
Optimization of Cultivation Conditions for Tetraselmis striata and Biomass Quality Evaluation for Fish Feed Production

 , ,
, ,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Culture Conditions
2.2. Tested Growth Substrates
2.3. Optimization of Growth Conditions
2.4. Analytical Methods and Calculations
2.4.1. Medium Analyses
2.4.2. Biomass Analyses
Estimation of Dry Biomass and Biomass Yields
Lipid and Carbohydrate Estimation
Lipid Fractionation and Fatty Acid Analysis
Protein and Amino Acid Profile Estimation
Pigment Estimation
2.5. Statistical Analysis
3. Results and Discussion
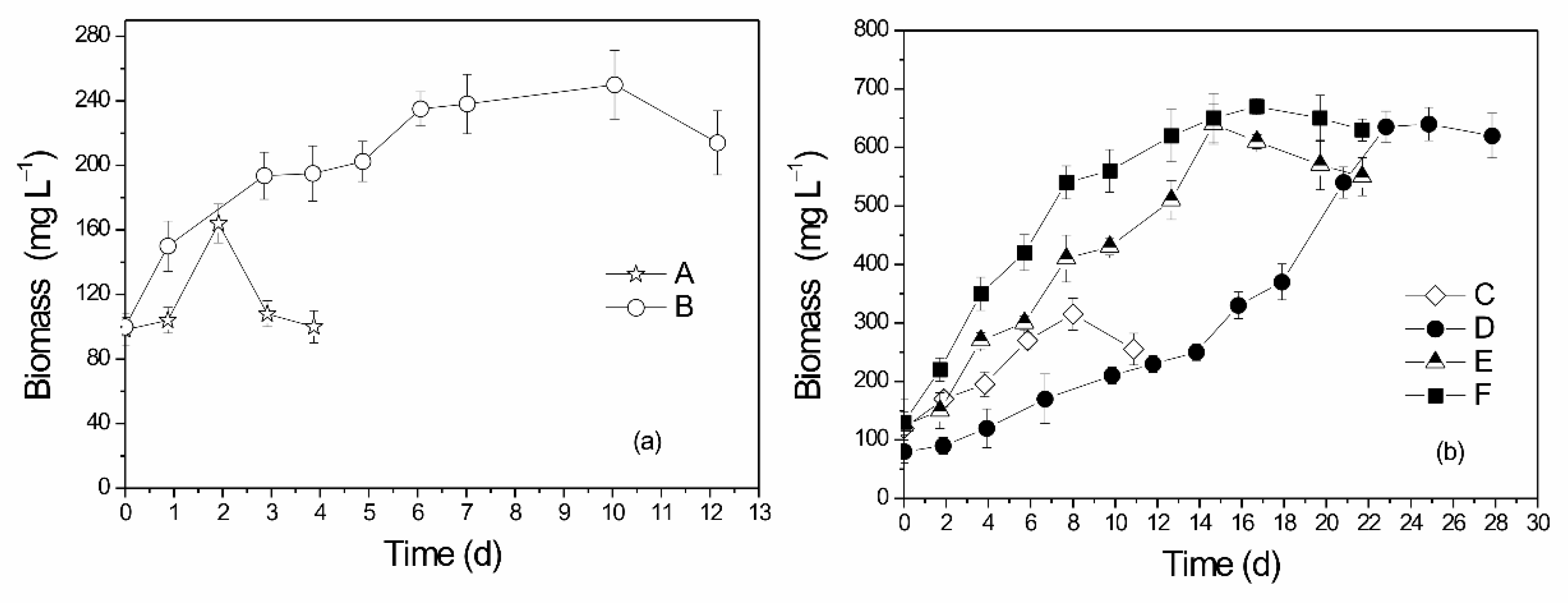
3.1. Substrate Optimization
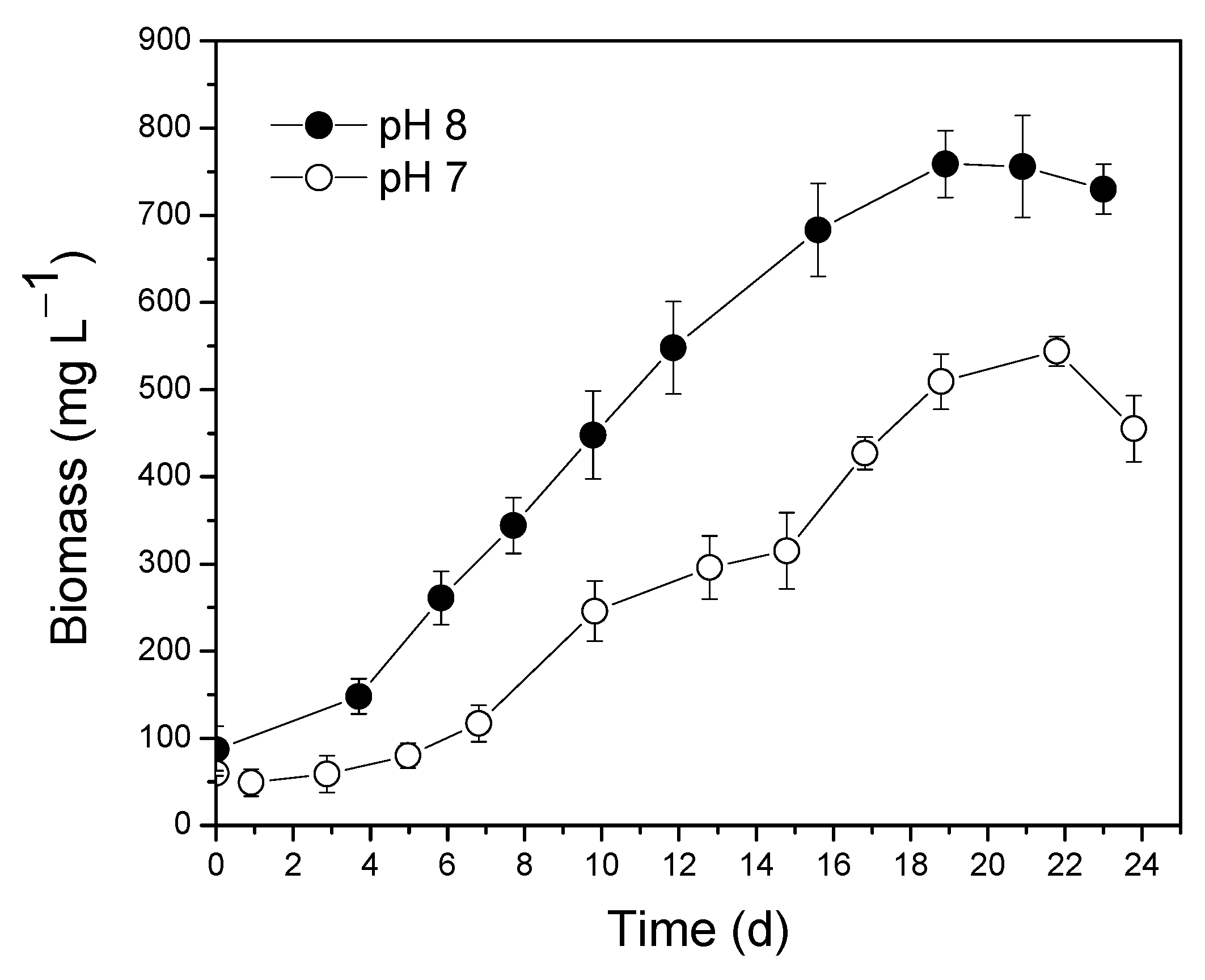
3.2. Optimization of Substrate pH
Effect of pH on Biomass Composition
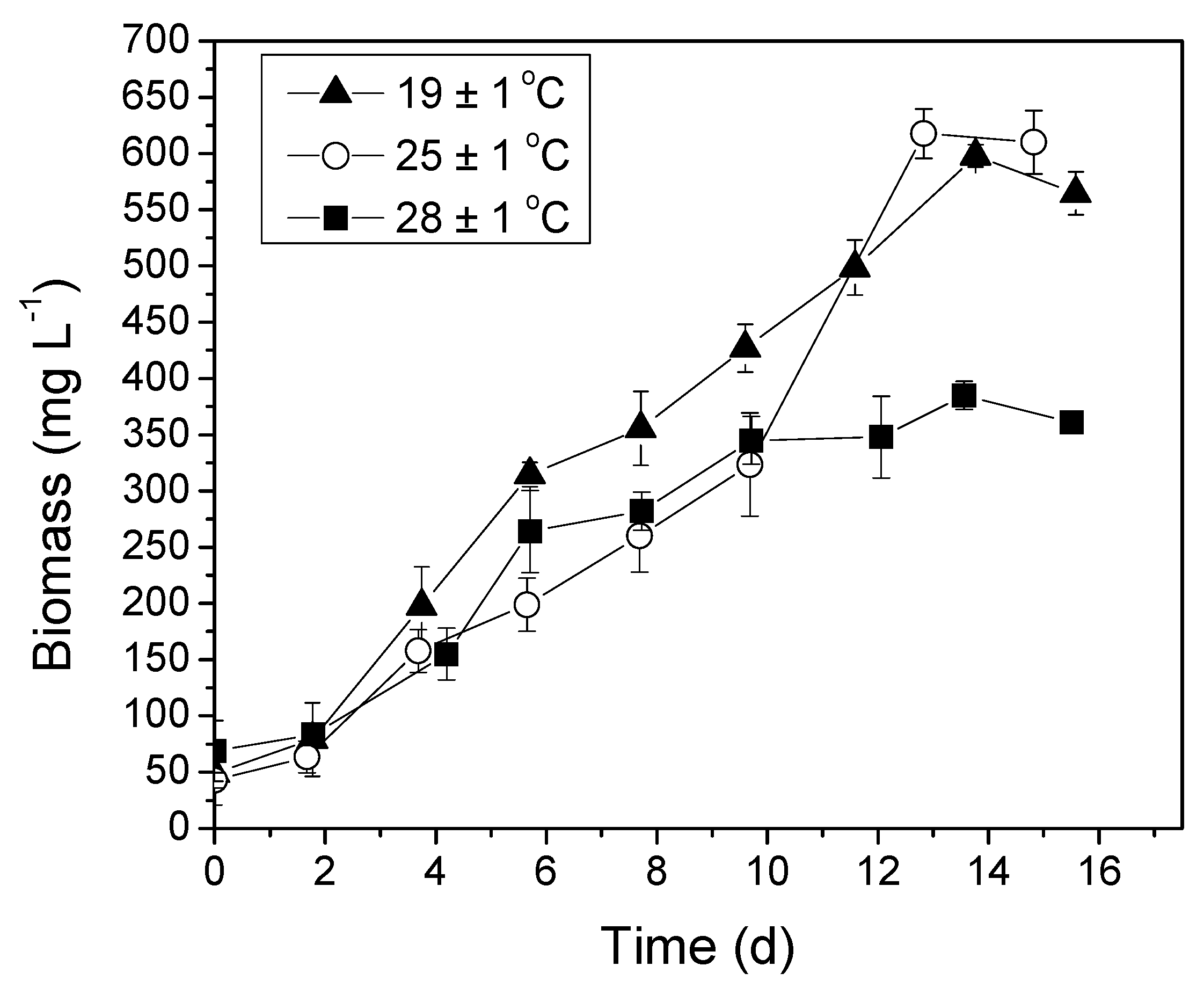
3.3. Temperature Optimization
Effect of Temperature on Biomass Composition
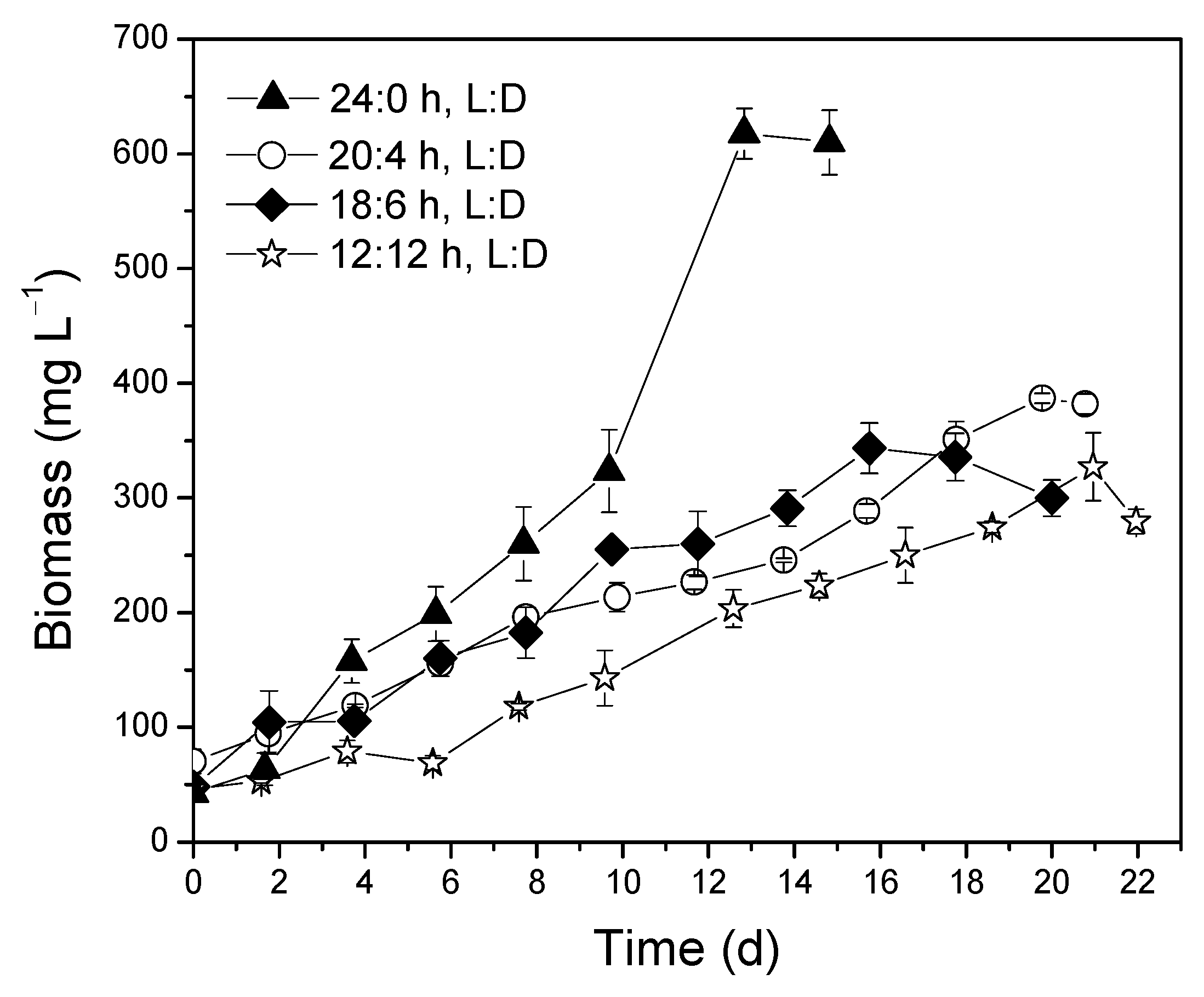
3.4. Optimization of Photoperiod
Effect of Photoperiod on Biomass Composition
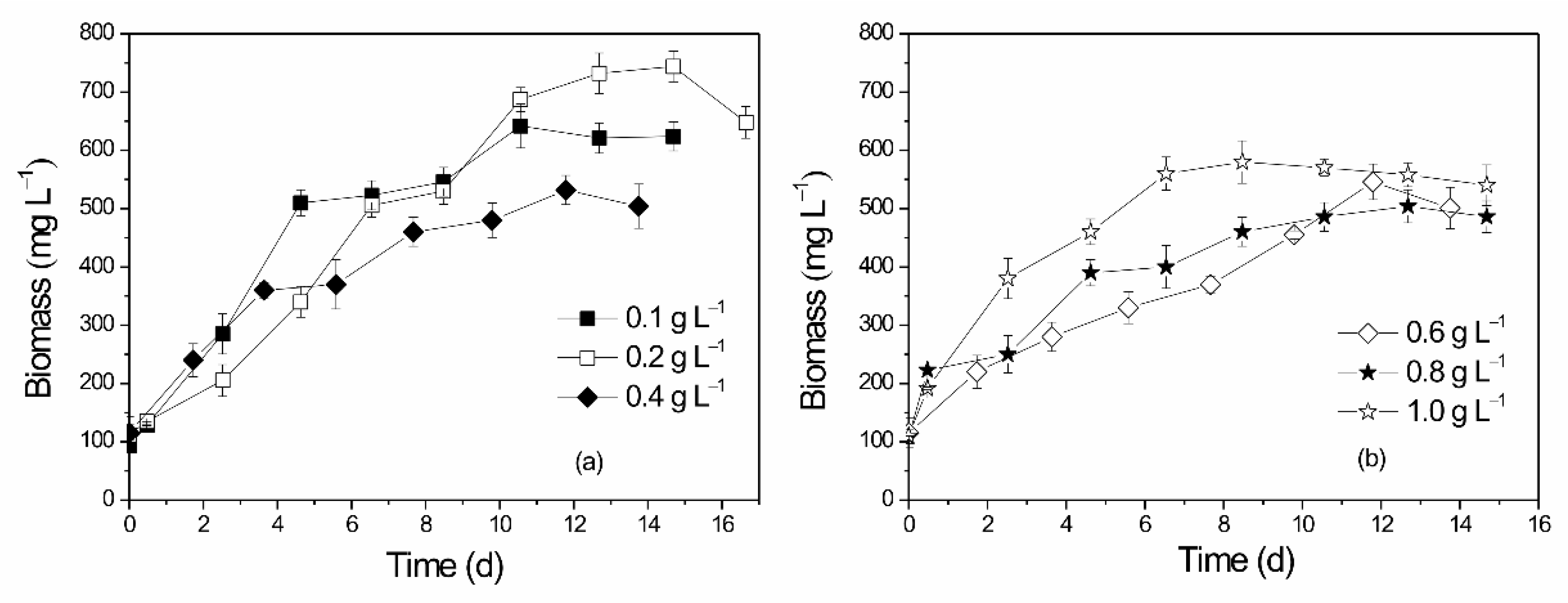
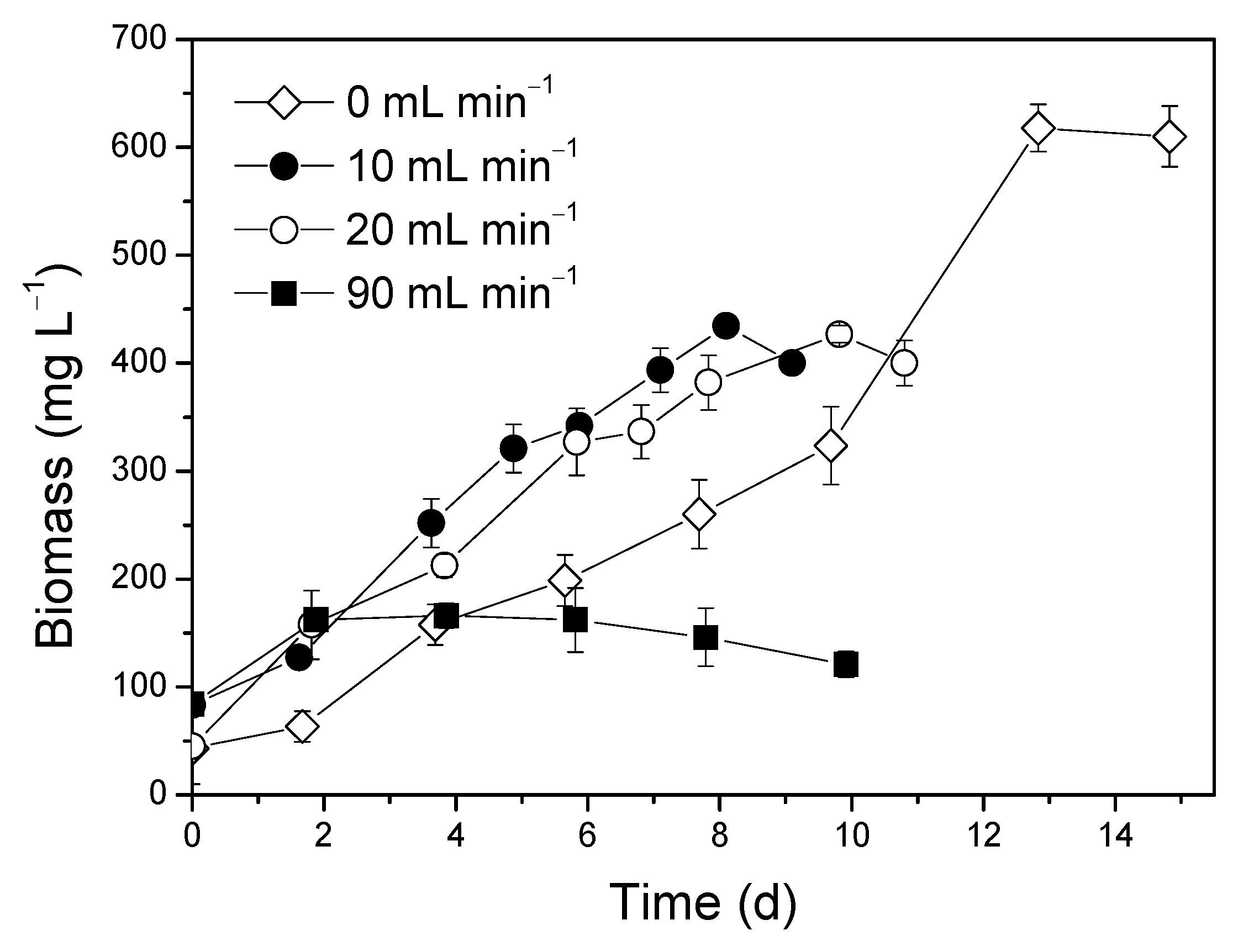
3.5. Optimization of CO2 Flow Rate
Effect of CO2 on Biomass Composition
3.6. Effect of Different Growth Conditions on Fatty Acid Composition
3.6.1. Effect of pH on Fatty Acid Composition
3.6.2. Effect of Temperature on Fatty Acid Composition
3.6.3. Effect of Photoperiod on Fatty Acid Composition
3.6.4. Effect of CO2 Supply on Fatty Acid Composition
3.7. Effect of Different Growth Conditions on the Amino Acid Profile of T. striata
3.8. Effect of Different Growth Conditions on Carotenoid Profile of T. striata
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, F.; Leng, Y.; Lu, Q.; Zhou, W. The application of microalgae biomass and bio-products as aquafeed for aquaculture. Algal Res. 2021, 60, 102541. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. The State of World Fisheries and Aquaculture. Sustainability in Action; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Shah, M.R.; Lutzu, G.A.; Alam, A.; Sarker, P.; Kabir Chowdhury, M.A.; Parsaeimehr, A.; Liang, Y.; Daroch, M. Microalgae in aquafeeds for a sustainable aquaculture industry. J. Appl. Phycol. 2018, 30, 197–213. [Google Scholar] [CrossRef]
- Imamoglu, E.; Demirel, Z.; Conk Dalay, M. Process optimization and modeling for the cultivation of Nannochloropsis sp. and Tetraselmis striata via response surface methodology. J. Phycol. 2015, 51, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Kaparapu, J. Application of Microalgae in Aquaculture. Phykos 2018, 48, 21–26. [Google Scholar]
- Ma, R.; Wang, B.; Chua, E.T.; Zhao, X.; Lu, K.; Ho, S.H.; Shi, X.; Liu, L.; Xie, Y.; Lu, Y.; et al. Comprehensive utilization of marine microalgae for enhanced co-production of multiple compounds. Mar. Drugs 2020, 18, 467. [Google Scholar] [CrossRef]
- Kim, G.; Lee, C.H.; Lee, K. Enhancement of lipid production in marine microalga Tetraselmis sp. through salinity variation. Korean J. Chem. Eng. 2015, 33, 230–237. [Google Scholar] [CrossRef]
