1. Introduction
Wastewater treatment is essential for public health and environmental protection. This is a specific process for removing various contaminants from wastewater. The idea is to safely release treated water into the environment or reuse it. In this regard, practical research related to the hydraulic efficiency of green–blue flood control scenarios for vegetated rivers is currently being expanded [
1]. The extent to which wastewater needs to be treated is determined by government water management standards and the specifics of the environment [
2]. The treatment process takes place in wastewater treatment plants or a bioreactor. The main methods used for wastewater treatment are mechanical, biological, and chemical. Biological wastewater treatment is performed under aerobic or anaerobic conditions for biomass growth depending on the specific microorganisms used as well as on the nature and concentrations of the contained pollutants [
3,
4]. The choice of a particular method depends on the type of polluted water and the nature and concentrations of the pollutants.
The main pollutants of industrial and domestic wastewater are plant nutrients, individual or mixtures of synthetic organic chemicals, inorganic chemicals including heavy metals, pathogenic organisms, oil, sediments, radioactive substances, etc. [
5].
Some of the most toxic and common mixtures of synthetic organic chemicals are phenol and its derivatives including resorcinol, hydroquinone, 3-nitrophenol, 2,6-dinitrophenol, 3-chlorophenol,
p-cresol (4-methylphenol), benzene, etc. Numerous scientific studies and practical applications have proven the potential of several microorganisms to biodegrade mixtures of phenol and its derivatives to environmentally friendly substances [
6,
7,
8]. It has been found that very good results in the biodegradation of phenolic compounds are achieved by the strains
Arthrobacter, Aspergillus awamori, Burkholderia, Candida tropicalis, Pseudomonas, Rhodococcus, Trametes hirsute, Trichosporon cutaneum, just to mention a few [
9,
10,
11,
12].
Generally speaking, the kinetics describing microbial growth and biodegradation of phenolic components (substrates) are of great importance for studying the peculiarities of the wastewater treatment process [
13]. Kinetic models are developed on the basis of expert knowledge and laboratory research with specific strains and particular water pollutants. The kinetic models provide opportunities to study various characteristics of the wastewater treatment process and support the successful and effective achievement of the prescribed environmental criteria [
14].
Specific microorganisms’ growth rates in a substrate mixture of two or more phenolic components are known as sum kinetic models (without interaction); multiplication kinetic models; sum kinetic models which incorporate purely competitive substrate kinetics; sum kinetics with interaction parameters (SKIP); elimination capacity-sum kinetics with interaction parameter (EC-SKIP); self-inhibition EC-SKIP (SIEC-SKIP), etc. [
15,
16,
17,
18,
19,
20].
The SKIP models are a successful extension of the sum kinetics models, because they take into account the mutual influence of the components in the substrate mixture [
21,
22]. The SKIP models demonstrate excellent prediction of the biodegradation with different microbial strains of the mixture of phenol and its derivatives: benzene, toluene, and phenol biodegradation by
Pseudomonas putida F1 [
23]; ethylbenzene and styrene by
R. pyridinovorans PYJ-1 [
22]; phenol and
p-cresol mixture degradation by
Aspergillus awamori strain [
24], etc.
In [
25], a mathematical model was proposed for phenol and
p-cresol mixture degradation in a continuously stirred tank bioreactor. The model was presented by a system of three nonlinear ordinary differential equations as follows
where
is the biomass specific growth rate, described by the SKIP (sum kinetics with interaction parameters) model in the form
In the analytic expression of
, the interaction parameters
and
indicate the degree to which the substrates
p-cresol and phenol affect the biodegradation of phenol and
p-cresol, respectively. Two inhibition parameters,
and
, are also included in the biomass specific growth rate
.
The state variables
X,
,
, and the model parameters are described in
Table 1. The numerical values in the last column were validated by laboratory experiments using the
Aspergillus awamori strain and given in [
24].
In this paper, we modify model (
1) by introducing a discrete time delay incorporated into the biomass growth response
The constant
stands for the time delay in the conversion of the consumed substrate into viable biomass. The term
represents the biomass of microorganisms that consumes nutrients at time
and survives in the bioreactor for
units of time, necessary to complete the conversion of substrate into viable biomass [
26,
27].
Bioreactor (chemostat) competition models involving time delay, recently, have been widely discussed and investigated in the literature. According to [
28], the conversion process of nutrients to biomass always requires a fixed length of time and thus the corresponding chemostat model should be described by a system of differential equations with discrete delays. To our knowledge, the first attempt to introduce a discrete time delay was in [
29], where a linear bioreactor model was discussed. Other early models in this regard can be found in [
30], where the effects of time delay and growth rate inhibition in the bacterial treatment of wastewater were discussed, as well as in [
31], where the appearance of a time delay was motivated by the existence of a lag phase in the growth response of microorganisms in the medium. A basic and well known chemostat model incorporating a time delay was proposed and studied in [
32]. This model contained three nonlinear ordinary differential equations, describing competition of two species on a single nutrient, and established the validity of the Competitive Exclusion Principle (CEP). CEP means that the species with the lowest break-even concentration can survive in the chemostat, and it drives other species to extinction. Two-species competition was also considered in [
33], where CEP was proved as well. On the other hand, introducing a time delay explicitly into the model equations can help to explain the transient oscillation behavior of the bioreactor dynamics often observed in practical experiments. Detailed investigations in this direction can be found in [
34] on a ‘single biomass/single substrate’ model.
Recently, various mathematical delayed models involving different numbers of species growing on one or more substrates and involving different kinds of specific growth rates (monotone, such as the Monod law, or nonmonotone, such as the Haldane law, etc.) have been discussed, see [
26,
27,
35] and the references therein. The latter papers consider competition of
n species of microorganisms on a single substrate. In [
26], sufficient conditions are presented which enhance the CEP and thus the global asymptotic stability of the model solutions. This was calculated for general nonmonotone response (specific growth rate) functions. The same time delayed model was modified in [
27] by introducing specific species death rates, different from the dilution rate in the chemostat. Under certain conditions, it was shown that the single species survival equilibrium is globally asymptotically stable. It was demonstrated numerically that the differential removal rates can lead to damped oscillation in the solutions. Similar results were established for the same model in [
35]. There, the competitive exclusion was established by applying the method of Liapunov functionals under some technical assumptions on the biomass response functions.
In this paper, we perform mathematical analysis of the time delayed model (
3)–(5). In
Section 2, we find the equilibrium points of the model and investigate their local asymptotic stability and occurrence of Hopf bifurcations with respect to the delay
. The basic properties of the model solutions as well as the global stabilizability of the model dynamics are presented in
Section 3.
Section 4 includes numerical examples, supporting the theoretical results. Concluding remarks are presented in
Section 5.
