An Increase of Abundance and Transcriptional Activity for Acinetobacter junii Post Wastewater Treatment

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Wastewater Sampling
2.2. Tangential Flow Filtration
2.3. Nucleic Acid Extraction and Metagenomics Sequence Analysis
2.4. cDNA Synthesis
2.5. RT-qPCR
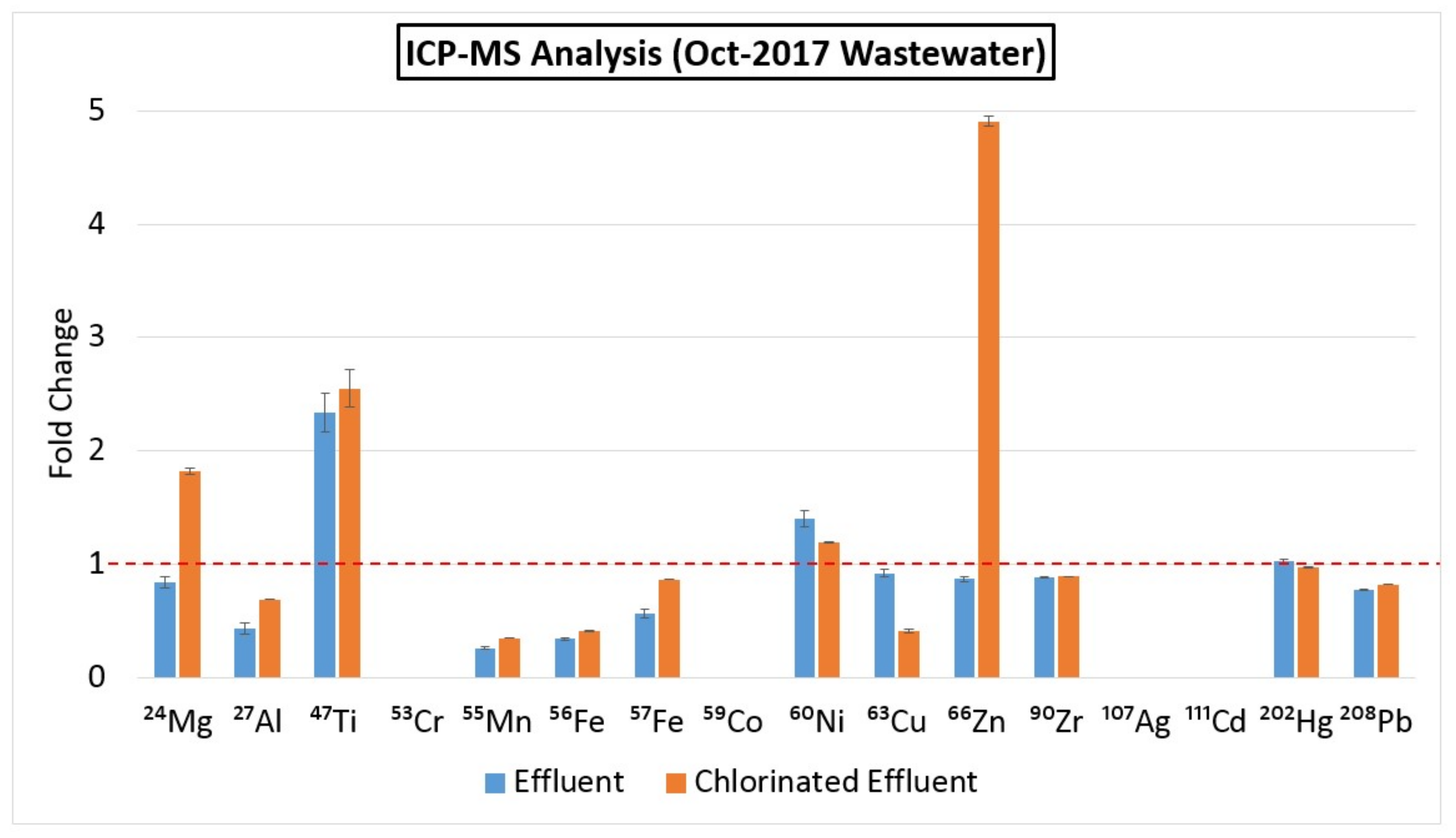
2.6. ICP-MS
3. Results
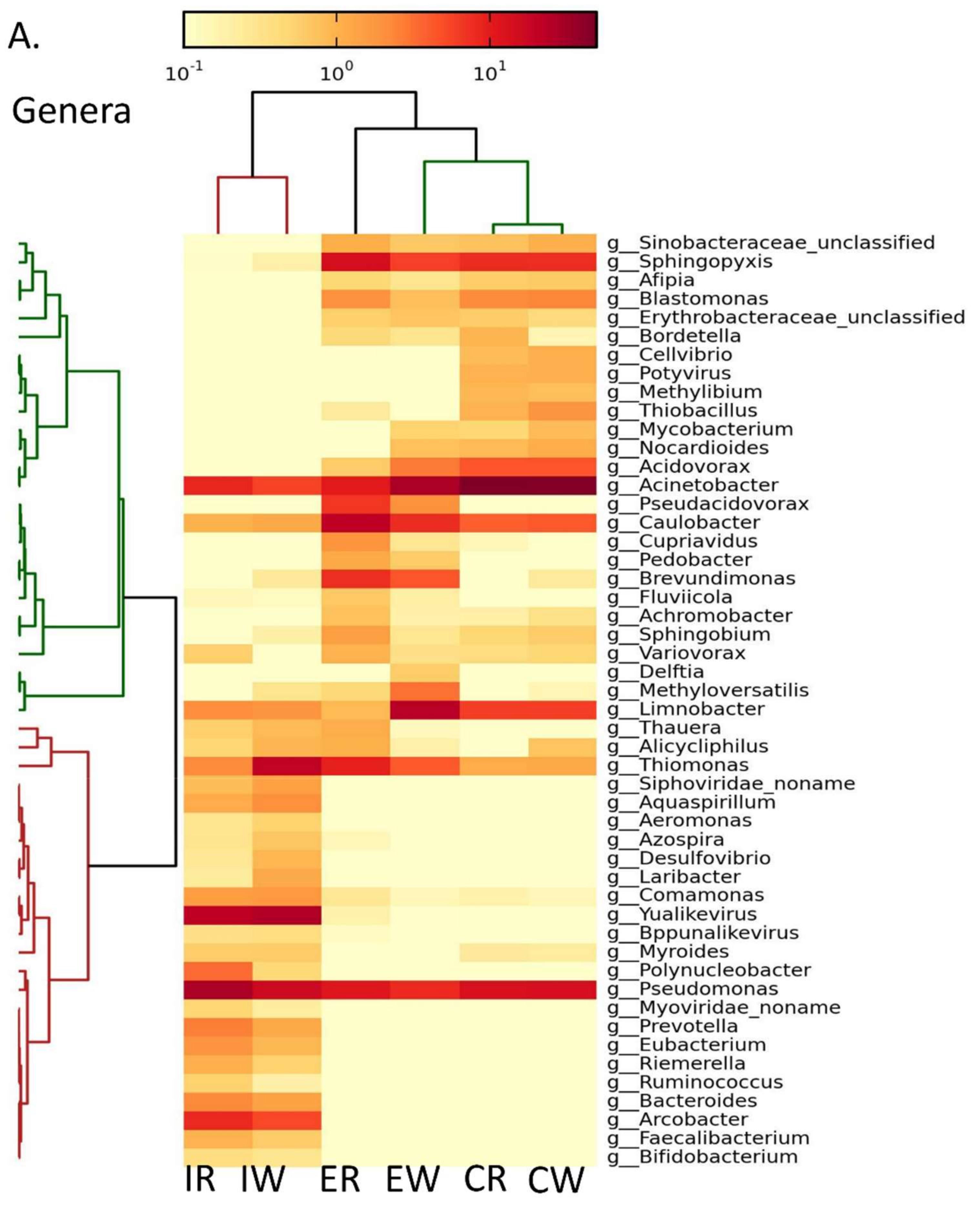
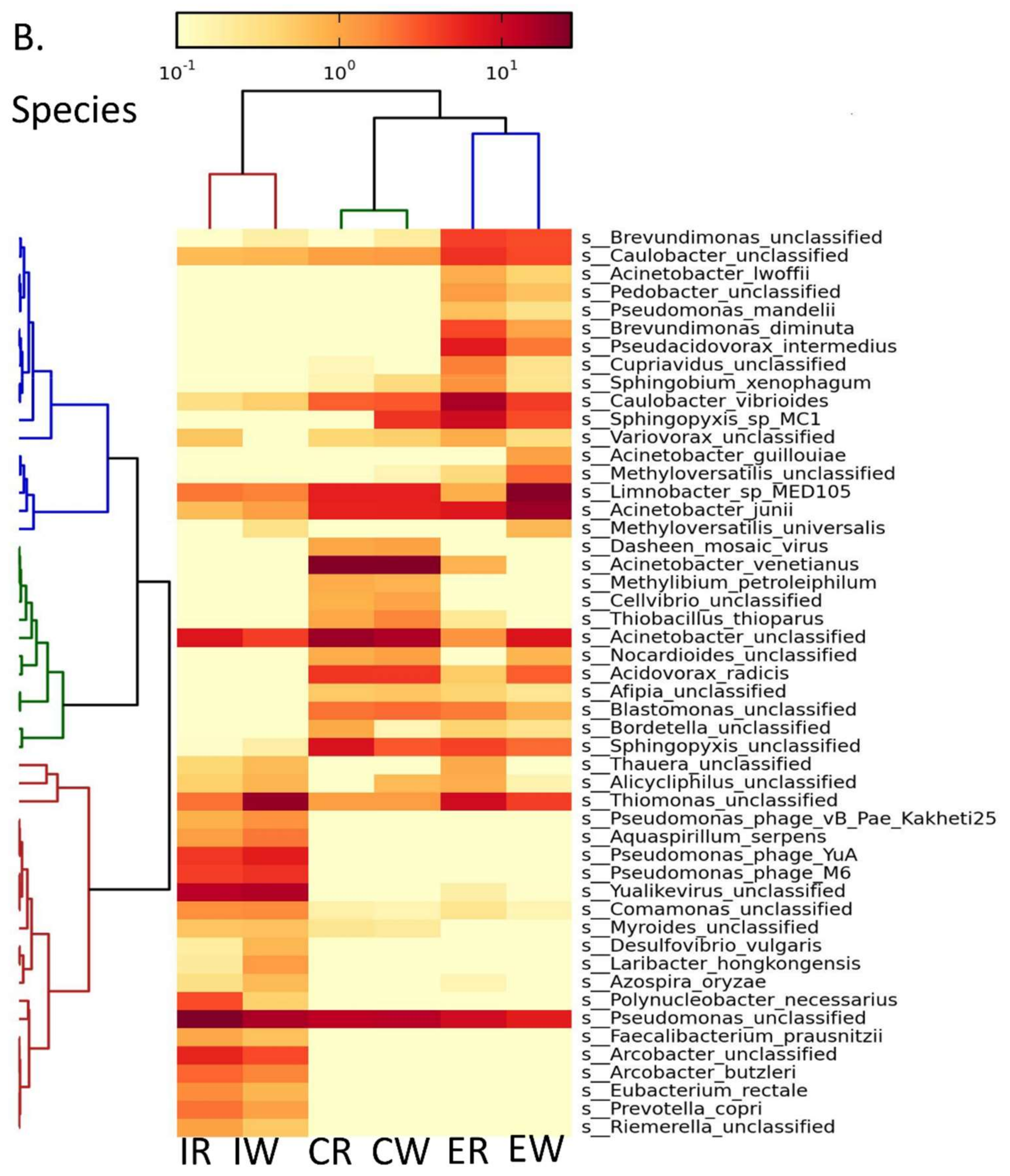
3.1. Metagenomics Analysis Revealed Enrichment of Acinetobacter spp. Post Wastewater Treatment
3.2. Gene Expression Analysis Revealed an Increase in Gene Expression of Acinetobacter junii Post MBR and Chlorination
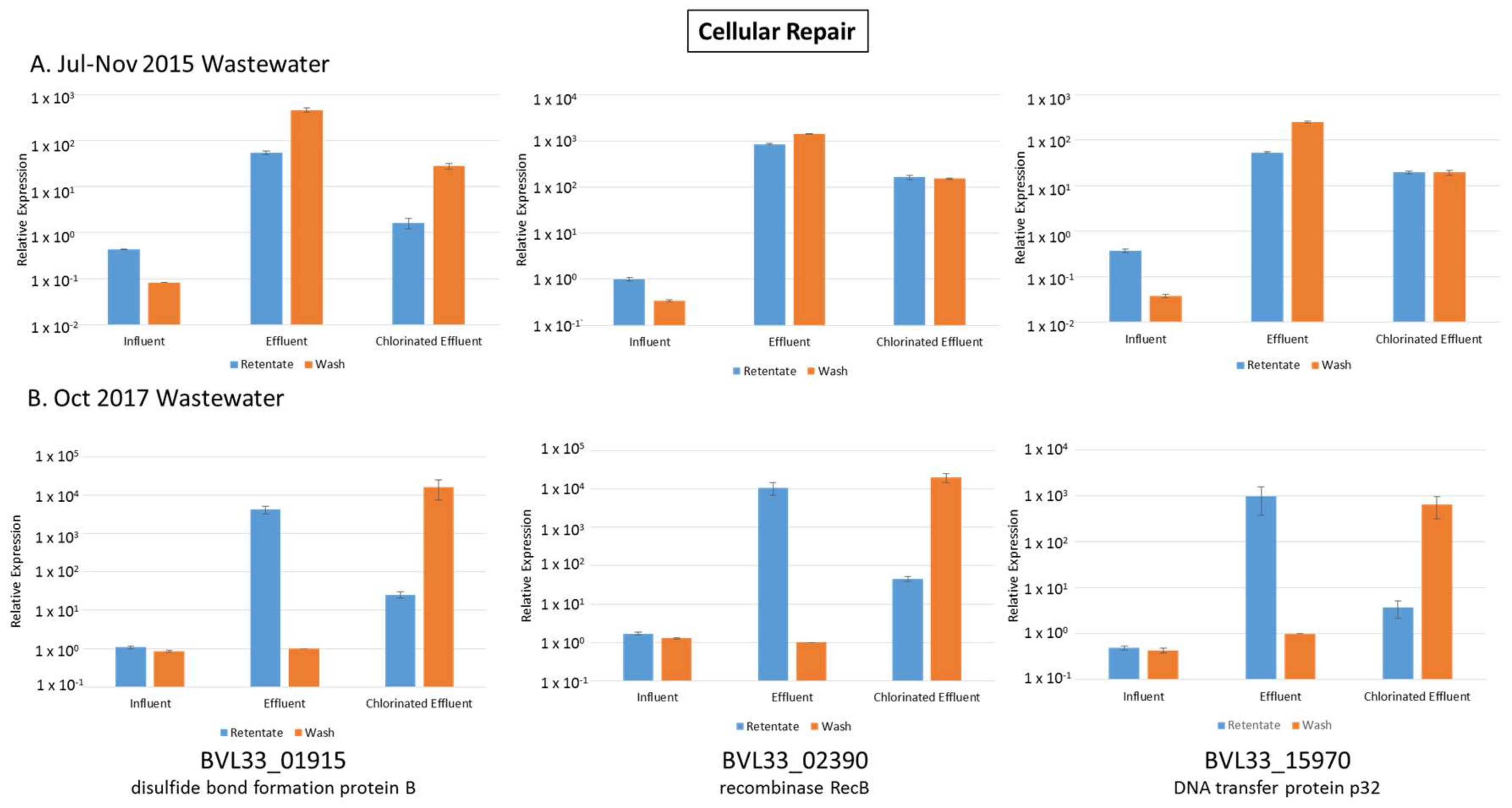
3.3. Cellular Repair Genes Upregulated in Both Effluent and Chlorinated Effluent
3.4. Metabolism Genes Upregulated in Both Effluent and Chlorinated Effluent
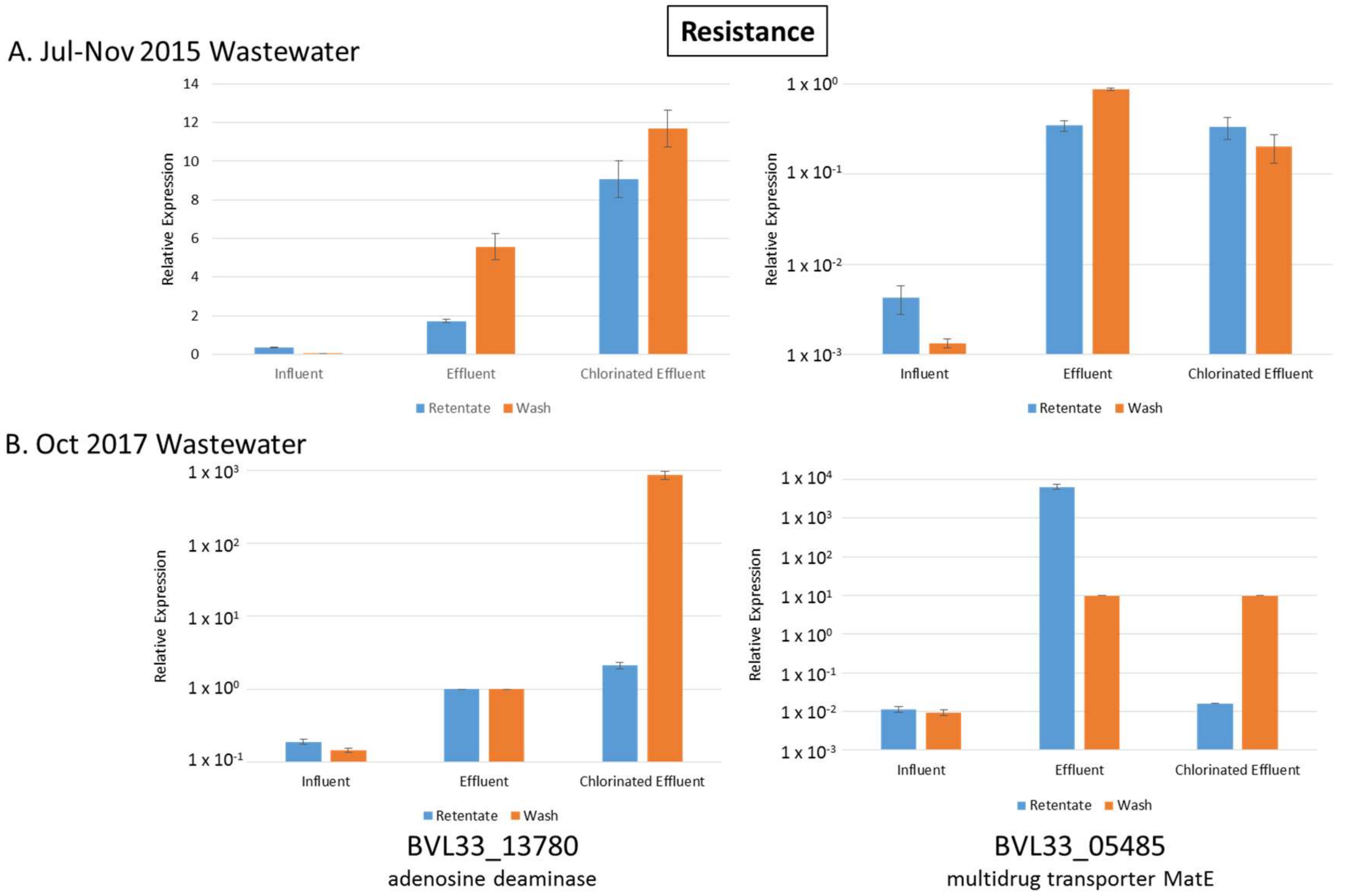
3.5. Resistance Genes Upregulated in Both Effluent and Chlorinated Effluent
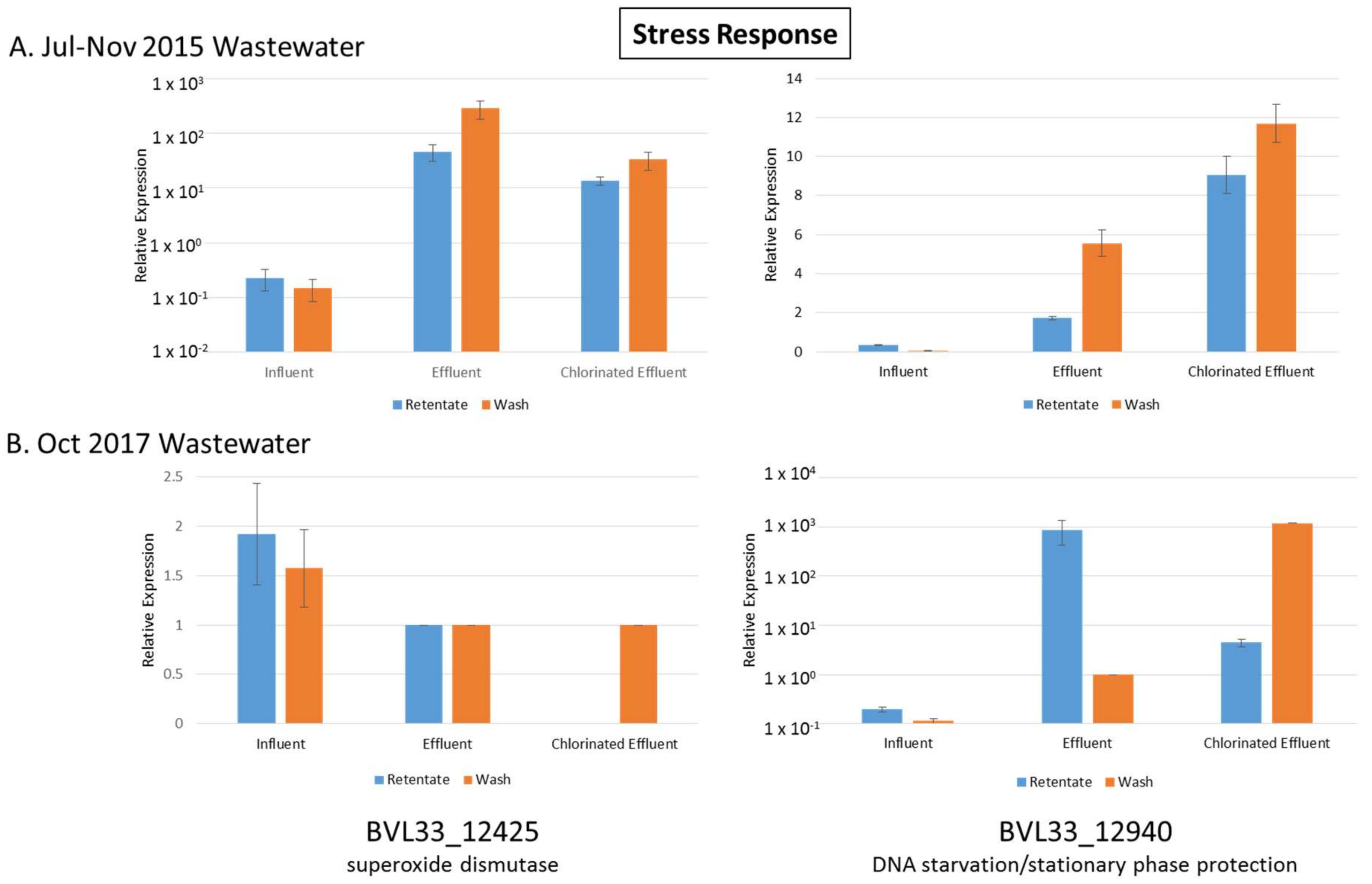
3.6. Stress Response Genes Upregulated in Both Effluent and Chlorinated Effluent
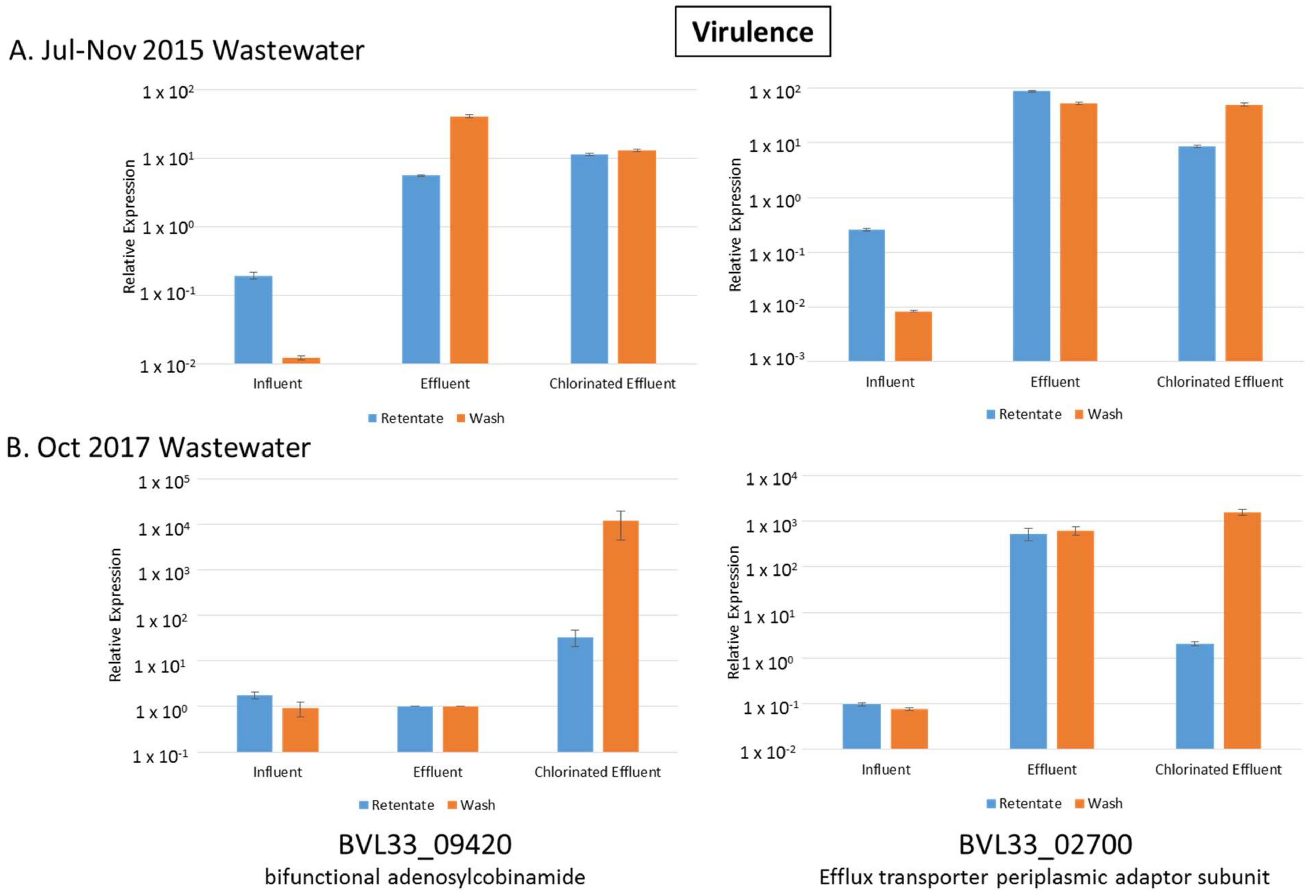
3.7. Virulence, Disease and Defense Genes Upregulated in Both Effluent and Chlorinated Effluent
3.8. Genes Upregulated Exclusively in the Effluent
3.9. Genes Upregulated Exclusively in the Chlorinated Effluent
3.10. Differential Upregulation of Genes in Retentate and Wash Samples
3.11. Validation of RNA-seq Data with RT-qPCR
3.12. Metal ion Concentrations in March 2016 and October 2017 Wastewater Samples
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shomar, B. Water Resources, Water Quality and Human Health in Regions of Extreme Stress: Middle East. J. Earth Sci. Clim. Chang. 2013, 4, 1–12. [Google Scholar] [CrossRef]
- Al-Jasser, A.O. Saudi wastewater reuse standards for agricultural irrigation: Riyadh treatment plants effluent compliance. J. King Saud Univ. Eng. Sci. 2011, 23, 1–8. [Google Scholar] [CrossRef]
