
Effects of Sulfamethoxazole on Growth and Antibiotic Resistance of A Natural Microbial Community


 ,
, 


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Anaerobic Digestate Samples
2.3. “Concentration-Dependent” Experiments
2.4. DNA Extraction and qPCR Analysis
2.5. Analytical Determination of SMX
2.6. Statistical Analysis
3. Results and Discussions
3.1. SMX, intI1 Gene, and SMX-ARGs in the Digestate
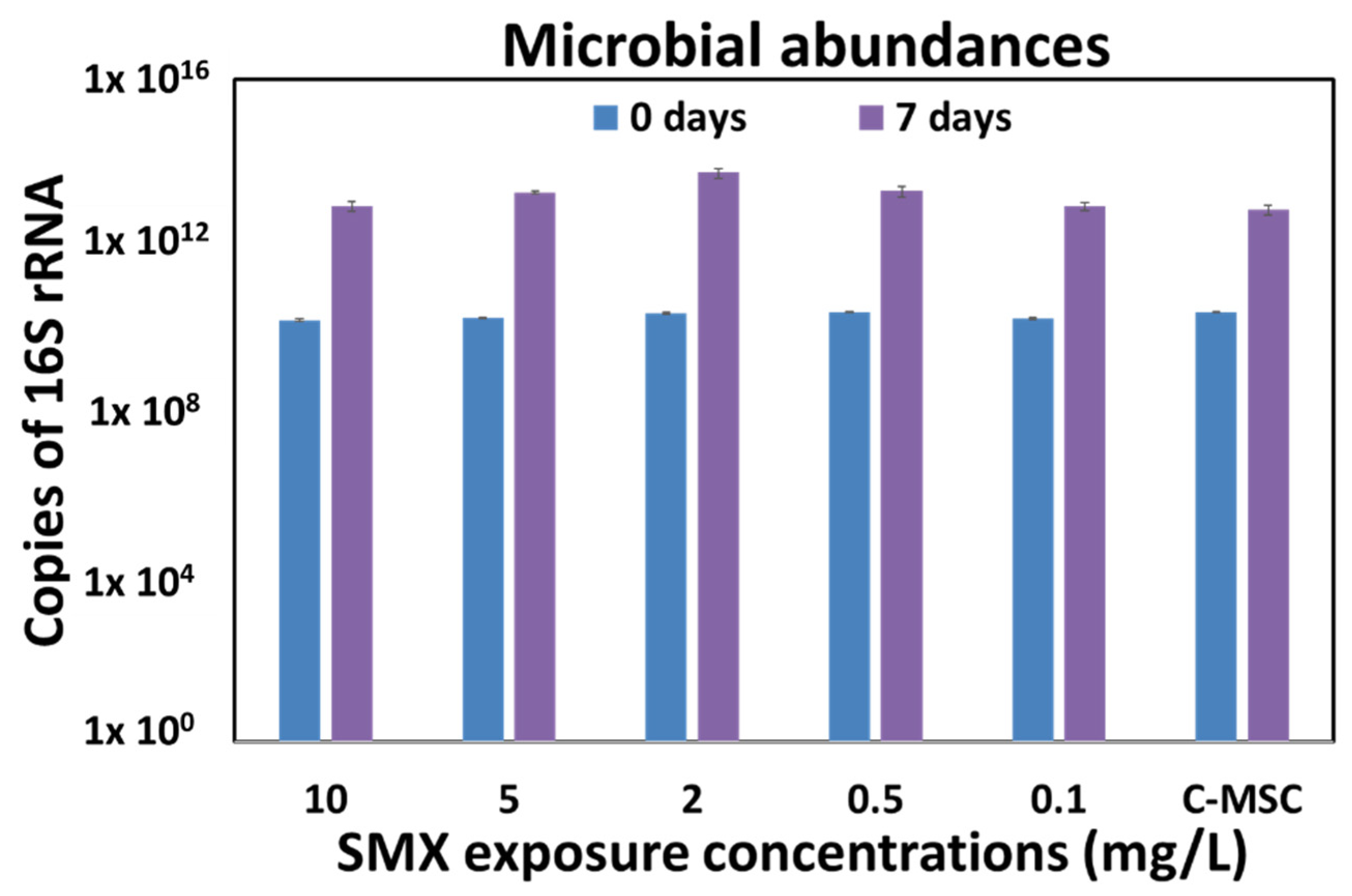
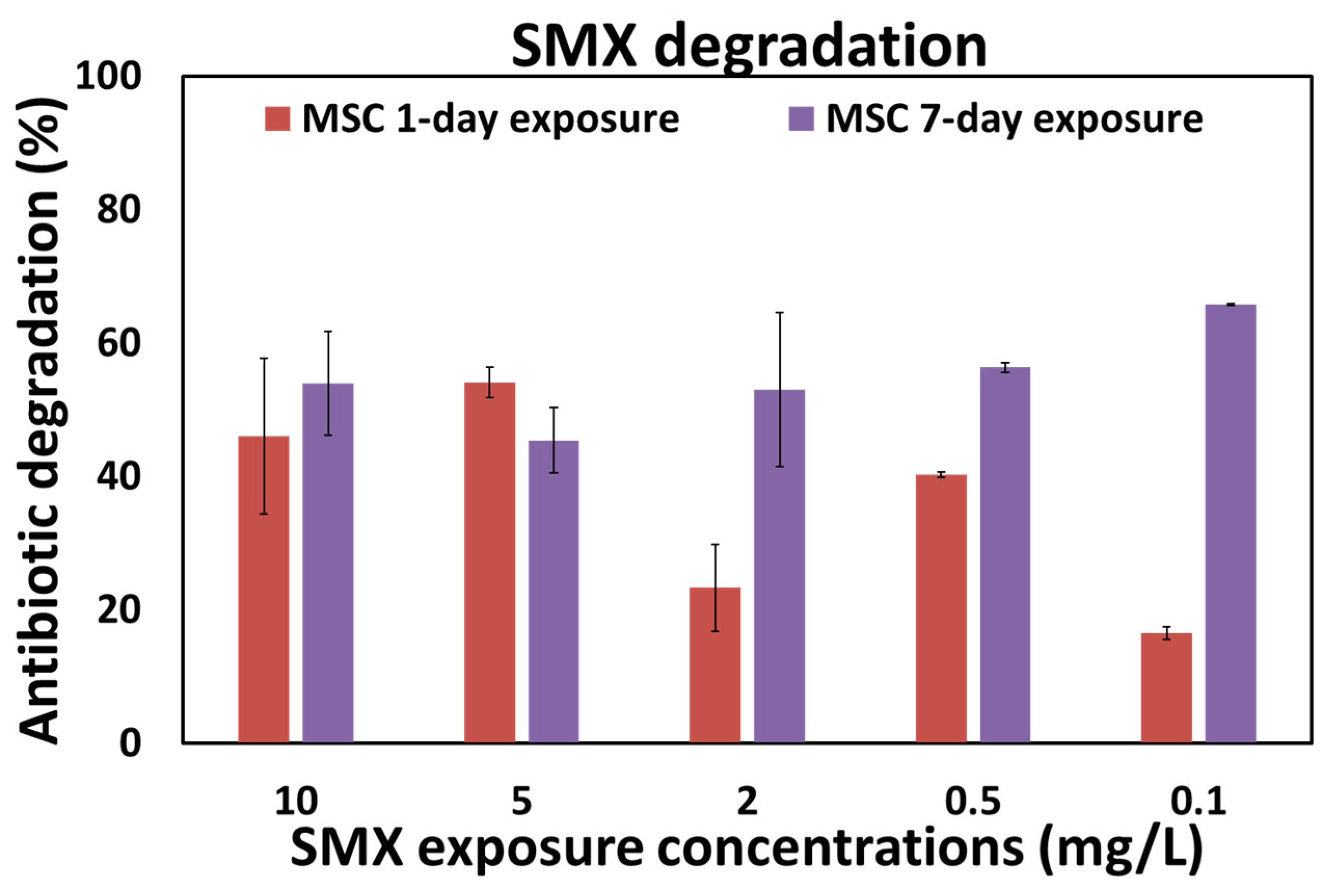
3.2. “Concentration-Dependent” Experiments
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hawkey, P.M. The origins and molecular basis of antibiotic resistance. Br. Med. J. 1998, 317, 657–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berti, A.D.; Hirsch, E.B. Tolerance to antibiotics affects response. Science 2020, 367, 2019–2021. [Google Scholar] [CrossRef] [PubMed]
- García, J.; García-Galán, M.J.; Day, J.W.; Boopathy, R.; White, J.R.; Wallace, S.; Hunter, R.G. A review of emerging organic contaminants (EOCs), antibiotic resistant bacteria (ARB), and antibiotic resistance genes (ARGs) in the environment: Increasing removal with wetlands and reducing environmental impacts. Bioresour. Technol. 2020, 307, 123228. [Google Scholar] [CrossRef] [PubMed]
- Nadimpalli, M.L.; Marks, S.J.; Montealegre, M.C.; Gilman, R.H.; Pajuelo, M.J.; Saito, M.; Tsukayama, P.; Njenga, S.M.; Kiiru, J.; Swarthout, J.; et al. Urban informal settlements as hotspots of antimicrobial resistance and the need to curb environmental transmission. Nat. Microbiol. 2020, 5, 787–795. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Environmental factors influencing the development and spread of antibiotic resistance. FEMS Microbiol. Rev. 2018, 42. [Google Scholar] [CrossRef]
- Lee, K.; Kim, D.W.; Lee, D.H.; Kim, Y.S.; Bu, J.H.; Cha, J.H.; Thawng, C.N.; Hwang, E.M.; Seong, H.J.; Sul, W.J.; et al. Mobile resistome of human gut and pathogen drives anthropogenic bloom of antibiotic resistance. Microbiome 2020, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Collignon, P. Antibiotic resistance: Are we all doomed? Intern. Med. J. 2015, 45, 1109–1115. [Google Scholar] [CrossRef]
- Finley, R.L.; Collignon, P.; Larsson, D.G.J.; Mcewen, S.A.; Li, X.Z.; Gaze, W.H.; Reid-Smith, R.; Timinouni, M.; Graham, D.W.; Topp, E. The scourge of antibiotic resistance: The important role of the environment. Clin. Infect. Dis. 2013, 57, 704–710. [Google Scholar] [CrossRef] [Green Version]