- Kim, G.; Mujtaba, G.; Lee, K. Effects of nitrogen sources on cell growth and biochemical composition of marine chlorophyte Tetraselmis sp. for lipid production. Algae 2016, 31, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Molina, E.; Martínez, E.; Sańchez, S.; García, F.; Contreras, A. The influence of temperature and the initial N:P ratio on the growth of microalgae Tetraselmis sp. Process Biochem. 1991, 26, 183–187. [Google Scholar] [CrossRef]
- Danquah, M.K.; Harun, R.; Halim, R.; Forde, G.M. Cultivation medium design via elemental balancing for Tetraselmis suecica. Chem. Biochem. Eng. Q. 2010, 24, 361–369. [Google Scholar]
- Carneiro, M.; Pôjo, V.; Malcata, F.X.; Otero, A. Lipid accumulation in selected Tetraselmis strains. J. Appl. Phycol. 2019, 31, 2845–2853. [Google Scholar] [CrossRef]
- Dammak, M.; Hadrich, B.; Miladi, R.; Barkallah, M.; Hentati, F.; Hachicha, R.; Laroche, C.; Michaud, P.; Fendi, I.; Abdelkafi, S. Effects of nutritional conditions on growth and biochemical composition of Tetraselmis sp. Lipids Health Dis. 2017, 16, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Khatoon, H.; Abdu Rahman, N.; Banerjee, S.; Harun, N.; Suleiman, S.S.; Zakaria, N.H.; Lananan, F.; Abdul Hamid, S.H.; Endut, A. Effects of different salinities and pH on the growth and proximate composition of Nannochloropsis sp. and Tetraselmis sp. isolated from South China Sea cultured under control and natural condition. Int. Biodeterior. Biodegr. 2014, 95, 11–18. [Google Scholar] [CrossRef]
- Rukminasari, N.; Omar, S.B.A.; Lukman, M. Effects of Increasing Temperature and Nitrate Concentration on Cell Abundance, Growth Rate, Biomass and Free Fatty Acid of Tetraselmis sp. In Proceedings of the 2nd International Symposium on Marine Science and Fisheries, Makassar, Indonesia, 22 June 2019. [Google Scholar]
- Chaisutyakorn, P.; Praiboon, J.; Kaewsuralikhit, C. The effect of temperature on growth and lipid and fatty acid composition on marine microalgae used for biodiesel production. J. Appl. Phycol. 2018, 30, 37–45. [Google Scholar] [CrossRef]
- Sas, A.A.; Turki, A.J.; Affan, A.; Al-Taisan, W.A.; Das, S.K.; Su, S.N.; Cob, Z.C. The Influence of Temperature and Nutrient Concentrations on Growth Rate, Biomass, Chlorophyll-a, and Biochemical Compositions of Tetraselmis suecica (Chlorophyta). In Proceedings of the IOP Conference Series: Earth and Environmental Science, Selangor, Malaysia, 10 November 2021. [Google Scholar]
- Yusof, N.S.; Yeong, Y.S.; Zakeri, H.A.; Wahid, M.E.A.; Ghafar, S.N.A.; Yusuf, N. Photoperiod influenced the growth and antioxidative responses of Chlorella vulgaris, Isochrysis galbana, and Tetraselmis chuii. J. Appl. Pharm. Sci. 2021, 11, 125–134. [Google Scholar]
- Pedro, S.I.H. Evaluation of Photoperiod Effect on the Growth and Protein Content of Microalgae. Ph.D. Thesis, School of Tourism and Maritime Technology, Polytechnic Institute of Leiria, Peniche, Portugal, 2015. [Google Scholar]
- Meseck, S.L.; Alix, J.H.; Wikfors, G.H. Photoperiod and light intensity effects on growth and utilization of nutrients by the aquaculture feed microalga, Tetraselmis chui (PLY429). Aquaculture 2005, 246, 393–404. [Google Scholar] [CrossRef]