- Webb, A.L.; Taboada, E.N.; Selinger, L.B.; Boras, V.F.; Inglis, G.D. Efficacy of wastewater treatment on Arcobacter butzleri density and strain diversity. Water Res. 2016, 105, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Al-Jassim, N.; Ansari, M.I.; Harb, M.; Hong, P.-Y. Removal of bacterial contaminants and antibiotic resistance genes by conventional wastewater treatment processes in Saudi Arabia: Is the treated wastewater safe to reuse for agricultural irrigation? Water Res. 2015, 73, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Sui, Q.; Jiang, C.; Yu, D.; Chen, M.; Zhang, J.; Wang, Y.; Wei, Y. Performance of a sequencing-batch membrane bioreactor (SMBR) with an automatic control strategy treating high-strength swine wastewater. J. Hazard. Mater. 2018, 342, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Dolfing, J.; Xie, B. Bacterial perspectives on the dissemination of antibiotic resistance genes in domestic wastewater bio-treatment systems: Beneficiary to victim. Appl. Microbiol. Biotechnol. 2018, 102, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Lekunberri, I.; Balcazar, J.L.; Borrego, C.M. Metagenomic exploration reveals a marked change in the river resistome and mobilome after treated wastewater discharges. Environ. Pollut. 2017, 234, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Hu, S.; Zhang, Y.; Zhao, X.; Pan, W. Influence of sewage treatment plant effluent discharge into multipurpose river on its water quality: A quantitative health risk assessment of Cryptosporidium and Giardia. Environ. Pollut. 2017, 233, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Ajonina, C.; Buzie, C.; Rubiandini, R.H.; Otterpohl, R. Microbial pathogens in wastewater treatment plants (WWTP) in Hamburg. J. Toxicol. Environ. Health Part A 2015, 78, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Naidoo, S.; Olaniran, A.O. Treated wastewater effluent as a source of microbial pollution of surface water resources. Int. J. Environ. Res. Public Health 2014, 11, 249–270. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Zhang, T. Detecting human bacterial pathogens in wastewater treatment plants by a high-throughput shotgun sequencing technique. Environ. Sci. Technol. 2013, 47, 5433–5441. [Google Scholar] [CrossRef] [PubMed]
- Judd, S. The status of membrane bioreactor technology. Trends Biotechnol. 2008, 26, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Iorhemen, O.T.; Hamza, R.A.; Tay, J.H. Membrane bioreactor (MBR) technology for wastewater treatment and reclamation: Membrane fouling. Membranes 2016, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Pollice, A.; Laera, G.; Saturno, D.; Giordano, C.; Sandulli, R. Optimal sludge retention time for a bench scale MBR treating municipal sewage. Water Sci. Technol. 2008, 57, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Anastasi, E.M.; Wohlsen, T.D.; Stratton, H.M.; Katouli, M. Survival of Escherichia coli in two sewage treatment plants using UV irradiation and chlorination for disinfection. Water Res. 2013, 47, 6670–6679. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.S. Disinfectant resistance mechanisms, cross-resistance, and co-resistance. Int. Biodeterior. Biodegrad. 2003, 51, 271–276. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, A.Z.; He, M.; Li, D.; Chen, J. Subinhibitory concentrations of disinfectants promote the horizontal transfer of multidrug resistance genes within and across genera. Environ. Sci. Technol. 2017, 51, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Reactive Oxygen Species (ROS). 2015. Available online: http://www.biology-pages.info/R/ROS.html (accessed on 10 January 2018).
- Jumat, M.; Hasan, N.; Subramanian, P.; Heberling, C.; Colwell, R.; Hong, P.-Y. Membrane bioreactor-based wastewater treatment plant in Saudi Arabia: Reduction of viral diversity, load, and infectious capacity. Water 2017, 9, 534. [Google Scholar] [CrossRef]