- Grenni, P.; Ancona, V.; Barra Caracciolo, A. Ecological effects of antibiotics on natural ecosystems: A review. Microchem. J. 2018, 36, 25–39. [Google Scholar] [CrossRef]
- Le Page, G.; Gunnarsson, L.; Snape, J.; Tyler, C.R. Integrating human and environmental health in antibiotic risk assessment: A critical analysis of protection goals, species sensitivity and antimicrobial resistance. Environ. Int. 2017, 109, 155–169. [Google Scholar] [CrossRef]
- Chow, L.K.M.; Ghaly, T.M.; Gillings, M.R. A survey of sub-inhibitory concentrations of antibiotics in the environment. J. Environ. Sci. 2021, 99, 21–27. [Google Scholar] [CrossRef]
- Gullberg, E.; Albrecht, L.M.; Karlsson, C.; Sandegren, L.; Andersson, D.I. Selection of a multidrug resistance plasmid by sublethal levels of antibiotics and heavy metals. MBio 2014, 5, e01918-14. [Google Scholar] [CrossRef] [Green Version]
- Gullberg, E.; Cao, S.; Berg, O.G.; Ilbäck, C.; Sandegren, L.; Hughes, D.; Andersson, D.I. Selection of resistant bacteria at very low antibiotic concentrations. PLoS Pathog. 2011, 7, e1002158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarei-Baygi, A.; Harb, M.; Wang, P.; Stadler, L.B.; Smith, A.L. Microbial community and antibiotic resistance profiles of biomass and effluent are distinctly affected by antibiotic addition to an anaerobic membrane bioreactor. Environ. Sci. Water Res. Technol. 2020, 6, 724–736. [Google Scholar] [CrossRef]
- Yan, M.; Xu, C.; Huang, Y.; Nie, H.; Wang, J. Tetracyclines, sulfonamides and quinolones and their corresponding resistance genes in the Three Gorges Reservoir, China. Sci. Total Environ. 2018, 631–632, 840–848. [Google Scholar] [CrossRef]
- Ma, J.; Cui, Y.; Li, A.; Zhang, W.; Liang, J.; Wang, S.; Zhang, L. Evaluation of the fate of nutrients, antibiotics, and antibiotic resistance genes in sludge treatment wetlands. Sci. Total Environ. 2020, 712, 136370. [Google Scholar] [CrossRef]
- Gillings, M.R.; Gaze, W.H.; Pruden, A.; Smalla, K.; Tiedje, J.M.; Zhu, Y.G. Using the class 1 integron-integrase gene as a proxy for anthropogenic pollution. ISME J. 2015, 9, 1269–1279. [Google Scholar] [CrossRef]
- Hernando-Amado, S.; Sanz-García, F.; Blanco, P.; Martínez, J.L. Fitness costs associated with the acquisition of antibiotic resistance. Essays Biochem. 2017, 61, 37–48. [Google Scholar] [PubMed]
