
Transpiration of a Tropical Dry Deciduous Forest in Yucatan, Mexico

 , , ,
, , , 

Abstract
:1. Introduction
2. Materials and Methods
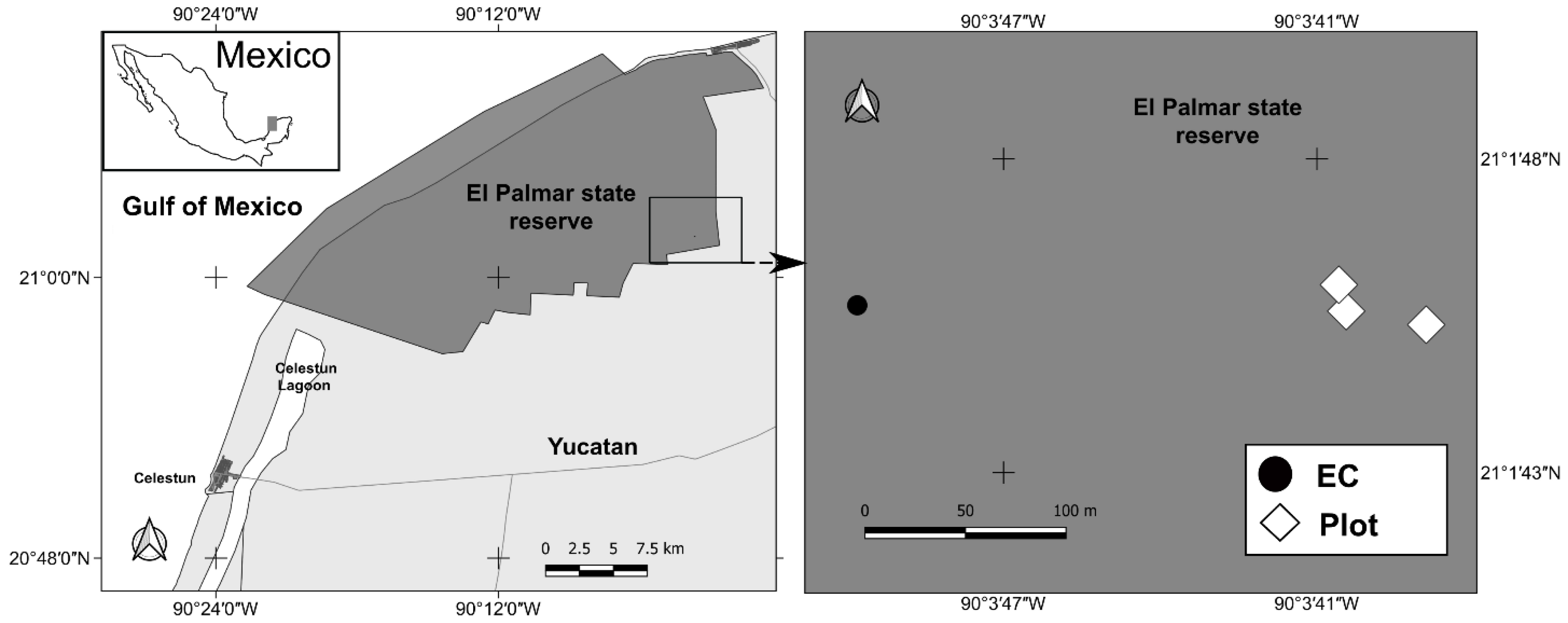
2.1. Study Area
2.2. Environmental Conditions
2.3. Sap Flux
2.4. Estimation of Stand Transpiration
3. Results
3.1. Environmental Conditions
3.2. Soil Water Content
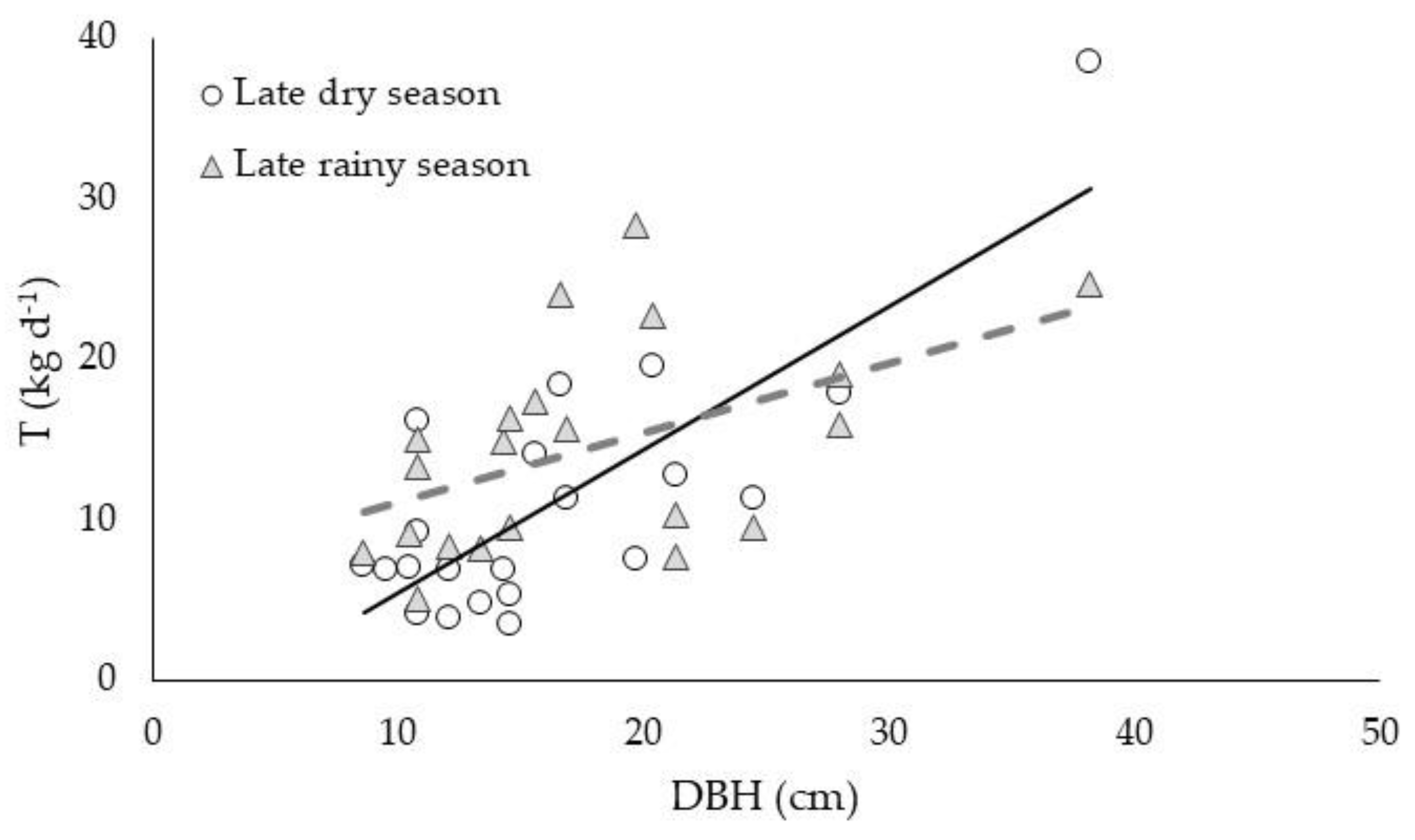
3.3. Allometric Relationships
3.4. DBH Categories
3.5. Stand Level
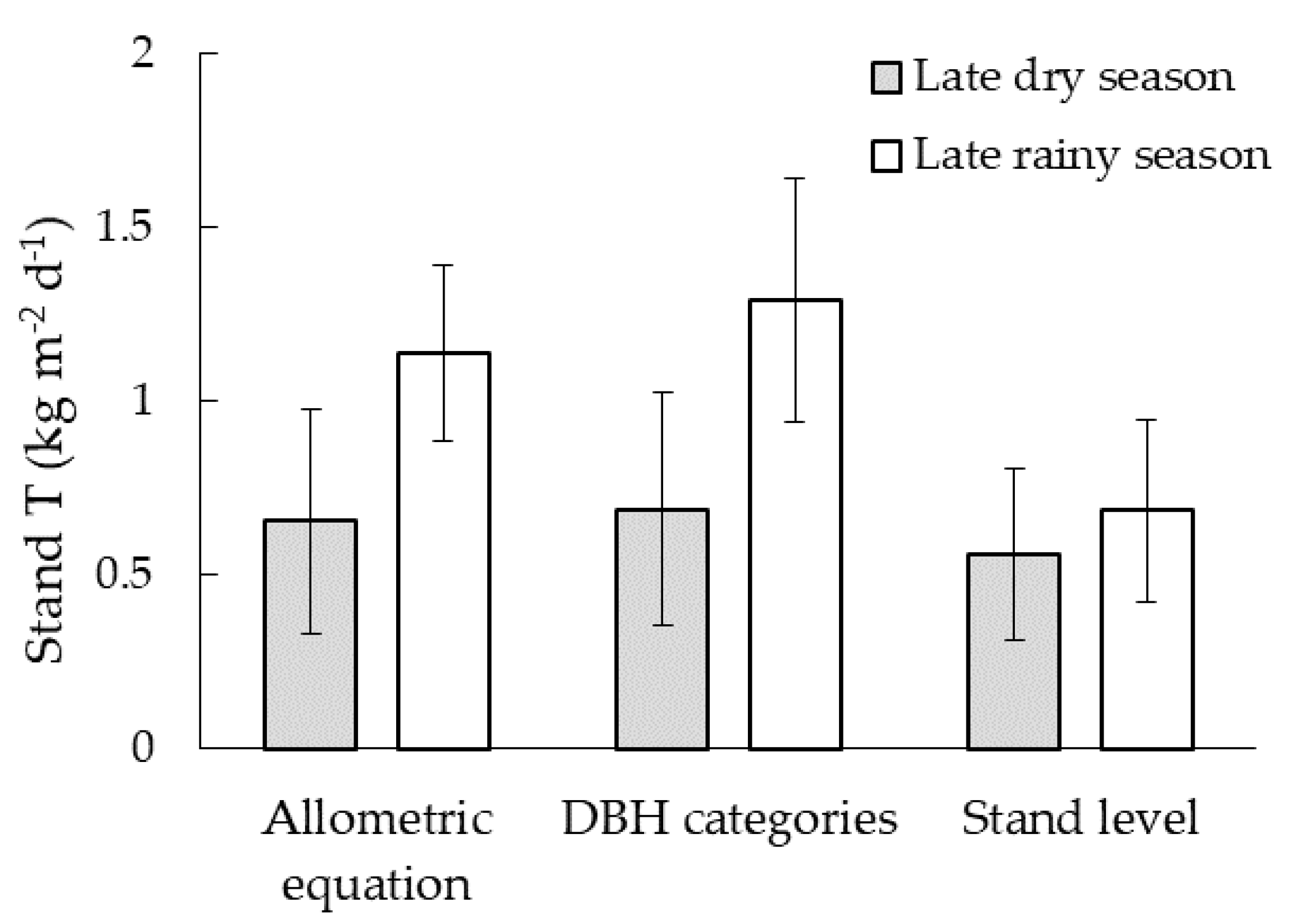
3.6. Comparison of Upscaling Methods
4. Discussion
Transpiration and Evapotranspiration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Healy, R.W.; Winter, T.C.; LaBaugh, J.W.; Franke, O.L. Water Budgets: Foundations for Effective Water-Resources and Environmental Management; US Geological Survey: Reston, VA, USA, 2007; Volume 1308.
- Bosch, J.M.; Hewlett, J.D. A review of catchment experiments to determine the effect of vegetation changes on water yield and evapotranspiration. J. Hydrol. 1982, 55, 3–23. [Google Scholar] [CrossRef]
- Oren, R.; Phillips, N.; Katul, G.; Ewers, B.; Pataki, D.E. Scaling xylem sap flux and soil water flux in forests. Ann. Sci. For. 1998, 55, 191–216. [Google Scholar] [CrossRef]
- Williams, D.G.; Cable, W.; Hultine, K.; Heodjes, J.C.B.; Yepez, E.A.; Simonneaux, V.; Er-Raki, S.; Boulet, G.; de Bruin, H.A.R.; Chehbouni, A.; et al. Evapotranspiration components determined by stable isotope, sap flow and eddy covariance techniques. Agric. For. Meteorol. 2004, 125, 241–258. [Google Scholar] [CrossRef]