- Molina, E.; Martinez, M.E.; Sanchez, S.; Garcia, F.; Contreras, A. The influence of flow rate and the composition of the CO2/air gaseous mixture on discontinuous growth of Tetraselmis sp. Appl. Microbiol. Biotechnol. 1990, 34, 103–107. [Google Scholar] [CrossRef]
- Kassim, M.A.; Meng, T.K. Carbon dioxide (CO2) biofixation by microalgae and its potential for biorefinery and biofuel production. Sci.Total Environ. 2016, 584–585, 1121–1129. [Google Scholar] [CrossRef]
- Ferriols, V.M.E.N.; Saclauso, C.A.; Fortes, N.R.; Toledo, N.A.; Pahila, I.G. Effect of elevated carbon dioxide and phosphorus levels on nitrogen uptake, lipid content and growth of Tetraselmis sp. J. Fish. Aquat. Sci. 2013, 8, 659–672. [Google Scholar]
- Zheng, Y.; Chen, Z.; Lu, H.; Zhang, W. Optimization of carbon dioxide fixation and starch accumulation by Tetraselmis subcordiformis in a rectangular airlift photobioreactor. Afr. J. Biotechnol. 2011, 10, 1888–1901. [Google Scholar]
- Rico, M.; González, A.G.; Santana-Casiano, M.; González-Dávila, M.; Pérez-Almeida, N.; de Tangil, M.S. Production of Primary and Secondary Metabolites Using Algae. In Prospects and Challenges in Algal Biotechnology; Tripathi, B.N., Kumar, D., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2017; pp. 311–326. [Google Scholar]
- Boopathy, A.B.; Jayakumar, T.; Chinnasamy, S.; Rajaram, M.G.; Mohan, N.; Nagaraj, S.; Rengasamy, R.; Manubolu, M.; Sheu, J.R.; Chang, C.C. Biomass and Lipid Production Potential of an Indian Marine Algal Isolate Tetraselmis striata BBRR1. Energies 2020, 13, 341. [Google Scholar] [CrossRef] [Green Version]
- Patidar, S.K.; Kim, S.H.; Kim, J.H.; Park, J.; Park, B.S.; Han, M.S. Pelagibaca bermudensis promotes biofuel competence of Tetraselmis striata in a broad range of abiotic stressors: Dynamics of quorum-sensing precursors and strategic improvement in lipid productivity. Biotechnol. Biofuels. 2018, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Zevallos-Feria, S. Influence of photoperiod on Tetraselmis striata (Chlorodendraceae) in benthic cultivation system. The Biologist (Lima) 2017, 15, 387–395. [Google Scholar]
- Geider, R.; La Roche, J. Redfield revisited: Variability of C:N:P in marine microalgae and its biochemical basis. Eur. J. Phycol. 2002, 57, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Pai, S.C.; Tsau, Y.J.; Yang, T.I. pH and buffering capacity problems involved in the determination of ammonia in saline water using the indophenol blue spectrophotometric method. Anal. Chim. Acta. 2001, 434, 209–216. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Tsonis, S.P. A modified method for the examination of chemical oxygen demand in sea water. WIT Trans. Ecol. Environ. 1993, 2, 1743–3541. [Google Scholar]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Folch, J.; Lees, M.; Sloane-Stanley, G.A. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1951, 226, 497–509. [Google Scholar] [CrossRef]
- Bellou, S.; Aggelis, G. Biochemical activities in Chlorella sp. and Nannochloropsis salina during lipid and sugar synthesis in a lab-scale open pond simulating reactor. J. Biotechnol. 2012, 164, 318–329. [Google Scholar] [CrossRef]
