Functional Groups of Phytoplankton and Their Relationship with Environmental Factors in the Restored Uzarzewskie Lake

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
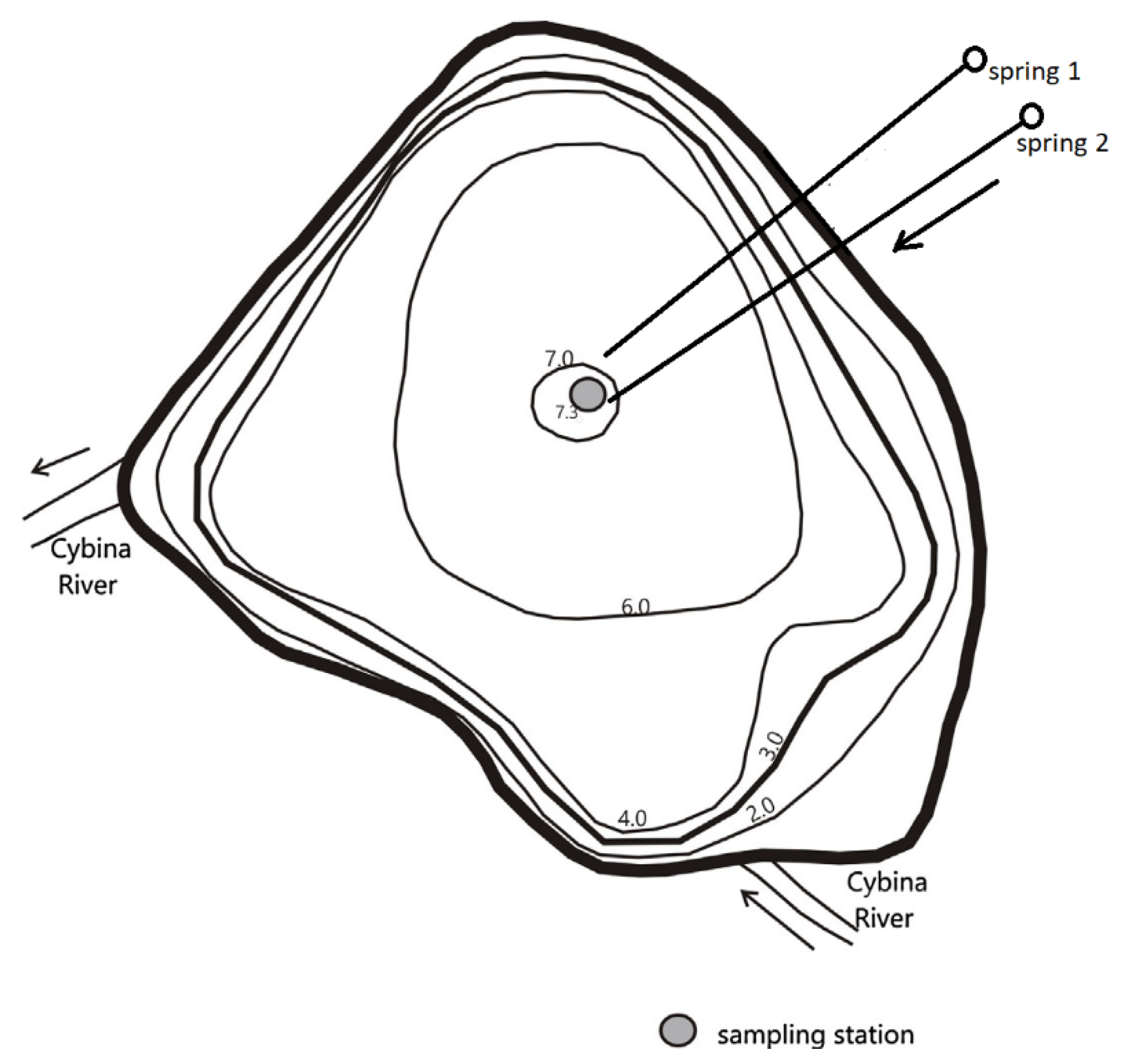
2.1. Uzarzewskie Lake and Its Restoration
2.2. Sampling and Analysis
3. Results
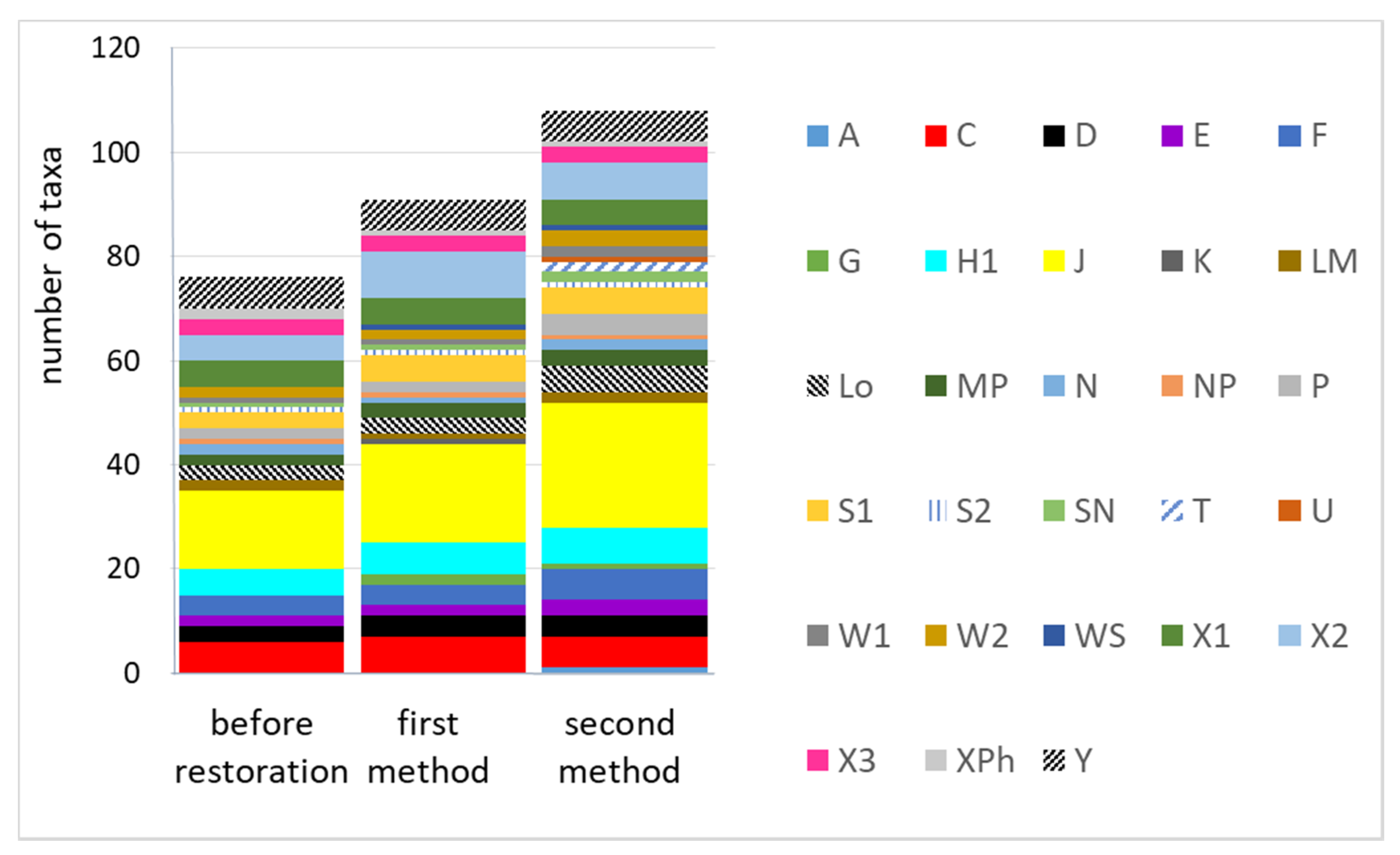
3.1. The Number of Species in Functional Groups in the Three Periods of Study
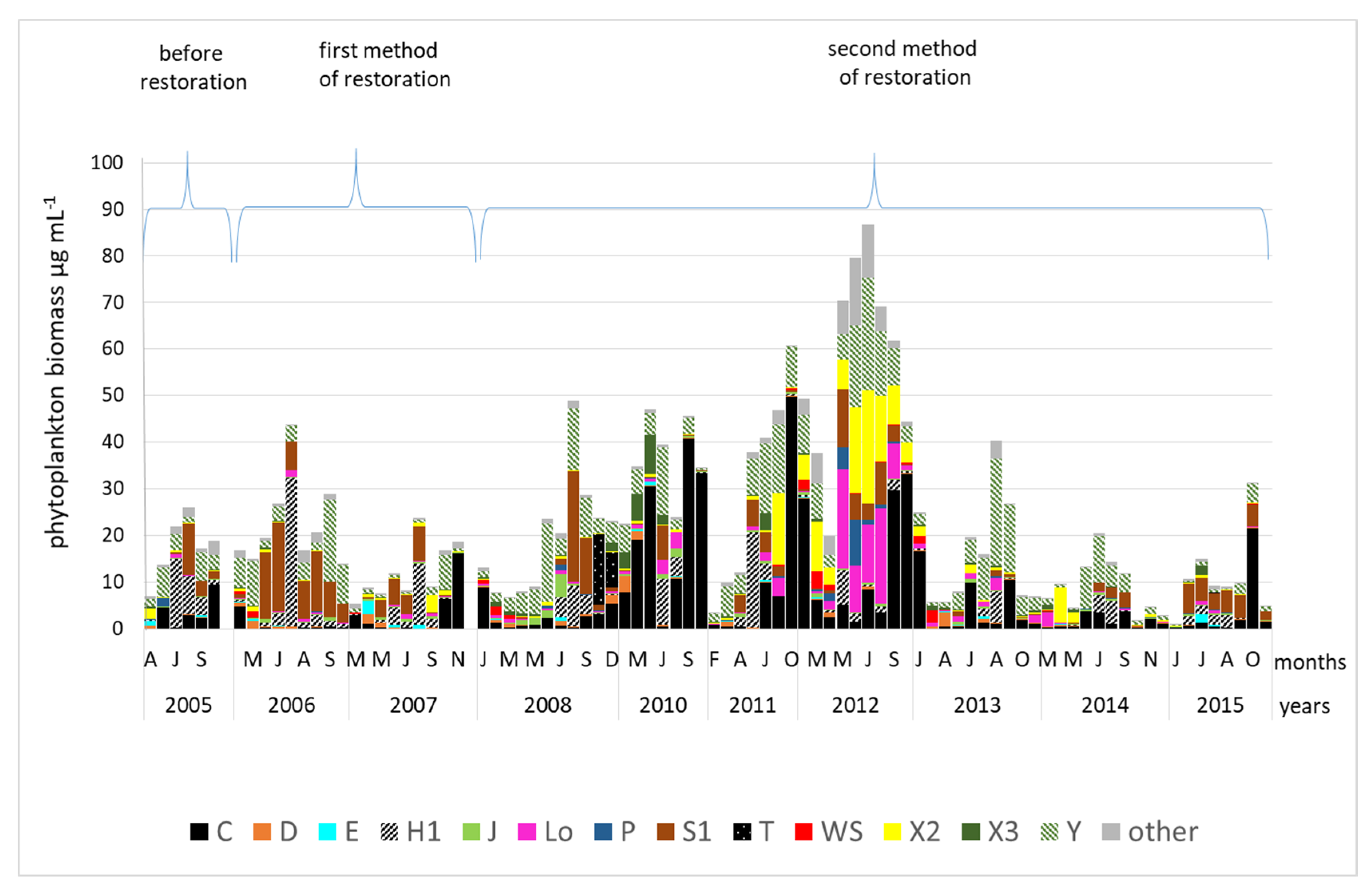
3.2. Changes in the Biomass of Functional Groups in the Three Periods of Study
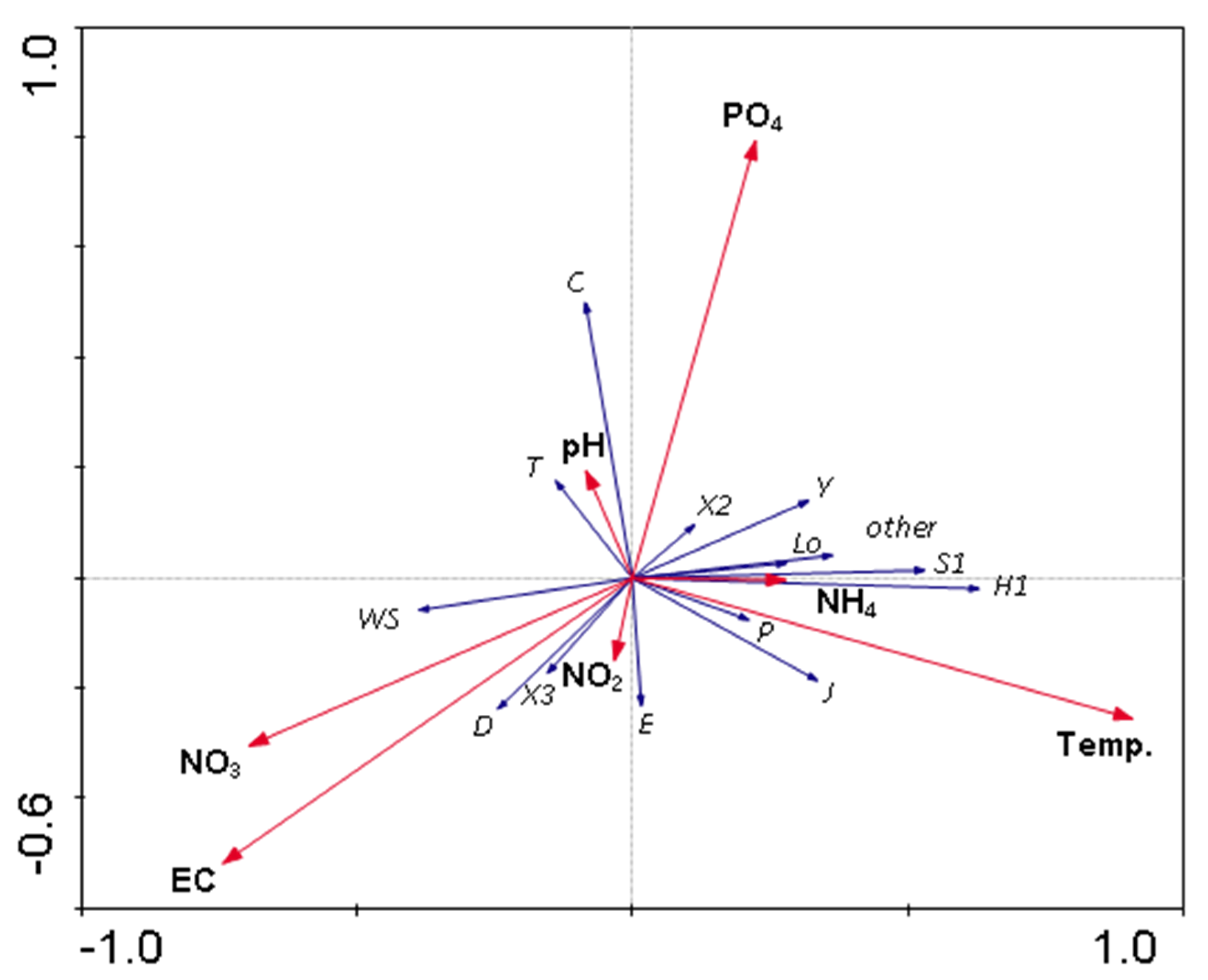
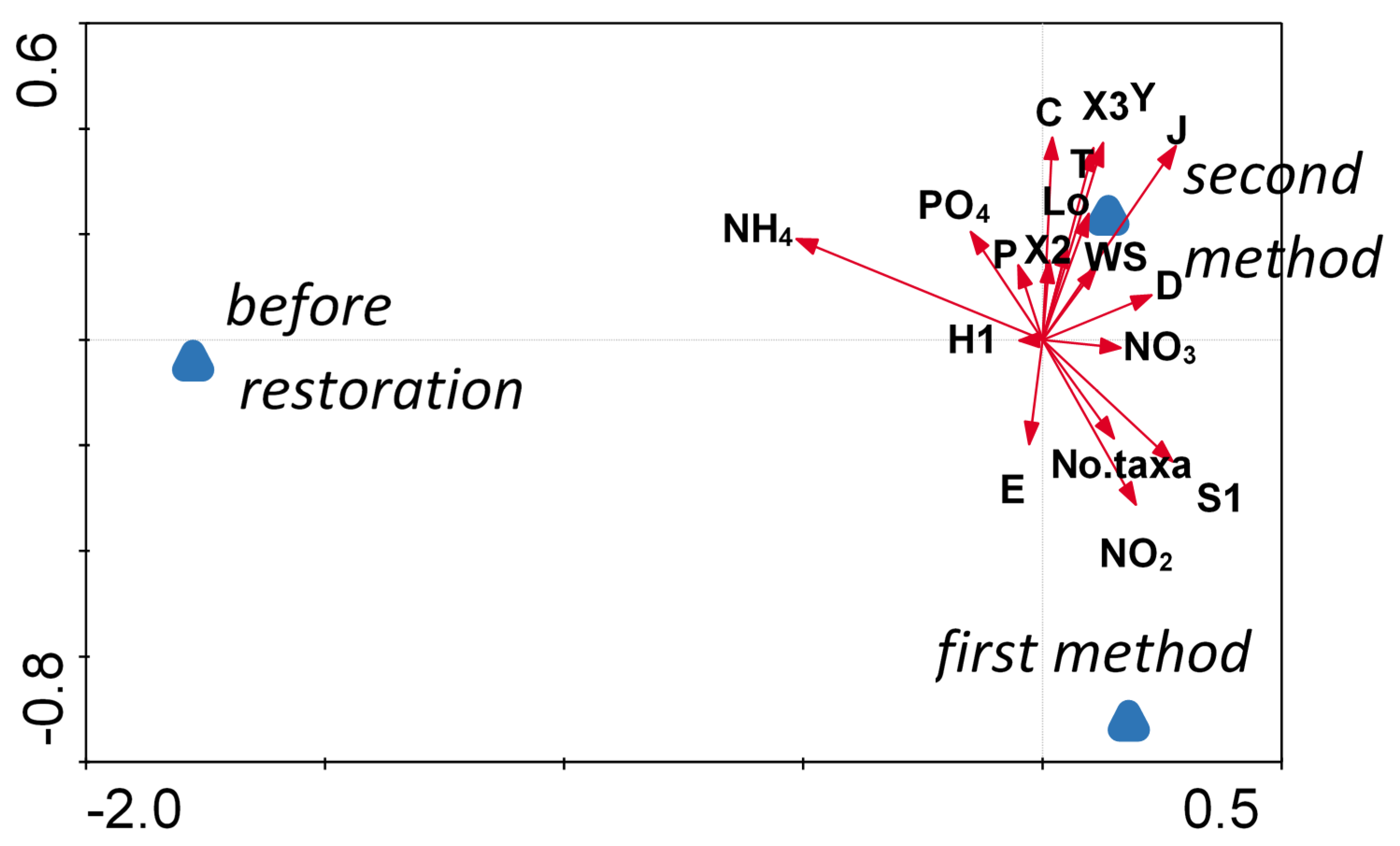
3.3. The Relations of the Phytoplankton to Environmental Factors in Three Periods of Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Paerl, H.W.; Fulton, R.S.; Moisander, P.H.; Dyble, J. Harmful freshwater algal blooms with an emphasis on cyanobacteria. Sci. World J. 2001, 1, 76–113. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471. [Google Scholar] [CrossRef] [PubMed]
- WFD. Directive 2000/60/ec of the European Parliament and of the Council 22.12.2000. Off. J. Eur. Communities 2000, L327, 1–72. [Google Scholar]
- Kozak, A.; Gołdyn, R. Zooplankton versus phyto- and bacterioplankton in the Maltanski Reservoir (Poland) during an extensive biomanipulation experiment. J. Plankton Res. 2004, 26, 37–48. [Google Scholar] [CrossRef]
- Kozak, A.; Gołdyn, R.; Dondajewska, R.; Kowalczewska-Madura, K.; Holona, T. Changes in phytoplankton and water quality during sustainable restoration of an urban lake used for recreation and water supply. Water 2017, 9, 713. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, C.S.; Huszar, V.; Kruk, C.; Naselli-Flores, L.; Melo, S. Towards a functional classification of the freshwater phytoplankton. J. Plankton Res. 2002, 24, 417–428. [Google Scholar] [CrossRef]
- Abonyi, A.; Leitão, M.; Lançon, A.M.; Padisák, J. Phytoplankton functional groups as indicators of human impacts along the River Loire (France). Hydrobiologia 2012, 698, 233–249. [Google Scholar] [CrossRef]
- Padisak, J.; Crossetti, L.O.; Naselli-Flores, L. Use and misuse in the application of the phytoplankton functional classification: A critical review with updates. Hydrobiologia 2009, 621, 1–19. [Google Scholar] [CrossRef]
