Nitrogen and Phosphorus Uptake Dynamics in Tropical Cerrado Woodland Streams

 , , and
, , and
Abstract
:1. Introduction
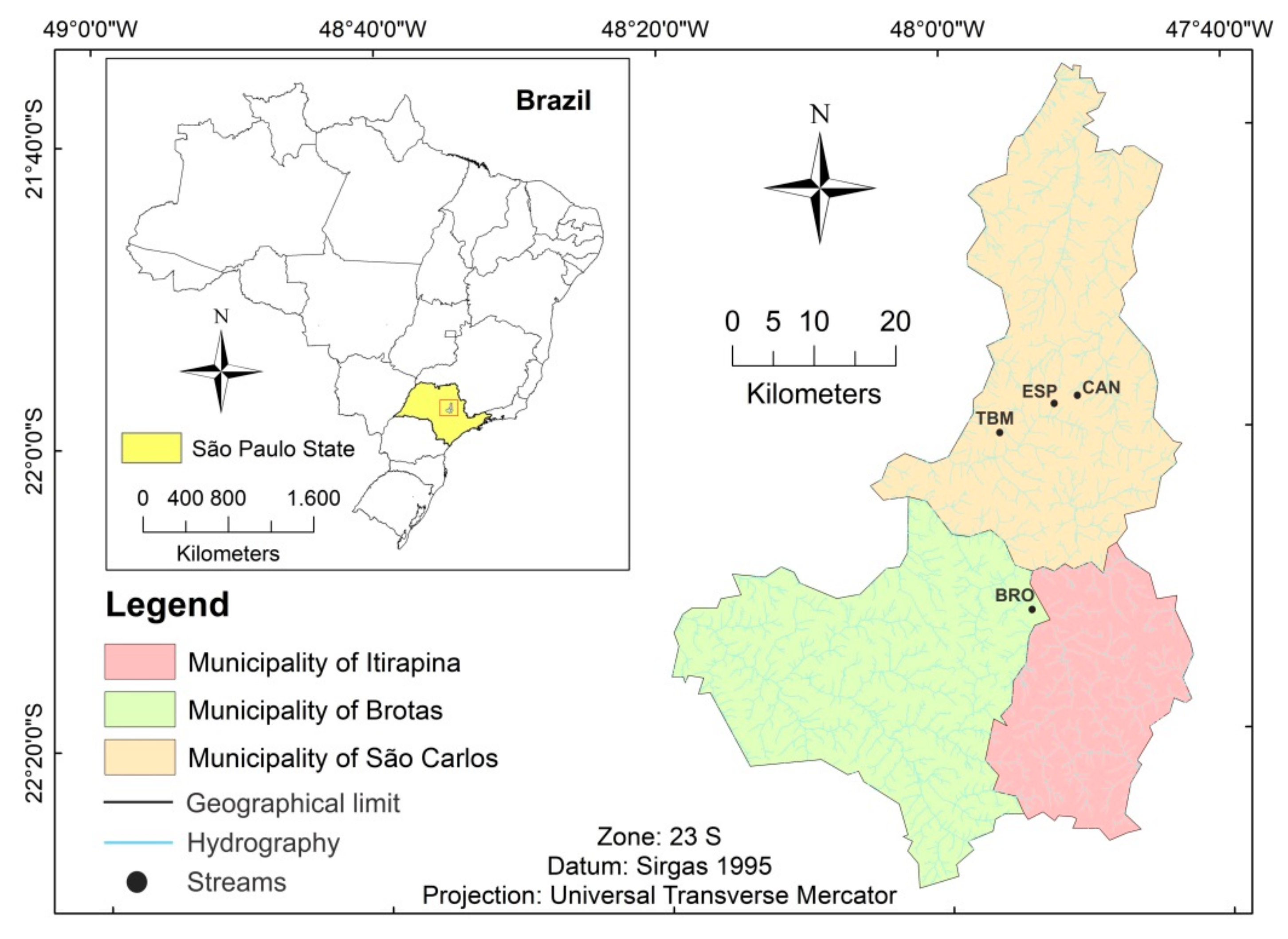
2. Methods
3. Results
3.1. General Characterization of the Streams
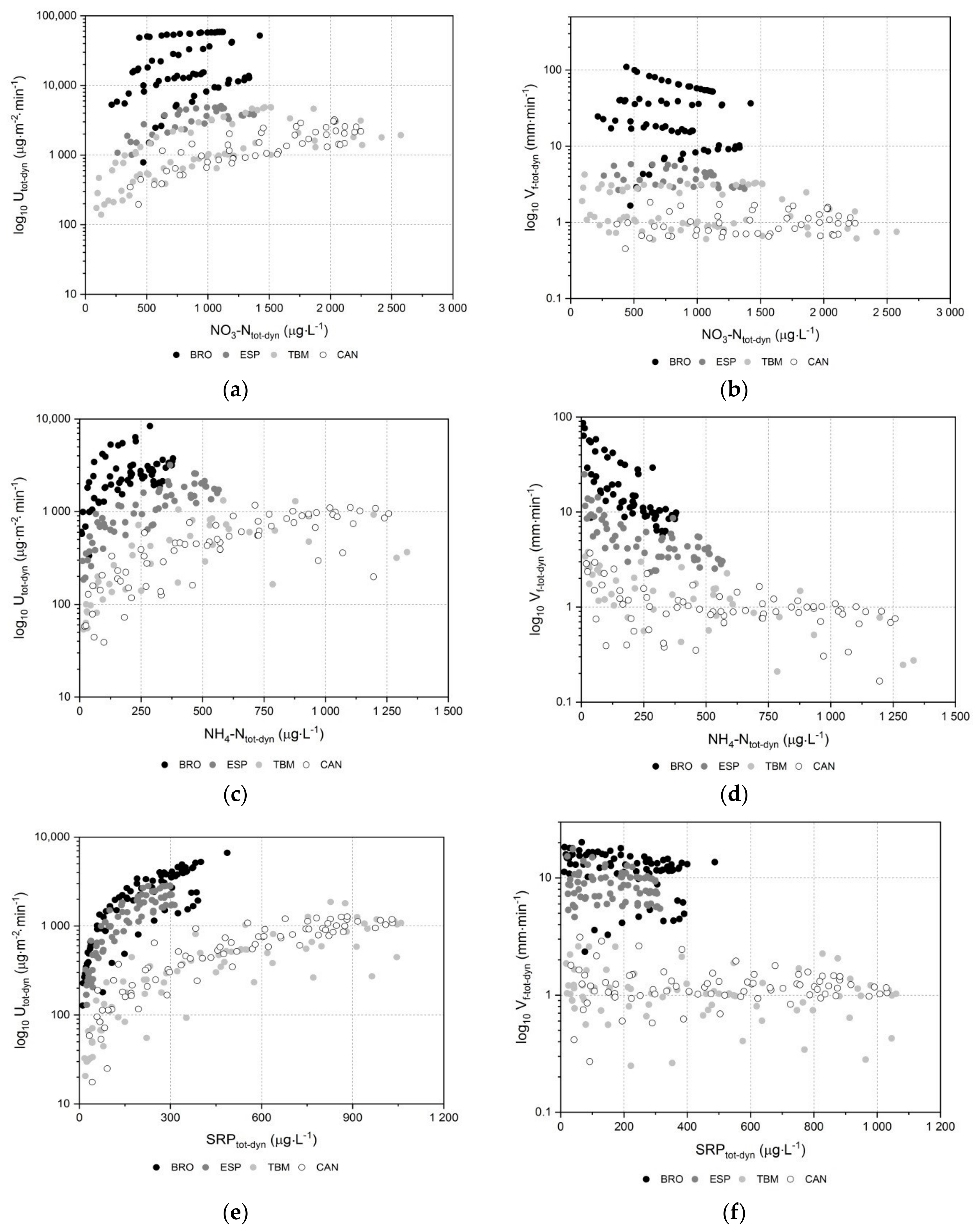
3.2. Nutrient Uptake Metrics and Kinetics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Newbold, J.D.; O’Neill, R.V.; Elwood, J.W.; Van Winkle, W. Nutrient Spiraling in Streams: Implications for Nutrient Limitation and Invertebrate Activity. Am. Nat. 1982, 120, 628–652. [Google Scholar] [CrossRef]
- Stream Solute Workshop. Concepts and Methods for Assessing Solute Dynamics in Stream Ecosystems. J. N. Am. Benthol. Soc. 1990, 9, 95–119. [Google Scholar] [CrossRef]
- Rabalais, N.N. Nitrogen in Aquatic Ecosystems. AMBIO J. Hum. Environ. 2002, 31, 102–112. [Google Scholar] [CrossRef]
- Vanni, M.J.; Renwick, W.H.; Bowling, A.M.; Horgan, M.J.; Christian, A.D. Nutrient stoichiometry of linked catchment-lake systems along a gradient of land use. Freshw. Biol. 2011, 56, 791–811. [Google Scholar] [CrossRef]
- Matson, P.A.; McDowell, W.H.; Townsend, A.R.; Vitousek, P.M. The globalization of N deposition: Ecosystem consequences in tropical environments. Biogeochemistry 1999, 46, 67–83. [Google Scholar] [CrossRef]
- Bennett, E.M.; Carpenter, S.R.; Caraco, N.F. Human Impact on Erodable Phosphorus and Eutrophication: A Global Perspective. AIBS Bull. 2001, 51, 227–231. [Google Scholar] [CrossRef]
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the Nitrogen Cycle: Recent Trends, Questions, and Potential Solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in Freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.J.O.; Tank, J.L. Ecosystem metabolism controls nitrogen uptake in streams in Grand Teton National Park, Wyoming. Limnol. Oceanogr. 2003, 48, 1120–1128. [Google Scholar] [CrossRef] [Green Version]
- Fellows, C.S.; Valett, H.M.; Dahm, C.N.; Mulholland, P.J.; Thomas, S.A. Coupling nutrient uptake and energy flow in headwater streams. Ecosystems 2006, 9, 788–804. [Google Scholar] [CrossRef]
- Gücker, B.; Pusch, M.T. Regulation of nutrient uptake in eutrophic lowland streams. Limnol. Oceanogr. 2006, 51, 1443–1453. [Google Scholar] [CrossRef]
- Valett, H.M.; Crenshaw, C.L.; Wagner, P.F.; Oct, N. Stream Nutrient Uptake, Forest Succession and Biogeochemical Theory. Ecology 2002, 83, 2888–2901. [Google Scholar] [CrossRef]
- Gücker, B.; Boëchat, I. Stream morphology controls ammonium retention. Ecology 2004, 85, 2818–2827. [Google Scholar] [CrossRef]
- Ensign, S.H.; Doyle, M.W. Nutrient spiraling in streams and river networks. J. Geophys. Res. Biogeosci. 2006, 111, 1–13. [Google Scholar] [CrossRef]
- Bernhardt, E.S.; Hall, R.O.; Likens, G.E. Whole-system estimates of nitrification and nitrate uptake in streams of the Hubbard Brook Experimental Forest. Ecosystems 2002, 5, 419–430. [Google Scholar] [CrossRef]
- Rodríguez-Cardona, B.; Wymore, A.S.; McDowell, W.H. DOC:NO3− ratios and NO3− uptake in forested headwater streams. J. Geophys. Res. Biogeosci. 2016, 121, 205–217. [Google Scholar] [CrossRef]