2. Equilibrium Points, Local Stability, and Bifurcations
In the mathematical analysis of the model (
3)–(5), we assume that the influent concentrations
and
are constant and consider the dilution rate
D and the delay
as varying parameters.
The equilibrium points of (
3)–(5) are obtained as solutions of the following system of transcendental equations
Obviously, (with ) is an equilibrium point of the model for all and and is called the boundary or washout equilibrium.
In what follows, we look for solutions of (
6)–(8) such that
.
Similar to [
25], multiplying Equation (7) by
, Equation (8) by
, and summing them together leads to
Denoting
we obtain
The latter expression is biologically relevant if and only if
. Using the numerical values in the last column of
Table 1, it follows that this is satisfied:
.
Now, we substitute
from (
11) into the expression of
and obtain the specific growth rate
as a function of
only. Denote for simplicity
or explicitly
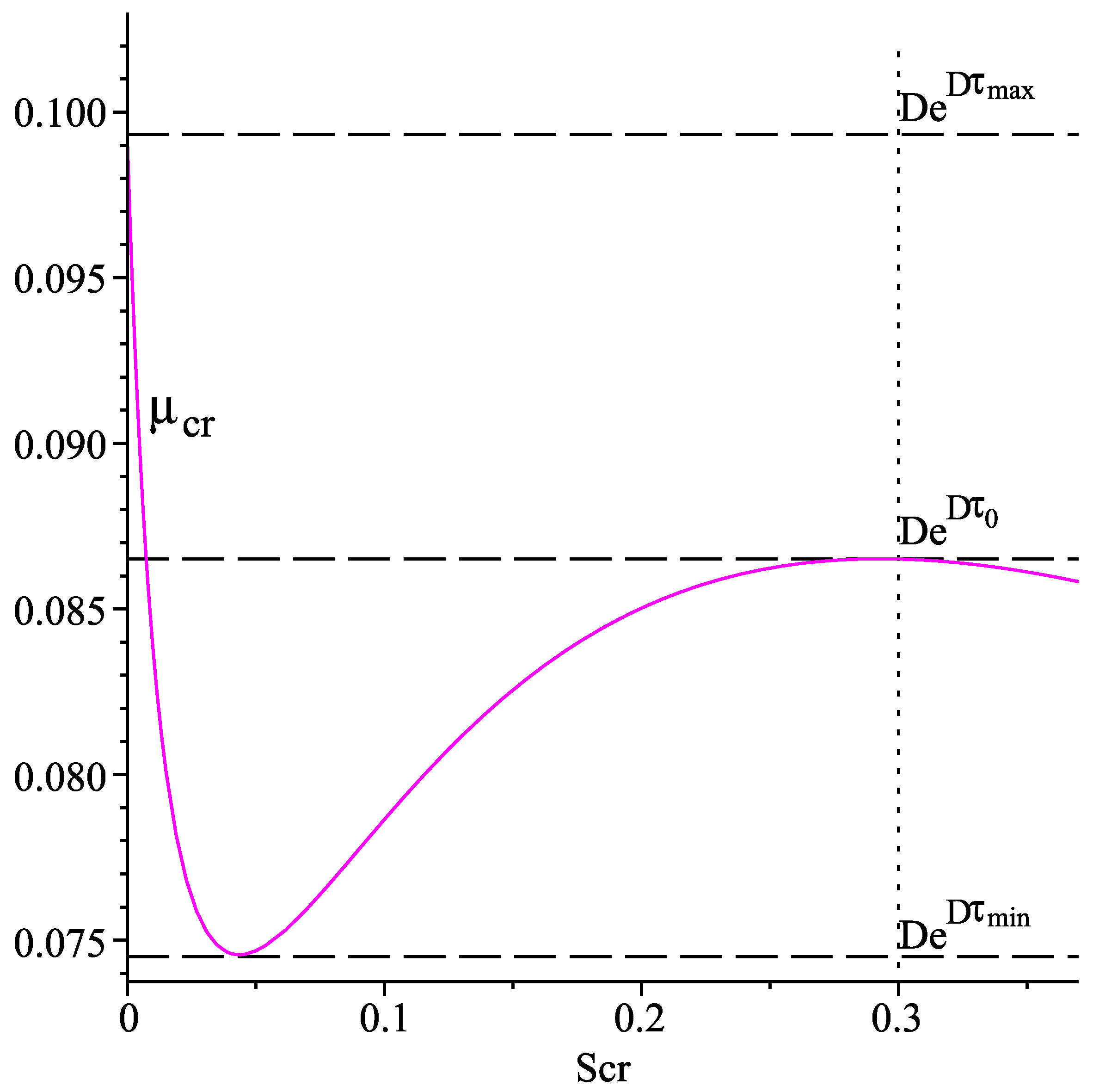
Figure 1 shows the graph of the function
for
, using the numerical values in the last column of
Table 1.
From Equation (
6), we learn that the steady state component with respect to
is the solution of the equation
If there exists a root
of the latter, such that
, then the equilibrium components with respect to
X and
are determined by the formulae
Denote . Obviously, all components of depend on the control parameter D and on the delay .
The graph of suggests the following properties of the latter, which are used in the further investigations.
Properties of
(P1) for all , and .
(P2) There exists a point such that if , and if .
(P3) The following inequalities hold true: .
For any
define
Obviously,
is a decreasing function of
. Moreover, for any
, the following inequalities are fulfilled:
Based on the above considerations we draw the following conclusions:
If
, then there exist two positive roots
,
of Equation (
14) such that
, and
,
.
If
, then there is only one positive root
of Equation (
14) such that
and
.
Therefore, the model (
3)–(5) possesses up to two interior (with positive components) equilibrium points depending on the values of
and
D. Denote them by
where
and
,
, are determined according to (
15) after replacing the
with
and
, respectively.
In what follows, we study the local asymptotic stability of the model equilibrium points. Here, we use the linearization technique for differential equations with a discrete time delay (see [
36]).
The characteristic polynomial corresponding to the Jacobian matrix
J of (
3)–(5) is defined by
, where
is any complex number, and
is the
–identity matrix:
The following presentation holds true
Obviously,
are always solutions of
, i.e., eigenvalues of the Jacobian matrix
evaluated at the equilibrium point
,
. The third eigenvalue
of (
17) is the root of the equation
the latter being evaluated at the components of
,
.
Straightforward calculations show that
where
Therefore, the characteristic Equation (
18) can be rewritten in the form
where
means
.
In the next subsections, we study the local asymptotic stability and existence of Hopf bifurcations of the equilibrium points with respect to the delay .
