Soil Hydrothermal Characteristics among Three Typical Vegetation Types: An Eco-Hydrological Analysis in the Qilian Mountains, China



Abstract
:1. Introduction
2. Materials and Methods
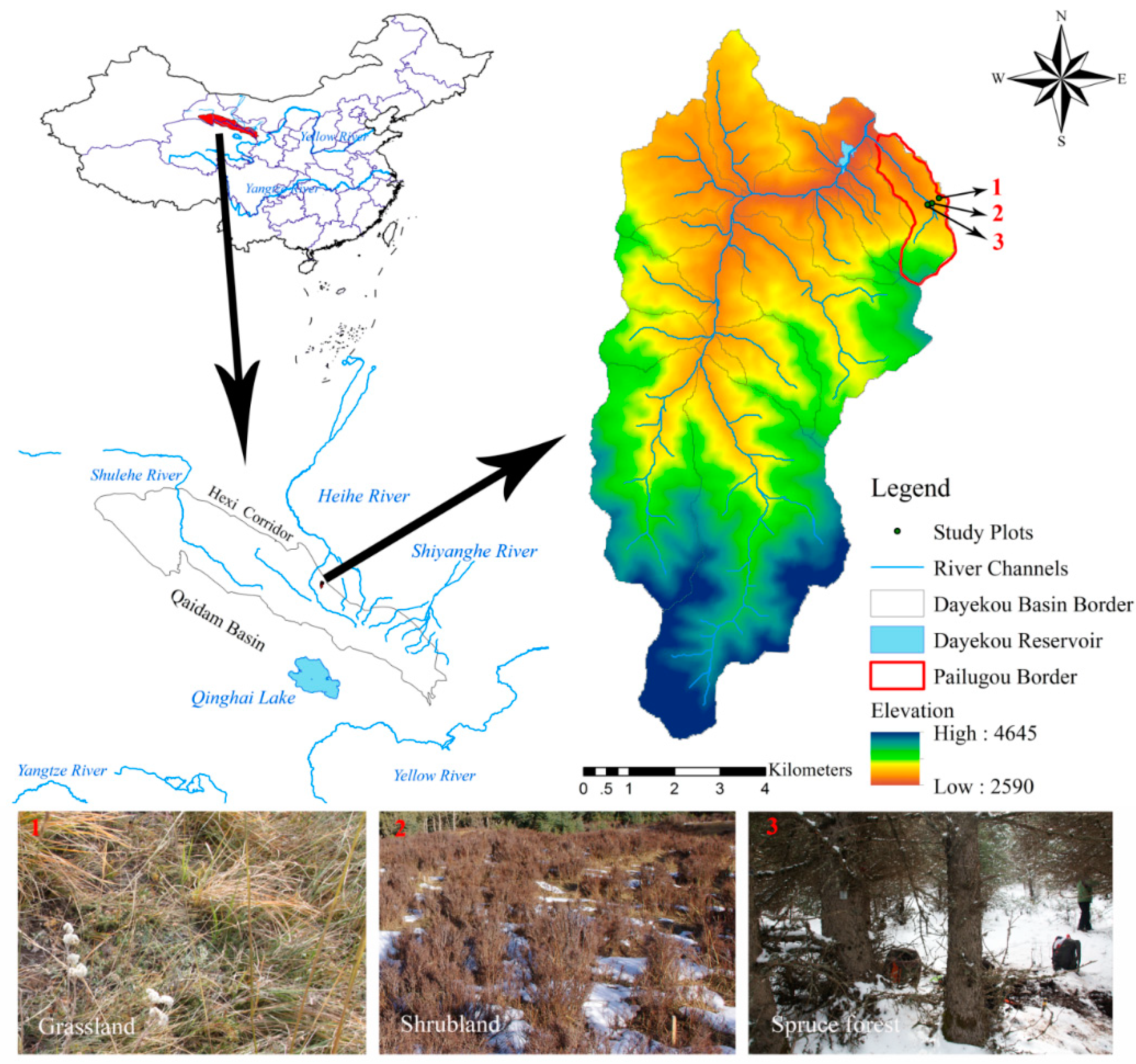
2.1. Study Site
2.2. Experimental Design and Data Measurements
2.3. Water Loss (Evapotranspiration and Leakage)
2.4. Statistical Analysis
3. Results
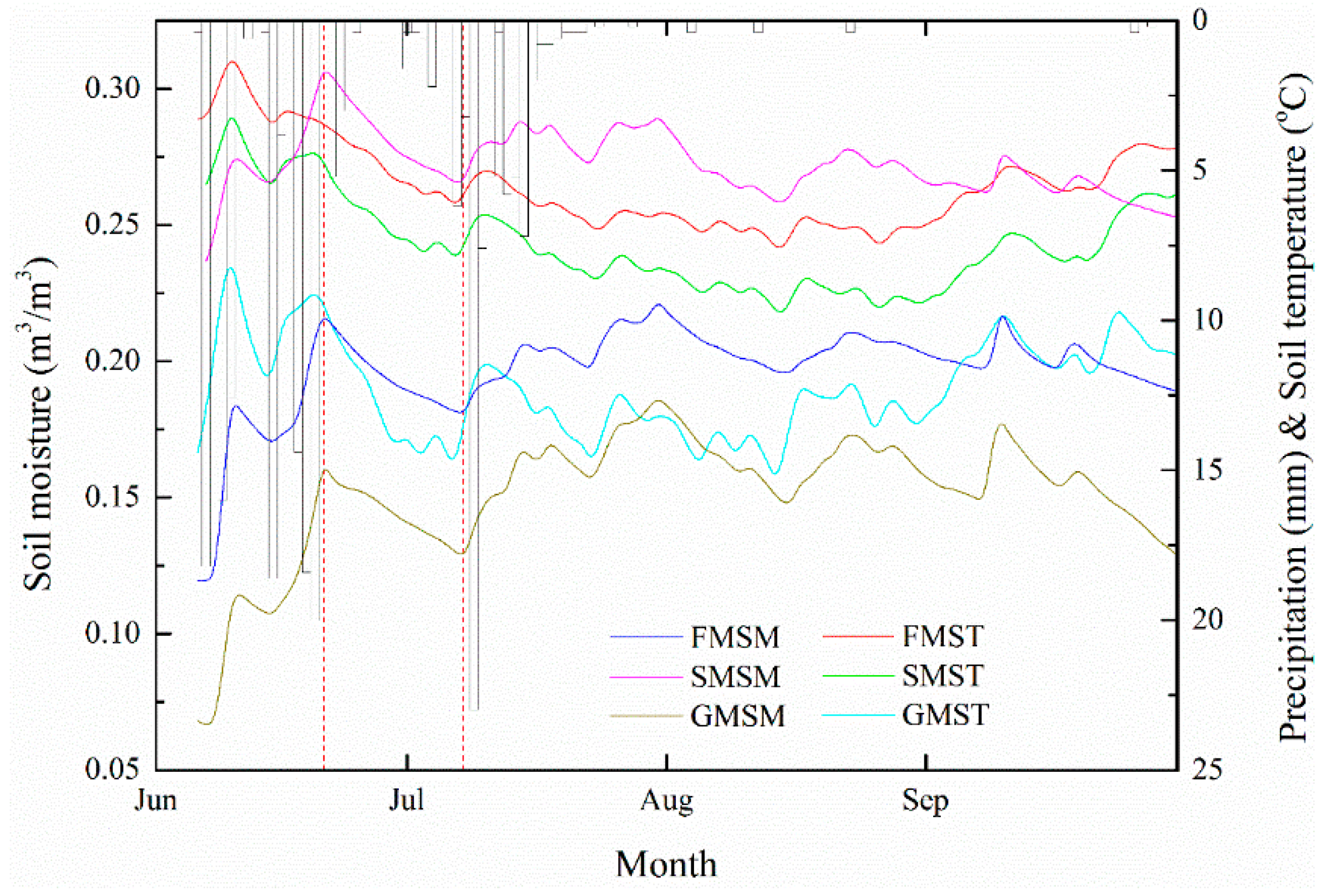
3.1. Soil Moisture Dynamics
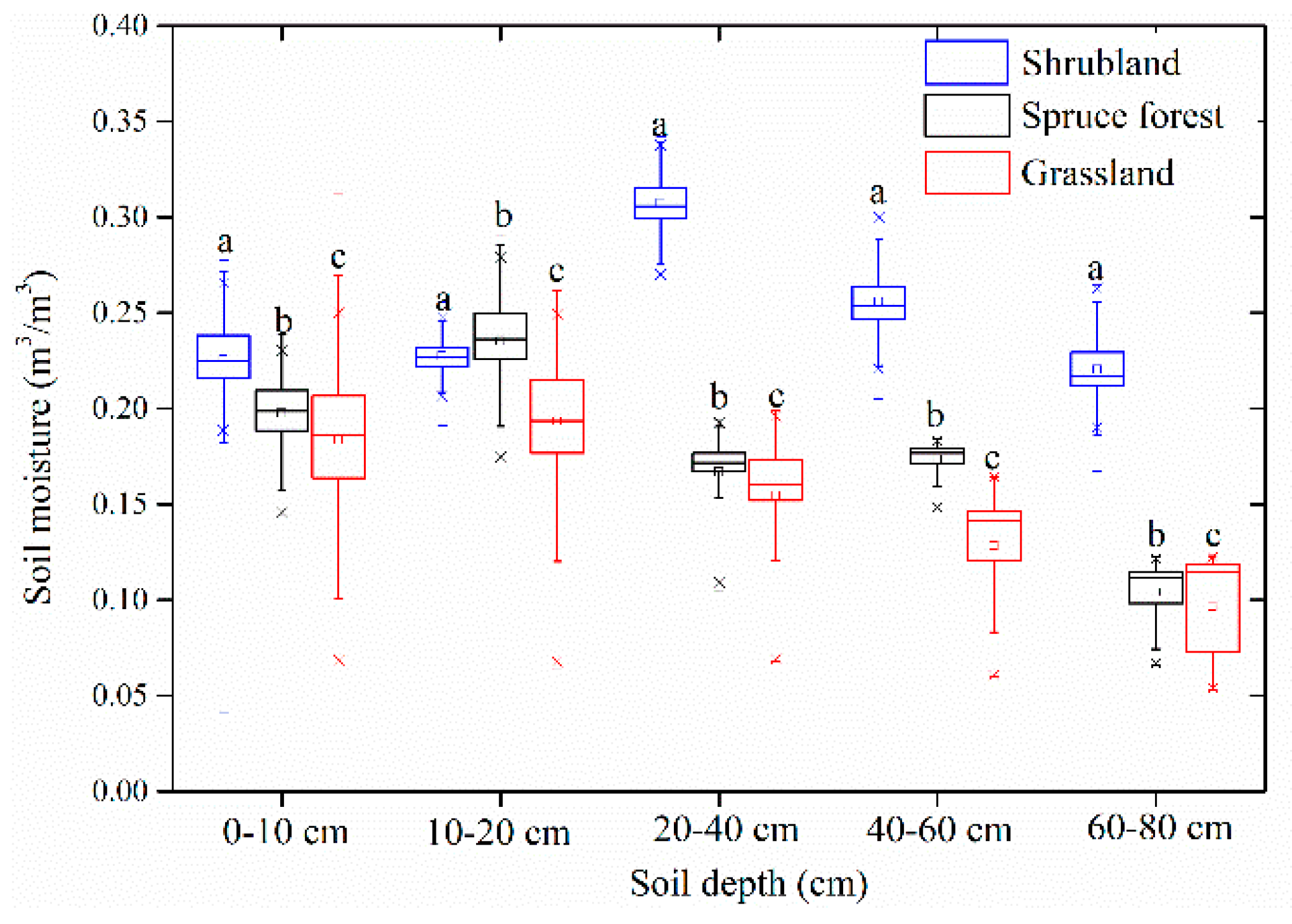
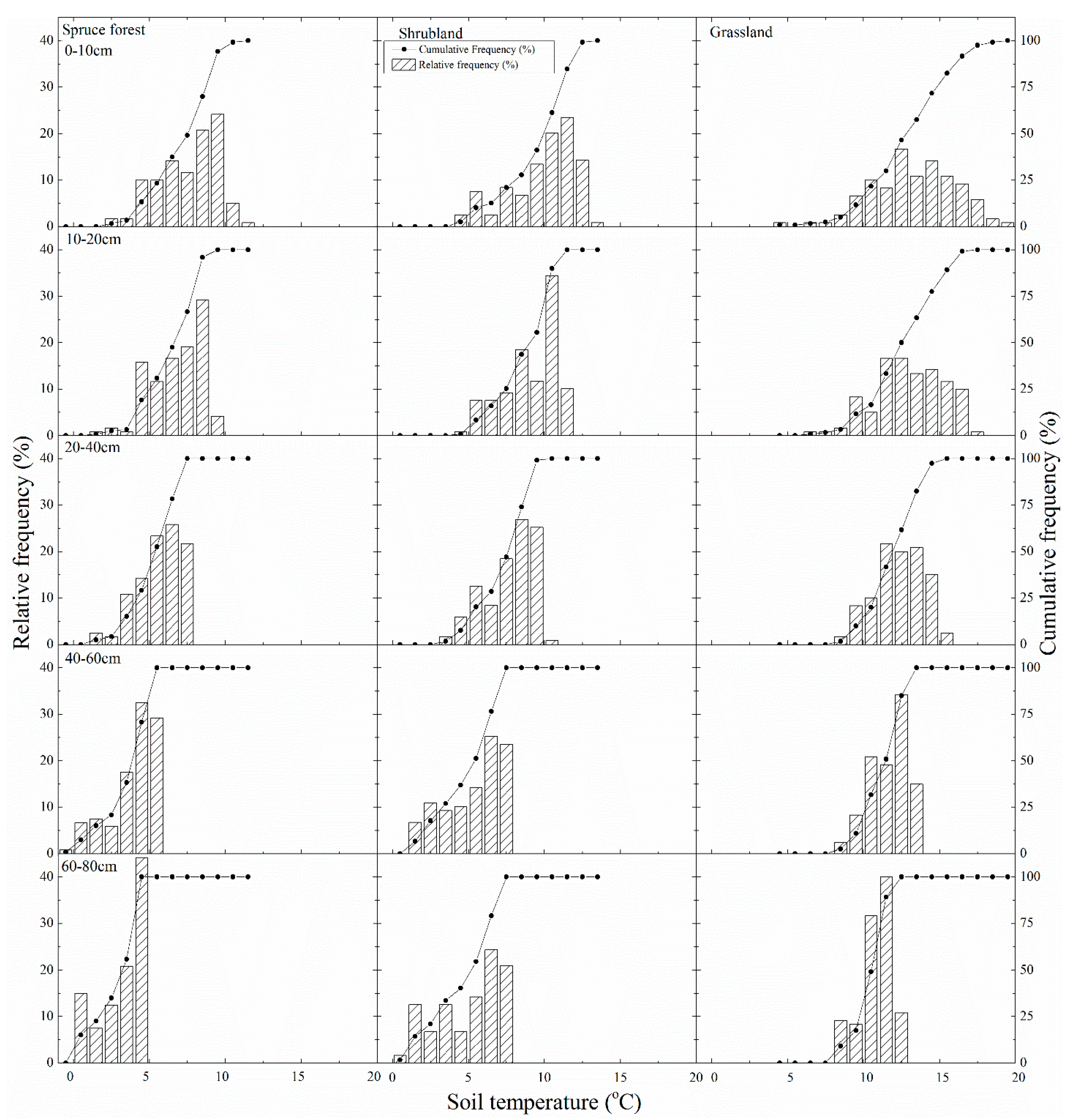
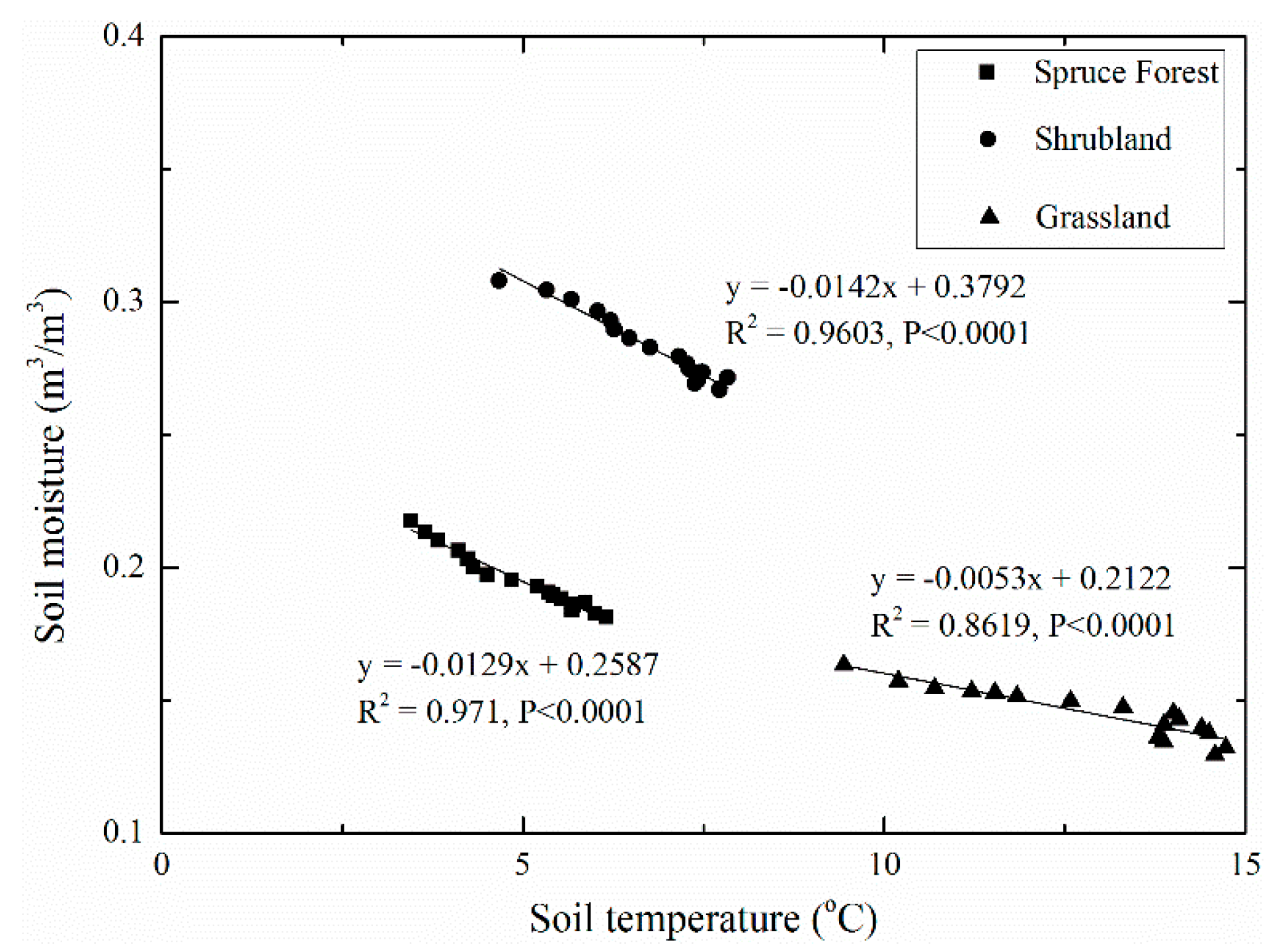
3.2. Comparison of Soil Moisture and Temperature under Three Typical Vegetation Types
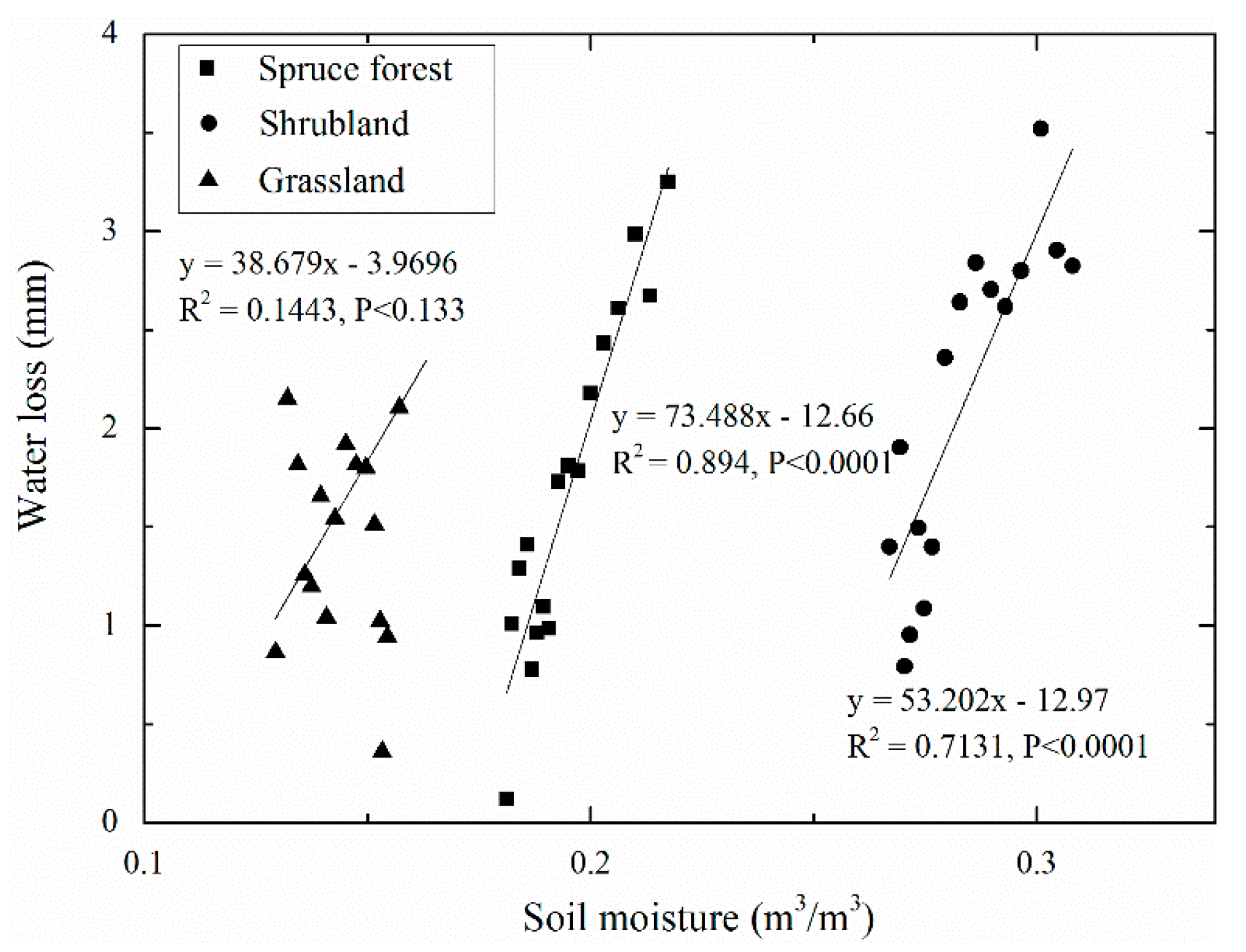
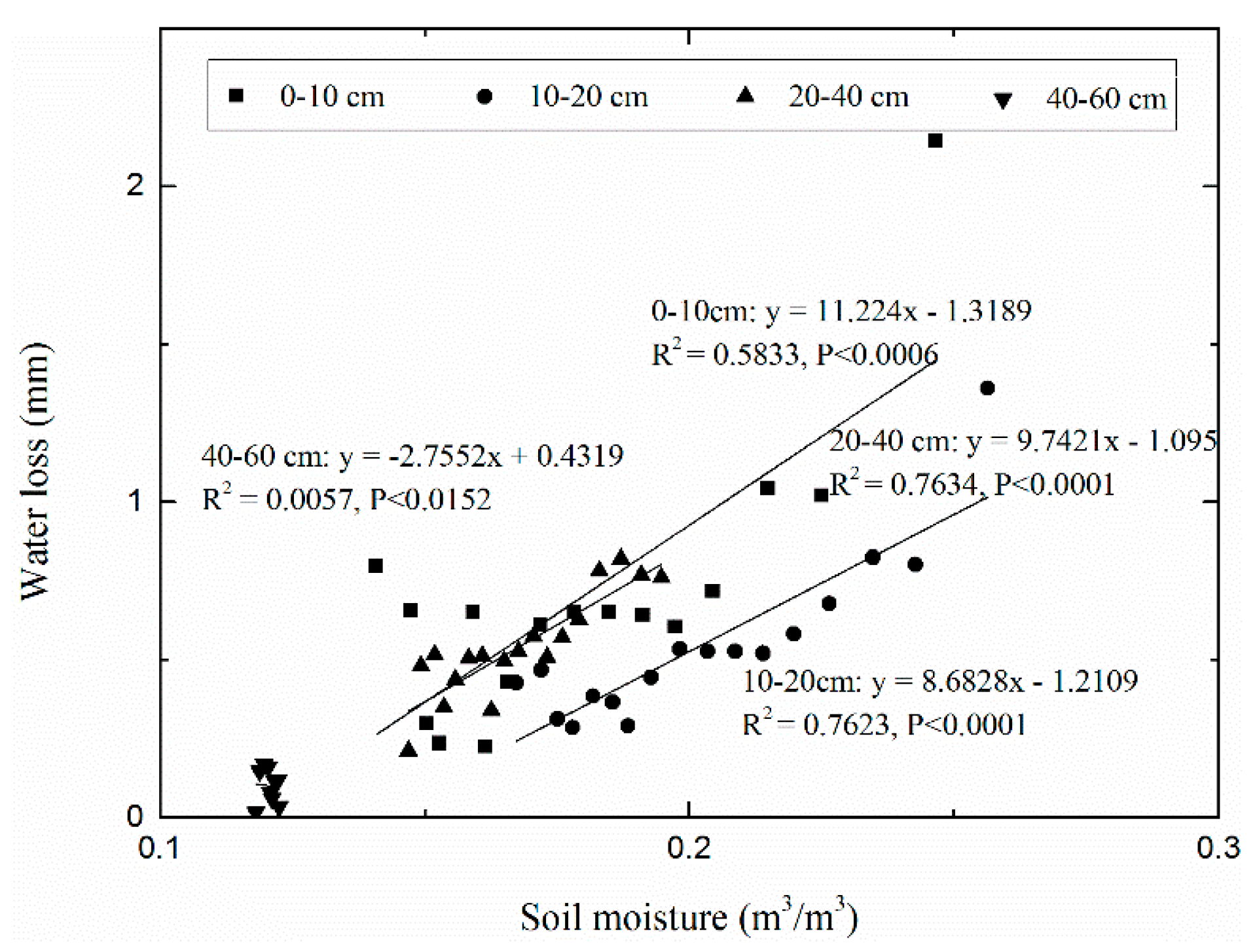
3.3. Water Loss and Its Relationship to Soil Moisture and Temperature
4. Discussion
4.1. Soil Moisture Dynamics under the Three Vegetation Types
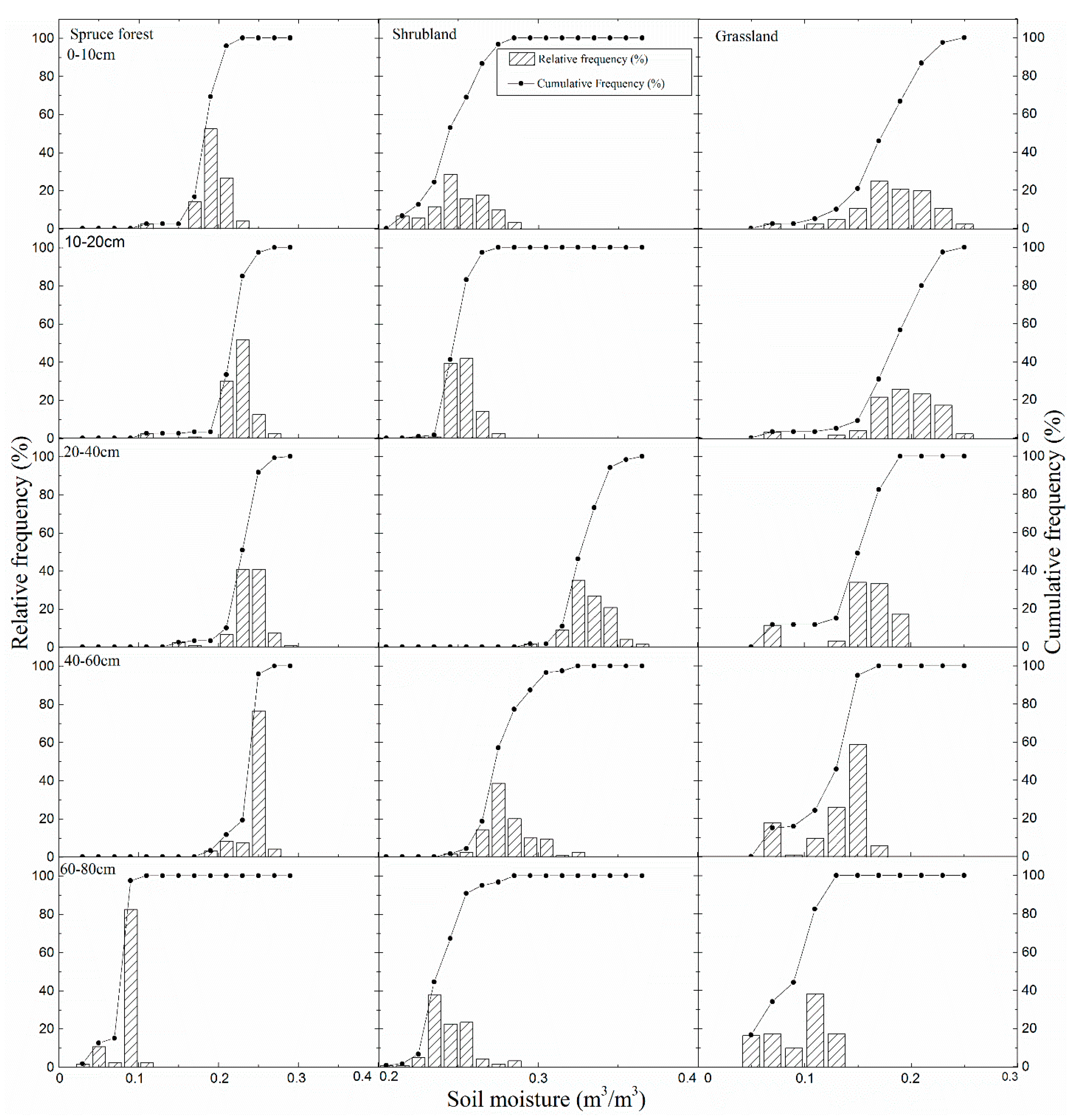
4.2. Soil Moisture and Soil Temperature Probability Distribution and Response to Climate Change
4.3. Water Loss and Its Driving Factors