- Kozak, A.; Gołdyn, R. Variation in Phyto- and Zooplankton of Restored Lake Uzarzewskie. Pol. J. Environ. Stud. 2014, 23, 1201–1209. [Google Scholar]
- Kowalczewska-Madura, K.; Gołdyn, R.; Dera, M. Spatial and seasonal changes of phosphorus internal loading in two lakes with different trophy. Ecol. Eng. 2015, 74, 187–195. [Google Scholar] [CrossRef]
- Dondajewska, R.; Kozak, A.; Kowalczewska-Madura, K.; Budzyńska, A.; Gołdyn, R.; Podsiadłowski, S.; Tomkowiak, A. The response of a shallow hypertrophic lake to innovative restoration measures—Uzarzewskie Lake case study. Ecol. Eng. 2018, 121, 72–82. [Google Scholar] [CrossRef]
- Gołdyn, R.; Kowalczewska-Madura, K.; Dondajewska, R.; Budzyńska, A.; Domek, P.; Romanowicz, W.; Sengupta, M.; Dalwani, R. (Eds.) Functioning of hypertrophic Uzarzewskie Lake ecosysytem. In Proceedings of Taal 2007: The12th World Lake Conference, Andhra Pradesh, India, 8 July 2008; ILEC: Jaipur, India, 2008; pp. 2233–2236. [Google Scholar]
- Kozak, A. Community Structure and Dynamics of Phytoplankton in Lake Uzarzewskie. Teka Komisji Ochrony Przyrody i Kształtowania Środowiska Przyrodniczego 2009, 6, 146–152. [Google Scholar]
- Kozak, A.; Gołdyn, R. Macrophyte response to the protection and restoration measures of four water bodies. Int. Rev. Hydrobiol. 2016, 101, 1–13. [Google Scholar] [CrossRef]
- Budzyńska, A.; Gołdyn, R.; Zagajewski, P.; Dondajewska, R.; Kowalczewska-Madura, K. The dynamics of a Planktothrix agardhii population in a shallow dimictic lake. Oceanol. Hydrobiol. Stud. 2009, 38, 7–19. [Google Scholar]
- Gołdyn, R.; Podsiadłowski, S.; Dondajewska, R.; Kozak, A. The sustainable restoration of lakes—Towards the challenges of the Water Framework Directive. Ecohydrol. Hydrobiol. 2014, 14, 68–74. [Google Scholar] [CrossRef]
- Utermőhl, H. Zur Vervollkommnung der quantitativen Phytoplankton-Methodik. Mitteilungen Internationale Vereinigung Theoretische und Angewandte Limnologie 1958, 9, 1–38. [Google Scholar]
- Hillebrand, H.; Dürselen, C.-D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination, version 4.5; Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Kozak, A.; Gołdyn, R.; Dondajewska, R. Phytoplankton composition and abundance in restored Maltanski Reservoir under the influence of physico-chemical variables and zooplankton grazing pressure. PLoS ONE 2015, 10, e0124738. [Google Scholar] [CrossRef] [Green Version]