2.1. The Interior Equilibrium
The interior equilibrium
exists for
and
Proposition 1. For the model (3)–(5), the equilibrium point is locally asymptotically stable for all . Proof. The characteristic Equation (
19) evaluated at
is
Using the fact that
, we obtain
We have in this case
, so that
holds true. Denote
. Then the characteristic Equation (
20) takes the form
Applying Theorem 1.4, Chapter 3 in [
37], it is enough to show that there are no purely imaginary roots
,
, of the latter equation. Indeed, using the presentation
we obtain
Separating the real and the imaginary parts leads to
Squaring both sides in the latter equations and adding them implies
The last inequality shows that there are no pure imaginary roots of (
20). This means that the interior equilibrium
is locally asymptotically stable whenever it exists, i.e., m for all
. The proof is completed. □
2.2. The Interior Equilibrium
The equilibrium
exists for
and
Since
, the characteristic Equation (
19) has the form
Proposition 2. For the model (3)–(5), the equilibrium point is locally asymptotically unstable for all . Proof. Using (
21), denote
Since
and
, this implies that
possesses at least one positive real root, and therefore
is locally asymptotically unstable. □
Below, we study conditions under which the coexistence equilibrium
is nonhyperbolic and undergoes local Hopf bifurcations with respect to the delay
. The investigations use the same techniques as in [
34]. We present them here in the notations of our model (
3)–(5) for completeness and the reader’s convenience. The proofs of the results can be found in
Appendix A.
Denote for convenience
According to Property (P2) of
, it follows that
for
holds true. Then the characteristic Equation (
21) is presented by
We are looking for solutions of (
23) with respect to
of the form
,
. For
,
, and using the equality
, we obtain
Separating the real and the imaginary part of the latter leads to
Further, the equalities
imply
Then we have from (
24)
We are looking for positive solutions with respect to
of equations (
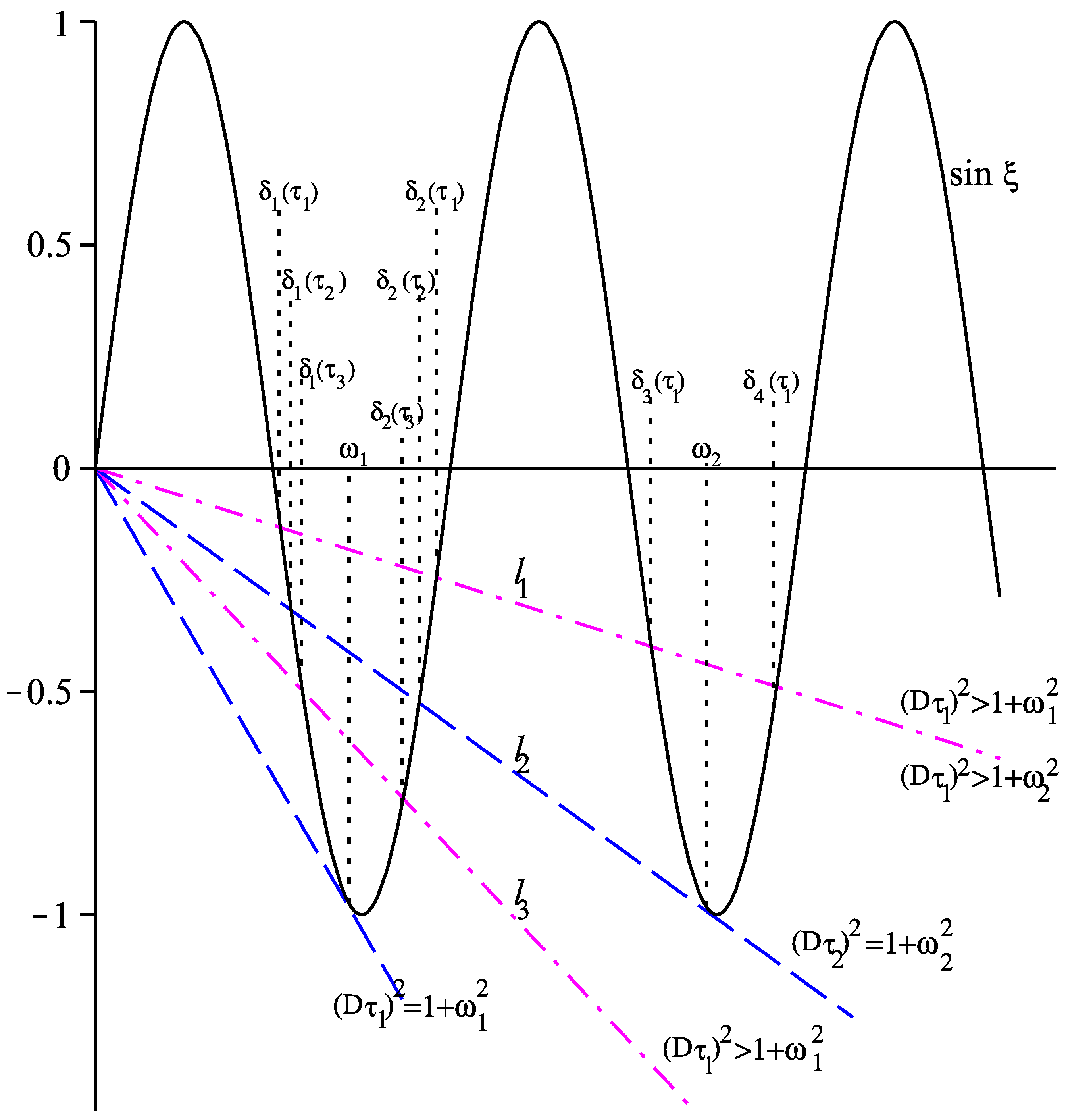
26). As in [
34], we first investigate solutions of
where
. For any fixed
there will be a unique solution of (
27) if the line
is tangent to the curve
. Denote by
this unique solution in the interval
, see
Figure 2. Using the equality
and solving for
, we obtain
. Clearly, if
then (
27) has no solutions in the interval
, and if
then there are exactly two solutions of (
27) in the interval
, one less than
and one larger than
(cf.
Figure 2).
Assume that
for some integer
. Let
be the unique solution of
for
and
be the unique solution of
for
. Then
is strictly decreasing,
is strictly increasing, and (cf. [
34])
Differentiating, with respect to
, the equality
and using the implicit function theorem, we obtain consecutively
Similarly, one obtains
Since
is increasing for
,
, using the relations
we find that
for
. Similarly, one can show that
for
holds true. Therefore,
Define the function
Obviously,
is defined and nonnegative for all
if and only if
holds true. If
for some values of
, then
is not defined on the whole interval
.
The function
plays a significant role in investigating the existence of Hopf bifurcations of
(cf. [
34]). However, since
, it can be used to detect other types of bifurcations of
. Indeed,
If there exists
such that
, then
, i.e.,
. This means that
is eventually a root of the characteristic Equation (
23), so that the equilibrium
becomes nonhyperbolic at
leading to some kind of bifurcation.