- Association Francaise de Normalisation. Collection of French Standards for Fats, Oleaginous Grains and Derived Products, 3rd ed.; French Association for Standardization, Paris (AFNOR): Paris, France, 1984; p. 95. [Google Scholar]
- Official Methods of Analysis of AOAC International. Agriculture Chemicals; Contaminants, Drugs, 16th ed.; AOAC International: Arlington, VA, USA, 1995; Volume 1. [Google Scholar]
- Kotzamanis, Y.; Kouroupakis, E.; Ilia, V.; Haralabous, J.; Papaioannou, N.; Papanna, K.; Richards, R.; Gisbert, E. Effects of high-level fishmeal replacement by plant proteins supplemented with different levels of lysine on growth performance and incidence of systemic non-infectious granulomatosis in meagre (Argyrosomus regius). Aquac. Nutr. 2018, 24, 1738–1751. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. In Handbook of Food Analytical Chemistry; Wiley Publishes: Hoboken, NJ, USA, 2005; Volume 2, pp. 171–178. [Google Scholar]
- Mohamed, M.S.; Tan, J.S.; Kadkhodaei, S.; Mohamad, R.; Mokhtar, M.N.; Ariff, A.B. Kinetics and modeling of microalga Tetraselmis sp. FTC 209 growth with respect to its adaptation toward different trophic conditions. Biochem. Eng. J. 2014, 88, 30–41. [Google Scholar] [CrossRef]
- Dammak, M.; Haase, S.M.; Miladi, R.; Ben Amor, F.; Barkallah, M.; Gosset, D.; Pichon, C.; Huchzermyer, B.; Fendri, I.; Denis, M.; et al. Enhanced lipid and biomass production by a newly isolated and identified marine microalga. Lipids Health Dis. 2016, 15, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Abu-Rezq, T.S.; Al-Musallam, L.; Al-Shimmari, J.; Dias, P. Optimum production conditions for different high-quality marine algae. Hydrobiologia 1999, 403, 97–107. [Google Scholar] [CrossRef]
- Laroche, C. Exopolysaccharides from Microalgae and Cyanobacteria: Diversity of Strains, Production Strategies, and Applications. Mar. Drugs 2022, 20, 336. [Google Scholar] [CrossRef]
- Kamil, W.A. Low cost technique for Tetraselmis chuii (Butcher) production in different nutritive conditions as a live feed diet for shrimp hatchery. Egypt. J. Aquat. Res. 2009, 35, 355–362. [Google Scholar]
- Farahin, A.W.; Natrah, I.; Nagao, N.; Yusoff, F.M.; Shariff, M.; Banerjee, S.; Katamaya, T.; Nakakuni, M.; Koyama, M.; Nakasaki, K.; et al. Tolerance of Tetraselmis tetrathele to High Ammonium Nitrogen and Its Effect on Growth Rate, Carotenoid and Fatty Acids Productivity. Front. Bioeng. Biotechnol. 2021, 9, 568776. [Google Scholar] [CrossRef]
- Li, X.; Li, W.; Zhai, J.; Wei, H.; Wang, Q. Effect of ammonium nitrogen on microalgal growth, biochemical composition and photosynthetic performance in mixotrophic cultivation. Bioresour. Technol. 2019, 273, 368–376. [Google Scholar] [CrossRef]
- Kokkinos, N.; Lazaridou, A.; Stamatis, N.; Orfanidis, S.; Mitropoulos, A.C.; Christoforidis, A.; Nikolaou, N. Biodiesel production from selected microalgae strains and determination of its properties and combustion specific characteristics. J. Eng. Sci. Technol. Rev. 2015, 8, 1–6. [Google Scholar] [CrossRef]
- Kumar, S.; Saramma, A.V. Effect of Salinity and pH Ranges on the Growth and Biochemical Composition of Marine Microalga-Nannochloropsis salina. Int. J. Agric. Environ. Biotechnol. 2018, 11, 651–660. [Google Scholar] [CrossRef]