- Dodds, W.K.; López, A.J.; Bowden, W.B.; Gregory, S.; Grimm, N.B.; Hamilton, S.K.; Hershey, A.E.; Martí, E.; McDowell, W.H.; Meyer, J.L.; et al. N uptake as a function of concentration in streams. J. N. Am. Benthol. Soc. 2002, 21, 206–220. [Google Scholar] [CrossRef]
- Earl, S.R.; Valett, H.M.; Webster, J.R. Nitrogen saturation in stream ecosystems. Ecology 2006, 87, 3140–3151. [Google Scholar] [CrossRef]
- Tromboni, F.; Thomas, S.A.; Gücker, B.; Neres-Lima, V.; Lourenço-Amorim, C.; Moulton, T.P.; Silva-Junior, E.F.; Feijó-Lima, R.; Boëchat, I.G.; Zandonà, E. Nutrient limitation and the stoichiometry of nutrient uptake in a tropical rainforest stream. J. Geophys. Res. Biogeosci. 2018. [Google Scholar] [CrossRef]
- O’Brien, J.M.; Dodds, W.K.; Wilson, K.C.; Murdock, J.N.; Eichmiller, J. The saturation of N cycling in Central Plains streams: 15N experiments across a broad gradient of nitrate concentrations. Biogeochemistry 2007, 84, 31–49. [Google Scholar] [CrossRef] [Green Version]
- Hall, R.O.; Bernhardt, E.S.; Likens, G.E. Relating Nutrient Uptake with Transient Storage in Forested Mountain Streams. Limnol. Oceanogr. 2008, 47, 255–265. [Google Scholar] [CrossRef]
- Martí, E.; Fonollà, P.; Von Schiller, D.; Sabater, F.; Argerich, A.; Ribot, M.; Riera, J.L. Variation in stream C, N and P uptake along an altitudinal gradient: A space-for-time analogue to assess potential impacts of climate change. Hydrol. Res. 2009, 40, 123. [Google Scholar] [CrossRef]
- Ribot, M.; Von Schiller, D.; Peipoch, M.; Sabater, F.; Grimm, N.B.; Martí, E. Influence of nitrate and ammonium availability on uptake kinetics of stream biofilms. Freshw. Sci. 2013, 32, 1155–1167. [Google Scholar] [CrossRef] [Green Version]
- Hoellein, T.J.; Tank, J.L.; Rosi-Marshall, E.J.; Entrekin, S.A.; Lamberti, G.A. Controls of nutrient variation on spatial and temporal variation of uptake in three Michigan headwater streams. Limnology 2007, 52, 1964–1977. [Google Scholar] [CrossRef]
- Von Schiller, D.; Martí, E.; Riera, J.L.; Ribot, M.; Argerich, A.; Fonolla, P.; Sabater, F. Inter-annual, Annual, and Seasonal Variation of P and N Retention in a Perennial and an Intermittent Stream. Ecosystems 2008, 11, 670–687. [Google Scholar] [CrossRef] [Green Version]
- Bechtold, H.A.; Marcarelli, A.M.; Baxter, C.V.; Inouye, R.S. Effects of N, P, and organic carbon on stream biofilm nutrient limitation and uptake in a semi-arid watershed. Limnol. Oceanogr. 2012, 57, 1544–1554. [Google Scholar] [CrossRef] [Green Version]
- Piper, L.R.; Cross, W.F.; McGlynn, B.L. Colimitation and the coupling of N and P uptake kinetics in oligotrophic mountain streams. Biogeochemistry 2017, 132, 165–184. [Google Scholar] [CrossRef]
- Hill, B.H.; Mccormick, F.H.; Harvey, B.C.; Johnson, S.L.; Warren, M.L.; Elonen, C.M. Microbial enzyme activity, nutrient uptake and nutrient limitation in forested streams. Freshw. Biol. 2010, 55, 1005–1019. [Google Scholar] [CrossRef]