Since
(see (
16) and (
22)), it follows that
, and so
. Thus,
can serve as another bifurcation value for the equilibrium
.
Theorem 1. [34] Let be the largest integer such that holds true. Then the following assertions are valid: If is a solution of (26), then the curve intersects one of the curves , , , at . Let intersects or at for some . Then is a solution of (26), if and only if If the solutions ofare isolated, then (26) possesses a finite number of positive solutions. Theorem 1 implies that if and , , do not intersect for , then, the equilibrium is hyperbolic, i.e., does not undergo any bifurcation with respect to .
Theorem 2. [34] Let be a root of the characteristic Equation (23) such that , . Then Corollary 1. [34] Let the assumptions of Theorem 1(i) be satisfied. Then the following assertions are valid: There exists a unique integer j, , , or for some , such that the curves and intersect at .
If or , then The next theorem reports the final result on the existence of local Hopf bifurcations of the interior equilibrium .
Theorem 3. [34] Let be a solution of (26). Then the following assertions hold true: (i) There exists a unique integer such that .
(ii) If , where or for some integer i, , and or , then the equilibrium undergoes a Hopf bifurcation at . All bifurcating periodic solutions are positive and unstable and have periods in the intervals 2.3. The Washout Equilibrium
For
(with
), we obtain from (
19) the following characteristic equation
Proposition 3. For the model (3)–(5), the equilibrium point is locally asymptotically stable for and locally asymptotically unstable for . Proof. Consider the characteristic Equation (
40), and define the function
First, let
, or equivalently,
. We have
This means that there is at least one negative root of the characteristic Equation (
40), thus
is locally asymptotically stable.
In the case when
, i.e., when
, we have
so there is at least one positive root of the characteristic Equation (
40), which means that
is locally asymptotically unstable. This completes the proof. □
Although the washout equilibrium
is not of practical interest, in
Section 3 we shall prove that it is also globally asymptotically stable if
. Global stability of
means total washout of biomass in the reactor and breakdown of the degradation process.
In the same way as in [
34], one can show existence of Hopf bifurcations of
when it is locally unstable, i.e., for
. In this case, due to the zero
X-component, the periodic solutions bifurcating from
cannot be nonnegative. Any periodic solution surrounding
will involve negative and positive values, i.e., the
X-component of any such periodic solution will have a zero in the interval
for certain
and for any
and will change sign there. Such kinds of periodic solutions are not biologically relevant, and we skip the corresponding investigations. The interested reader can consult [
34], as well as [
38], for more information.
3. Global Properties of the Time-Delayed Model Solutions
Consider the time-delayed model (
3)–(5). Denote by
the set of all nonnegative real numbers and by
the Banach space of continuous functions
. Define
and assume that the initial data for model (
3)–(5) belong to
.
According to the theory of functional differential equations (cf. [
36,
37,
39]), for each
there exists a unique solution
of (
3)–(5) defined on
such that
for each
.
Denote
, where
K and
are defined by (
10). After multiplying Equation (5) by
and adding it to Equation (4) we obtain
The latter equality means that
,
, so
. Then, system (
3)–(5) can be written in the form
Since
for any
, and we are interested in the asymptotic behavior of the dynamics, we consider the limiting system
where
, cf. (
12). The initial conditions for (
41) belong to the set
Let us fix an arbitrary
, and denote
. If no confusion arises, we shall also use the simpler notation
, as well as
, instead of
. As mentioned before, the solution
of (
41) is defined and exists for all
.
Theorem 4. If for all , then for all .
Let the following inequalities be fulfilledThen the solution of (41) is positive for all and is uniformly bounded. Proof . is obvious. The plane
is invariant for the model (
41). Therefore, we shall consider solutions
with
for all
.
Assume that
. Then, from the second equation of (
41), we have
This implies that
for all
.
Further, applying the variation-of-constant formula, we obtain
which implies that
for each
.
Denote
Then
The latter equality implies
where
exponentially as
. This means that all nonnegative solutions are uniformly bounded and thus exist for all
. The proof is completed. □
Remark 1. Condition (42) can be explained by the complicated expression of the SKIP model , and, in particular, by the fact that is strongly positive (see Property (P1)). Usually, in bioreactor models, the specific growth rate is assumed to satisfy . In particular, if we assume that are simultaneously fulfilled, i.e., initially the whole mixture of phenol and p-cresol is not available in the bioreactor, then condition (42) is not necessary, and it can be proved as above that all model solutions are strongly positive and uniformly bounded for all time . The equilibrium points
,
, and
of the reduced model (
41) take the form
In what follows, we prove that the interior equilibrium point is globally asymptotically stable whenever it exists, i.e., for .
Lemma 1. Let be a positive solution of (41). If , then there exists time such that for all . Proof. Let
. Assume that there exists time
such that
for all
. Then
thus,
is a strictly decreasing function. Since
is uniformly continuous in
, applying Barbălat’s Lemma [
40], we obtain
Since
and
, the above equality implies that
and
as
. Define the function (cf. Lemma 2.2 in [
26])
Then
Since, in this case,
, we find that
for all sufficiently large
. So, there exists
such that
as
. However, this is impossible according to the definition of
, and because we have already shown that
as
. Hence, there exists a sufficiently large time
such that
. Moreover, if there exists
such that
then
The last inequality shows that
for all
. The proof is completed. □
Lemma 2. Let be a positive solution of (41) and be its interior equilibrium point. DenoteThen and hold true. Proof. Assume that
. Choose an arbitrary
According to Theorem 4 (see (
43)), there exists time
such that for all
the following inequalities hold true
Lemma 1 implies that there exists time
such that
for all
. Since
, there exists
such that
for all
. We set (cf. Lemma 3.5 in [
26])
Then
,
,
for all
, and
From (
46) and the choice of
we obtain
and further, taking into account the monotonicity of
(Property (P2)), it follows
The last equality contradicts (
47), which means that
.
Assume now that
, i.e., that the limit of
exists as
. We shall show that
holds true. By Barbălat’s Lemma, we obtain that
, i.e.,
It follows then that
as
, which means that
as
and thus
Hence, if
, then
holds true.
Now assume that
, i.e., the limit of
exists as
. We shall show that
is fulfilled. Applying Barbălat’s Lemma yields
, i.e.,
which means that
From here, it follows that there exists the limit of
as
, i.e.,
is satisfied.
Next, we show that the equalities
and
are simultaneously fulfilled. Thus, we shall use some ideas from the proofs of Lemma 4.3 in [
26] and Theorem 3.1 in [
27].