- Oki, T.; Kanae, S. Global hydrological cycles and world water resources. Science 2006, 313, 1068–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drexler, J.Z.; Snyder, R.L.; Spano, D.; Paw, U.K.T. A review of models and micrometeorological methods used to estimate wetland evapotranspiration. Hydrol. Process. 2004, 18, 2071–2101. [Google Scholar] [CrossRef]
- Bolbotín-Nesvará, C.; Calera-Belmonte, A.; González-Piqueras, J.; Campos-Rodríguez, I.; López-González, M.L.; Torres-Prieto, E. Comparación de los sistemas covarianza y relación de Bowen en la evapotranspiración de un viñedo bajo clima semi-árido. Agrociencias 2011, 45, 87–103. [Google Scholar]
- Evett, S.R.; Kustas, W.P.; Gowda, P.H.; Anderson, M.C.; Prueger, J.H.; Howell, T.A. Overview of the bushland evapotranspiration and agricultural remote sensing experiment 2008 (BEAREX08): A field experiment evaluating methods for quantifying ET at multiple scales. Adv. Water Resour. 2012, 50, 4–19. [Google Scholar] [CrossRef]
- Rezaei, M.; Valipour, M.; Valipour, M. Modelling evapotranspiration to increase the accuracy of the estimations based on the climatic parameters. Water Conserv. Sci. Eng. 2016, 1, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Kingston, D.G.; Todd, M.C.; Taylor, R.G.; Thompson, J.R.; Arnell, N.W. Uncertainty in the estimation of potential evapotranspiration under climate change. Geophys. Res. Lett. 2009, 36, L20403. [Google Scholar] [CrossRef] [Green Version]
- Mahrt, L. Flux Sampling Errors for Aircraft and Towers. J. Atmos. Ocean. Technol. 1998, 15, 416–429. [Google Scholar] [CrossRef]
- Deb Burman, P.K.; Sarma, D.; Morrison, R.; Karipot, A.; Chakraborty, S. Seasonal variation of evapotranspiration and its effect on the surface energy budget closure at a tropical forest over north-east India. J. Earth Syst. Sci. 2019, 128, 127. [Google Scholar] [CrossRef] [Green Version]