- Matsumura-Tundisi, T.; Tundisi, J. Plankton richness in a eutrophic reservoir (Barra Bonita Reservoir, SP, Brazil). Hydrobiologia 2005, 542, 367–378. [Google Scholar] [CrossRef]
- Connell, J. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1304–1310. [Google Scholar] [CrossRef] [Green Version]
- Padisak, J.; Reynolds, C.S.; Sommer, U. (Eds.) Intermediate Disturbance Hypothesis in Phytoplankton Ecology; Developments in Hydrobiology 81; Springer: Berlin/Heidelberg, Germany, 1991. [Google Scholar]
- Li, Z.; Wang, S.; Guo, J.; Fang, F.; Gao, X.; Long, M. Responses of phytoplankton diversity to physical disturbance under manual operation in a large reservoir, China. Hydrobiologia 2012, 684, 45–56. [Google Scholar] [CrossRef]
- Demir, A.M.; Fakioğlu, Ö.; Dural, B. Phytoplankton functional groups provide a quality assessment method by the Q assemblage index in Lake Mogan (Turkey). Turk. J. Bot. 2014, 38, 169–179. [Google Scholar] [CrossRef]
- Soylu, E.N.; Gönülol, A. Functional Classification and Composition of Phytoplankton in Liman Lake. Turk. J. Fish. Aquat. Sci. 2010, 10, 53–60. [Google Scholar] [CrossRef]
- Maraşlıoğlu, F.; Gönülol, A. Phytoplankton Community, Functional Classification and Trophic State Indices of Yedikır Dam Lake (Amasya). J. Biol. Environ. Sci. 2014, 8, 133–141. [Google Scholar]
- Reynolds, C.S. The Ecology of Freshwater Phytoplankton; Cambridge University Press: Cambridge, UK, 1984. [Google Scholar]
- Napiórkowska-Krzebietke, A. Diversity and dynamics of phytoplankton in lakes Licheńskie and Slesińskie in 2004–2005. Arch. Pol. Fish. 2009, 17, 253–265. [Google Scholar] [CrossRef]
- Lenard, T.; Ejankowski, W.; Poniewozik, M. Responses of Phytoplankton Communities in Selected Eutrophic Lakes to Variable Weather Conditions. Water 2019, 11, 1207. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, C.S.; Descy, J.-P. The production, biomass and structure of phytoplankton in large rivers. Large Rivers 1996, 10, 161–187. [Google Scholar] [CrossRef]
- Reynolds, C.S.; Descy, J.P.; Padisak, J. Are phytoplankton dynamics in rivers so different from those in shallow lakes? Hydrobiologia 1994, 289, 1–7. [Google Scholar] [CrossRef]
