1. Introduction
Shallow and small aquatic environments, whose functioning may differ from better recognized ecosystems of lakes, consist of many different types, which include not only typical ponds, but also small ditches, relatively permanent standing or slow-flowing drainage canals, temporary pools, or puddles. Despite being prone to anthropogenic stress in the direct catchment area [
1,
2], ponds often create a refuge for diverse organisms [
2]. The structure of the community and diversity of aquatic organisms can reflect environmental conditions [
3]. Due to the high susceptibility of zooplankton to changes in the environment, especially to human impact in the catchment area of aquatic systems, these animals are often used to determine a variety of environmental features [
4,
5]. Knowledge of zooplankton response to changes is therefore very practical and necessary for both the assessment of habitat quality as well as the establishment of criteria for conservation.
Observing a constant global change of natural environment there is a pressing need to understand, and also assess, the speed and direction of changes. Therefore, some monitoring tools must be worked out and the application of biological communities can serve this purpose. Zooplankton are known to be sensitive to changes in the environment; thus, they have been proposed to be a very valuable indicator of water quality [
6], including the trophic state of water [
7]. I presume that it can also identify anthropogenic activity, contributing to varying levels of human-induced stress in the direct catchment of a pond, in water bodies located within the agricultural landscape [
8,
9].
Not only single species, but also communities of organisms can be a valid tool for monitoring changes in the ecosystem. Thus, for examining the response of zooplankton diversity to changes in environmental parameters, I employed biocoenotic features based on diversity indices (number of species and Shannon–Weaver index) in order to find out which method of diversity analysis will be more suitable for assessing the level of human impact. A biodiversity-based approach is very effective, especially for analyzing the total number of species in a community and/or measurement based on biodiversity indexes. The Shannon–Weaver index is one of the most commonly used in the examination of both the terrestrial environment and freshwater systems [
10,
11]. This is why these two approaches were taken into account in this study carried out on small water bodies.
Another approach that can be used for environmental assessment is the application of groups of organisms, which play a similar role in the ecosystem [
12]. Studying functional categories can refer to niche occupation or habitat requirements, such as ecological groups of zooplankton. Pelagic and littoral species are those that are evolutionary adapted to living in the open water zone, or among macrophytes, respectively. The first group possesses specific adaptations to living in the homogenous conditions of the open water, where they can also be particularly prone to high numbers of potential predators. The second group is well adapted, both morphologically and metabolically, to living in a heterogenic area of macrophytes. Littoral species evolved strategies that enable them to move in often very complex habitats, but also have to forage for specific food resources in a highly qualified way [
13]. They must also cope with unfavorable abiotic conditions that often prevail among plant beds [
14], and this group of species is expected to greatly contribute to the overall diversity.
Using the data set from a survey of 296 small water bodies, I explore the responses of zooplankton community metrics (rotifers, cladocerans, and copepods), referring to the diversity and occurrence of two ecological groups (pelagic and littoral), to limnological variables, including abiotic and biotic features, and also to the site effect (macrophyte site occurrence and biometric features of a plant habitat). This is done to compare the effectiveness of these two approaches for distinguishing the effect of anthropogenic pressure in the catchment area of ponds located within an agricultural landscape.
Along with mid-field woodlots, ditches, oxbows, and wetlands—but also small rivers and streams—ponds are ecosystems of great importance for the preservation and enrichment of biodiversity. They provide an optimal habitat for various plants and animals [
2], often being a source of rare and unique species, e.g., oligochaets, molluscs, beetles, gastropods, large branchiopods, annelids, as well as algae species and zooplankton [
11,
15]. A great range of habitats, especially created by hydromacrophytes, make ponds a valuable aquatic ecosystem, and despite their small area and depth, they will particularly contribute to the enrichment of biodiversity since favorable life conditions for organisms representing various life strategies [
16] are created. Macrophyte beds offer pelagic zooplankters concealment from predators, both planktivorous fish and invertebrates, and provide littoral species with a number of various niches, which altogether increase total biodiversity. Plant build reflects the degree of spatial complexity and at the same time the availability of ecological niches for inhabiting organisms. Therefore, usually the most complex elodeids host the most diverse communities [
17,
18,
19] compared to architecturally simpler stands of helophytes or nymphaeids. Thus, I hypothesized that species diversity will be highest in the habitats of the greatest complexity understood as plant density. There is also a high potential that along with the increase of ecological niches, due to variation in macrophyte cover, more species of high conservation value, such as those that are rare or infrequent on a local, regional, or a worldwide scale [
20,
21,
22], will occur.