- Wilson, K.B.; Hanson, P.J.; Mulholland, P.J.; Baldocchi, D.D.; Wullschleger, S.D. A comparison of methods for determining forest evapotranspiration and its components: Sap-flow, soil water budget, eddy covariance and catchment water balance. Agric. For. Meteorol. 2001, 106, 153–168. [Google Scholar] [CrossRef]
- Burba, G. Eddy Covariance Method for Scientific, Industrial, Agricultural, and Regulatory Applications; LI-COR Biosciences: Lincoln, NE, USA, 2013; ISBN 978-0-615-76827-4. [Google Scholar]
- Gutierrez-Jurado, H.A.; Uuh-Sonda, J.M.; Méndez-Barroso, L.M.; Figueroa-Espinoza, B.; Yepez, E.A.; Rojas-Robles, N.E. Evapotranspiration partitioning of tropical deciduous forests of North America: A model-based comparison of two eddy-covariance sites with contrasting hydroclimatic regimes. In Proceedings of the AGU Fall Meeting 2021, New Orleans, LA, USA, 13-17 December 2021. [Google Scholar]
- Salazar-Martínez, D.; Holwerda, F.; Holmes, T.R.H.; Yépez, E.A. Evaluation of remote sensing-based evapotranspiration products at low-latitude eddy covariance sites. J. Hydrol. under review. 2022.
- Jasechko, S.; Sharp, Z.D.; Gibson, J.J.; Birks, S.J.; Yi, Y.; Fawcett, P.F. Terrestrial water fluxes dominated by transpiration. Nature 2013, 496, 347–350. [Google Scholar] [CrossRef]
- Nelson, J.A.; Pérez-Priego, O.; Zhou, S.; Poyatos, R.; Zhang, Y.; Blanken, P.D.; Gimeno, T.E.; Wohlfahrt, G.; Desai, A.R.; Gioli, B.; et al. Ecosystem transpiration and evaporation: Insights from three water flux partitioning methods across FLUXNET sites. Glob. Chang. Biol. 2020, 26, 6916–6930. [Google Scholar] [CrossRef]