- Dondajewska, R.; Kowalczewska-Madura, K.; Gołdyn, R.; Kozak, A.; Messyasz, B.; Cerbin, S. Long-term water quality changes as a result of a sustainable restoration-a case study of Dimictic Lake Durowskie. Water 2019, 11, 616. [Google Scholar] [CrossRef] [Green Version]
- Dondajewska, R.; Kozak, A.; Rosińska, J.; Gołdyn, R. Water quality and phytoplankton structure changes under the influence of effective microorganisms (EM) and barley straw–Lake restoration case study. Sci. Total Environ. 2019, 660, 1355–1366. [Google Scholar] [CrossRef]
- Grabowska, M.; Mazur-Marzec, H. The influence of hydrological conditions on phytoplankton community structure and cyanopeptide concentration in dammed lowland river. Environ. Monit. Assess. 2016, 188, 488. [Google Scholar] [CrossRef] [PubMed]
- Herrero, A.; Muro-Pastor, A.M.; Flores, E. Nitrogen control in cyanobacteria. J. Bacteriol. 2001, 183, 411–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, A.; Flores, E. Genetic responses to carbon and nitrogen availability in Anabaena. Environ. Microbiol. 2019, 21, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Kozak, A.; Celewicz-Gołdyn, S.; Kuczyńska-Kippen, N. Cyanobacteria in small water bodies: The effect of habitat and catchment area conditions. Sci. Total Environ. 2019, 646, 1578–1587. [Google Scholar] [CrossRef]
- Pełechata, A.; Pełechaty, M.; Pukacz, A. Factors influencing cyanobacteria community structure in Chara-lakes. Ecol. Indic. 2016, 71, 477–490. [Google Scholar] [CrossRef]
- Yatigammana, S.K.; Ileperuma, O.A.; Perera, M.B.U. Water pollution due to a harmful algal bloom: A preliminary study from two drinking water reservoirs in Kandy, Sri Lanka. J. Natl. Sci. Found. Sri Lanka 2011, 39. [Google Scholar] [CrossRef] [Green Version]
- Crossetti, L.O.; Beckerb, V.; de Souza Cardosoc, L.; Rodriguesd, L.R.; da Costad, L.S.; da Motta-Marquesd, D. Is phytoplankton functional classification a suitable tool to investigate spatial heterogeneity in a subtropical shallow lake? Limnologica 2013, 43, 157–163. [Google Scholar] [CrossRef]
- Kozak, A. Seasonal changes occurring over four years in a reservoir’s phytoplankton composition. Pol. J. Environ. Stud. 2005, 14, 437–444. [Google Scholar]