Rare species are often attributed to the littoral area [
10]. Macrophyte-dominated zones hosting complex zooplankton assemblages may be found in any type of pond, as well as in anthropogenically-modified water bodies. Some researchers [
23], examining only eutrophic ponds, have stated that extensive, often monospecific macrophyte stands, will provide an environment suitable not only for species associated with macrophytes, but also for migrating pelagic species; thus, maintaining high biodiversity of heleoplankton. A good example of a vegetation zone that is ascertained even in degraded areas, and can be an indicator of high trophic conditions, is
Ceratophyllum demersum stands [
24,
25]. It can therefore be expected that rare species should occur not only in more natural ponds, but also in those with human-induced perturbation, although high anthropogenic impact may be a modifying force responsible for the occurrence of certain species or their prevalence. Therefore, I predicted that in two types of ponds, distinguished on the basis of anthropogenic impact, different frequencies of such species will be found where traces of human impact are stronger. It can also be presumed that more of such species will occur in ponds of low human impact.
The main purpose of the study was to define the response of zooplankton to human-induced transformation, where two groups of ponds were distinguished: (1) Ponds of low human alteration (LowHI) and (2) Ponds of high human alteration (HighHI). I aimed to define whether zooplankton community indices will be a responsive tool responding to environmental parameters in these two groups of ponds, differing in the level of human stress. This may prove that zooplankton, which is often used for trophic state assessment, can also be implemented to assess the level of anthropogenic impacts in the direct basin of a water body. As the type of catchment area, in terms of the level of agricultural coverage and forestation, can determine the specificity of zooplankton [
8,
9], only field ponds were taken into account in order to limit the effect of type of surroundings on the structure of the occurring organisms.
2. Materials and Methods
Research on the occurrence of zooplankton community indices was carried out in the optimum period of the summer season (July–August), between years 2011 and 2018 on 296 small water bodies situated within four large districts of central-western Poland (Provinces: Dolnośląskie, Kujawsko-Pomorskie, Lubuskie, Wielkopolskie). All of the ponds were located within rural catchment areas with the domination of agricultural land and also meadows, pastures, or within household and rural settlements. The water bodies that were chosen for the examination were divided into two main groups, depending on the level of observed anthropogenic impact in the direct catchment area. The first group consisted of ponds of a high level of anthropogenic transformation—HighHI (e.g., ponds situated within reach of rural settlements, such as farm buildings, cattle and pig farms, or manure storage areas). The second group of ponds was of a low level of anthropogenic transformation— LowHI (e.g., ponds situated within an agricultural landscape of very low human alterations, such as meadows, pastures, or wasteland meadows of extensive types of use). Ponds that were difficult to quantify in one of the groups were not adopted for this examination. Divisions of water bodies, based on a quantitative categorization of water bodies in reference, e.g., to trophic state exist. These methods assess the consequences of negative human impact, which can be measured by, e.g., specific levels of nutrients, primary production, or water transparency. However, in this study the division of two groups of ponds was done by assessing the observed level of anthropogenic impact in the surrounding area and was aimed to find out whether this will be reflected in the abiotic and biotic parameters of water. The studied water bodies also varied in respect to the surface area (between 0.001 and 3.4 ha) and to origin (post-glacial ponds, oxbows, and artificial water bodies, such as clay-pits, peat-pits). All of the studied ponds were shallow: LowHI—mean depth 1.4 m and HighHI—1.2 m. Fish were present in 75% of the investigated water bodies.
In order to provide a full spectrum of species inhabiting the ponds, samples were taken from various habitats. The open water area (total number of samples = 274) and vegetated sites (171) were studied. Among helophytes (59 sites) dominating stands of Phragmites australis (Cav.) Trin. ex Steud, Schoenoplectus lacustris (L.) Palla, Typha angustifolia L., and Typha latifolia L. were recorded; among elodeids (106 sites) Ceratophyllum demersum L., Ceratophyllum submersum L., stoneworts (among them e.g., Chara hispida L. and Chara tomentosa L.), milfoil (Myriophyllum spicatum L. and Myriophyllum verticillatum L.); nymphaeids (6 sites) with Potamogeton natans L.
The unequal number of stations, representing particular types, reflect the natural conditions prevailing in the water bodies, where the open water area was present in almost each water body, while particular vegetated sites occurred less frequently. Four types of stations (the open water area, elodeids, helophytes, nymphaeids) that were undergoing the examination had a similar proportion in reference to two groups of ponds – LowHI vs. HighHI ponds (elodeids—24% and %22% of all stations, helophytes—13% and 11%, nymphaeids—2 and 1%, open water—61% and 66%, respectively).