- Klümper, U.; Recker, M.; Zhang, L.; Yin, X.; Zhang, T.; Buckling, A.; Gaze, W.H. Selection for antimicrobial resistance is reduced when embedded in a natural microbial community. ISME J. 2019, 13, 2927–2937. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Yang, Q.; Wang, R.; Wang, R.; Wang, Q.; Xin, Y. Evolution of antibiotic resistance and the relationship between the antibiotic resistance genes and microbial compositions under long-term exposure to tetracycline and sulfamethoxazole. Int. J. Environ. Res. Public Health 2019, 16, 4681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, A.K.; Zhang, L.; Snape, J.; Gaze, W.H. Comparing the selective and co-selective effects of different antimicrobials in bacterial communities. Int. J. Antimicrob. Agents 2019, 53, 1–7. [Google Scholar] [CrossRef]
- Di Lenola, M.; Barra Caracciolo, A.; Grenni, P.; Ancona, V.; Rauseo, J.; Laudicina, V.A.; Uricchio, V.F.; Massacci, A. Effects of Apirolio Addition and Alfalfa and Compost Treatments on the Natural Microbial Community of a Historically PCB-Contaminated Soil. Water Air Soil Pollut. 2018, 229, 143. [Google Scholar] [CrossRef]
- Barra Caracciolo, A.; Bottoni, P.; Crobe, A.; Fava, L.; Funari, E.; Giuliano, G.; Silvestri, C. Microbial degradation of two carbamate insecticides and their main metabolites in soil. Chem. Ecol. 2002, 18, 245–255. [Google Scholar] [CrossRef]
- Murray, A.K.; Zhang, L.; Yin, X.; Zhang, T.; Buckling, A.; Snape, J.; Gaze, W.H. Novel insights into selection for antibiotic resistance in complex microbial communities. MBio 2018, 9, e00969-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EUCAST (The European Committee on Antimicrobial Susceptibility Testing). Breakpoint Tables for Interpretation of MICs and Zone Diameters; Version 10.0; EUCAST: Växjö, Sweden, 2020. [Google Scholar]
- Göbel, A.; McArdell, C.S.; Suter, M.J.F.; Giger, W. Trace determination of macrolide and sulfonamide antimicrobials, a human sulfonamide metabolite, and trimethoprim in wastewater using liquid chromatography coupled to electrospray tandem mass spectrometry. Anal. Chem. 2004, 76, 4756–4764. [Google Scholar] [CrossRef] [PubMed]
- Göbel, A.; Thomsen, A.; McArdell, C.S.; Alder, A.C.; Giger, W.; Theiß, N.; Löffler, D.; Ternes, T.A. Extraction and determination of sulfonamides, macrolides, and trimethoprim in sewage sludge. J. Chromatogr. A 2005, 1085, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Spataro, F.; Ademollo, N.; Pescatore, T.; Rauseo, J.; Patrolecco, L. Antibiotic residues and endocrine disrupting compounds in municipal wastewater treatment plants in Rome, Italy. Microchem. J. 2019, 148, 634–642. [Google Scholar] [CrossRef]