- Schade, J.D.; Macneill, K.; Thomas, S.A.; Camille Mcneely, F.; Welter, J.R.; Hood, J.; Goodrich, M.; Power, M.E.; Finlay, J.C. The stoichiometry of nitrogen and phosphorus spiralling in heterotrophic and autotrophic streams. Freshw. Biol. 2011, 56, 424–436. [Google Scholar] [CrossRef]
- Gibson, C.A.; Reilly, C.M.O.; Conine, A.L.; Lipshutz, S.M. Nutient uptake across a gradient of nutrient concentration and ratios at the landscape scale. J. Geophys. Res. Biogeosci. 2015, 120, 326–340. [Google Scholar] [CrossRef]
- Wymore, A.S.; Coble, A.A.; Rodríguez-Cardona, B.; McDowell, W.H. Nitrate uptake across biomes and the influence of elemental stoichiometry: A new look at LINX II. Glob. Biogeochem. Cycles 2016, 30, 1183–1191. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Cole, J.J.; Finzi, A.C.; Holland, E.A. Introduction to coupled biogeochemical cycles. Front. Ecol. Environ. 2011, 9, 5–8. [Google Scholar] [CrossRef] [Green Version]
- Marklein, A.R.; Houlton, B.Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol. 2012, 193, 696–704. [Google Scholar] [CrossRef] [PubMed]
- UNFPA. The State of World Population. 2017. Available online: https://www.unfpa.org/sites/default/files/sowp/downloads/UNFPA_PUB_2017_EN_SWOP.pdf (accessed on 1 August 2018).
- MEA. Millennium Ecosystem Assessment. Ecosystems and Human Well Being: Synthesis; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Tromboni, F.; Dodds, W.K. Relationships between Land Use and Stream Nutrient Concentrations in a Highly Urbanized Tropical Region of Brazil: Thresholds and Riparian Zones. Environ. Manag. 2017, 60, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Gücker, B.; Silva, R.C.S.; Graeber, D.; Monteiro, J.A.F.; Brookshire, E.N.J.; Chaves, R.C.; Boëchat, I.G. Dissolved nutrient exports from natural and human-impacted Neotropical catchments. Glob. Ecol. Biogeogr. 2016, 25, 378–390. [Google Scholar] [CrossRef]
- Mulholland, P.J.; Tank, J.L.; Webster, J.R.; Bowden, W.B.; Dodds, W.K.; Gregory, S.V.; Grimm, N.B.; Hamilton, S.K.; Johnson, S.L.; Mart, E.; et al. Can uptake length in streams be determined by nutrient addition experiments? Results from an interbiome comparison study. J. N. Am. Benthol. Soc. 2002, 21, 544–560. [Google Scholar] [CrossRef]
- Solomon, C.T.; Hotchkiss, E.R.; Moslemi, J.M.; Ulseth, A.J.; Stanley, E.H.; Hall, R.O., Jr.; Flecker, A.S. Sediment size and nutrients regulate denitrification in a tropical stream. J. N. Am. Benthol. Soc. 2009, 29, 480–490. [Google Scholar] [CrossRef]
- Potter, J.D.; McDowell, W.H.; Merriam, J.L.; Peterson, B.J.; Thomas, S.M. Denitrification and total nitrate uptake in streams of a tropical landscape. Ecol. Appl. 2010, 20, 2104–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koenig, L.E.; Song, C.; Wollheim, W.M.; Rüegg, J.; Mcdowell, W.H. Nitrification increases nitrogen export from a tropical river network. Freshw. Sci. 2017, 36, 698–712. [Google Scholar] [CrossRef]
- Newbold, J.D.; Bott, T.L.; Kaplan, L.A.; Dow, C.L.; Jackson, J.K.; Aufdenkampe, A.K.; Martin, L.A.; Van Horn, D.J.; de Long, A.A. Uptake of nutrients and organic C in streams in New York City drinking-water-supply watersheds. J. N. Am. Benthol. Soc. 2006, 25, 998–1017. [Google Scholar] [CrossRef]