Let
be an arbitrary fixed number. The Fluctuation Lemma [
41] implies that there exists a sequence
such that for each
m we have
The equality
leads to
Since
can be arbitrarily small, it follows that
The latter inequality yields
and hence
Similarly, one can show that
. This and (
48) lead to
Applying the Fluctuation Lemma again, there exists a sequence
such that for each
k we have
Then
and therefore
In the same way one can show that
Hence,
holds true. This and (
49) lead to
. Using (
49) again, we find that
is valid. The proof is completed. □
Theorem 5. Let and be an arbitrary element with such that (42) is fulfilled. Then, the corresponding positive solution converges asymptotically towards . Proof. Lemmas 1 and 2 imply that the solution
is convergent as
. Let
,
. Applying Barbălat’s Lemma, we obtain
and hence
From here, it follows that
, because
is the locally asymptotically stable equilibrium for
according to Proposition 1. The proof is completed. □
The next Theorem 6 proves that the locally asymptotically stable washout equilibrium
is also globally asymptotically stable. The proof uses similar ideas to Theorem 2.3 of [
26].
Theorem 6. Let and be an arbitrary element such that (42) is fulfilled. Then the corresponding positive solution of (41) converges asymptotically towards . Proof. Let . According to the definition of , this means that .
Suppose that
for all sufficiently large
t. Then using the function
in (
44) we obtain from (
45)
The last inequality follows from the fact that
is an increasing function for
. Therefore,
as
for some
. Since
is bounded and uniformly continuous, it follows by Barbălat’s Lemma [
40] that
as
, i.e.,
Assume that there is a sequence such that . Then , which implies that , a contradiction. Therefore, for all sufficiently large t.
Assume that
for all sufficiently large
. Then it follows from the second equation of (
41) that
for all large
t, which implies that
for some
. If
, then
because
is decreasing for
. Therefore,
as
for some
. Barbălat’s Lemma then implies that
as
, which means that
leading to
as
. If
, then obviously
. Therefore,
is globally asymptotically stable for the model (
41). The proof is completed. □
4. Numerical Simulation
Here, we illustrate the theoretical results from the previous sections on three numerical examples for different values of the parameters
D and
. We use the numerical values of the model parameters in
Table 1.
We notice that if is fulfilled.
The next computer simulations illustrate the occurrence of Hopf bifurcations of the interior equilibrium
, investigated in
Section 2.2. First we have to determine the tangent point
of
and the line
, which coincides with the unique solution of
, i.e., with the unique solution of
in the interval
for some
(cf. [
34]). We obtain the following values
For this value of
D we obtain
thus,
holds true. According to Theorem 1, we have
, thus, there are two solutions
and
, and the equilibrium
undergoes Hopf bifurcations at the following four values of
:
Figure 3 visualizes the curve
and the solutions
and
as well as the above intersection points.
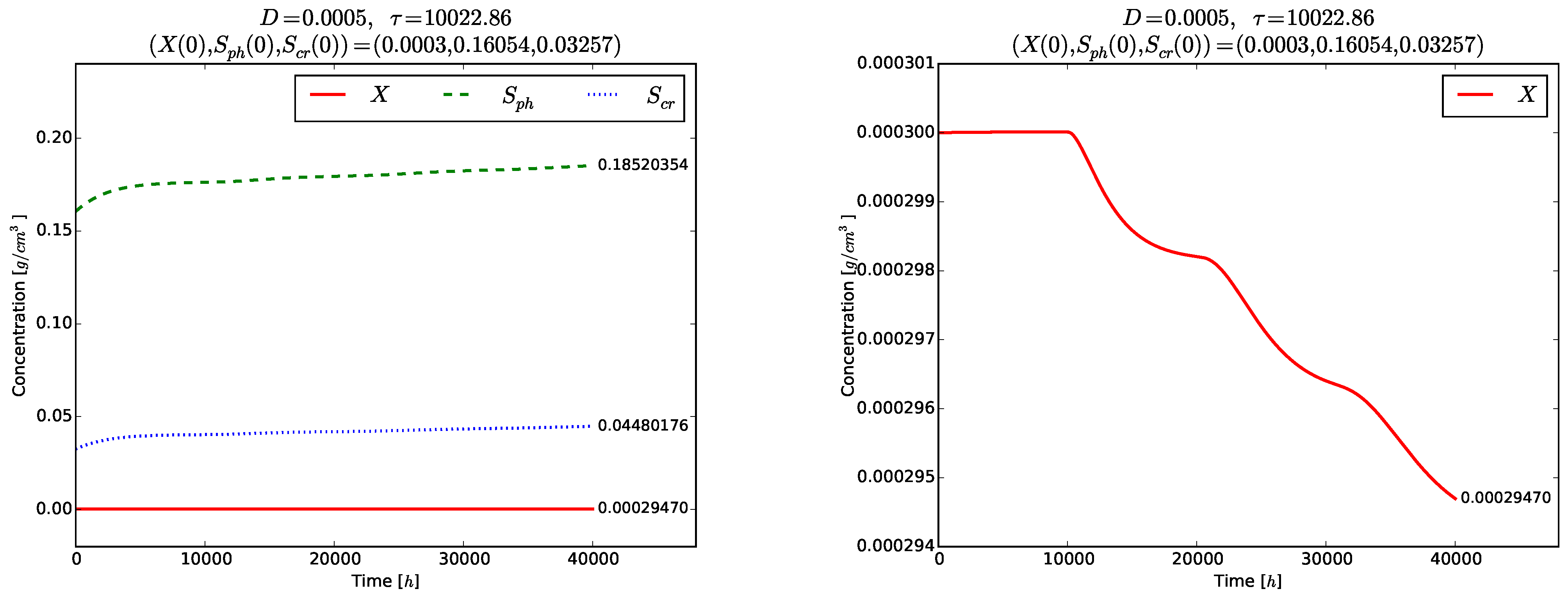
We take
. At this value of the delay, the two interior steady states are
According to Proposition 2, is locally asymptotically unstable for all . Theorem 5 implies that the equilibrium is globally asymptotically stable for .