- Thompson, M.; Ellison, S.L.R.; Wood, R. Harmonized guidelines for single-laboratory validation of methods of analysis (IUPAC Technical Report). Pure Appl. Chem. 2002, 74, 835–855. [Google Scholar] [CrossRef]
- Gurmessa, B.; Pedretti, E.F.; Cocco, S.; Cardelli, V.; Corti, G. Manure anaerobic digestion effects and the role of pre- and post-treatments on veterinary antibiotics and antibiotic resistance genes removal efficiency. Sci. Total Environ. 2020, 721, 137532. [Google Scholar] [CrossRef]
- Mazzurco Miritana, V.; Massini, G.; Visca, A.; Grenni, P.; Patrolecco, L.; Spataro, F.; Rauseo, J.; Garbini, G.L.; Signorini, A.; Rosa, S.; et al. Effects of Sulfamethoxazole on the Microbial Community Dynamics During the Anaerobic Digestion Process. Front. Microbiol. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Mohring, S.A.I.; Strzysch, I.; Fernandes, M.R.; Kiffmeyer, T.K.; Tuerk, J.; Hamscher, G. Degradation and elimination of various sulfonamides during anaerobic fermentation: A promising step on the way to sustainable pharmacy? Environ. Sci. Technol. 2009, 43, 2569–2574. [Google Scholar] [CrossRef]
- Gillings, M.; Boucher, Y.; Labbate, M.; Holmes, A.; Krishnan, S.; Holley, M.; Stokes, H.W. The evolution of class 1 integrons and the rise of antibiotic resistance. J. Bacteriol. 2008, 190, 5095–5100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adelowo, O.O.; Helbig, T.; Knecht, C.; Reincke, F.; Mäusezahl, I.; Müller, J.A. High abundances of class 1 integrase and sulfonamide resistance genes, and characterisation of class 1 integron gene cassettes in four urban wetlands in Nigeria. PLoS ONE 2018, 13, e0208269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Mao, D.; Rysz, M.; Zhou, Q.; Zhang, H.; Xu, L.; Alvarez, P.J.J. Trends in antibiotic resistance genes occurrence in the Haihe River, China. Environ. Sci. Technol. 2010, 44, 7220–7225. [Google Scholar] [CrossRef]
- Stanton, I.C.; Murray, A.K.; Zhang, L.; Snape, J.; Gaze, W.H. Evolution of antibiotic resistance at low antibiotic concentrations including selection below the minimal selective concentration. Commun. Biol. 2020, 3, 1–11. [Google Scholar] [CrossRef]
- Karci, A.; Balcioǧlu, I.A. Investigation of the tetracycline, sulfonamide, and fluoroquinolone antimicrobial compounds in animal manure and agricultural soils in Turkey. Sci. Total Environ. 2009, 407, 4652–4664. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhou, Q.; Luo, Y. Occurrence and source analysis of typical veterinary antibiotics in manure, soil, vegetables and groundwater from organic vegetable bases, northern China. Environ. Pollut. 2010, 158, 2992–2998. [Google Scholar] [CrossRef]