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Eagleson, P.S. Ecohydrology: Darwinian Expression of Vegetation Form and Function, 2nd ed; Cambridge University Press: Cambridge, UK, 2004; pp. 170–206. [Google Scholar]
- Ma, Y.J.; Li, X.Y.; Guo, L.; Lin, H. Hydropedology: Interactions between pedologic and hydrologic processes across spatiotemporal scales. Earth-Sci. Rev. 2017, 171, 181–195. [Google Scholar] [CrossRef]
- Rodriguez-Iturbe, I.; Porporato, A. Ecohydrology of Water-controlled Ecosystems: Soil Moisture and Plant Dynamics; Cambridge University Press: Cambridge, UK, 2005; pp. 1–47. [Google Scholar]
- Berryman, E.M.; Barnard, H.R.; Adams, H.R.; Burns, M.A.; Gallo, E.; Brooks, P.D. Complex terrain alters temperature and moisture limitations of forest soil respiration across a semiarid to subalpine gradient. J. Geophys. Res. 2015, 120, 707–723. [Google Scholar] [CrossRef]
- D’Odorico, P.; Laio, F.; Porporato, A.; Ridolfi, L.; Rinaldo, A.; Rodriguez-Iturbe, I. Ecohydrology of terrestrial ecosystems. Bioscience 2010, 60, 898–907. [Google Scholar] [CrossRef]