- Mulholland, P.J.; Helton, A.M.; Poole, G.C.; Hall, R.O.; Hamilton, S.K.; Peterson, B.J.; Tank, J.L.; Ashkenas, L.R.; Cooper, L.W.; Dahm, C.N.; et al. Stream denitrification across biomes and its response to anthropogenic nitrate loading. Nature 2008, 452, 202–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, C.A.; O’Reilly, C.M. Organic matter stoichiometry influences nitrogen and phosphorus uptake in a headwater stream. Freshw. Sci. 2012, 31, 395–407. [Google Scholar] [CrossRef]
- Bernot, M.J.; Tank, J.L.; Royer, T.V.; David, M.B. Nutrient uptake in streams draining agricultural catchments of the midwestern United States. Freshw. Biol. 2006, 51, 499–509. [Google Scholar] [CrossRef]
- Niyogi, D.K.; Simon, K.S.; Townsend, C.R. Land use and stream ecosystem functioning: Nutrient uptake in streams that contrast in agricultural development. Arch. Hydrobiol. 2004, 160, 471–486. [Google Scholar] [CrossRef]
- Covino, T.; McGlynn, B.; McNamara, R. Land use/land cover and scale influences on in-stream nitrogen uptake kinetics. J. Geophys. Res. Biogeosci. 2012, 117, 1–13. [Google Scholar] [CrossRef]
- Fonseca, B.M.; De Mendonça-Galvão, L.; Padovesi-Fonseca, C.; De Abreu, L.M.; Fernandes, A.C.M. Nutrient baselines of Cerrado low-order streams: Comparing natural and impacted sites in Central Brazil. Environ. Monit. Assess. 2014, 186, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Covino, T.P.; McGlynn, B.L.; McNamara, R.A. Tracer Additions for Spiraling Curve Characterization (TASCC): Quantifying stream nutrient uptake kinetics from ambient to saturation. Limnol. Oceanogr. Methods 2010, 8, 484–498. [Google Scholar] [CrossRef] [Green Version]
- Solórzano, L. Determination of ammonia in natural waters by phenol hypochlorite method. Limnol. Oceanogr. 1969, 14, 799–801. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water and Wastewater, 22nd ed.; American Water Works Assn: Washington, DC, USA, 2012. [Google Scholar]
- Lemmon, P.E. A New Instrument for Measuring Forest Overstory Density. J. For. 1957, 55, 667–668. [Google Scholar]
- Lemmon, P.E. A Spherical Densiometer for Estimating Forest Overstory Density. For. Sci. 1956, 2, 314–320. [Google Scholar]
- Webster, J.R.; Valett, H.M. Solute Dynamics. Methods in Stream Ecology; Hauer, F.R., Lamberti, G., Eds.; Elsevier Inc.: New York, NY, USA, 2006; pp. 169–185. ISBN 9780123329080. [Google Scholar]
- Alexander, R.B.; Smith, R.A.; Schwarz, G.E. Effect of stream channel size on the delivery of nitrogen to the Gulf of Mexico. Nature 2000, 403, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Webster, J.R.; Mulholland, P.J.; Tank, J.L.; Valett, H.M.; Dodds, W.K.; Peterson, B.J.; Bowden, W.B.; Dahm, C.N.; Findlay, S.; Gregory, S.V. Others Factors affecting ammonium uptake in streams–an inter-biome perspective. Freshw. Biol. 2003, 48, 1329–1352. [Google Scholar] [CrossRef]