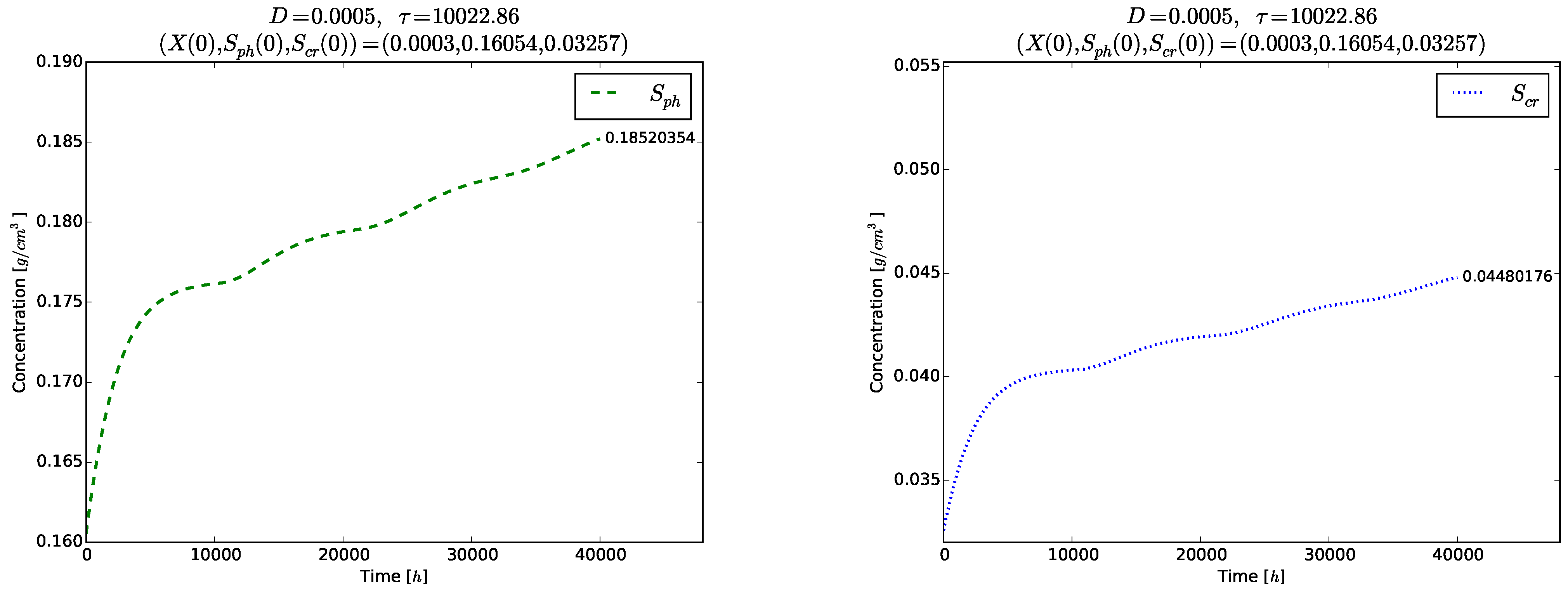
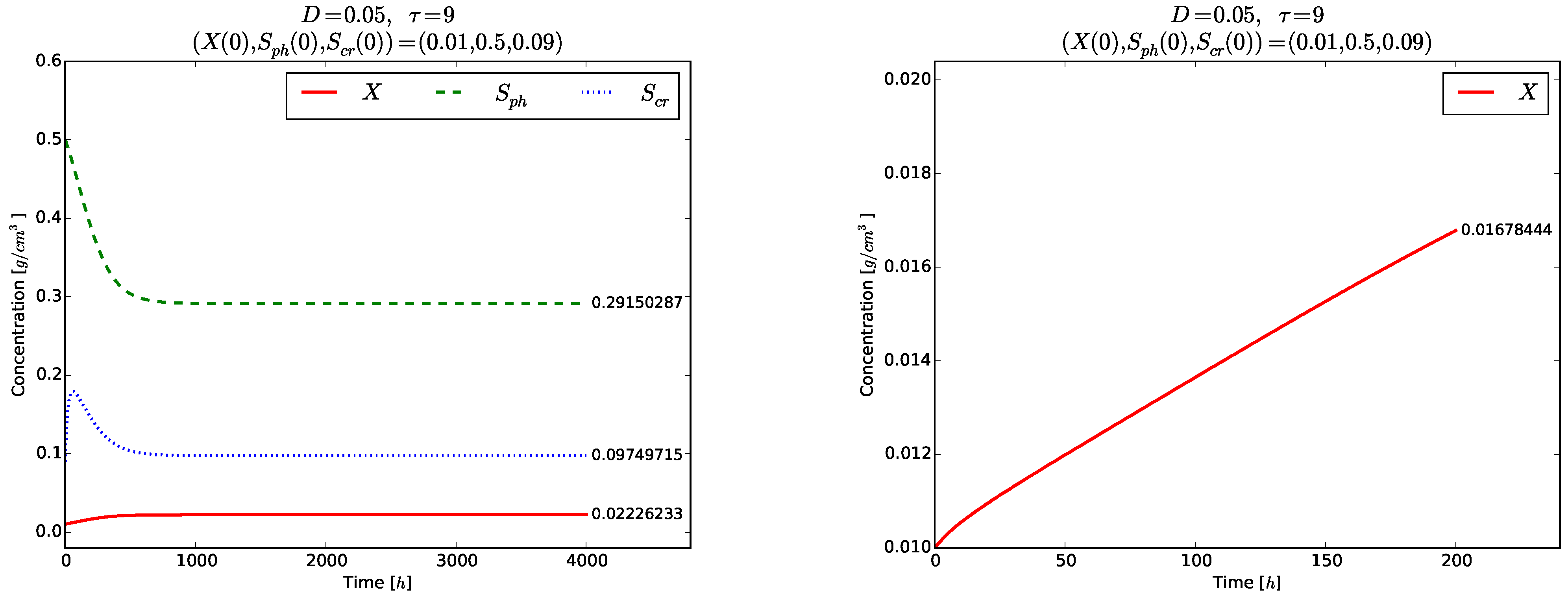
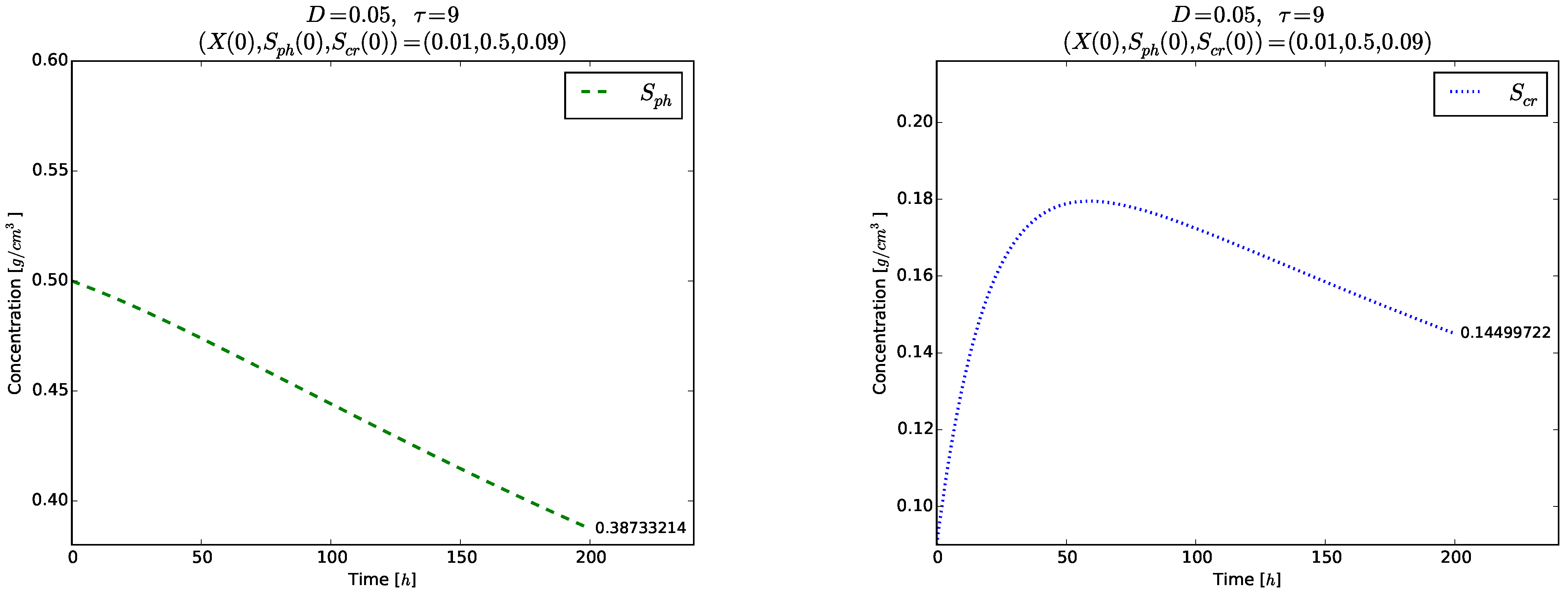
Figure 4 and
Figure 5 show the time evolution of the solutions of the model (
3)–(5). Although the initial conditions
are chosen near to
, the solutions tend to the globally asymptotically stable equilibrium
. Transient oscillations in the time evolution of the phase
,
and
are observed in the right plot of
Figure 4 and in
Figure 5.