- He, Z.B.; Zhao, M.M.; Zhu, X.; Du, J.; Chen, L.F.; Lin, P.F.; Li, J. Temporal stability of soil water storage in multiple soil layers in high-elevation forests. J. Hydrol. 2019, 569, 532–545. [Google Scholar] [CrossRef]
- Sun, F.; Lü, Y.; Wang, J.; Hu, J.; Fu, B. Soil moisture dynamics of typical ecosystems in response to precipitation: A monitoring-based analysis of hydrological service in the Qilian Mountains. Catena 2015, 129, 63–75. [Google Scholar] [CrossRef]
- Zou, C.B.; Barron-Gafford, G.A.; Breshears, D.D. Effects of topography and woody plant canopy cover on near-ground solar radiation: Relevant energy inputs for ecohydrology and hydropedology. Geophys. Res. Lett. 2007, 34, L24S21. [Google Scholar] [CrossRef]
- Legates, D.R.; Mahmood, R.; Levia, D.F.; DeLiberty, T.L.; Quiring, S.M.; Houser, C.; Nelson, F.E. Soil moisture: A central and unifying theme in physical geography. Prog. Phys. Geog. 2010, 35, 65–86. [Google Scholar] [CrossRef]
- He, Z.B.; Zhao, W.Z.; Liu, H.; Chang, X.X. The response of soil moisture to rainfall event size in subalpine grassland and meadows in a semi-arid mountain range: A case study in northwestern China’s Qilian Mountains. J. Hydrol. 2012, 420–421, 183–190. [Google Scholar] [CrossRef]