- Tijerina, C.L. Uso Eficiente del Agua en Unidades de Riego Para el Desarrollo Rural. Diplomado; Colegio de Postgraduados: Montecillo, México, 1992. [Google Scholar]
- Huxman, T.E.; Wilcox, B.P.; Breshears, D.D.; Scott, R.L.; Snyder, K.A.; Small, E.E.; Hultine, K.; Pockman, W.T.; Robert, B. Ecohydrological implications of woody plant encroachment. Ecology 2005, 86, 308–319. [Google Scholar] [CrossRef]
- Casagrande, E.; Recanati, F.; Melià, P. Assessing the influence of vegetation on the water budget of tropical areas. IFAC-PapersOnLine 2018, 51, 1–6. [Google Scholar] [CrossRef]
- Jones, H. Radiation. In Plants and Microclimate: A Quantitative Approach to Environmental Plant Physiology; Cambridge University Press: Cambridge, UK, 2013; pp. 9–46. [Google Scholar]
- Wullschleger, S.D.; King, A.W. Radial variation in sap velocity as a function of stem diameter and sapwood thickness in yellow-poplar trees. Tree Physiol. 2000, 20, 511–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granier, A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiol. 1987, 3, 309–320. [Google Scholar] [CrossRef]
- Granier, A.; Biron, P.; Bréda, N.; Pontailler, J.-Y.; Saugier, B. Transpiration of trees and forest stands: Short and longterm monitoring using sap flow methods. Glob. Chang. Biol. 1996, 2, 265–274. [Google Scholar] [CrossRef]
- Gonzalez García, M.A.; Paz Gonzalez, A.; Castelao Gegunde, A. La medida térmica del flujo de savia aplicada al estudio de la extracción de agua por los arboles: Revisión bibliográfica y puesta a punto del método. Investig. Agrícola Sist. Recur. For. 1995, 4, 1–16. [Google Scholar]
- Lu, P.; Urban, L.; Zhao, P. Granier’s Thermal Dissipation Probe (TDP) method for measuring sap flow in trees: Theory and practic’. Acta Bot. Sin. 2004, 46, 631–646. [Google Scholar]