- Harb, M.; Hong, P.-Y. Molecular-based detection of potentially pathogenic bacteria in membrane bioreactor (MBR) systems treating municipal wastewater: A case study. Environ. Sci. Pollut. Res. 2017, 24, 5370–5380. [Google Scholar] [CrossRef] [PubMed]
- Hrenovic, J.; Goic-Barisic, I.; Kazazic, S.; Kovacic, A.; Ganjto, M.; Tonkic, M. Carbapenem-resistant isolates of Acinetobacter baumannii in a municipal wastewater treatment plant, Croatia, 2014. Euro Surveill. 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Marrs, C.F.; Simon, C.; Xi, C. Wastewater treatment contributes to selective increase of antibiotic resistance among Acinetobacter spp. Sci. Total Environ. 2009, 407, 3702–3706. [Google Scholar] [CrossRef] [PubMed]
- Oerther, D.B.; Pernthaler, J.; Schramm, A.; Amann, R.; Raskin, L. Monitoring Precursor 16S rRNAs of Acinetobacter spp. in Activated Sludge Wastewater Treatment Systems. Appl. Environ. Microbiol. 2000, 66, 2154–2165. [Google Scholar] [CrossRef] [PubMed]
- Joly-Guillou, M.L. Clinical impact and pathogenicity of Acinetobacter. Clin. Microbiol. Infect. 2005, 11, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Linde, H.J.; Hahn, J.; Holler, E.; Reischl, U.; Lehn, N. Septicemia due to Acinetobacter junii. J. Clin. Microbiol. 2002, 40, 2696–2697. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.T.; Lee, Y.T.; Huang, L.J.; Chen, T.L.; Yu, K.W.; Fung, C.P.; Cho, W.L.; Liu, C.Y. Clinical characteristics of patients with Acinetobacter junii infection. J. Microbiol. Immunol. Infect. 2009, 42, 47–53. [Google Scholar] [PubMed]
- Cayo, R.; Yanez San Segundo, L.; Perez del Molino Bernal, I.C.; Garcia de la Fuente, C.; Bermudez Rodriguez, M.A.; Calvo, J.; Martinez-Martinez, L. Bloodstream infection caused by Acinetobacter junii in a patient with acute lymphoblastic leukaemia after allogenic haematopoietic cell transplantation. J. Med. Microbiol. 2011, 60 Pt 3, 375–377. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.M.; Tsimring, L.; Hasty, J. Inter-species population dynamics enhance microbial horizontal gene transfer and spread of antibiotic resistance. eLife 2017, 6, E25950. [Google Scholar] [CrossRef] [PubMed]