The method for collecting samples differed, depending on the type of habitat. Zooplankton material, of a volume of 10 L, was collected from among macrophytes with a plexiglass core sampler (∅ 50 mm), going vertically through a plant stand. The open water samples, of a volume of 20 L, were taken with a calibrated vessel. All samples were collected from the surface layer of water (0–30 cm). Collected samples were concentrated using a 45-µm plankton net and finally fixed with 96% ethanol. Specified methods for sample collection were described in previous papers [
26,
27].
Zooplankton was classified into two groups in reference to their evolutionary habitat requirements: pelagic and littoral groups. Within the pelagic group species indicating eutrophic conditions of water (e.g.,
Anuraeopsis fissa (Gosse),
Brachionus angularis Gosse,
Filinia longiseta (Ehrenberg),
Keratella cochlearis f.
tecta (Gosse),
Keratella quadrata (Müller),
Pompholyx sulcata Hudson,
Trichocerca pusilla (Lauterborn),
Bosmina longirostris (O.F. Müller),
Chydorus sphaericus (O.F. Müller), and
Diaphanosoma brachyurum (Liévin)) were separated, according to [
7,
28,
29]. Moreover, due to the fact that cladocerans size varies greatly, and also because different responses of particular body-size groups can be expected [
30], the plankton-associated fraction of this group of animals was divided into small-bodied cladocerans and large-bodied cladocerans. Moreover, rare species for the Polish fauna were established and their frequency was calculated. Only species that reached a high frequency (over 5% of the examined samples) underwent analyses.
Physical features of water (temperature, oxygen saturation, conductivity, and pH) were measured in-situ each time and at each sampling station. Water was also sampled for measurements of nutrient concentration (total phosphorus and dissolved inorganic nitrogen, which was the sum of nitrate, nitrite, and ammonium nitrogen) and chlorophyll a concentration. Detailed methods for nutrient determination and chlorophyll a concentration (corrected for pheopigments) were described in the previous paper [
31].
The biometric character of each macrophyte bed was determined by calculating the stem density (length) and dry plant biomass (mass) occupied by plants in a water column (volume) in 1 L of water. Plant material, collected in triplicate, was obtained by cutting all stems from the surface of an area of 0.0625 m
2 and a height of 0.25 m. For a better description of the procedures connected with plant stand measurements, see [
25].
The identification of rare species occurring in the studied ponds has been made on the basis of literature [
32], where rare and infrequently occurring species are listed.
Species diversity, given as the Shannon–Weaver index [
33], was used to measure the species diversity attributed to each pond and station.
On the basis of detrended correspondence analysis (DCA), which revealed that the gradients of length of species and the environmental variables were shorter than three, but longer than two standard deviations, Redundancy Analysis (RDA) was performed [
34] using CANOCO 4.5 for Windows (Wageningen UR, Netherlands). RDA was applied to analyze zooplankton indices (number of species, Shannon–Weaver index, % participation of pelagic and littoral species in the total rotifer abundance, abundance) distribution patterns in relation to physical-chemical factors (conductivity, chlorophyll
a, dissolved inorganic nitrogen, pH, total phosphorus, Secchi disc transparency), level of anthropogenic transformation, the presence of fish, the level of overshading (percent of pond’s surface coverage created by nymphaeids and pleustophytes) and type of habitat (open water zone and dominating macrophyte species representing particular ecological groups of macrophytes). In order to obtain normality of variance, data used in the RDA analyses were log (x+1) transformed, except for the pH, as it is already a negative logarithm of the hydrogen ion. The statistical significance in RDA was assessed by Monte-Carlo permutation tests (9999 permutations) [
34].
The relationships between environmental factors and the frequency of rare species in the studied water bodies were calculated using the Spearman rank correlation coefficient. The Mann–Whitney test was used in order to determine the effect of anthropogenic transformation in the vicinity of the ponds and the Kruskal–Wallis test to determine the effect of a specific type of habitat on the percent participation of rare species in the zooplankton total abundance in a particular water body. For these two analyses, only rare species, whose general frequency in the examined material was higher than 5%, were taken into consideration for statistical analyses.
3. Results
The study ponds revealed a wide range of measured features in respect to the two groups of small water bodies, distinguished on the basis of human impact in the direct catchment area. Neither their morphology (area), nutrient level, nor the spatial characteristics of their vegetated stands, reflected in the density of macrophyte bed, differed between the two types of ponds. Significantly higher values for HighHI ponds were obtained for pH, oxygen saturation (OS), conductivity, and water surface overshading caused by pleustophytes and nymphaeids. At the same time water transparency measured as Secchi disc visibility was higher in LowHI ponds (
Table 1). Fish were present in three-quarters of the studied ponds and there was a similar share in two types of water bodies (73% in LowHI and 83% in HighHI).