- Zhang, Z.; Tian, F.; Hu, H.; Yang, P. A comparison of methods for determining field evapotranspiration: Photosynthesis system, sap flow, and eddy covariance. Hydrol. Earth Syst. Sci. 2014, 18, 1053–1072. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Garcia, C.; Andrade, J.L.; Sima, J.L.; Us-Santamarıa, R.; Jackson, P.C. Sapwood to heartwood ratio affects whole-tree water use in dry forest legume and non-legume trees. Trees 2012, 26, 1317–1330. [Google Scholar] [CrossRef]
- León Palomo, M.A. Flujo de Agua en Árboles de Una Selva Mediana Subcaducifolia en Yucatán, México. Bachelor’s Thesis, Instituto Tecnológico de Conkal, Conkal, Mexico, 2013. [Google Scholar]
- CONAGUA. Actualización de la Disponibilidad Media Anual de Agua en el Acuífero Península de Yucatán (3105), Estado de Yucatán’, Diario Oficial de la Federación. Available online: https://sigagis.conagua.gob.mx/gas1/Edos_Acuiferos_18/yucatan/DR_3105.pdf (accessed on 4 March 2020).
- Uuh-Sonda, J.M.; Figueroa-Espinoza, B.; Gutiérrez-Jurado, H.A.; Méndez-Barroso, L.A. Ecosystem productivity and evapotranspiration dynamics of a seasonally dry tropical forest of the Yucatan Peninsula. J. Geophys. Res. Biogeosci. 2019, 126, e2019JG005629. [Google Scholar] [CrossRef]
- Uuh-Sonda, J.M.; Gutiérrez-Jurado, H.A.; Figueroa-Espinoza, B.; Méndez-Barroso, L.A. On the ecohydrology of the Yucatan Peninsula: Evapotranspiration and carbon intake dynamics across an eco-climatic gradient. Hydrol. Process. 2018, 32, 2806–2828. [Google Scholar] [CrossRef]
- INEGI. Anuario Estadístico y Geográfico de Yucatán 2017; Instituto Nacional de Estadística y Geografía: Aguascalientes, México.