- Zaher, S.S.; Helal, A.M. How culture medium pH range influence phytoplankton growth performance and biochemical content. Egypt. J. Aquat. Biol. Fish. 2020, 24, 103–116. [Google Scholar] [CrossRef]
- Dammak, M.; Hadrich, B.; Barkallah, M.; Hentati, F.; Ben Hlima, H.; Pichon, C.; Denis, M.; Fendi, I.; Michaud, P.; Abdelkafi, S. Modelling Tetraselmis sp. growth-kinetics and optimizing bioactive-compound production through environmental conditions. Bioresour. Technol. 2018, 249, 510–518. [Google Scholar] [CrossRef]
- Chen, S.Y.; Pan, L.Y.; Hong, M.J.; Lee, A.C. The effects of temperature on the growth of and ammonia uptake by marine microalgae. Bot. Stud. 2012, 53, 125–133. [Google Scholar]
- Sheng, J.; Kim, H.W.; Badalamenti, J.P.; Zhou, C.; Sridharakrishnan, S.; Krajimalnik-Brown, R.; Rittman, B.E.; Vannela, R. Effects of temperature shifts on growth rate and lipid characteristics of Synechocystis sp. PCC6803 in a bench-top photobioreactor. Bioresour. Technol. 2011, 102, 11218–11225. [Google Scholar] [CrossRef]
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; Del Borghi, M. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculate and Chlorella vulgaris for biodiesel production. Chem. Eng. Process. Process Intensif. 2009, 48, 1146–1151. [Google Scholar] [CrossRef]
- Wei, L.; Huang, X.; Huang, Z. Temperature effects on lipid properties of microalgae Tetraselmis subcordiformis and Nannochloropsis oculata as biofuel resources. Chin. J. Oceanol. Limnol. 2015, 33, 99–106. [Google Scholar] [CrossRef]
- Arkronrat, W.; Deemark, P.; Oniam, V. Growth performance and proximate composition of mixed cultures of marine microalgae (Nannochloropsis sp. & Tetraselmis sp.) with monocultures. Songklanakarin J. Sci. Technol. 2016, 38, 1–5. [Google Scholar]
- Schulze, P.S.C.; Carvalho, C.F.; Pereira, H.; Gangadhar, K.N.; Schüler, L.M.; Santos, T.F.; Varela, J.C.; Barreira, L. Urban wastewater treatment by Tetraselmis sp. CTP4 (Chlorophyta). Bioresour. Technol. 2017, 223, 175–183. [Google Scholar] [CrossRef]
- Schüler, L.M.; Bombo, G.; Duarte, P.; Santos, T.F.; Maia, I.B.; Pinheiro, F.; Marques, J.; Jacinto, R.; Schulze, P.S.C.; Pereira, H.; et al. Carotenoid biosynthetic gene expression, pigment and n-3 fatty acid contents in carotenoid-rich Tetraselmis striata CTP4 strains under heat stress combined with high light. Bioresour. Technol. 2021, 337, 125385. [Google Scholar] [CrossRef]
- Tsolcha, O.N.; Tekerlekopoulou, A.G.; Akratos, C.S.; Aggelis, G.; Genitsaris, S.; Moustaka-Gouni, M.; Vayenas, D.V. Biotreatment of raisin and winery wastewaters and simultaneous biodiesel production using a Leptolyngbya-based microbial consortium. J. Clean. Prod. 2017, 148, 185–193. [Google Scholar] [CrossRef]
- Fakhri, M.; Sanudi; Arifin, N.B.; Ekawati, A.W.; Yuniarti, A.; Hariati, A.M. Effect of photoperiod regimes on growth, biomass and pigment content of Nannochloropsis sp. BJ17. Asian J. Microbiol. Biotechnol. Environ. Sci. 2017, 19, 263–267. [Google Scholar]
- Chauton, M.S.; Winge, P.; Brembu, T.; Vadstein, O.; Bones, A.M. Gene regulation of carbon fixation, storage, and utilization in the diatom Phaeodactylum tricornutum acclimated to light/dark cycles. Plant Physiol. 2013, 161, 1034–1048. [Google Scholar] [CrossRef] [Green Version]