The number of species and Shannon–Weaver index values were significantly higher in LowHI ponds for each zooplankton group. Moreover, LowHI ponds had a significantly higher participation of large pelagic cladocerans and of the littoral fraction of copepods. In all other cases, the percent participation of pelagic and littoral zooplankton was evenly distributed between the two groups of ponds (
Table 1). The pelagic group of zooplankton contained 40% of eutrophic species on average (34% of rotifers and 45% of crustaceans) in the open water area.
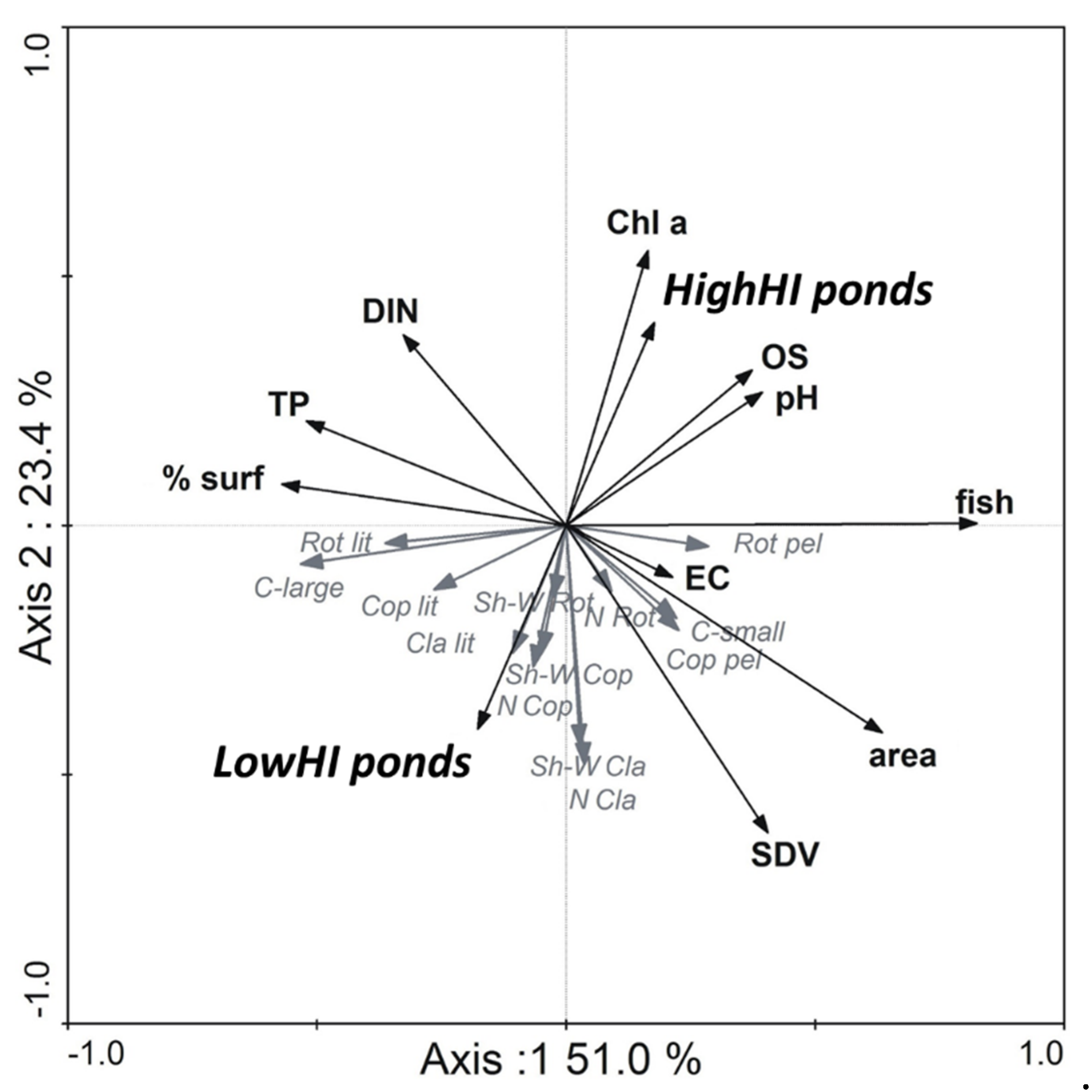
Redundancy analysis (RDA) used to define the relationships between zooplankton indices and abiotic parameters generated two axes, accounting for 51.0% and 23.4%. A total of twelve environmental variables (
Table 2) as well as 13 zooplankton indices were chosen to perform the RDA.