- Miranda, F.; Hernandez-Xolocotzi, E. Los Tipos de Vegetación de México y su Clasificación; Sociedad Botánica de México: Chapingo, México, 1963; p. 179. [Google Scholar]
- Trejo, I. Análisis de la diversidad de la selva baja caducifolia en México. In Sobre la Diversidad Biológica: El significado de las Diversidades alfa, beta y Gamma; m3m-Monografías 3ercer Milenio, vol. 4. SEA, CONABIO; Halffter, G., Soberón, J., Koleff, P., Melic., A., Eds.; Grupo DIVERSITAS & CONACYT: Zaragoza, España, 2005; pp. 111–122. [Google Scholar]
- Canche Escamilla, G. Uso de Biomasa Leñosa Como Biocomustible Sólido Para la Generación de Energía Eléctrica en Zonas Rurales; Informe Final Fondo sectorial CONACYT-Secretaría de Energía-Sustentabilidad Energética: Proyecto, México, 2019. [Google Scholar]
- Smith, D.M.; Allen, S.J. Measurement of sap flow in plant stems. J. Exp. Bot. 1996, 47, 1833–1844. [Google Scholar] [CrossRef] [Green Version]
- Meinzer, F.C.; Goldstein, G.; Andrade, J.L. Regulation of water flux through tropical forest canopy trees: Do universal rules apply? Tree Physiol. 2001, 21, 19–26. [Google Scholar] [CrossRef]
- Bucci, S.J.; Goldstein, G.; Meinzer, F.C.; Scholz, F.G.; Franco, A.C.; Bustamante, M. Functional convergence in hydraulic architecture and water relations of tropical savanna trees: From leaf to whole plant. Tree Physiol. 2004, 24, 891–899. [Google Scholar] [CrossRef] [Green Version]
- Andrade, J.L.; Meinzer, F.C.; Goldstein, G.; Schnitzer, S.A. Water uptake and transport in lianas co-occurring trees of a seasonally dry tropical forest. Trees 2005, 19, 282–289. [Google Scholar] [CrossRef]
- Bucci, S.J.; Scholz, F.G.; Goldstein, G.; Hoffmann, W.A.; Meinzer, F.C.; Franco, A.C.; Giambelluca, T.; Miralles-Wilhelm, F. Controls on stand transpiration and soil water utilization along a tree density gradient in a Neotropical savanna. Agric. For. Meteorol. 2008, 148, 839–849. [Google Scholar] [CrossRef]
- CONAFOR. Innovación Forest@l-Glosario. Available online: http://www.conafor.gob.mx/innovacion_forestal/?page_id=436 (accessed on 26 April 2021).
- Cochard, H.; Coll, L.; Le Roux, X.; Améglio, T. Unraveling the Effects of Plant Hydraulics on Stomatal Closure during Water Stress in Walnut. Plant Physiol. 2002, 128, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Dawson, T.E. Determining water use by trees and forests from isotopic, energy balance and transpiration analyses: The roles of tree size and hydraulic lift. Tree Physiol. 1996, 16, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-López, R.; Ojeda-Bustamante, W.; López Andrade, A.P.; Catalán-Valencia, E.A. Heat Pulse Method and Sap Flow for Measuring Transpiration in Cacao. Rev. Chapingo-Ser. Zonas Áridas 2013, 12, 85–96. [Google Scholar] [CrossRef]
- Vertessy, R.A.; Benyon, R.G.; O´Sullivan, S.K.; Gribben, P.R. Relationships between stem diameter, sapwood area, leaf area and transpiration in a young mountain ash forest. Tree Physiol. 1995, 15, 559–567. [Google Scholar] [CrossRef]
- Vertessy, R.A.; Hatton, T.J.; Reece, P.; Sullivan, S.K.O.; Benyon, R.G. Estimating stand water use of large mountain ash trees and validation of the sap flow measurement technique. Tree Physiol. 1997, 19, 747–756. [Google Scholar] [CrossRef] [Green Version]
- Loustau, D.; Berbigier, P.; Roumagnac, P.; Arruda-Pacheco, C.; David, J.C.; Ferreira, M.I.; Pereira, J.S.; Tavares, R. Transpiration of a 64-year-old maritime pine stand in Portugal. Oecologia 1996, 107, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Saugier, B.; Granier, A.; Pontailler, J.Y.; Dufrene, E.; Baldocchi, D.D. Transpiration of a boreal pine forest measured by branch bag, sap flow and micrometeorological methods. Tree Physiol. 1997, 17, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Heilman, J.L.; Litvak, M.E.; McInnes, K.J.; Kjelgaard, J.F.; Kamps, R.H.; Schwinning, S. Water-storage capacity controls energy partitioning and water use in karst ecosystems on the Edwards Plateau, Texas. Ecohydrology 2014, 7, 127–138. [Google Scholar] [CrossRef]
- Green, S.R.; McNaughton, K.G.; Clothier, B.E. Observations of night-time water use in kiwifruit vines and apple trees. Agric. For. Meteorol. 1989, 48, 251–261. [Google Scholar] [CrossRef]
- Valdez-Hernández, M.; Andrade, J.L.; Jackson, P.C.; Rebolledo-Vieyra, M. Phenology of five tree species of a tropical dry forest in Yucatan, Mexico: Effects of environmental and physiological factors. Plant Soil 2010, 329, 155–171. [Google Scholar] [CrossRef]
- Goldstein, G.; Andrade, J.L.; Meinzer, F.C.; Holbrook, N.M.; Cavelier, J.; Jackson, P.; Celis, A. Stem water storage and diurnal patterns of water use in tropical forest canopy trees. Plant Cell Environ. 1998, 21, 397–406. [Google Scholar] [CrossRef]
- Werden, L.K.; Waring, B.G.; Smith-Martin, C.M.; Powers, J.S. Tropical dry forest trees and lianas differ in leaf economic spectrum traits but have overlapping water-use strategies. Tree Physiol. 2018, 38, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Servicio Meteorológico Nacional. Monitor de Sequía. Available online: https://smn.conagua.gob.mx/es/climatologia/monitor-de-sequia/monitor-de-sequia-en-mexico (accessed on 20 February 2020).