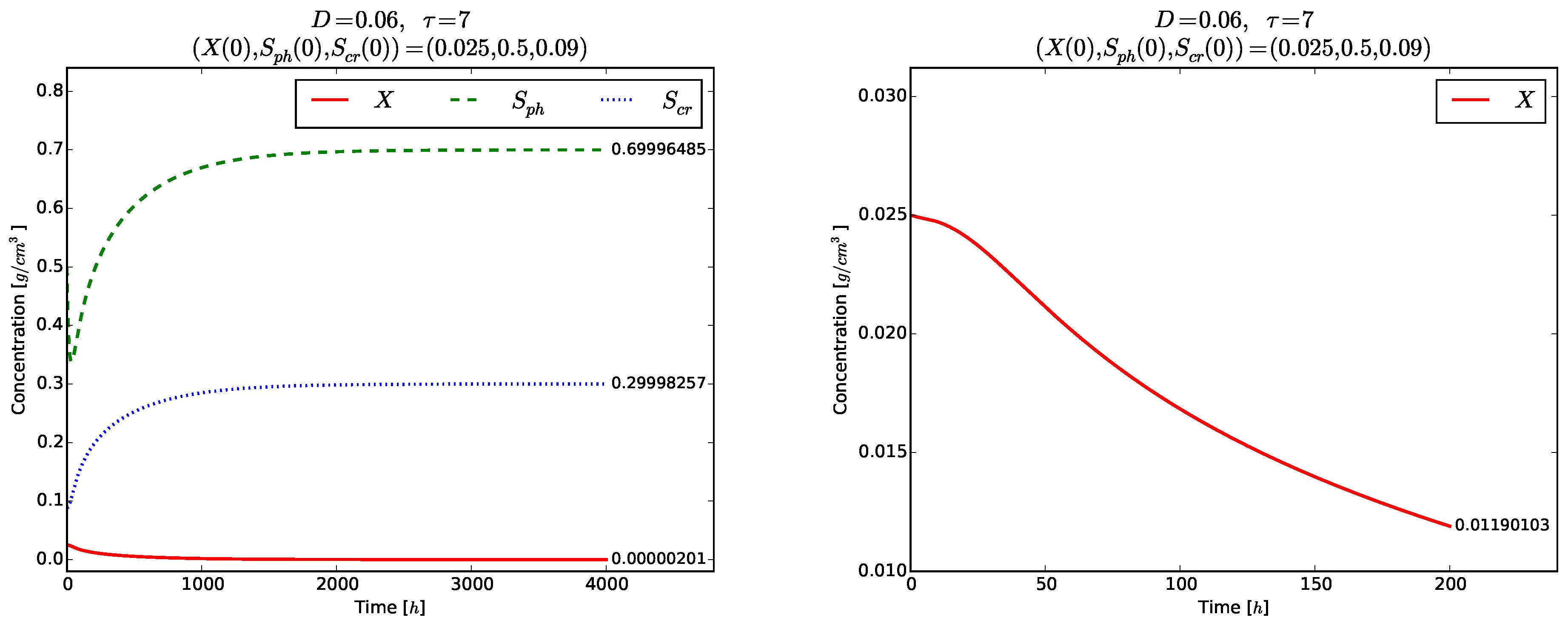
The next two examples demonstrate the global stability of the equilibrium points and with respect to D and .
For this value of
D we obtain
We choose and fix
. Then, there exist the two interior equilibria
Proposition 2 implies that
is locally asymptotically unstable, and
is the global attractor for the model (
3)–(5) according to Theorem 5.