The results of RDA analysis indicated a division of zooplankton indices in reference to environmental features (
Figure 1). The first group gathered around LowHI ponds and consisted of metrics responsible for the high taxonomic structure of crustaceans (increasing number of species of cladocerans and copepods) and of the littoral community indices of all zooplankton groups. Among them, large-bodied cladocerans of pelagic origin (daphnids) also occurred. This group of indices was associated with total phosphorus concentration. Moreover, the rotifer littoral community was also attributed to ponds of a high level of shade caused by the layer of pleustophytes and nymphaeids. At the same time, these community metrics negatively correlated with chlorophyll a concentration and the presence of fish.
The second group of zooplankton features, which was in opposition to LowHI ponds, was clustered close to HighHI ponds. Both rotifer number of species and diversity, as well as densities of the rotifer pelagic community, and that of small pelagic cladocerans and pelagic copepods, revealed the strongest affinity with the presence of fish and high concentrations of chlorophyll a.
There was also an additional group of factors, which had a strong impact on the pelagic community of crustaceans. Both pelagic copepods and a small-sized fraction of pelagic cladocerans were found to be strongly structured by the increasing area of small water bodies as well as by high water transparency, measured by Secchi disc visibility. These communities were associated with waters of low overshading. Environmental parameters, such as water conductivity, pH, and dissolved oxygen content were of minor effect in reference to the zooplankton pelagic community (
Figure 1).
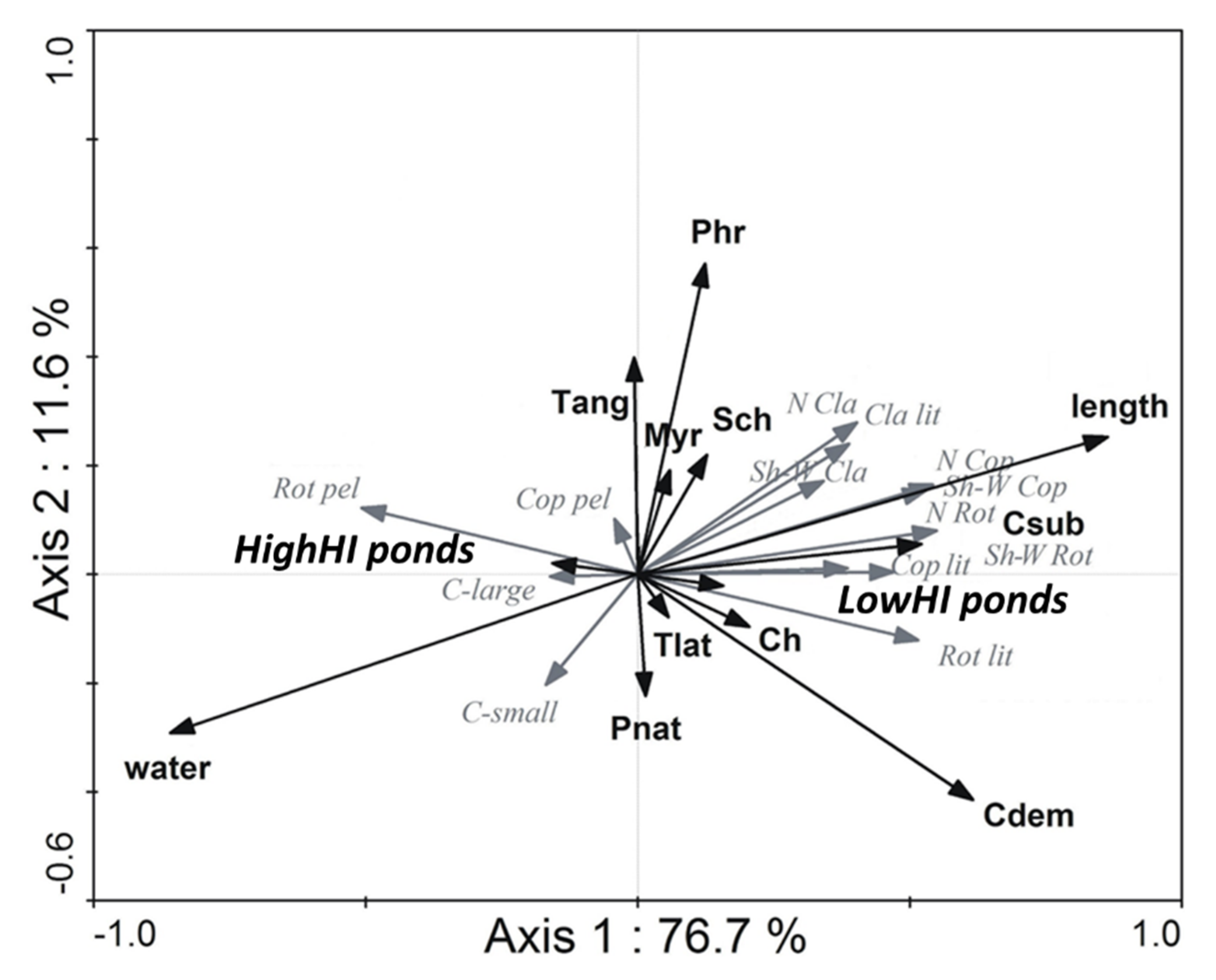
The second RDA analysis with habitat type and biometric features, reflecting the heterogeneity level of a plant stand, (76.7 and 11.6% of explained variance) took into consideration twelve variables (with eight of high significance: open water area, length of macrophyte stems, stands of
Ceratophyllum demersum,
Ceratophyllum submersum,
Potamogeton natans, charophytes, level of anthropogenic transformation) as well as 13 zooplankton indices. This analysis also showed a clear division of zooplankton indices in reference to human-induced changes in the direct catchment area (
Figure 2). The open water area was attributed to HighHI ponds and pelagic rotifers and cladocerans (both the small-sized fraction as well as the large-bodied community) prevailed here. The second group of habitats, consisting of stands located within elodeids, was in close association with LowHI ponds. Community indices of all zooplankton groups, such as number of species, Shannon–Weaver diversity index, share of littoral community, reached their highest values, being particularly associated with the presence of