- Rosa, R.D.S.; Aguiar, A.C.F.; Boëchat, I.G.; Gücker, B. Impacts of fish farm pollution on ecosystem structure and function of tropical headwater streams. Environ. Pollut. 2013, 174, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Saltarelli, W.A.; Dodds, W.K.; Tromboni, F.; Calijuri, M.C.; Neres-lima, V.; Jordão, C.E.; Cunha, D.G.F. Variation of stream metabolism along a tropical environmental gradient. J. Limnol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Merriam, J.L.; McDowell, W.H.; Tank, J.L.; Wollheim, W.M.; Crenshaw, C.L.; Johnson, S.L. Characterizing nitrogen dynamics, retention and transport in a tropical rainforest stream using an in situ 15N addition. Freshw. Biol. 2002, 47, 143–160. [Google Scholar] [CrossRef]
- Small, G.E.; Ardón, M.; Duff, J.H.; Jackman, A.P.; Ramírez, A.; Triska, F.J.; Pringle, C.M. Phosphorus retention in a lowland Neotropical stream following an eight-year enrichment experiment. Freshw. Sci. 2016, 35, 1–11. [Google Scholar] [CrossRef]
- Schanz, F.; Juon, H. Two different methods of evaluating nutrient limitations of periphyton bioassays, using water from the River Rhine and eight of its tributaries. Hydrobiologia 1983, 102, 187–195. [Google Scholar] [CrossRef]
- Dodds, W.; Whiles, M. Nitrogen, sulfur, phosphorus, and other nutrients. In Freshwater Ecology; Dodds, W.K., Whiles, M., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2010; ISBN 978-0-12-374724-2. [Google Scholar]
- Newbold, J.D.; Elwood, J.W.; O’Neill, R.V.; Sheldon, A.L. Phosphorus Dynamics in a Woodland Stream Ecosystem: A Study of Nutrient Spiralling. Ecology 1983, 64, 1249–1265. [Google Scholar] [CrossRef]
- Meyer, J.L. The Role of Sediments and Bryophytes in Phosphorus Dynamics in a Head Water Stream Ecosystem. Limnol. Oceanogr. 1979, 24, 365–375. [Google Scholar] [CrossRef]
- Demars, B.O.L. Whole-stream phosphorus cycling: Testing methods to assess the effect of saturation of sorption capacity on nutrient uptake length measurements. Water Res. 2008, 42, 2507–2516. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.M.; Stanley, E.H.; Lottig, N.R. Quantifying phosphorus uptake using pulse and steady-state approaches in streams. Limnol. Oceanogr. Methods 2009, 7, 498–508. [Google Scholar] [CrossRef] [Green Version]
- House, W.A.; Warwick, M.S. Interactions of phosphorus with sediments in the River Swale, Yorkshire, UK. Hydrol. Process. 1999, 13, 1103–1115. [Google Scholar] [CrossRef]
- Reddy, K.R.; Kadlec, R.H.; Flaig, E.; Gale, P.M. Phosphorus retention in streams and wetlands: A review. Crit. Rev. Environ. Sci. Technol. 1999, 29, 83–146. [Google Scholar] [CrossRef]
- House, W.H.; Denison, F.H.; Armitage, P. Comparison of the uptake of inorganic P to a suspended and stream bed-sediment. Water Res. 1995, 29, 767–779. [Google Scholar] [CrossRef]
- Silva, J.S.O.; da Bustamante, M.M.C.; Markewitz, D.; Krusche, A.V.; Ferreira, L.G. Effects of land cover on chemical characteristics of streams in the Cerrado region of Brazil. Biogeochemistry 2011, 105, 75–88. [Google Scholar] [CrossRef]
- Mortimer, C.H. Chemical exchanges betweeen sediments and water in the Great Lakes—Speculations on probable regulatory mechanisms. Limnol. Oceanogr. 1971, 16, 387–404. [Google Scholar] [CrossRef]