- Al Atrouni, A.; Joly-Guillou, M.-L.; Hamze, M.; Kempf, M. Reservoirs of non-baumannii Acinetobacter species. Front. Microbiol. 2016, 7, 49. [Google Scholar] [CrossRef] [PubMed]
- Wiedmann-al-Ahmad, M.; Tichy, H.V.; Schön, G. Characterization of Acinetobacter type strains and isolates obtained from wastewater treatment plants by PCR fingerprinting. Appl. Environ. Microbiol. 1994, 60, 4066–4071. [Google Scholar] [PubMed]
- Doughari, H.J.; Ndakidemi, P.A.; Human, I.S.; Benade, S. The ecology, biology and pathogenesis of Acinetobacter spp.: An overview. Microbes Environ. 2011, 26, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Montaña, S.; Cittadini, R.; del Castillo, M.; Uong, S.; Lazzaro, T.; Almuzara, M.; Barberis, C.; Vay, C.; Ramírez, M.S. Presence of New Delhi metallo-β-lactamase gene (NDM-1) in a clinical isolate of Acinetobacter junii in Argentina. New Microbes New Infect. 2016, 11, 43–44. [Google Scholar] [CrossRef] [PubMed]
- Guardabassi, L.; Dalsgaard, A.; Olsen, J.E. Phenotypic characterization and antibiotic resistance of Acinetobacter spp. isolated from aquatic sources. J. Appl. Microbiol. 1999, 87, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Molina-Romero, D.; Baez, A.; Quintero-Hernández, V.; Castañeda-Lucio, M.; Fuentes-Ramírez, L.E.; Bustillos-Cristales, M.d.R.; Rodríguez-Andrade, O.; Morales-García, Y.E.; Munive, A.; Muñoz-Rojas, J. Compatible bacterial mixture, tolerant to desiccation, improves maize plant growth. PLoS ONE 2017, 12, E0187913. [Google Scholar] [CrossRef] [PubMed]
- Bastian, R.; Murray, D. 2012 Guidelines for Water Reuse; EPA Office of Research and Development: Washington, DC, USA, 2012.