- Visca, A.; Barra Caracciolo, A.; Grenni, P.; Patrolecco, L.; Rauseo, J.; Massini, G.; Miritana, V.M.; Spataro, F. Anaerobic Digestion and Removal of Sulfamethoxazole, Enrofloxacin, Ciprofloxacin and Their Antibiotic Resistance Genes in a Full-Scale Biogas Plant. Antibiotic 2021, 10, 502. [Google Scholar] [CrossRef]
- Radke, M.; Lauwigi, C.; Heinkele, G.; Múrdter, T.E.; Letzel, M. Fate of the antibiotic sulfamethoxazole and its two major human metabolites in a water sediment test. Environ. Sci. Technol. 2009, 43, 3135–3141. [Google Scholar] [CrossRef]
- Larcher, S.; Yargeau, V. Biodegradation of sulfamethoxazole by individual and mixed bacteria. Appl. Microbiol. Biotechnol. 2011, 91, 211–218. [Google Scholar] [CrossRef]
- Larcher, S.; Yargeau, V. Biodegradation of sulfamethoxazole: Current knowledge and perspectives. Appl. Microbiol. Biotechnol. 2012, 96, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Cycoń, M.; Mrozik, A.; Piotrowska-Seget, Z. Antibiotics in the soil environment—Degradation and their impact on microbial activity and diversity. Front. Microbiol. 2019, 10, 338. [Google Scholar] [CrossRef]
- Topp, E.; Chapman, R.; Devers-Lamrani, M.; Hartmann, A.; Marti, R.; Martin-Laurent, F.; Sabourin, L.; Scott, A.; Sumarah, M. Accelerated Biodegradation of Veterinary Antibiotics in Agricultural Soil following Long-Term Exposure, and Isolation of a Sulfamethazine-degrading Microbacterium sp. J. Environ. Qual. 2013, 42, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Rauseo, J.; Barra Caracciolo, A.; Ademollo, N.; Cardoni, M.; Di Lenola, M.; Gaze, W.H.; Stanton, I.C.; Grenni, P.; Pescatore, T.; Spataro, F.; et al. Dissipation of the antibiotic sulfamethoxazole in a soil amended with anaerobically digested cattle manure. J. Hazard. Mater. 2019, 378, 120769. [Google Scholar] [CrossRef]
- Norman, A.; Hansen, L.H.; Sørensen, S.J. Conjugative plasmids: Vessels of the communal gene pool. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2275–2289. [Google Scholar] [CrossRef] [Green Version]
- Grenni, P.; Patrolecco, L.; Rauseo, J.; Spataro, F.; Di Lenola, M.; Aimola, G.; Zacchini, M.; Pietrini, F.; Di Baccio, D.; Stanton, I.C.; et al. Sulfamethoxazole persistence in a river water ecosystem and its effects on the natural microbial community and Lemna minor plant. Microchem. J. 2019, 149, 103999. [Google Scholar] [CrossRef]