- Kozak, A.; Kowalczewska-Madura, K. Pelagic Phytoplankton of Shallow Lakes in the Promno Landscape Park. Pol. J. Environ. Stud. 2010, 19, 587–592. [Google Scholar]
- Kozak, A.; Rosińska, R.; Gołdyn, R. Changes in the phytoplankton structure due to prematurely limited restoration treatments. Pol. J. Environ. Stud. 2018, 27, 1097–1103. [Google Scholar] [CrossRef]
- Paerl, H.W.; Gardner, W.S.; Havens, K.E.; Joyner, A.R.; McCarthy, M.J.; Newell, S.E.; Qin, B.; Scott, J.T. Mitigating cyanobacterial harmful algal blooms in aquatic ecosystems impacted by climate change and anthropogenic nutrients. Harmful Algae 2016, 54, 213–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantzouki, E.; Campbell, J.; Loon, E.V.; Visser, P.; Konstantinou, I.; Antoniou, M.; Giuliani, G.; Machado-Vieira, D.; Cromie, H.; Ibelings, B.W.; et al. A European Multi Lake Survey dataset of environmental variables, phytoplankton pigments and cyanotoxins. Sci. Data 2018, 5, 180226. [Google Scholar] [CrossRef] [Green Version]
- Mantzouki, E.; Lürling, M.; Fastner, J.; De Senerpont Domis, L.; Wilk-Woźniak, E.; Koreivienė, J.; Seelen, L.; Teurlincx, S.; Carey, C.C.; Ibelings, B.W.; et al. Temperature effects explain continental scale distribution of cyanobacterial toxins. Toxins 2018, 10, 156. [Google Scholar] [CrossRef] [Green Version]
- Pálffy, K.; Vőrős, L. Phytoplankton functional composition shows higher seasonal variability in a large shallow lake after a eutrophic past. Ecosphere 2019, 10, e02684. [Google Scholar] [CrossRef]
- Kozak, A. Changes in the structure of phytoplankton in the lowest part of the Cybina River and the Maltański Reservoir. Oceanol. Hydrobiol. Stud. 2010, 39, 85–94. [Google Scholar] [CrossRef]
- Budzyńska, A.; Gołdyn, R. Domination of invasive Nostocales (Cyanoprokaryota) at 52° N latitude. Phycol. Res. 2017, 65, 322–332. [Google Scholar] [CrossRef]
- Budzyńska, A.; Rosińska, J.; Pełechata, A.; Toporowska, M.; Napiórkowska-Krzebietke, A.; Kozak, A.; Messyasz, B.; Pęczuła, W.; Kokociński, M.; Szeląg-Wasielewska, E.; et al. Environmental factors driving the occurence of the invasive cyanobacterium Sphaerospermopsis aphanizomenoides (Nostocales) in temperate lakes. Sci. Total Environ. 2019, 650, 1338–1347. [Google Scholar] [CrossRef]