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Truong, D.T.; Franzosa, E.A.; Tickle, T.L.; Scholz, M.; Weingart, G.; Pasolli, E.; Tett, A.; Huttenhower, C.; Segata, N. MetaPhlAn2 for enhanced metagenomic taxonomic profiling. Nat. Methods 2015, 12, 902. [Google Scholar] [CrossRef] [PubMed]
- Baggerly, K.A.; Deng, L.; Morris, J.S.; Aldaz, C.M. Differential expression in SAGE: Accounting for normal between-library variation. Bioinformatics 2003, 19, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Broniek, G.; Langwińska-Wośko, E.; Szaflik, J.; Wróblewska, M. Acinetobacter junii as an aetiological agent of corneal ulcer. Infection 2014, 42, 1051–1053. [Google Scholar] [CrossRef] [PubMed]
- Cydzik-Kwiatkowska, A.; Zielińska, M. Bacterial communities in full-scale wastewater treatment systems. World J. Microbiol. Biotechnol. 2016, 32, 66. [Google Scholar] [CrossRef] [PubMed]
- Spies, M.; Kowalczykowski, S.C. Homologous recombination by RecBCD and RecF pathways. In The Bacterial Chromosome 2005; ASM Press: Washington, DC, USA, 2005; pp. 389–403. [Google Scholar]
- Dillingham, M.S.; Kowalczykowski, S.C. RecBCD enzyme and the repair of double-stranded DNA breaks. Microbiol. Mol. Biol. Rev. MMBR 2008, 72, 642–671. [Google Scholar] [CrossRef] [PubMed]
- Kickstein, E.; Harms, K.; Wackernagel, W. Deletions of recBCD or recD influence genetic transformation differently and are lethal together with a recJ deletion in Acinetobacter baylyi. Microbiology 2007, 153 Pt 7, 2259–2270. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Finkel, S.E. Dps protects cells against multiple stresses during stationary phase. J. Bacteriol. 2004, 186, 4192–4198. [Google Scholar] [CrossRef] [PubMed]
- Boubrik, F.; Rouviere-Yaniv, J. Increased sensitivity to gamma irradiation in bacteria lacking protein, H.U. Proc. Natl. Acad. Sci. USA 1995, 92, 3958–3962. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Waters, R. Escherichia coli strains lacking protein HU are UV sensitive due to a role for HU in homologous recombination. J. Bacteriol. 1998, 180, 3750–3756. [Google Scholar] [PubMed]
- Miyabe, I.; Zhang, Q.M.; Kano, Y.; Yonei, S. Histone-like protein HU is required for recA gene-dependent DNA repair and SOS induction pathways in UV-irradiated Escherichia coli. Int. J. Radiat. Biol. 2000, 76, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Knowles, T.J.; Scott-Tucker, A.; Overduin, M.; Henderson, I.R. Membrane protein architects: The role of the BAM complex in outer membrane protein assembly. Nat. Rev. Microbiol. 2009, 7, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Knowles, T.J.; Browning, D.F.; Jeeves, M.; Maderbocus, R.; Rajesh, S.; Sridhar, P.; Manoli, E.; Emery, D.; Sommer, U.; Spencer, A.; et al. Structure and function of BamE within the outer membrane and the beta-barrel assembly machine. EMBO Rep. 2011, 12, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, N.; Falcone, B.; Kahne, D.; Silhavy, T.J. Chemical conditionality: A genetic strategy to probe organelle assembly. Cell 2005, 121, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Denks, K.; Vogt, A.; Sachelaru, I.; Petriman, N.-A.; Kudva, R.; Koch, H.-G. The Sec translocon mediated protein transport in prokaryotes and eukaryotes. Mol. Membr. Biol. 2014, 31, 58–84. [Google Scholar] [CrossRef] [PubMed]
- Maillard, A.P.; Chan, K.K.Y.; Duong, F. Preprotein translocation through the Sec translocon in bacteria. In Protein Movement Across Membranes; Springer: Boston, MA, USA, 2005; pp. 19–32. [Google Scholar]
- Fekkes, P.; Driessen, A.J.M. Protein targeting to the bacterial cytoplasmic membrane. Microbiol. Mol. Biol. Rev. 1999, 63, 161–173. [Google Scholar] [PubMed]
- Hunt, M.C.; Siponen, M.I.; Alexson, S.E.H. The emerging role of acyl-CoA thioesterases and acyltransferases in regulating peroxisomal lipid metabolism. Biochim. Biophys. Acta (BBA) 2012, 1822, 1397–1410. [Google Scholar] [CrossRef] [PubMed]
- Fixter, L.M.; Nagi, M.N.; Mccormack, J.G.; Fewson, C.A. Structure, distribution and function of wax esters in Acinetobacter calcoaceticus. Microbiology 1986, 132, 3147–3157. [Google Scholar] [CrossRef]
- Kalscheuer, R.; Steinbuchel, A. A novel bifunctional wax ester synthase/acyl-CoA:diacylglycerol acyltransferase mediates wax ester and triacylglycerol biosynthesis in Acinetobacter calcoaceticus ADP1. J. Biol. Chem. 2003, 278, 8075–8082. [Google Scholar] [CrossRef] [PubMed]
- Ishige, T.; Tani, A.; Sakai, Y.; Kato, N. Wax ester production by bacteria. Curr. Opin. Microbiol. 2003, 6, 244–250. [Google Scholar] [CrossRef]
- Chaiyen, P.; Suadee, C.; Wilairat, P. A novel two-protein component flavoprotein hydroxylase. Eur. J. Biochem. 2001, 268, 5550–5561. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.; Sar, P. Real-time PCR based analysis of metal resistance genes in metal resistant Pseudomonas aeruginosa strain J007. J. Basic Microbiol. 2016, 56, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Monsieurs, P.; Moors, H.; Van Houdt, R.; Janssen, P.J.; Janssen, A.; Coninx, I.; Mergeay, M.; Leys, N. Heavy metal resistance in Cupriavidus metallidurans CH34 is governed by an intricate transcriptional network. Biometals 2011, 24, 1133–1151. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Jia, S.; Zhang, X.X.; Zhang, T.; Cheng, S.; Li, A. Metagenomic insights into chlorination effects on microbial antibiotic resistance in drinking water. Water Res. 2013, 47, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Karumathil, D.P.; Yin, H.-B.; Kollanoor-Johny, A.; Venkitanarayanan, K. Effect of chlorine exposure on the survival and antibiotic gene expression of multidrug resistant Acinetobacter baumannii in Water. Int. J. Environ. Res. Public Health 2014, 11, 1844–1854. [Google Scholar] [CrossRef] [PubMed]