- Patrolecco, L.; Rauseo, J.; Ademollo, N.; Grenni, P.; Cardoni, M.; Levantesi, C.; Luprano, M.L.; Barra Caracciolo, A. Persistence of the antibiotic sulfamethoxazole in river water alone or in the co-presence of ciprofloxacin. Sci. Total Environ. 2018, 640–641, 1438–1446. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| SMX | intI1 Gene | sul1 Gene | sul2 Gene | |
|---|---|---|---|---|
| Average values | 2.40 | 3.03 ×10−4 | 2.71 ×10−4 | 8.06 ×10−2 |
| s.e. | 0.50 | 1.20 ×10−3 | 2.02 ×10−3 | 0.03 |
| sul1 Gene | sul2 Gene | intI1 Gene | ||||
|---|---|---|---|---|---|---|
| 0 Days | 7 Days | 0 Days | 7 Days | 0 Days | 7 Days | |
| C-MSC | 2.6 × 10−4 | 8.3 × 10−8 | 8.1 × 10−2 | 2.0 × 10−4 | 3.2 × 10−4 | 2.8 × 10−6 |
| 10 mg/L | 2.7 × 10−4 | 3.9 × 10−8 | 8.1 × 10−2 | 1.9 × 10−4 | 2.32 × 10−8 | 3.4 × 10−7 |
| 5 mg/L | 2.7 × 10−4 | 0 | 8.0 × 10−2 | 0 | 2.32 × 10−8 | 2.3 × 10−6 |
| 2 mg/L | 2.8 × 10−4 | 3.1 × 10−10 | 8.3 × 10−2 | 3.1 × 10−7 | 2.32 × 10−8 | 1.5 × 10−4 |
| 0.5 mg/L | 2.5 × 10−4 | 7.3 × 10−10 | 8.1 × 10−2 | 4.7 × 10−5 | 2.32 × 10−8 | 1.2 × 10−5 |
| 0.1 mg/L | 2.3 × 10−4 | 1.1 × 10−6 | 8.0 × 10−2 | 3.9 × 10−4 | 2.32 × 10−8 | 7.8 × 10−7 |
| Factors | p-Value |
|---|---|
| (A) SMX concentrations | 0.00376 ** |
| (B) time | 0.01253 * |
| (C) specific gene | 1.25 × 10−7 *** |
| A × B Interaction | 0.01084 * |
| A × C Interaction | 3.72 × 10−6 *** |
| B × C Interaction | 0.10162 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rauseo, J.; Barra Caracciolo, A.; Spataro, F.; Visca, A.; Ademollo, N.; Pescatore, T.; Grenni, P.; Patrolecco, L. Effects of Sulfamethoxazole on Growth and Antibiotic Resistance of A Natural Microbial Community. Water 2021, 13, 1262. https://doi.org/10.3390/w13091262
Rauseo J, Barra Caracciolo A, Spataro F, Visca A, Ademollo N, Pescatore T, Grenni P, Patrolecco L. Effects of Sulfamethoxazole on Growth and Antibiotic Resistance of A Natural Microbial Community. Water. 2021; 13(9):1262. https://doi.org/10.3390/w13091262
Chicago/Turabian StyleRauseo, Jasmin, Anna Barra Caracciolo, Francesca Spataro, Andrea Visca, Nicoletta Ademollo, Tanita Pescatore, Paola Grenni, and Luisa Patrolecco. 2021. "Effects of Sulfamethoxazole on Growth and Antibiotic Resistance of A Natural Microbial Community" Water 13, no. 9: 1262. https://doi.org/10.3390/w13091262
APA StyleRauseo, J., Barra Caracciolo, A., Spataro, F., Visca, A., Ademollo, N., Pescatore, T., Grenni, P., & Patrolecco, L. (2021). Effects of Sulfamethoxazole on Growth and Antibiotic Resistance of A Natural Microbial Community. Water, 13(9), 1262. https://doi.org/10.3390/w13091262