- Hsieh, C.H.; Ishikawa, K.; Sakai, Y.; Ishikawa, T.; Ichise, S.; Yamamoto, Y.; Kuo, T.C.; Park, H.D.; Yamamura, N.; Kumagai, M. Phytoplankton community reorganization driven by eutrophication and warming in Lake Biwa. Aquat. Sci. 2010, 72, 467–483. [Google Scholar] [CrossRef]
- De Senerpont Domis, L.N.; Elser, J.J.; Gsell, A.S.; Huszar, V.L.; Ibelings, B.W.; Jeppesen, E.; Kosten, S.; Mooij, W.M.; Roland, F.; Sommer, U.; et al. Plankton dynamics under different climatic conditions in space and time. Freshw. Biol. 2012, 58, 463–482. [Google Scholar] [CrossRef]
- Lürling, M.; Eshetu, F.; Faassen, E.J.; Kosten, S.; Huszar, V.L.M. Comparison of cyanobacterial and green algal growth rates at different temperatures. Freshw. Biol. 2012, 58, 1–8. [Google Scholar] [CrossRef]
- Singh, S.P.; Singh, P. Effect of temperature and light on the growth of algae species: A review. Renew. Sustain. Energy Rev. 2015, 50, 431–444. [Google Scholar] [CrossRef]
- Donald, D.B.; Bogard, M.J.; Finlay, K.; Bunting, L.; Leavitt, P.R. Phytoplankton-Specific Response to Enrichment of Phosphorus-Rich Surface Waters with Ammonium, Nitrate, and Urea. PLoS ONE 2013, 8, e53277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmaso, N. Ecological patterns of phytoplankton assemblages in Lake Garda: Seasonal, spatial and historical features. J. Limnol. 2002, 61, 95–115. [Google Scholar] [CrossRef]
- Sommer, U. The periodicity of phytoplankton in Lake Constance (Bodensee) in comparison to other deep lakes of central Europe. Hydrobiologia 1986, 138, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Tapolczai, K.; Anneville, O.; Padisák, J.; Salmaso, N.; Morabito, G.; Zohary, T.; Tadonléké, R.; Rimet, F. Occurrence and mass development of Mougeotia spp. (Zygnemataceae) in large, deep lakes. Hydrobiologia 2015, 745, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Kowalczewska-Madura, K.; Dondajewska, R.; Gołdyn, R.; Podsiadłowski, S. The influence of restoration measures on phosphorus internal loading from the sediments of a hypereutrophic lake. Environ. Sci. Pollut. Res. 2017, 24, 14417–14429. [Google Scholar] [CrossRef] [Green Version]
- Kowalczewska-Madura, K.; Dondajewska, R.; Gołdyn, R.; .Kozak, A.; Messyasz, B. Internal Phosphorus Loading from the Bottom Sediments of a Dimictic Lake During Its Sustainable Restoration. Water Air Soil Pollut. 2018, 229, 280. [Google Scholar] [CrossRef] [Green Version]