- Noy-Meir, I. Desert ecosystems: Environment and producers. Annu. Rev. Ecol. Evol. S. 1973, 4, 25–51. [Google Scholar] [CrossRef]
- Laio, F.; Porporato, A.; Ridolfi, L.; Rodrguez-Iturbe, I. Plants in water-controlled ecosystems: Active role in hydrologic processes and response to water stress II. Probabilistic soil moisture dynamics. Adv. Water Res. 2001, 24, 707–723. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, W.W.; Wang, L.X.; Liu, Y.X.; Liu, Y.; Feng, Q. Relationship between soil water content and soil particle size on typical slopes of the Loess Plateau during a drought year. Sci. Total Environ. 2019, 648, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Migala, K.; Wojtun, B.; Szymanski, W.; Muskala, P. Soil moisture and temperature variation under different types of tundra vegetation during the growing season: A case study from the Fuglebekken catchment, SW Spitsbergen. Catena 2014, 116, 10–18. [Google Scholar] [CrossRef]
- Wang, S.; Fu, B.; Gao, G.; Liu, Y.; Zhou, J. Responses of soil moisture in different land cover types to rainfall events in a re-vegetation catchment area of the Loess Plateau, China. Catena 2013, 101, 122–128. [Google Scholar] [CrossRef]