- Angelo, A.D.J.D.; Webster, J.R.; Benfield, E.F. Mechanisms of Stream Phosphorus Retention: An Experimental Study. J. N. Am. Benthol. Soc. 1991, 10, 225–237. [Google Scholar] [CrossRef]
- Valett, H.M.; Thomas, S.A.; Mulholland, P.J.; Webster, J.R.; Dahm, C.N.; Fellows, C.S.; Crenshaw, C.L.; Peterson, C.G. Endogenous and exogenous control of ecosystem function: N cycling in headwater streams. Ecology 2008, 89, 3515–3527. [Google Scholar] [CrossRef] [PubMed]
- Dodds, W.K.; Priscu, J.C.; Ellis, B.K. Seasonal uptake and regeneration of inorganic nitrogen and phosphorus in a large oligotrophic lake: Size-fractionation and antibiotic treatment. J. Plankton Res. 1991, 13, 1339–1358. [Google Scholar] [CrossRef]
- Pastor, A.; Peipoch, M.; Cañas, L.; Chappuis, E.; Ribot, M.; Gacia, E.; Riera, J.L.; Martí, E.; Sabater, F. Nitrogen stable isotopes in primary uptake compartments across streams differing in nutrient availability. Environ. Sci. Technol. 2013, 47, 10155–10162. [Google Scholar] [CrossRef] [PubMed]
- Oviedo-Vargas, D.; Royer, T.V.; Johnson, L.T. Dissolved organic carbon manipulation reveals coupled cycling of carbon, nitrogen, and phosphorus in a nitrogen-rich stream. Limnol. Oceanogr. 2013, 58, 1196–1206. [Google Scholar] [CrossRef] [Green Version]
- Diemer, L.A.; McDowell, W.H.; Wymore, A.S.; Prokushkin, A.S. Nutrient uptake along a fire gradient in boreal streams of Central Siberia. Freshw. Sci. 2015, 34, 1443–1456. [Google Scholar] [CrossRef]
- Appling, A.P.; Heffernan, J.B. Nutrient Limitation and Physiology Mediate the Fine-Scale (De)coupling of Biogeochemical Cycles. Am. Nat. 2014, 184, 384–406. [Google Scholar] [CrossRef] [PubMed]
- Tank, J.L.; Reisinger, A.J. Nutrient Limitation and Uptake Rates in Streams and Rivers of the Greater Yellowstone Ecosystem. Univ. Wyoming Natl. Park Serv. Res. Cent. Annu. Rep. 2013, 36, 153–159. [Google Scholar]
- King, S.A.; Heffernan, J.B.; Cohen, M.J. Nutrient flux, uptake, and autotrophic limitation in streams and rivers. Freshw. Sci. 2014, 33, 85–98. [Google Scholar] [CrossRef]
- Jarvie, H.P.; Smith, D.R.; Norton, L.R.; Edwards, F.K.; Bowes, M.J.; King, S.M.; Scarlett, P.; Davies, S.; Dils, R.M.; Bachiller-Jareno, N. Phosphorus and nitrogen limitation and impairment of headwater streams relative to rivers in Great Britain: A national perspective on eutrophication. Sci. Total Environ. 2018, 621, 849–862. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Stream (Site Code) | Geographic Coordinates | Drainage Area (km2) | General Characteristics |
|---|---|---|---|
| Broa (BRO) | 22°11′40.93″ S 47°53′55.78″ W | 2.68 | Natural vegetation, forested and dense riparian zone, natural substrate, located upstream from a natural wetland |
| Canchim (CAN) | 21°57′54.69″ S 47°50′38.02″ W | 1.17 | Natural vegetation, forested and dense riparian zone, natural substrate, located next to the spring |
| Espraiado (ESP) | 21°58′46.75″ S 47°52′23.11″ W | 2.49 | Natural vegetation, forested and dense riparian zone, meandric channel, natural substrate |