- Niu, F.; Röll, A.; Meijide, A.; Hölscher, D. Rubber tree transpiration in the lowlands of Sumatra. Ecohydrology 2017, 10, e1882. [Google Scholar] [CrossRef]
- Tan, P.Y.; Wong, N.H.; Tan, C.L.; Jusuf, S.K.; Schmiele, K.; Chiam, Z.Q. Transpiration and cooling potential of tropical urban trees from different native habitats. Sci. Total Environ. 2020, 705, 135764. [Google Scholar] [CrossRef]
- Running, S.; Mu, Q.; Zhao, M. MOD16A3 MODIS/Terra Net Evapotranspiration Yearly L4 Global 500m SIN Grid V006 [Data Set]. NASA EOSDIS Land Processes DAAC. Available online: https://doi.org/10.5067/MODIS/MOD16A3.006 (accessed on 10 July 2021).
- Baldocchi, D.D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: Past, present and future. Glob. Chang. Biology 2003, 9, 479–492. [Google Scholar] [CrossRef] [Green Version]
- Mu, Q.; Heinsch, F.A.; Zhao, M.; Running, S.W. Development of a global evapotranspiration algorithm based on MODIS and global meteorology data. Remote Sens. Environ. 2007, 111, 519–536. [Google Scholar] [CrossRef]
- Mu, Q.; Zhao, M.; Running, S.W. Improvements to a MODIS global terrestrial evapotranspiration algorithm. Remote Sens. Environ. 2011, 115, 1781–1800. [Google Scholar] [CrossRef]
- Terminel, M.L.V.; Yépez, E.A.; Tarín, T.; Zazueta, C.A.R.; Payan, J.G.; Rodríguez, J.C.; Watts, C.J.; Vivoni, E.R. Understory contribution to water vapor and CO2 fluxes from a subtropical shrubland in northwestern Mexico. Tecnol. Cienc. Agua 2020, 11, 130–170. [Google Scholar] [CrossRef]
- Iida, S.I.; Shimizu, T.; Tamai, K.; Kabeya, N.; Shimizu, A.; Ito, E.; Ohnuki, Y.; Chann, S.; Levia, D.F. Evapotranspiration from the understory of a tropical dry deciduous forest in Cambodia. Agric. For. Meteorol. 2020, 295, 108170. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Family | ndry | nrainy | IVI (%) |
|---|---|---|---|---|
| Caesalpinia sp. Bursera simaruba (L.) Sarg. Lysiloma latisiliquum (L.) Benth. | Fabaceae | 1 | – | – |
| Burseraceae | 3 | 3 | 49.62 | |
| Fabaceae | 2 | 3 | 35.41 | |
| Gymnopodium floribundum Rolfe Caesalpinia gaumeri Greenm. | Polygonaceae | 3 | 3 | 25.4 |
| Fabaceae | 2 | 2 | 22.98 | |
| Havardia albicans (Kunth) Britton & Rose Neea choriophylla Standl. Piscidia piscipula (L.) Sarg. Cordia gerascanthus L. | Fabaceae | 2 | 2 | 15.33 |
| Nyctiginaceae | 2 | 1 | 11.96 | |
| Fabaceae | 1 | 1 | 10.12 | |
| Cordiaceae | 1 | 1 | 9.38 | |
| Plumeria obtusa L. Erythroxylum rotundifolium Lunan | Apocynaceae | 2 | 2 | 7.49 |
| Erythroxylaceae | 1 | 1 | 5.17 | |
| TOTAL | 20 | 19 | 192.86 |
| Month | Temperature (°C) | Relative Humidity (RH) (%) | Maximum Solar Radiation (W m–2) | Vapor Pressure Deficit (kPa) |
|---|---|---|---|---|