- Olaizola, M.; Duerr, E.O.; Freeman, D.W. Effect of CO2 enhancement in an outdoor algal production system using Tetraselmis. J. Appl. Phycol. 1991, 3, 363–366. [Google Scholar] [CrossRef]
- Tahir, A.; Rukminasari, N.; Yaqin, K.; Lukman, M. Increasing CO2 concentration impact upon nutrient absorption and removal efficiency of supra intensive shrimp pond wastewater by marine microalgae Tetraselmis chui. Int. J. Phytoremediation 2020, 23, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Bondioli, P.; Della Bella, L.; Rivolta, G.; Chini Zittelli, G.; Bassi, N.; Rodolfi, L.; Casini, D.; Prussi, M.; David, C.; Tredici, M.R. Oil production by the marine microalgae Nannochloropsis sp. F&M-M24 and Tetraselmis suecica F&M-M33. Bioresour. Technol. 2012, 114, 567–572. [Google Scholar] [PubMed]
- Kandasamy, L.C.; Neves, M.A.; Demura, M.; Nakajima, M. The effects of total dissolved carbon dioxide on the growth rate, biochemical composition, and biomass productivity of nonaxenic microalgal polyculture. Sustainability 2021, 13, 2267. [Google Scholar] [CrossRef]
- Varshney, P.; Beardall, J.; Bhattacharya, S.; Wangikar, P.P. Effect of elevated carbon dioxide and nitric oxide on the physiological responses of two green algae, Asterarcys quadricellulare and Chlorella sorokiniana. J. Appl. Phycol. 2020, 32, 189–204. [Google Scholar] [CrossRef]
- Qi, M.; Yao, C.; Sun, B.; Cao, X.; Fei, Q.; Liang, B.; Ran, W.; Xiang, Q.; Zhang, Y.; Lan, X. Application of an in situ CO2-bicarbonate system under nitrogen depletion to improve photosynthetic biomass and starch production and regulate amylose accumulation in a marine green microalga Tetraselmis subcordiformis. Biotechnol. Biofuels 2019, 12, 1–21. [Google Scholar] [CrossRef]
- White, D.A.; Pagarette, A.; Rooks, P.; Ali, S.T. The effect of sodium bicarbonate supplementation on growth and biochemical composition of marine microalgae cultures. J. Appl. Phycol. 2013, 25, 153–165. [Google Scholar] [CrossRef]
- Patil, L.; Kaliwal, B. Effect of CO2 Concentration on Growth and Biochemical Composition of Newly Isolated Indigenous Microalga Scenedesmus bajacalifornicus BBKLP-07. Appl. Biochem. Biotechnol. 2017, 182, 335–348. [Google Scholar] [CrossRef]
- Darwish, R.; Gedi, M.A.; Akepach, P.; Assaye, H.; Zaky, A.S.; Gray, D.A. Chlamydomonas reinhardtii is a potential food supplement with the capacity to outperform Chlorella and Spirulina. Appl. Sci. 2020, 10, 6736. [Google Scholar] [CrossRef]
- Almutairi, A.W.; Toulibah, H.E. Effect of Salinity and pH on Fatty Acid Profile of The Green Algae Tetraselmis suecica. J. Pet. Environ. Biotechnol. 2017, 8, 1–6. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef]
- Mandotra, S.P.; Kumar, M.; Suseela, M.R.; Nayaka, S.; Ramteke, P.W. Evaluation of fatty acid profile and biodiesel properties of microalga Scenedesmus abundans under the influence of phosphorus, pH and light intensities. Bioresour. Technol. 2016, 201, 222–229. [Google Scholar] [CrossRef]