According to Theorem 4(ii) the inequality
should be fulfilled to ensure existence of positive model solutions. The right plot in
Figure 6 visualizes the time evolution of
towards the globally asymptotically stable equilibrium
. As expected, no oscillations are observed in the time evolution of each one of the variables
(right plot in
Figure 6), as well as of
and
even for the short time
,
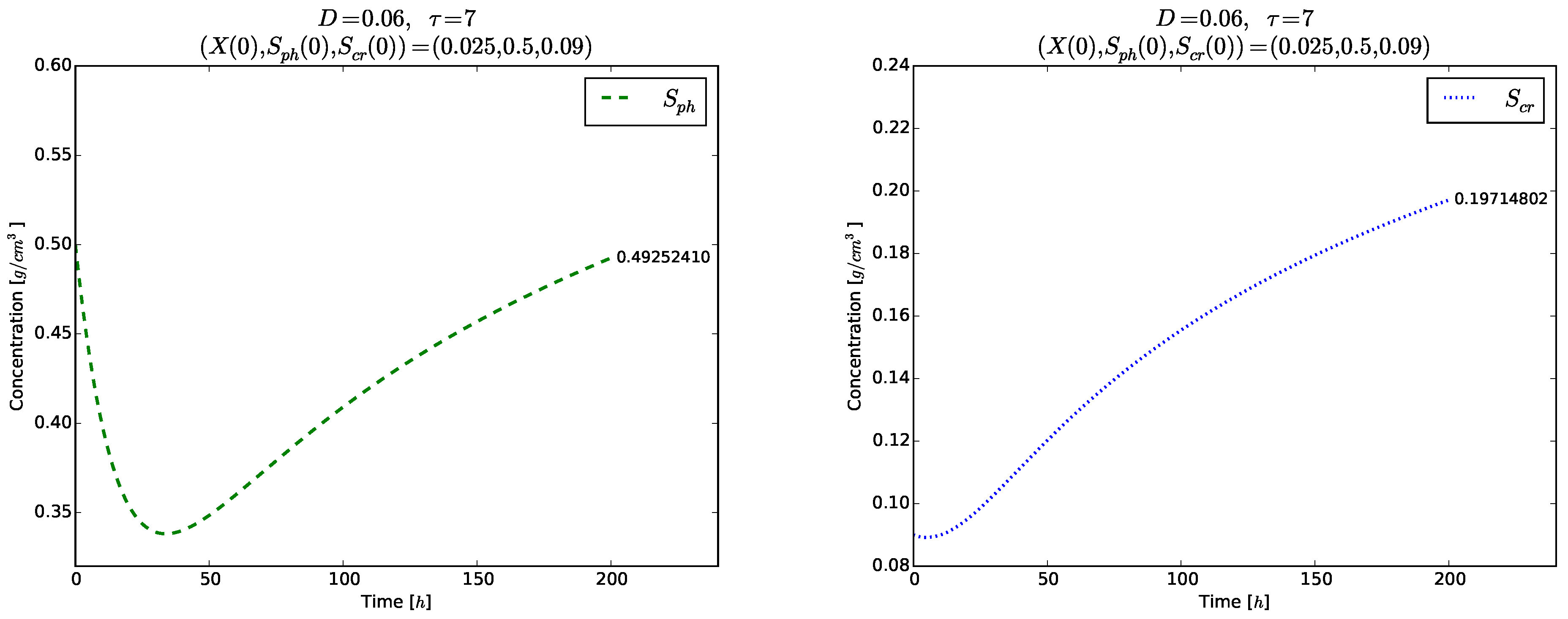
Figure 7.
Taking
we obtain
In this case,
is the global attractor of the model (Theorem 6), and
is locally asymptotically unstable (Proposition 2).
According to Theorem 4
, the inequality
has to be satisfied so that the model (
3)–(5) possesses positive solutions. The left plot in
Figure 8 illustrates the global stability of the washout equilibrium
. The right plot of
Figure 8 and
Figure 9 show the evolution of each phase variable
,
,
for a shorter time
. Again, no transient oscillations can be seen in these plots.
5. Conclusions and Future Work
We considered a time-delayed model, describing a phenol and 4-methylphenol (
p-cresol) mixture biodegradation in a continuously stirred tank bioreactor with a SKIP specific growth rate. It was based on a previously proposed and studied dynamical model, presented in [
25]. The discrete time delay
was introduced in the microorganisms’ growth response to indicate the delay in the conversion of the consumed nutrient into viable biomass.
We presented the mathematical analysis of the proposed time-delayed model. First, in
Section 2, the equilibrium points were determined, and their local asymptotic stability was investigated in dependence on the delay
(and on the dilution rate
D). Three equilibrium points were found, namely a boundary (washout) equilibrium
with
and two interior (coexistence) equilibria
and
with
. For any fixed
, values of
were determined,
, and it was shown that
exists and is locally asymptotically stable for (Proposition 1);
exists and is locally asymptotically unstable for (Proposition 2);
exists for all and is locally asymptotically stable (unstable) if () (Proposition 3).
Then, we showed that the locally unstable equilibrium
underwent local Hopf bifurcations at certain critical values of the delay parameter
(Theorems 1–3). To prove this, we exploited a known approach presented in [
34]. The occurrence of some transient oscillations as a result of the Hopf bifurcations was demonstrated by Example 1 in
Section 4, see
Figure 4 and
Figure 5. In this example, the delay
was chosen to be at one of the four existing bifurcating values, namely
h, which is approximately 418 days. According to Theorem 3, the period of the bifurcating solutions lies in the interval
h, which corresponds to an interval of approximately
days, with a width of 318 days. Practically, this oscillating behavior is difficult to observe, not only because the periodic solutions are unstable but also due to the rather large bifurcating periods, especially in realtime laboratory experiments.
Finally, in
Section 3, we established the existence and uniqueness of positive model solutions in Theorem 4. We reduced the 3-dimensional model (
3)–(5) into a limiting 2-dimensional dynamical system (
41). Although the reduced model was very similar to well known bioreactor models, the main difference was in the specific properties of the SKIP function
and, in particular, of
. We proved in Theorem 5 the global asymptotic stability of the coexistence equilibrium point
with respect to
whenever it exists, i.e., for
. However, if
, then the washout equilibrium
is globally asymptotically stable (Theorem 6). These results mean practically either long-term sustainability of the biodegradation process when
is the global attractor or process breakdown due to total washout of the biomass in the reactor when
is the global attractor. Numerical examples in
Section 4 (Examples 2 and 3) confirmed the latter theoretical results.
The time delayed model (
3)–(5), investigated in this paper, shows many similarities in its dynamic properties in comparison with the previously studied [
25] model (
1) without a time delay. These are, for example, the global attractivity of the two equilibrium points
and
. The main difference is the existence of Hopf bifurcations around the unstable equilibria at certain critical values of
, which serve as sources of transient oscillations in practical experiments.
As mentioned before, the model parameters in
Table 1 were obtained from laboratory experiments for phenol and
p-cresol mixture degradation. New experimental work is planned in the future to eventually account for the time delay in the biomass growth response and the model will be validated by the new data.
The proposed model (
3)–(5) could be successfully used on a bioreactor scale-up, and this is also planned in the future. It is very likely to change the values of the model parameters, which will lead to adaptation of the investigations and to new conclusions. However, the following theoretical results are independent of the particular parameter values: (i) existence of a washout steady state
and of at least one interior (coexistence) equilibrium
; and (ii) the inversely proportional relationship between the dilution rate
D and the time delay
. Generally speaking, this means that relatively large values of
D and small values of
may cause biomass washout, due to the global stability of
, and thus to process breakdown in the bioreactor. On the contrary, smaller values of
D and larger values of
lead to a stable process and biodegradation sustainability due to the global attractivity of
. Since
D is the controllable input in the bioreactor, these results can be very useful for the experimenter in order to obtain answers long before the physical prototype of the actual system is built and tested.
A next step in future investigations will be to treat more complex chemical compounds involving more than two substrates. The challenging element will be the design of the biomass specific growth rate as a SKIP-type model. This could be performed in dependence on experimental results showing the activity and the mutual interaction of the involved substances.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}