- Wang, S.; Fu, B.J.; Gao, G.Y.; Yao, X.L.; Zhou, J. Soil moisture and evapotranspiration of different land cover types in the Loess Plateau, China. Hydrol. Earth Syst. Sc. 2012, 16, 2883–2892. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Nie, X.F.; Zhou, X.B.; Liao, K.H.; Li, H.P. Soil moisture response to rainfall at different topographic positions along a mixed land-use hillslope. Catena 2014, 119, 61–70. [Google Scholar] [CrossRef]
- Katul, G.; Porporato, A.; Oren, R. Stochastic dynamics of plant-water interactions. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 767–791. [Google Scholar] [CrossRef]
- Asbjornsen, H.; Goldsmith, G.R.; Alvarado-Barrientos, M.S.; Rebel, K.; Van Osch, F.P.; Rietkerk, M.; Chen, J.Q.; Gotsch, S.; Tobon, C.; Geissert, D.R.; et al. Ecohydrological advances and applications in plant-water relations research: A review. J. Plant Ecol. 2011, 4, 3–22. [Google Scholar] [CrossRef]
- D’Odorico, P.; Porporato, A. Preferential states in soil moisture and climate dynamics. Proc. Natl. Acad. Sci. USA 2004, 101, 8848–8851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seneviratne, S.I.; Corti, T.; Davin, E.L.; Hirschi, M.; Jaeger, E.B.; Lehner, I.; Orlowsky, B.; Teuling, A.J. Investigating soil moisture–climate interactions in a changing climate: A review. Earth-Sci. Rev. 2010, 99, 125–161. [Google Scholar] [CrossRef]
- Bell, J.E.; Sherry, R.; Luo, Y. Changes in soil water dynamics due to variation in precipitation and temperature: An ecohydrological analysis in a tallgrass prairie. Water Resour. Res. 2010, 46, W03523. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Li, X.Y. Soil moisture and temperature dynamics in typical alpine ecosystems: A continuous multi-depth measurements-based analysis from the Qinghai-Tibet Plateau, China. Hydrol. Res. 2018, 49, 194–209. [Google Scholar] [CrossRef]
- Stocker, T.F.; Dahe, Q.; Plattner, G.K. Climate Change 2013: The Physical Science Basis. The Working Group I contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (IPCC). 2013. Available online: http://www.ipcc.ch/report/ar5/wg1/ (accessed on 11 May 2015).