| March | 24.7 ± 2.6 (a) | 75.5 ± 12.3 (a) | 1045.3 | 0.68 ± 0.41 (a) |
| April | 26.2 ± 2.9 (b) | 74.2 ± 13.5 (a) | 1082.0 | 0.76 ± 0.47 (b) |
| September | 27.4 ± 1.8 (c) | 84.4 ± 7.2 (b) | 1041.1 | 0.47 ± 0.24 (c) |
| October | 27.5 ± 2.0 (c) | 84.1 ± 8.7 (b) | 993.5 | 0.49 ± 0.30 (c) |
| ANOVA | F = 202.2 p < 0.0001 | F = 167.6 p < 0.0001 | F = 2.4799 p = 0.0594 | F = 97.006 p < 0.0001 |
| Season | DBH (cm) | TDBH (kg d-1) | N (Cluster) | TCluster (kg d–1) | T (kg m–2 d–1) |
|---|---|---|---|---|---|
| Late dry season | 5 – 8.5 | 2.55 ± 1.09 | 86 | 219.30 ± 93.74 | 0.100 ± 0.043 |
| 8.6 – 18.8 | 8.44 ± 4.94 | 102 | 860.88 ± 503.88 | 0.391 ± 0.229 | |
| 18.9 – 29 | 14.5 ± 5.51 | 25 | 362.50 ± 137.75 | 0.165 ± 0.063 | |
| >29 | 38.5 ± 2.2 | 2 | 77 ± 4.4 | 0.035 ± 0.020 | |
| 215 | 1519.68 ± 739.77 | 0.691 ± 0.336 | |||
| Late rainy season | 5 – 8.5 | 9.72 ± 0.53 | 86 | 835.92 ± 45.58 | 0.380 ± 0.021 |
| 8.6 – 18.8 | 14.5 ± 5.51 | 102 | 1479 ± 562.02 | 0.672 ± 0.255 | |
| 18.9 – 29 | 19.26 ± 6.83 | 25 | 481.50 ± 170.75 | 0.219 ± 0.078 | |
| >29 | 24.7 ± 4.3 | 2 | 49.40 ± 8.6 | 0.022 ± 0.004 | |
| 215 | 2845.82 ± 784.95 | 1.294 ± 0.350 |
| Season | MODIS–Estimated Daily Average ET (kg m–2 d–1) | Daily Average T/ET Allometric Relationships | Daily Average T/ETDBH Categories | Daily Average T/ET Stand Level | Seasonal Precipitation (mm) |
|---|---|---|---|---|---|
| Late dry season | 1.35 ± 0.24 | 0.48 | 0.51 | 0.42 | 111 |
| Late rainy season | 3.36 ± 0.35 | 0.34 | 0.38 | 0.20 | 248 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salas-Acosta, E.R.; Andrade, J.L.; Perera-Burgos, J.A.; Us-Santamaría, R.; Figueroa-Espinoza, B.; Uuh-Sonda, J.M.; Cejudo, E. Transpiration of a Tropical Dry Deciduous Forest in Yucatan, Mexico. Atmosphere 2022, 13, 271. https://doi.org/10.3390/atmos13020271
Salas-Acosta ER, Andrade JL, Perera-Burgos JA, Us-Santamaría R, Figueroa-Espinoza B, Uuh-Sonda JM, Cejudo E. Transpiration of a Tropical Dry Deciduous Forest in Yucatan, Mexico. Atmosphere. 2022; 13(2):271. https://doi.org/10.3390/atmos13020271
Chicago/Turabian StyleSalas-Acosta, Evelyn Raquel, José Luis Andrade, Jorge Adrián Perera-Burgos, Roberth Us-Santamaría, Bernardo Figueroa-Espinoza, Jorge M. Uuh-Sonda, and Eduardo Cejudo. 2022. "Transpiration of a Tropical Dry Deciduous Forest in Yucatan, Mexico" Atmosphere 13, no. 2: 271. https://doi.org/10.3390/atmos13020271
APA StyleSalas-Acosta, E. R., Andrade, J. L., Perera-Burgos, J. A., Us-Santamaría, R., Figueroa-Espinoza, B., Uuh-Sonda, J. M., & Cejudo, E. (2022). Transpiration of a Tropical Dry Deciduous Forest in Yucatan, Mexico. Atmosphere, 13(2), 271. https://doi.org/10.3390/atmos13020271