Ceratophyllum demersum,
Ceratophyllum submersum, and charoids stands. Increasing values of biometric features of macrophyte beds were responsible for the rise of the above-mentioned zooplankton metrics. The presence of helophytes had no effect on zooplankton diversity (
Figure 2;
Table 2).
In the samples 34, rotifer species and 11 of cladocerans were rare or of infrequent occurrence in the Polish fauna. No rare species among copepods were recorded. There were 9 rotifer species (Cephalodella carina Wulfert, Cephalodella gibboides Wulfert, Euchlanis triquetra Ehrenberg, Lecane elsa Hauer, Lecane furcata Murray, Lecane inermis Bryce, Lecane nana Murray, Lecane pyriformis Daday, Trichocerca vernalis Hauer) and 4 cladoceran species (Alona rustica Scott, Chydorus gibbus G.O. Sars, Chydorus ovalis Kurz, Tretocephala ambigua Lilljeborg) that reached a considerably high frequency (they were present in >5% of the samples) in the examined material. The majority of such species were littoral-associated taxa, e.g., representatives of the genus Cephalodella, Euchlanis, Lecane, Lepadella, Alona. However, some of them were also pelagic-associated organisms, e.g., Brachionus falcatus Zacharias, Brachionus polyacanthus (Ehrenberg), Brachionus leydigi Cohn, or Keratella paludosa (Lucks).
Rare rotifers contributed on average 1.46% of the total rotifer abundance, while crustaceans 2.26%. A higher frequency of both rotifers and cladocerans was recorded for LowHI ponds (MW-Z = −3.12; p < 0.01 and MW-Z = −2.49; p < 0.05, respectively).
Looking at the relationships between frequency of rare species and environmental parameters, it was noticed, especially in case of rotifers, that overshading and chlorophyll a concentration (r = 0.22; p < 0.05 and r = 0.22; p < 0.05, respectively) had a positive effect on the prevalence of such species, while level of water oxygenation and conductivity (r = −0.21; p < 0.05 and r = −0.30; p < 0.001, respectively) had a negative effect. The frequency of rare species also positively correlated with the total number of species and values of the Shannon–Weaver index for both rotifers and cladocerans (r = 0.67; p < 0.001, r = 0.36; p < 0.001 and r = 0.33; p < 0.001, r = 0.23; p < 0.01, respectively).
The frequency of both rotifer and cladoceran rare species revealed a significant variation between particular habitats in small water bodies (KW-H = 95.24; p < 0.01 and KW-H = 38.63; p < 0.01, respectively). It was noticed that rotifer rare species were, on average, most frequent among charoids, Ceratophyllum submersum, Potamogeton natans, and Schoenoplectus lacustris, while rare cladocerans most often chose macrophyte sites among Typha angustifolia, Schoenoplectus lacustris, and stoneworts.
