1 | 0.59 ± 0.02 |
| GS2-TR2 | 27.2 ± 2.5 | 3093 ± 299 | 5.1 ± 0.6 | 0.65 ± 0.04 * | 14.8 ± 0.2 * | 1338 ± 42 | 3.6 ± 0.2 | 0.67 ± 0.01 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, Y.; Shi, A.; Jia, X.; Zhang, Z.; Ma, X.; Gu, M.; Meng, X.; Wang, X. Nitrogen Supply and Leaf Age Affect the Expression of TaGS1 or TaGS2 Driven by a Constitutive Promoter in Transgenic Tobacco. Genes 2018, 9, 406. https://doi.org/10.3390/genes9080406
Wei Y, Shi A, Jia X, Zhang Z, Ma X, Gu M, Meng X, Wang X. Nitrogen Supply and Leaf Age Affect the Expression of TaGS1 or TaGS2 Driven by a Constitutive Promoter in Transgenic Tobacco. Genes. 2018; 9(8):406. https://doi.org/10.3390/genes9080406
Chicago/Turabian StyleWei, Yihao, Aibo Shi, Xiting Jia, Zhiyong Zhang, Xinming Ma, Mingxin Gu, Xiaodan Meng, and Xiaochun Wang. 2018. "Nitrogen Supply and Leaf Age Affect the Expression of TaGS1 or TaGS2 Driven by a Constitutive Promoter in Transgenic Tobacco" Genes 9, no. 8: 406. https://doi.org/10.3390/genes9080406
APA StyleWei, Y., Shi, A., Jia, X., Zhang, Z., Ma, X., Gu, M., Meng, X., & Wang, X. (2018). Nitrogen Supply and Leaf Age Affect the Expression of TaGS1 or TaGS2 Driven by a Constitutive Promoter in Transgenic Tobacco. Genes, 9(8), 406. https://doi.org/10.3390/genes9080406