| Mineirinho tributary (TBM) | 22°00′12.78″ S 47°55′40.82″ W | 0.82 | Relatively degraded vegetation, natural substrate, presence of erosive features and pluvial runoff |
| Site | DOC (mg·L−1) | DO (mg·L−1) | pH | EC (µS·cm−1) | T (°C) | Q (L·s−1) | v (m·s−1) | H (cm) | w (m) | CC (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| BRO | 1.3–3.8 | 6.9–7.9 | 4.4–5.3 | 4–6 | 16.5–21.6 | 7.5–11.2 | 0.10–0.30 | 4–16 | 0.6–1.0 | 90–98 |
| ESP | 1.4–3.3 | 5.5–7.4 | 4.9–5.6 | 11–19 | 15–20.6 | 5.0–19.2 | 0.06–0.26 | 5–48 | 0.5–0.6 | 93–98 |
| TBM | 1.2–2.4 | 7.1–8.3 | 5.8–6.1 | 19–28 | 17.5–22.8 | 3.4–6.9 | 0.10–0.60 | 1–5 | 1.0–1.2 | 77–91 |
| CAN | 1.6–2.7 | 7.5–8.5 | 5.9–6.5 | 21–32 | 15–20.5 | 2.0–4.0 | 0.04–0.25 | 1–8 | 0.9–1.1 | 92–97 |
| Site | NH4–N | NO3–N | SRP | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sw-amb (m) | Uamb (µg·m−2·min−1) | Vf-amb (mm·min−1) | Camb (µg·L−1) | Sw-amb (m) | Uamb (µg·m−2·min−1) | Vf-amb (mm·min−1) | Camb (µg·L−1) | Sw-amb (m) | Uamb (µg·m−2·min−1) | Vf-amb (mm·min−1) | Camb (µg·L−1) | |
| All sites | 70 | 75 | 7.5 | 6 | 150 | 593 | 2.4 | 315 | 144 | 35 | 4.3 | 13 |
| (32–253) | (5–593) | (0.8–68) | (5–36) | (7–418) | (56–46,373) | (0.5–123) | (37–457) | (55–559) | (9–149) | (0.5–17) | (1–23) | |
| BRO | 44 | 251 | 24.3 | 8 | 63 | 8829 | 32.0 | 357 | 64 | 53 | 16.0 | 3 |
| (32–59) | (121–593) | (16–68) | (5–24) | (8–418) | (714–46,373) | (2–123) | (177–382) | (55–105) | (10–79) | (7–17) | (1–5) | |
| ESP | 90 | 112 | 9 | 8 | 249 | 1131 | 4.0 | 322 | 138 | 87 | 8.0 | 11 |
| (57–152) | (41–274) | (7–25) | (5–36) | (139–313) | (667–2038) | (3.6–5.6) | (183–407) | (42–154) | (60–149) | (7–12) | (8–13) | |
| TBM | 116 | 13 | 3 | 6 | 194 | 264 | 1.3 | 142 | 283 | 12 | 1.0 | 13 |
| (62–172) | (10–20) | (1.6–3) | (5–6.7) | (95–351) | (56–519) | (1–3) | (37–392) | (156–560) | (9–15) | (0.6–1) | (8–15) | |
| CAN | 193 | 8 | 0.9 | 10 | 132 | 313 | 1.0 | 316 | 201 | 24 | 1.0 | 21 |
| (35–253) | (76–96) | (0.7–6) | (5–12) | (131–373) | (155–328) | (0.5–1.6) | (202–457) | (77–391) | (10–36) | (0.5–1.5) | (18–23) | |
| Dependent Variable | Independent Variable | B | SE | p-Value | Adjusted R2 |
|---|---|---|---|---|---|
| Uamb NO3–N (µg·m−2·min−1) | Intercept | 2.20 | 3.50 | 0.54 | 0.482 |
| SRPamb | −1.33 | 0.52 | 2 × 102 | ||
| NO3amb | 1.14 | 0.57 | 3 × 102 | ||
| Uamb NH4–N (µg·m−2·min−1) | Intercept | −12.25 | 13.15 | 0.37 | 0.824 |
| SRPamb | −1.64 | 0.24 | 3 × 105 | ||
| DO | −4.68 | 1.52 | 1 × 102 | ||
| %CC | 6.61 | 2.65 | 3 × 102 | ||
| Uamb SRP (µg·m−2·min−1) | Intercept | 24.28 | 4.20 | 1 × 104 | 0.816 |
| DO | −4.60 | 1.13 | 2 × 103 | ||
| Q | 0.47 | 0.21 | 4 × 102 | ||
| V | −3.08 | 0.99 | 1 × 102 | ||
| T | −3.85 | 0.92 | 1 × 103 | ||
| Vf-amb NO3–N (mm·min−1) | Intercept | 5.31 | 0.97 | 2 × 104 | 0.508 |
| SRPamb | −1.12 | 0.36 | 1 × 102 | ||
| v | −5.31 | 2.20 | 3 × 102 | ||
| Vf-amb NH4–N (mm·min−1) | Intercept | 13.03 | 3.00 | 1 × 103 | 0.731 |
| SRPamb | −1.35 | 0.22 | 8 × 104 | ||
| DO | −3.74 | 1.39 | 2 × 102 | ||
| Vf-amb SRP (mm·min−1) | Intercept | 21.34 | 3.10 | 3 × 105 | 0.889 |
| SRPamb | −1.05 | 0.12 | 2 × 106 | ||