- Sakarika, M.; Kornaros, M. Effect of pH on growth and lipid accumulation kinetics of the microalga Chlorella vulgaris grown heterotrophically under sulfur limitation. Bioresour. Technol. 2016, 219, 694–701. [Google Scholar] [CrossRef]
- Brindhadevi, K.; Mathimani, T.; Renec, E.R.; Shanmugamd, S.; Lan Chie, N.T.; Pugazhendhie, A. Impact of cultivation conditions on the biomass and lipid in microalgae with an emphasis on biodiesel. Fuel 2021, 284, 119058. [Google Scholar] [CrossRef]
- Schüler, L.M.; Schulze, P.S.C.; Pereira, H.; Barreira, I.; León, R.; Varela, J. Trends and strategies to enhance triacylglycerols and high-value compounds in microalgae. Algal Res. 2017, 25, 263–273. [Google Scholar] [CrossRef]
- Wang, Y.; He, B.; Sun, Z.; Chen, Y.F. Chemically enhanced lipid production from microalgae under low sub-optimal temperature. Algal Res. 2016, 16, 20–27. [Google Scholar] [CrossRef]
- Mai, T.D.; Lee-Chang, K.J.; Jameson, I.D.; Hoang, T.; Cai, N.B.A.; Pham, H.Q. Fatty acid profiles of selected microalgae used as live feeds for shrimp postlarvae in Vietnam. Aquac. J. 2021, 1, 26–38. [Google Scholar] [CrossRef]
- Chua, E.T.; Dal’Molin, C.; Thomas-Hall, S.; Netzel, M.E.; Netzel, G.; Schenk, P.M. Cold and dark treatments induce omega-3 fatty acid and carotenoid production in Nannochloropsis oceanica. Algal Res. 2020, 51, 102059. [Google Scholar] [CrossRef]
- Lim, K.C.; Zaleha, K. Effect of photoperiod on the cellular fatty acid composition of three tropical marine microalgae. Malays. J. Appl. Biol. 2013, 42, 41–49. [Google Scholar]
- Ma, C.; Zhang, Y.; Ho, S.; Xing, D.; Ren, N.; Liu, B. Cell growth and lipid accumulation of a microalgal mutant Scenedesmus sp. Z-4 by combining light/dark cycle with temperature variation. Biotechnol. Biofuels 2017, 10, 260–272. [Google Scholar] [CrossRef] [Green Version]
- Vendruscolo, R.G.; Fagundes, M.B.; Maroneze, M.M.; do Nascimento, T.C.; de Menezes, C.R.; Barin, J.S.; Zepka, L.Q.; Jacob-Lopes, E.; Wagner, R. Scenedesmus obliquus metabolomics: Effect of photoperiods and cell growth phases. Bioprocess Biosyst. Eng. 2019, 42, 727–739. [Google Scholar] [CrossRef]
- Srirangan, S.; Sauer, M.L.; Howard, B.; Dvora, M.; Dums, J.; Backman, P.; Sederoff, H. Interaction of temperature and photoperiod increases growth and oil content in the marine microalgae Dunaliella viridis. PLoS ONE 2015, 10, e0127562. [Google Scholar] [CrossRef]
- De Jesus, P.C.C.; Mendes, M.A.; Perpétuo, E.A.; Basso, T.O.; do Nascimento, C.A.O. Extracellular carotenoid production and fatty acids profile of Parachlorella kessleri under increased CO2 concentrations. J. Biotechnol. 2011, 329, 151–159. [Google Scholar] [CrossRef]
- Farhadian, O.; Yusoff, F.; Mohamed, S. Nutritional values of Apocyclops dengizicus (Copepoda: Cyclopoida) fed Chaetocerous calcitrans and Tetraselmis tetrathele. Aquac. Res. 2008, 40, 74–82. [Google Scholar] [CrossRef]
- Xiang, Q.; Wei, X.; Yang, Z.; Xie, T.; Zhang, Y.; Li, D.; Pan, X.; Liu, X.; Zhang, X.; Yao, C. Acclimation to a broad range of nitrate strength on a euryhaline marine microalga Tetraselmis subcordiformis for photosynthetic nitrate removal and high-quality biomass production. Sci. Total Environ. 2021, 781, 146687. [Google Scholar] [CrossRef]