- Moustaka-Gouni, M.; Michaloudi, E.; Kormas, K.A.; Katsiapi, M.; Vardaka, E.; Genitsaris, S. Plankton changes as critical processes for restoration plans of lakes Kastoria and Koronia. Eur. Water 2012, 40, 43–51. [Google Scholar]
- Rosińska, J.; Kozak, A.; Dondajewska, R.; Gołdyn, R. Cyanobacteria blooms before and during the restoration process of a shallow urban lake. J. Environ. Manag. 2017, 198, 340–347. [Google Scholar] [CrossRef]
- Rosińska, J.; Kozak, A.; Dondajewska, R.; Kowalczewska-Madura, K.; Gołdyn, R. Water quality response to sustainable restoration measures-case study of urban Swarzędzkie Lake. Ecol. Indic. 2018, 84, 437–449. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Group | The Most Abundant Representatives |
|---|---|
| A | Acanthoceras zachariasii, Rhizosolenia longiseta |
| C | Asterionella formosa, Cyclotella meneghiniana, Stephanodiscus hantzschii |
| D | Nitzschia acicularis, Ulnaria acus |
| E | Dinobryon sp., Mallomonas sp. |
| F | Elakatothrix gelatinosa, Keratococcus sp., Oocystis spp. |
| G | Eudorina sp., Pandorina morum |
| H1 | Aphanizomenon gracile, Dolichospermum planctonicum, D. viguieri |
| J | Actinastrum hantzschii, Coelastrum astroideum, Crucigenia tetrapedia, Pediastrum sp., Scenedesmus sp., Tetraëdron sp., Tetrastrum sp., |
| K | Aphanocapsa sp. |
| LM | Microcystis aeruginosa, M. wessenbergii, Ceratium hirundinella |
| Lo | Peridiniopsis cunningtonii, Peridinium aciculiferum |
| MP | Achnanthes sp., Amphora ovalis, Nitzchia palea, Gomphonema sp. |
| N | Cosmarium bioculatum, Staurastrum sp. |
| NP | Ulnaria ulna |
| P | Closterium aciculare, Closterium acutum, Fragilaria crotonensis |
| S1 | Planktolyngbya limnetica, Limnothrix redekei, Planktothrix agardhii |
| S2 | Raphidiopsis raciborskii |
| SN | Dolichospermum compactum, Raphidiopsis sp. |
| T | Binuclearia lauterbornii, Mougeotia sp. |
| U | Uroglena sp. |
| W1 | Phacus longicauda, Euglena sp. ,Lepocinclis acus |
| W2 | Trachelomonas hispida |
| WS | Synura uvella |
| X1 | Ankistrodesmus gracilis, Monoraphidium irregulare, M. contortum, M. minutum, Schroederia setigera |
| X2 | Rhodomonas lacustris, Bicoeca planktonica, Ochromonas sp. |