4. Discussion
Zooplankton community indices were proved to be a sensitive tool for the identification of environmental features in two different types of ponds, distinguished on the basis of the level of human stress (LowHI vs. HighHI ponds) in the direct catchment area. It was found that both diversity measures, as well as the occurrence of the littoral fraction of zooplankton, revealed a similar response to environmental variables and to habitat type. Taxonomic richness and the Shannon–Weaver index values of each group of zooplankton were significantly higher in LowHI ponds, which were, on average, characterized by higher water transparency and a lower level of conductivity, pH, and OS. Moreover, large-bodied cladocerans and the littoral community of copepods reached higher values in less human-impacted ponds (LowHI). This shows that zooplankton, which is often used for trophic state assessment [
7,
28,
35], can also be implemented to assess the level of anthropogenic impacts in the direct basin of a water body. The less the surroundings of the pond are transformed, the more diverse the zooplankton community that inhabit a pond will be, and there is also a greater chance that large-bodied cladocerans will occur. The occurrence of large cladocerans is often attributed to small and productive ecosystems and the majority of ponds that occurred in the central-western part of Poland were generally eutrophic. This has been proved by the lack of significant variation in the content of nitrogen (DIN) and phytoplankton biomass level measured as chlorophyll a concentration (Chl a), between the two types of ponds selected in this study (LowHI vs. HighHI). Neither did the content of phosphorus (TP), very important for the level of eutrophication, differ between these two groups of ponds (0.58 and 0.53 mg L
−1, respectively). A higher proportion of large cladocerans, such as representatives of the
Daphnia genus (e.g.,
Daphnia pulex) in less human-impacted ponds, contributed to generally better water transparency. Moreover, [
36], who examined a set of 33 interconnected ponds in Belgium, obtained clear differences in the structure of zooplankton communities that were attributed to local biotic and abiotic factors. They observed a variation in the occurrence of large daphnids between the clear-water and turbid state ponds, with ponds in the clear-water state having low fish densities being chosen by large
Daphnia species. Moreover, the clear-water ponds had more species designated as littoral ones. At the same time, their group of ponds in the turbid-water state contained a high abundance of rotifers and cyclopoid copepods. The results of the RDA analysis that I conducted on 296 small water bodies has also confirmed the affinity of zooplankton diversity indices, and large-bodied cladocerans for LowHI, and a high level of shade caused by macrophyte cover built by pleustophytes and nymphaeids. These zooplankton indices were in opposition to the occurrence of fish in ponds. Even though fish had a similar share in the two types of water bodies, it was clear that their presence negatively affected zooplankton diversity measures and the occurrence of large-bodied cladocerans.
Moreover, the littoral fraction of all zooplankton groups was also found to prevail here. Limnetic communities inhabiting the open water areas of small water bodies have always received more scientific attention than communities inhabiting littoral areas. One of the main causes of the neglect of macrophyte-dominated areas in hydrobiological studies is associated with the difficult research methodology and labor-consuming identification of single littoral taxa, such as periphytic and benthic species. This can also be associated with littoral species being overlooked, mainly due to the improper methods applied for sampling [
37], as well as the extremely difficult taxonomic features of animals subjected to determination. However, in this study, a whole littoral community was treated as a group. Such a method, without a need for distinctive species determination, is much easier to implement, and I have proved that littoral zooplankton can serve as a sensitive indicator for identifying a low level of human impact on the aquatic environment.
Unlike the zooplankton diversity indices and the occurrence of littoral community and large-bodied cladocerans, the pelagic fraction of rotifers, copepods, and small-bodied cladocerans occurred in association with different environmental conditions. They were found to prevail in larger ponds with fish presence. Pelagic rotifers and small-bodied cladocerans are equipped with morphological or behavioral anti-predator mechanisms [
38], such as, e.g., strong lorica, long bristles, or they undertake migrations, which help them cope with predation and enable them to develop in water bodies with fish presence. At the same time, these organisms avoided ponds overgrown by macrophytes (pleustophytes and nymphaeids). The pelagic community of zooplankton contains typically eutrophic species, which confirms their prevalence in HighHI ponds. Therefore, it is evident that not only taxon-related indices (e.g., taxonomic composition), which are argued to be a valid instrument in environmental assessment [
39], or functional-based metrics (e.g., feeding-type groups) will indicate changes in the environment [
40]. Thus, it can be recommended that for examining small water bodies’ basic biocoenotic features, such as diversity indices, as well as the share of pelagic and littoral species can be a sensitive tool that responds to varying levels of human stress in a pond’s surroundings.