- Kohanski, M.A.; DePristo, M.A.; Collins, J.J. Sublethal antibiotic treatment leads to multidrug resistance via radical-induced mutagenesis. Mol. Cell 2010, 37, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Bogomolnaya, L.M.; Andrews, K.D.; Talamantes, M.; Maple, A.; Ragoza, Y.; Vazquez-Torres, A.; Andrews-Polymenis, H. The ABC-type efflux pump MacAB protects Salmonella enterica serovar typhimurium from oxidative stress. mBio 2013, 4, e00630-13. [Google Scholar] [CrossRef] [PubMed]
- Rumbo-Feal, S.; Gómez, M.J.; Gayoso, C.; Álvarez-Fraga, L.; Cabral, M.P.; Aransay, A.M.; Rodríguez-Ezpeleta, N.; Fullaondo, A.; Valle, J.; Tomás, M.; et al. Whole transcriptome analysis of Acinetobacter baumannii assessed by RNA-sequencing reveals different mRNA expression profiles in biofilm compared to planktonic cells. PLoS ONE 2013, 8, E72968. [Google Scholar] [CrossRef] [PubMed]
- Cray, J.A.; Bell, A.N.; Bhaganna, P.; Mswaka, A.Y.; Timson, D.J.; Hallsworth, J.E. The biology of habitat dominance; can microbes behave as weeds? Microb. Biotechnol. 2013, 6, 453–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balaban, N.Q.; Merrin, J.; Chait, R.; Kowalik, L.; Leibler, S. Bacterial persistence as a phenotypic switch. Science 2004, 305, 1622–1625. [Google Scholar] [CrossRef] [PubMed]
- Rittershaus, E.S.; Baek, S.H.; Sassetti, C.M. The normalcy of dormancy: Common themes in microbial quiescence. Cell Host Microbe 2013, 13, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, A.S.; Tang, Y.J.; Mukhopadhyay, A.; Martin, H.G.; Gin, J.; Benke, P.I.; Keasling, J.D. Study of stationary phase metabolism via isotopomer analysis of amino acids from an isolated protein. Biotechnol. Prog. 2010, 26, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Klappenbach, J.A.; Dunbar, J.M.; Schmidt, T.M. rRNA operon copy number reflects ecological strategies of bacteria. Appl. Environ. Microbiol. 2000, 66, 1328–1333. [Google Scholar] [CrossRef] [PubMed]
- Mahillon, J.; Chandler, M. Insertion sequences. Microbiol. Mol. Biol. Rev. MMBR 1998, 62, 725–774. [Google Scholar] [PubMed]
- Mugnier, P.D.; Poirel, L.; Nordmann, P. Functional analysis of insertion sequence ISAba1, responsible for genomic plasticity of Acinetobacter baumannii. J. Bacteriol. 2009, 191, 2414–2418. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.G.; Gianoulis, T.A.; Pukatzki, S.; Mekalanos, J.J.; Ornston, L.N.; Gerstein, M.; Snyder, M. New insights into Acinetobacter baumannii pathogenesis revealed by high-density pyrosequencing and transposon mutagenesis. Genes Dev. 2007, 21, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Rose, A. TnAbaR1: A novel Tn7-related transposon in Acinetobacter baumannii that contributes to the accumulation and dissemination of large repertoires of resistance genes. Biosci. Horiz. 2010, 3, 40–48. [Google Scholar] [CrossRef]
- Mussi, M.A.; Limansky, A.S.; Viale, A.M. Acquisition of resistance to carbapenems in multidrug-resistant clinical strains of Acinetobacter baumannii: Natural insertional inactivation of a gene encoding a member of a novel family of beta-barrel outer membrane proteins. Antimicrob. Agents Chemother. 2005, 49, 1432–1440. [Google Scholar] [CrossRef] [PubMed]
- Heritier, C.; Poirel, L.; Nordmann, P. Cephalosporinase over-expression resulting from insertion of ISAba1 in Acinetobacter baumannii. Clin. Microbiol. Infect. 2006, 12, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Block, A.; Alfano, J.R. Plant targets for Pseudomonas syringae type III effectors: Virulence targets or guarded decoys? Curr. Opin. Microbiol. 2011, 14, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Lee, K.C.; Kim, K.K.; Hwang, I.C.; Jang, C.; Kim, N.G.; Yeo, W.H.; Kim, B.S.; Yu, Y.M.; Ahn, J.S. Acinetobacter antiviralis sp. nov., from tobacco plant roots. J. Microbiol. Biotechnol. 2009, 19, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.S.; Jung, J.; Jeon, C.O.; Park, W. Acinetobacter oleivorans sp. nov. is capable of adhering to and growing on diesel-oil. J. Microbiol. 2011, 49, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Perez, S.; Lievens, B.; Jacquemyn, H.; Herrera, C.M. Acinetobacter nectaris sp. nov. and Acinetobacter boissieri sp. nov., isolated from floral nectar of wild Mediterranean insect-pollinated plants. Int. J. Syst. Evolut. Microbiol. 2013, 63 Pt 4, 1532–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fold Change | ||||||
|---|---|---|---|---|---|---|
| Locus Tag | Gene Product | Effluent Retentate | Effluent Wash | Chlorinated Effluent Retentate | Chlorinated Effluent Wash | Category |
| BVL33_15990 | 50S ribosomal protein L10 | 2.06 | 31.09 | 2.51 | 80.82 | Cellular Repair |
| BVL33_10015 | L-threonine dehydrogenase | 2.94 | 12.93 | 2.32 | 3.36 | Cellular Repair |
| BVL33_00375 | F0F1 ATP synthase subunit epsilon | 2.04 | 12.66 | 3.30 | 16.98 | Cellular Repair |
| BVL33_12120 | protoheme IX farnesyltransferase | 2.11 | 7.08 | 3.26 | 4.70 | Cellular Repair |
| BVL33_06310 | outer membrane protein assembly factor BamE | 2.45 | 5.89 | 2.96 | 3.42 | Cellular Repair |
| BVL33_12235 | cytochrome b | 2.71 | 5.14 | 2.45 | 4.75 | Cellular Repair |
| BVL33_02910 | bacterioferritin | 37.79 | 5.09 | 57.84 | 7.09 | Cellular Repair |
| BVL33_13450 | RNA-binding protein | 4.75 | 4.91 | 7.36 | 4.48 | Cellular Repair |
| BVL33_10260 | adenosine kinase | 3.72 | 4.74 | 3.67 | 3.52 | Cellular Repair |
| BVL33_01915 | disulfide bond formation protein B | 3.58 | 4.74 | 9.65 | 6.64 | Cellular Repair |
| BVL33_02390 | recombinase RecB | 5.06 | 4.66 | 8.33 | 6.01 | Cellular Repair |
| BVL33_10760 | NAD kinase | 2.99 | 4.37 | 4.90 | 3.95 | Cellular Repair |
| BVL33_01480 | iron-sulfur cluster insertion protein ErpA | 2.18 | 4.10 | 2.47 | 3.01 | Cellular Repair |
| BVL33_01875 | short-chain dehydrogenase | 6.64 | 3.61 | 2.28 | 3.27 | Cellular Repair |
| BVL33_14300 | gamma-glutamylcyclotransferase | 2.71 | 2.90 | 4.87 | 5.64 | Cellular Repair |
| BVL33_08930 | dehydratase | 2.25 | 2.62 | 9.67 | 4.05 | Cellular Repair |
| BVL33_15970 | DNA transfer protein p32 | 250.50 | 2.56 | 493.00 | 21.17 | Cellular Repair |
| BVL33_14275 | folate-binding protein YgfZ | 2.26 | 2.53 | 4.89 | 4.89 | Cellular Repair |