| X3 | Chrysochromulina parva, Chrysococcus sp. |
| XPh | Phacotus lenticularis |
| Y | Cryptomonas marssonii, C. ovata, C. reflexa |
| Variable | Var.N | LambdaA | p | F |
|---|---|---|---|---|
| Temp. | 6 | 0.11 | 0.002 | 29.98 |
| EC | 8 | 0.05 | 0.002 | 14.56 |
| NO2 | 12 | 0.02 | 0.002 | 5.39 |
| PO4 | 17 | 0.02 | 0.002 | 5.16 |
| pH | 7 | 0.01 | 0.032 | 2.75 |
| NH4 | 11 | 0.01 | 0.040 | 2.27 |
| NO3 | 13 | 0.01 | 0.056 | 2.09 |
| Variable | Var.N | LambdaA | p | F |
|---|---|---|---|---|
| NH4 | 11 | 0.08 | 0.002 | 14.49 |
| J | 58 | 0.05 | 0.004 | 10.11 |
| S1 | 66 | 0.05 | 0.004 | 8.69 |
| C | 52 | 0.02 | 0.020 | 4.75 |
| NO3 | 13 | 0.02 | 0.030 | 4.19 |
| Y | 78 | 0.02 | 0.030 | 4.50 |
| No. taxa | 83 | 0.02 | 0.030 | 4.10 |
| NO2 | 12 | 0.02 | 0.050 | 3.51 |
| T | 69 | 0.01 | 0.090 | 3.25 |
| Lo | 61 | 0.01 | 0.120 | 2.21 |
| X3 | 76 | 0.01 | 0.206 | 1.57 |
| E | 54 | 0.01 | 0.108 | 2.10 |
| WS | 73 | 0.01 | 0.292 | 1.14 |
| D | 53 | 0.00 | 0.290 | 1.14 |
| P | 65 | 0.01 | 0.390 | 0.88 |
| PO4 | 17 | 0.00 | 0.560 | 0.50 |
| H1 | 57 | 0.00 | 0.684 | 0.31 |
| X2 | 75 | 0.00 | 0.902 | 0.08 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozak, A.; Budzyńska, A.; Dondajewska-Pielka, R.; Kowalczewska-Madura, K.; Gołdyn, R. Functional Groups of Phytoplankton and Their Relationship with Environmental Factors in the Restored Uzarzewskie Lake. Water 2020, 12, 313. https://doi.org/10.3390/w12020313
Kozak A, Budzyńska A, Dondajewska-Pielka R, Kowalczewska-Madura K, Gołdyn R. Functional Groups of Phytoplankton and Their Relationship with Environmental Factors in the Restored Uzarzewskie Lake. Water. 2020; 12(2):313. https://doi.org/10.3390/w12020313
Chicago/Turabian StyleKozak, Anna, Agnieszka Budzyńska, Renata Dondajewska-Pielka, Katarzyna Kowalczewska-Madura, and Ryszard Gołdyn. 2020. "Functional Groups of Phytoplankton and Their Relationship with Environmental Factors in the Restored Uzarzewskie Lake" Water 12, no. 2: 313. https://doi.org/10.3390/w12020313
APA StyleKozak, A., Budzyńska, A., Dondajewska-Pielka, R., Kowalczewska-Madura, K., & Gołdyn, R. (2020). Functional Groups of Phytoplankton and Their Relationship with Environmental Factors in the Restored Uzarzewskie Lake. Water, 12(2), 313. https://doi.org/10.3390/w12020313