- Weltzin, J.F.; Loik, M.E.; Schwinning, S.; Williams, D.G.; Fay, P.A.; Haddad, B.M.; Harte, J.; Huxman, T.E.; Knapp, A.K.; Lin, G.H.; et al. Assessing the response of terrestrial ecosystems to potential changes in precipitation. Bioscience 2003, 53, 941–952. [Google Scholar] [CrossRef]
- Knapp, A.K.; Beier, C.; Briske, D.D.; Classen, A.T.; Luo, Y.; Reichstein, M.; Smith, M.D.; Smith, S.D.; Bell, J.E.; Fay, P.A.; et al. Consequences of more extreme precipitation regimes for terrestrial ecosystems. Bioscience 2008, 58, 811–821. [Google Scholar] [CrossRef]
- Gao, G.Y.; Zhan, H.B.; Feng, S.Y.; Fu, B.J.; Ma, Y.; Huang, G.H. A new mobile-immobile model for reactive solute transport with scale-dependent dispersion. Water Resour. Res. 2010, 46, W08533. [Google Scholar] [CrossRef]
- Vereecken, H.; Huisman, J.A.; Pachepsky, Y.; Montzka, C.; van der Kruk, J.; Bogena, H.; Weihermuller, L.; Herbst, M.; Martinez, G.; Vanderborght, J. On the spatio-temporal dynamics of soil moisture at the field scale. J. Hydrol. 2014, 516, 76–96. [Google Scholar] [CrossRef]
- Ryu, D.; Famiglietti, J.S. Characterization of footprint-scale surface soil moisture variability using Gaussian and beta distribution functions during the Southern Great Plains 1997 (SGP97) hydrology experiment. Water Resour. Res. 2005, 41, W12433. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, W.Z.; He, Z.B. Self-organized vegetation patterning effects on surface soil hydraulic conductivity: A case study in the Qilian Mountains, China. Geoderma 2013, 192, 362–367. [Google Scholar] [CrossRef]
- He, Z.B.; Fang, S.; Chen, L.F.; Du, J.; Zhu, X.; Lin, P.F. Spatial patterns in natural Piceacrassifolia forests of northwestern China, as basis for close-to-nature forestry. J. Mt. Sci. 2018, 9, 1909–1919. [Google Scholar] [CrossRef]
- Lin, P.F.; He, Z.B.; Du, J.; Chen, L.F.; Zhu, X.; Li, J. Impacts of climate change on reference evapotranspirationin the Qilian Mountains of China: Historical trends and projected changes. Int. J. Climatol. 2018, 38, 2980–2993. [Google Scholar] [CrossRef]
- Zheng, X.L.; Zhao, C.Y.; Peng, S.Z.; Jian, S.Q.; Liang, B.; Wang, X.P.; Yang, S.F.; Wang, C.; Peng, H.H.; Wang, Y. Soil CO2 efflux along an elevation gradient in Qinghai spruce forests in the upper reaches of the Heihe River, northwest China. Environ. Earth Sci. 2014, 71, 2065–2076. [Google Scholar] [CrossRef]
- Sala, O.E.; Lauenroth, W.K. Small rainfall events: An ecological role in semiarid regions. Oecologia 1982, 53, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.P.; Cui, Y.; Pan, Y.X.; Li, X.R.; Yu, Z.; Young, M.H. Effects of rainfall characteristics on infiltration and redistribution patterns in revegetation-stabilized desert ecosystems. J. Hydrol. 2008, 358, 134–143. [Google Scholar] [CrossRef]
- Crockford, R.H.; Richardson, D.P. Partitioning of rainfall into throughfall, stemflow and interception: Effect of forest type, ground cover and climate. Hydrol. Process 2000, 14, 2903–2920. [Google Scholar] [CrossRef]
- Stavi, I.; Lavee, H.; Ungar, E.D.; Sarah, P. Ecogeomorphicfeedbacks in semiarid rangelands: A review. Pedosphere 2009, 19, 217–229. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, C.; Xu, Z.; Wang, Y.; Peng, H. Effect of vegetation on soil water retention and storage in a semi-arid alpine forest catchment. J. Arid Land 2013, 5, 207–219. [Google Scholar] [CrossRef]
- D’Odorico, P.; Caylor, K.; Okin, G.S.; Scanlon, T.M. On soil moisture-vegetation feedbacks and their possible effects on the dynamics of dryland ecosystems. J. Geophys. Res. 2007, 112, G04010. [Google Scholar] [CrossRef]
- Ding, Y.; Huang, H.; Wang, L.; Zhang, Z.Q.; Zhang, W.H. Effect of different organic matter content on soil moisture dynamics. Appl. Mech. Mater. 2014, 477–478, 481–484. [Google Scholar] [CrossRef]
- Moran, M.S.; Hamerlynck, E.P.; Scott, R.L.; Stone, J.J.; Collins, C.D.H.; Keefer, T.O.; Bryant, R.; DeYoung, L.; Nearing, G.S.; Sugg, Z.; et al. Hydrologic response to precipitation pulses under and between shrubs in the Chihuahuan Desert, Arizona. Water Resour. Res. 2010, 46, W10509. [Google Scholar] [CrossRef]
- Pockman, W.T.; Small, E.E. The influence of spatial patterns of soil moisture on the grass and shrub responses to a summer rainstorm in a Chihuahuandesert ecotone. Ecosystems 2010, 13, 511–525. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, G.L.; Yang, J.L.; Li, D.C.; Zhao, Y.G.; Liu, F.; Yang, R.M.; Yang, F. Organic matter controls of soil water retention in an alpine grassland and its significance for hydrological processes. J. Hydrol. 2014, 519, 3086–3093. [Google Scholar] [CrossRef]