| BVL33_06815 | phosphoribosylglycinamide formyltransferase | 2.11 | 2.49 | 4.90 | 8.94 | Cellular Repair |
| BVL33_11785 | DNA-binding protein HU | 16.05 | 2.23 | 9.16 | 4.32 | Cellular Repair |
| BVL33_01730 | lipoprotein-34 precursor (NlpB) | 5.43 | 2.03 | 2.46 | 2.84 | Cellular Repair |
| BVL33_14490 | SPOR domain-containing protein | 329.60 | 2.13 | 425.30 | 3.80 | Cellular Repair & Metabolism |
| BVL33_14010 | acetyltransferase | 2.35 | 3.04 | 2.19 | 4.41 | Cellular Repair & Metabolism |
| BVL33_13315 | acyl-CoA thioesterase | 2.47 | 3.04 | 2.11 | 7.39 | Cellular Repair & Metabolism |
| BVL33_13885 | acyltransferase | 117.20 | 2.01 | 421.20 | 8.79 | Cellular Repair & Metabolism |
| BVL33_03425 | 30S ribosomal protein S17 | 2.27 | 46.72 | 3.25 | 7.78 | Metabolism |
| BVL33_03420 | 50S ribosomal protein L29 | 2.32 | 39.53 | 3.50 | 8.58 | Metabolism |
| BVL33_03375 | 30S ribosomal protein S10 | 2.08 | 30.42 | 2.28 | 5.55 | Metabolism |
| BVL33_03435 | 50S ribosomal protein L24 | 2.42 | 24.00 | 4.37 | 3.22 | Metabolism |
| BVL33_03500 | 30S ribosomal protein S4 | 3.41 | 13.28 | 2.47 | 16.66 | Metabolism |
| BVL33_14915 | acyl-CoA dehydrogenase | 4.44 | 7.56 | 2.46 | 3.19 | Metabolism |
| BVL33_04785 | integration host factor subunit alpha | 12.70 | 5.81 | 6.56 | 11.07 | Metabolism |
| BVL33_14350 | VOC family protein | 2.87 | 4.87 | 3.28 | 4.64 | Metabolism |
| BVL33_14925 | 2,4-dienoyl-CoA reductase | 3.46 | 4.51 | 4.17 | 5.50 | Metabolism |
| BVL33_04625 | 3-hydroxyisobutyrate dehydrogenase | 3.14 | 3.10 | 2.44 | 3.25 | Metabolism |
| BVL33_03235 | GTP cyclohydrolase II | 6.02 | 2.98 | 6.53 | 3.89 | Metabolism |
| BVL33_13355 | SCP-2 sterol transfer family protein | 3.01 | 6.90 | 3.02 | 13.07 | Metabolite Transport |
| BVL33_02100 | IS4 family transposase | 15.00 | 4.70 | 77.00 | 6.11 | Resistance |
| BVL33_15710 | Na+/H+ antiporter subunit C | 5.83 | 8.50 | 4.87 | 33.77 | Resistance |
| BVL33_10090 | MBL fold metallo-hydrolase | 7.89 | 5.56 | 14.22 | 8.98 | Resistance |
| BVL33_13770 | efflux transporter periplasmic adaptor subunit | 4.53 | 4.02 | 5.41 | 2.33 | Resistance |
| BVL33_13780 | adenosine deaminase | 6.49 | 3.83 | 3.28 | 2.04 | Resistance |
| BVL33_02685 | sodium:proton antiporter | 27.50 | 2.57 | 144.50 | 7.71 | Resistance |
| BVL33_01605 | efflux transporter periplasmic adaptor subunit | 4.76 | 2.33 | 2.46 | 3.82 | Resistance |
| BVL33_11410 | glutathione S-transferase | 3.46 | 2.77 | 3.26 | 2.52 | Resistance & Signalling |
| BVL33_11715 | glutathione S-transferase | 4.52 | 2.51 | 2.46 | 3.09 | Resistance & Signalling |
| BVL33_14240 | response regulator | 2.51 | 9.69 | 3.70 | 60.47 | Signalling |
| BVL33_02040 | ion channel protein Tsx | 3.37 | 7.82 | 9.72 | 2.61 | Signalling |
| BVL33_13200 | co-chaperone GroES | 2.70 | 6.16 | 2.82 | 4.36 | Signalling |
| BVL33_14235 | response regulator | 5.73 | 3.49 | 4.93 | 2.63 | Signalling |
| BVL33_14220 | hybrid sensor histidine kinase/response regulator | 4.35 | 2.94 | 2.00 | 2.01 | Signalling |
| BVL33_04800 | thiol reductase thioredoxin | 4.42 | 9.02 | 4.23 | 4.60 | Stress Response |
| BVL33_12425 | superoxide dismutase | 4.44 | 6.13 | 3.76 | 19.06 | Stress Response |
| BVL33_00220 | NADPH-dependent FMN reductase | 2.24 | 5.24 | 19.19 | 27.53 | Stress Response |
| BVL33_12940 | DNA starvation/stationary phase protection protein | 823.90 | 3.50 | 179.50 | 2.17 | Stress Response |
| BVL33_13710 | transcriptional regulator | 63.30 | 4.46 | 226.20 | 7.05 | Transcriptional Regulator |
| BVL33_07060 | serine protease | 3.54 | 2.27 | 2.11 | 3.06 | Transcriptional Regulator |
| BVL33_04615 | ribosome silencing factor RsfS | 2.76 | 9.01 | 2.47 | 17.79 | Translation Regulator |
| BVL33_14205 | entericidin, EcnA/B family | 5.45 | 11.83 | 2.47 | 12.14 | Virulence, Disease & Defense |
| BVL33_14745 | preprotein translocase subunit YajC | 14.79 | 8.03 | 4.87 | 10.43 | Virulence, Disease & Defense |
| BVL33_04950 | protein FilA | 17.07 | 5.88 | 4.21 | 9.87 | Virulence, Disease & Defense |
| BVL33_15450 | HopJ type III effector protein | 146.90 | 4.97 | 792.20 | 4.59 | Virulence, Disease & Defense |
| BVL33_09420 | bifunctional adenosylcobinamide kinase/adenosylcobinamide-phosphate guanylyltransferase | 63.40 | 2.24 | 170.20 | 13.80 | Virulence, Disease & Defense |
| Fold Change | ||||||
|---|---|---|---|---|---|---|
| Locus Tag | Gene Product | Effluent Retentate | Effluent Wash | Chlorinated Effluent Retentate | Chlorinated Effluent Wash | Category |
| BVL33_10620 | thiamine biosynthesis protein ThiS | −2.16 | −30.91 | −90.70 | −139.10 | Cellular Repair |
| BVL33_11020 | (4Fe-4S)-binding protein | −2.16 | −50.30 | −72.30 | −150.90 | Cellular Repair |
| BVL33_15925 | hypothetical protein | −4.31 | −104.94 | −192.00 | −2.43 | None |
| BVL33_00180 | hypothetical protein | −88.10 | −90.14 | −88.10 | −126.20 | None |
| BVL33_03255 | hypothetical protein | −2.16 | −4.66 | −83.20 | −3.89 | None |
| BVL33_06630 | amino acid transport protein | −88.10 | −6.40 | −88.10 | −6.40 | None |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jumat, M.R.; Haroon, M.F.; Al-Jassim, N.; Cheng, H.; Hong, P.-Y. An Increase of Abundance and Transcriptional Activity for Acinetobacter junii Post Wastewater Treatment. Water 2018, 10, 436. https://doi.org/10.3390/w10040436
Jumat MR, Haroon MF, Al-Jassim N, Cheng H, Hong P-Y. An Increase of Abundance and Transcriptional Activity for Acinetobacter junii Post Wastewater Treatment. Water. 2018; 10(4):436. https://doi.org/10.3390/w10040436
Chicago/Turabian StyleJumat, Muhammad Raihan, Muhammad Fauzi Haroon, Nada Al-Jassim, Hong Cheng, and Pei-Ying Hong. 2018. "An Increase of Abundance and Transcriptional Activity for Acinetobacter junii Post Wastewater Treatment" Water 10, no. 4: 436. https://doi.org/10.3390/w10040436
APA StyleJumat, M. R., Haroon, M. F., Al-Jassim, N., Cheng, H., & Hong, P. -Y. (2018). An Increase of Abundance and Transcriptional Activity for Acinetobacter junii Post Wastewater Treatment. Water, 10(4), 436. https://doi.org/10.3390/w10040436