Because small water bodies usually create a mosaic of microhabitats, where the open water area is often divided by various patches of aquatic vegetation, I determined which habitats favored high zooplankton diversity and the occurrence of certain ecological groups of animals. The second RDA analysis revealed a distinct division of zooplankton metrics depending on anthropogenic transformation in the direct catchment area and the level of habitat heterogeneity, expressed in the length of macrophyte stems. The open water area, associated with HighHI ponds, favored the occurrence of pelagic zooplankton, which contain species of eutrophic origin. The association of areas of open water zones with HighHI ponds is probably connected with significantly lower water transparency in this type of pond compared with LowHI ponds. This is why macrophytes have worse light conditions and do not develop as well as in ponds of lower human impact, where water transparency is higher. At the same time, it had a negative effect on all diversity indices and the littoral community of zooplankton, which found the most advantageous conditions among aquatic vegetation, especially in elodeids. In particular, the presence of stands built by
Ceratophyllum demersum,
Ceratophyllum submersum, and charoids was of significant importance. Stoneworts as a group, despite the varying tolerance between certain species, are very sensitive to water quality and often maintain the stabilization of a clear-water state [
41,
42]. Therefore, the occurrence of charoids was associated with LowHI ponds and their presence positively influenced the diversity increase of zooplankton. Contrary to this, hornworts usually appear in nutrient-rich waters [
43], such as the examined small water bodies located in the agricultural landscape of central-western Poland. Despite the clear niche partitioning of both
Ceratophyllum species, where
C. demersum occurs in clearer waters, and
C. submersum in lower light conditions [
44], both macrophyte species provided a very suitable habitat for diverse zooplankton communities and substratum for the development of littoral organisms. This shows that in case of small water bodies, which are often eutrophic systems, aquatic plants of a high level of complexity expressed by the length of macrophyte stems will contribute to the rise of overall zooplankton diversity, even when these plants are indicators of poor quality of water, such as hornworts. Other plant stands, such as helophytes (e.g.,
Typha latifolia,
Typha angustifolia,
Phragmites australis), which are of much lower structural complexity compared to elodeids [
45,
46], had a much lower impact on zooplankton diversity or the occurrence of the littoral community. Therefore, in order to maintain a high diversity of zooplankton and thereby improve the ecological status of small water bodies, it is recommended that a complex and dense cover of submerged macrophytes be encouraged and sustained.
It is not only the biocoenotic features of zooplankton that can be valuable for detecting the level of anthropogenic impact on a pond ecosystem; the occurrence of rare species can also serve this purpose. A higher frequency of rare species, which favored waters of low conductivity and a high level of overshading caused by the occurrence of pleustophytes and nymphaeids, was found in LowHI ponds. I also found a positive relationship between the total number of rotifer and crustacean species and values of the Shannon–Weaver index with the increasing frequency of rare species. This indicates that increased diversity in the zooplankton community will improve the chances that species of very high conservation value will occur. The occurrence of rare species in areas of a very low level of human transformation has also been ascertained for other small water bodies. The results of a study carried out on 126 ponds distributed over the area of Belgium show that a high share of forest in the immediate surroundings of ponds was associated with the clear water state. These ponds had a higher number of abundant water plant taxa, a higher number of water plant growth forms, and also were characterized by a higher diversity of biotope types [
47]. However, some studies have also presented data that rare species occur in water bodies situated within areas of higher anthropogenic impact, such as farmland or urban areas. Although it has been shown that effective conservation of species diversity in such areas should include typical human-impacted as well as semi-natural habitats, which could both cover the full range of occurring species [
48].
A minority of rare species were of pelagic origin, especially rotifers of the family Brachionidae, e.g., Brachionus falcatus, B. polyacanthus, B. leydigi, or Keratella paludosa, while littoral-associated taxa prevailed. This also confirms the major role of vegetated zones in even very small water bodies in increasing the overall biodiversity, and particularly supporting the occurrence of species of high ecological value.
The results of this study provide further evidence that diversity may play an extremely important role in maintaining the occurrence of rare species. In spite of the fact that macrophyte stations constituted 39% of the whole material in the case of LowHI ponds and 34% in HighHI ponds, rare species were mainly littoral organisms. Therefore, a complexity of habitat is of primary importance for the enrichment of the local fauna and finally for species which do not occur frequently. Analyzing particular habitats, where rare zooplankton was most frequent, it was noticed that they displayed significant preferences towards certain habitats. They chose stations located in elodeids, such as stoneworts (both rotifers and cladocerans) and Ceratophyllum submersum (rotifers), also among helophytes, such as Schoenoplectus lacustris (both rotifers and cladocerans) and Typha angustifolia (cladocerans), and finally nymphaeids such as Potamogeton natans (rotifers). The lack of correlations between biometric features of macrophyte stands, referring to their level of complexity, indicates the fact that rare species will frequently occur in various macrophyte-dominated habitats, irrespective of their build and density. These findings also support the importance of maintaining diverse macrophyte cover even within very small and shallow aquatic systems, such as ponds.





{kind=link}
{kind=link}