| DO | −3.71 | 0.84 | 1 × 103 | ||
| T | −2.97 | 0.73 | 2 × 103 | ||
| v | −2.77 | 0.79 | 4 × 103 |
| Stream | Best Kinetic Model (R2) for Vft-tot-dyn versus Ntot-dyn | ||
|---|---|---|---|
| NO3–N | NH4–N | SRP | |
| BRO | - | M-M (0.653) | - |
| ESP | FTO (0.649) | ELS (0.658) | - |
| TBM | - | FTO (0.453) | - |
| CAN | – | ELS (0.364) | - |
| Stream | Best Kinetic Model (R2) for Utot-dyn versus Ntot-dyn | ||
| NO3–N | NH4–N | SRP | |
| BRO | - | ELS (0.465) | ELS (0.787) |
| ESP | ELS (0.697) | FTO (0.795) | ELS (0.881) |
| TBM | FTO (0.596) | ELS (0.679) | FTO (0.831) |
| CAN | ELS (0.733) | FTO (0.702) | FTO (0.854) |
| Nutrient | Stream | Location | Sw-amb (m) | Uamb (µg·m−2·min−1) | Vf-amb (mm·min−1) | Reference |
|---|---|---|---|---|---|---|
| NO3–N | – | Temperate | 101–478 | 5.8–19.1 | 0.8–4.2 | [14] |
| Barra Pequena | Tropical | 467–499 | 94 | – | [19] | |
| Q Bisley | 1192 | 16 | 1.92 | [59] | ||
| – | 315–8480 | – | 0.13–0.94 | [40] | ||
| BRO | 6 (8–418) | 8829 (714–46,373) | 32 (2–123) | This study | ||
| ESP | 249 (139–313) | 1131 (667–2038) | 4 (3.6–5.6) | |||
| TBM | 194 (95–351) | 264 (56–519) | 1.3 (1–3) | |||
| CAN | 132 (131–373) | 313 (155–328) | 1 (0.5–1.6) | |||
| NH4–N | – | Temperate | 23–275 | 5.3–52.6 | 2.5–17.0 | [14] |
| Barra Pequena | Tropical | 138–501 | 5.45–14.2 | – | [19] | |
| Q. Bisley | 15–26 | 33–42 | 87.3 | [59] | ||
| R. Cipó | – | 1000 | – | [13] | ||
| – | – | – | 0.3–8.5 | [41] | ||
| BRO | 44 (32–59) | 251 (121–593) | 24.3 (16–68) | This study | ||
| ESP | 90 (57–152) | 112 (41–274) | 9 (7–25) | |||
| TBM | 116 (62–172) | 13 (10–20) | 3 (1.6–3) | |||
| CAN | 193 (35–253) | 8 (76–96) | 0.9 (0.7–6) | |||
| SRP | – | Temperate | 24–161 | 3.4–15.7 | 1.5–6.6 | [14] |
| Barra Pequena | Tropical | 461–1065 | 6.5–63 | – | [19] | |
| Carapa | – | 950–1150 | – | [60] | ||
| BRO | 64 (55–105) | 53 (10–79) | 16 (7–17) | This study | ||
| ESP | 138 (42–154) | 86 (60–149) | 8 (7–12) | |||
| TBM | 283 (156–560) | 12 (9–15) | 1 (0.6–1) | |||
| CAN | 201 (77–391) | 24 (10–36) | 1 (0.5–1.5) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reinaldo Finkler, N.; Tromboni, F.; Boëchat, I.G.; Gücker, B.; Gasparini Fernandes Cunha, D. Nitrogen and Phosphorus Uptake Dynamics in Tropical Cerrado Woodland Streams. Water 2018, 10, 1080. https://doi.org/10.3390/w10081080
Reinaldo Finkler N, Tromboni F, Boëchat IG, Gücker B, Gasparini Fernandes Cunha D. Nitrogen and Phosphorus Uptake Dynamics in Tropical Cerrado Woodland Streams. Water. 2018; 10(8):1080. https://doi.org/10.3390/w10081080
Chicago/Turabian StyleReinaldo Finkler, Nícolas, Flavia Tromboni, Iola Gonçalves Boëchat, Björn Gücker, and Davi Gasparini Fernandes Cunha. 2018. "Nitrogen and Phosphorus Uptake Dynamics in Tropical Cerrado Woodland Streams" Water 10, no. 8: 1080. https://doi.org/10.3390/w10081080
APA StyleReinaldo Finkler, N., Tromboni, F., Boëchat, I. G., Gücker, B., & Gasparini Fernandes Cunha, D. (2018). Nitrogen and Phosphorus Uptake Dynamics in Tropical Cerrado Woodland Streams. Water, 10(8), 1080. https://doi.org/10.3390/w10081080