- Caylor, K.K.; Shugart, H.H.; Rodriguez-Iturbe, I. Tree canopy effects on simulated water stress in Southern African savannas. Ecosystems 2005, 8, 17–32. [Google Scholar] [CrossRef]
- Porporato, A.; Laio, F.; Ridolfi, L.; Caylor, K.K.; Rodriguez-Iturbe, I. Soil moisture and plant stress dynamics along the Kalahari precipitation gradient. J. Geophys. Res. 2003, 108, 4127. [Google Scholar] [CrossRef]
- Porporato, A.; Laio, F.; Ridolfi, L.; Rodriguez-Iturbe, I. Plants in water-controlled ecosystems: Active role in hydrologic processes and response to water stress III. Vegetation water stress. Adv. Water Resour. 2001, 24, 725–744. [Google Scholar] [CrossRef]
- Jenerette, G.D.; Barron-Gafford, G.A.; Guswa, A.J.; McDonnell, J.J.; Villegas, J.C. Organization of complexity in water limited ecohydrology. Ecohydrology 2012, 5, 184–199. [Google Scholar] [CrossRef]
- D’Odorico, P.; Ridolfi, L.; Porporato, A.; Rodriguez-Iturbe, I. Preferential states of seasonal soil moisture: The impact of climate fluctuations. Water Resour. Res. 2000, 36, 2209–2219. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.R.; Hornberger, G.M. Inferred bimodality in the distribution of soil moisture at Big Meadows, Shenandoah National Park, Virginia. Geophys. Res. Lett. 2006, 33, L06407. [Google Scholar] [CrossRef]
- Teuling, A.J.; Uijlenhoet, R.; Troch, P.A. On bimodality in warm season soil moisture observations. Geophys. Res. Lett. 2005, 32, L13402. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, W.Z.; He, Z.B.; Zhang, L.J. Stochastic modelling of soil moisture dynamics in a grassland of Qilian Mountain at point scale. Sci. China Ser. D-Earth Sci. 2007, 50, 1844–1856. [Google Scholar] [CrossRef]
- Yin, X.Z.; Zhang, Q.; Xu, Q.Y.; Xue, W.X.; Guo, H.; Shi, Z.J. Characteristics of climate change in qilianmountains region in recent 50 years. Plateau Meteorol. 2009, 28, 85–90. (in Chinese). [Google Scholar]
- Turner, N.C.; Kramer, P.J. Adaptations of Plants to Water and High Temperature Stress; John Wiley: New York, NY, USA, 1980; pp. 1–20. [Google Scholar]
- Ferrante, D.; Oliva, G.E.; Fernández, R.J. Soil water dynamics, root systems, and plant responses in a semiarid grassland of Southern Patagonia. J. Arid Environ. 2014, 104, 52–58. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | Shrubland | Spruce Forest | Grassland | |||
|---|---|---|---|---|---|---|
| Mean | Standard Error | Mean | Standard Error | Mean | Standard Error | |
| Clay (%) | 0.71 | 0.08 | 0.81 | 0.02 | 2.20 | 0.17 |
| Silt (%) | 58.12 | 4.18 | 55.39 | 2.56 | 80.27 | 2.24 |
| Sand (%) | 41.17 | 4.23 | 43.80 | 2.58 | 17.53 | 2.40 |
| Bulk density (g/cm3) | 0.84 | 0.07 | 0.75 | 0.10 | 0.95 | 0.06 |
| Soil organic matter (g/kg) | 70.63 | 6.18 | 62.72 | 2.55 | 16.91 | 3.80 |
| Porosity (%) | 70.10 | 2.91 | 71.69 | 3.88 | 64.14 | 2.34 |
| Field capacity (%) | 47.83 | 3.13 | 45.29 | 5.99 | 39.77 | 1.71 |
| Soil Depth (cm) | Degrees of Freedom | Sum of Squares | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| 0–10 cm | 2 | 0.29653 | 0.14826 | 240.3421 | <0.0001 |
| 10–20 cm | 2 | 0.21465 | 0.10733 | 178.4466 | <0.0001 |
| 20–40 cm | 2 | 1.89643 | 0.94821 | 1766.88 | <0.0001 |
| 40–60 cm | 2 | 1.53584 | 0.76792 | 1702.211 | <0.0001 |
| 60–80 cm | 2 | 1.89761 | 0.9488 | 2647.561 | <0.0001 |
| Soil Depth (cm) | Degrees of Freedom | Sum of Squares | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| 0–10 cm | 2 | 1989.968 | 994.9841 | 188.9363 | <0.0001 |
| 10–20 cm | 2 | 2398.674 | 1199.337 | 322.1715 | <0.0001 |
| 20–40 cm | 2 | 2800.094 | 1400.047 | 597.3742 | <0.0001 |
| 40–60 cm | 2 | 3950.727 | 1975.363 | 762.2848 | <0.0001 |
| 60–80 cm | 2 | 3773.258 | 1886.629 | 707.2228 | <0.0001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Lü, D.; Sun, F.; Lü, Y.; Chen, Y.; Zhou, Q. Soil Hydrothermal Characteristics among Three Typical Vegetation Types: An Eco-Hydrological Analysis in the Qilian Mountains, China. Water 2019, 11, 1277. https://doi.org/10.3390/w11061277
Hu J, Lü D, Sun F, Lü Y, Chen Y, Zhou Q. Soil Hydrothermal Characteristics among Three Typical Vegetation Types: An Eco-Hydrological Analysis in the Qilian Mountains, China. Water. 2019; 11(6):1277. https://doi.org/10.3390/w11061277
Chicago/Turabian StyleHu, Jian, Da Lü, Feixiang Sun, Yihe Lü, Youjun Chen, and Qingping Zhou. 2019. "Soil Hydrothermal Characteristics among Three Typical Vegetation Types: An Eco-Hydrological Analysis in the Qilian Mountains, China" Water 11, no. 6: 1277. https://doi.org/10.3390/w11061277
APA StyleHu, J., Lü, D., Sun, F., Lü, Y., Chen, Y., & Zhou, Q. (2019). Soil Hydrothermal Characteristics among Three Typical Vegetation Types: An Eco-Hydrological Analysis in the Qilian Mountains, China. Water, 11(6), 1277. https://doi.org/10.3390/w11061277