- De Oliveira, M.; Monteiro, M.; Robbs, P.; Leite, S. Growth and Chemical Composition of Spirulina maxima and Spirulina platensis Biomass at Different Temperatures. Aquac. Int. 1999, 7, 261–275. [Google Scholar] [CrossRef]
- Schüler, L.; Gangadhar, K.; Duarte, P.; Placines, C.; Molina-Márquez, A.; Léon-Bañares, R.; Sousa, V.; Varela, J.; Barreira, L. Improvement of carotenoid extraction from a recently isolated, robust microalga, Tetraselmis sp. CTP4 (chlorophyta). Bioprocess Biosyst. Eng. 2020, 43, 785–796. [Google Scholar] [CrossRef]
- Giannelli, L.; Yamada, H.; Katsuda, T.; Yamaji, H. Effects of temperature on the astaxanthin productivity and light harvesting characteristics of the green alga Haematococcus pluvialis. J. Biosci. Bioeng. 2015, 119, 345–350. [Google Scholar] [CrossRef]
- Domínguez-Bocanegra, A.R.; Guerrero Legarreta, I.; Martinez Jeronimo, F.; Tomasini Campocosio, A. Influence of environmental and nutritional factors in the production of astaxanthin from Haematococcus pluvialis. Bioresour. Technol. 2004, 92, 209–214. [Google Scholar] [CrossRef]
- Olaizola, M. Commercial production of astaxanthin from Haematococcus pluvialis using 25,000 Liter photobioreactors. J. Appl. Phycol. 2000, 12, 499–506. [Google Scholar] [CrossRef]
- do Nascimento, T.; Nass, P.; Fernandes, A.; Vieira, K.; Wagner, R.; Jacob-Lopes, E.; Zepka, L. Exploratory data of the microalgae compounds for food purposes. Data Br. 2020, 29, 105182. [Google Scholar] [CrossRef] [PubMed]
- Vendruscolo, R.; Fernandes, A.; Fagundes, M.; Zepka, L.; de Menezes, C.; Jacob–Lopes, E.; Wagner, R. Development of a new method for simultaneous extraction of chlorophylls and carotenoids from microalgal biomass. J. Appl. Phycol. 2021, 33, 1987–1997. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial Concentrations (mg L−1) | ||||||
|---|---|---|---|---|---|---|
| Growth Substrate | NH4+-N | TN | PO43− | Total Sugars | Biomass | N:P |
| A 1 | 18.6 ± 0.2 | 34.3 ± 2.1 | 6.5 ± 1.0 | 19.6 ± 3.3 | 98 ± 10.0 | 5:1 |
| B 2 | 63.5 ± 1.4 | 72.4 ± 2.8 | 6.0 ± 0.2 | 29.6 ± 0.3 | 100 ± 5.6 | 12:1 |
| C 3 | 62.7 ± 2.6 | 75.0 ± 3.3 | 6.2 ± 0.3 | 17.2 ± 0.3 | 120 ± 28.3 | 12:1 |
| D 4 | 30.5 ± 2.3 | 44.4 ± 3.8 | 4.4 ± 0.1 | 8.0 ± 1.1 | 80 ± 19.7 | 10:0 |
| E 5 | 5.1 ± 2.2 | 16.1 ± 0.9 | 2.2 ± 0.1 | 14.8 ± 0.1 | 125 ± 16.1 | 7:3 |
| F 6 | 5.4 ± 0.5 | 16.2 ± 0.8 | 2.1 ± 0.01 | 10.1 ± 2.3 | 130 ± 39.0 | 7:7 |
| % Removal Rate | Maximum Biomass Productivity (mg L−1 d−1) | Specific Growth Rate (d−1) | % Maximum Lipid Content | ||||
|---|---|---|---|---|---|---|---|
| Growth Substrate | NH4+-N | TN | PO43− | Total Sugars | |||
| A 1 | 89.0 ± 0.3 | 34.8 ± 1.3 | 58.3 ± 1.2 | 0.0 | 37.5 ± 4.4 | 0.101 ± 0.01 | 16.0 ± 2.02 |
| B 2 | 85.4 ± 2.2 | 52.5 ± 4.3 | 87.2 ± 0.1 | 0.0 | 40.0 ± 2.3 | 0.129 ± 0.01 | 10.3 ± 0.8 |
| C 3 | 76.2 ± 1.0 | 51.1 ± 3.2 | 94.9 ± 0.8 | 0.0 | 47.6 ± 9.8 | 0.184 ± 0.01 | 13.7 ± 2.4 |
| D 4 | 98.4 ± 2.3 | 51.0 ± 0.4 | 97.8 ± 0.2 | 0.0 | 58.2 ± 10.7 | 0.227 ± 0.03 | 24.2 ± 2.8 |
| E 5 | 95.1 ± 0.1 | 54.6 ± 2.4 | 95.5 ± 0.1 | 0.0 | 62.0 ± 13.8 | 0.200 ± 0.05 | 16.4 ± 4.6 |
| F 6 | 97.2 ± 0.6 | 56.0 ± 0.5 | 97.5 ± 0.1 | 0.0 | 70.1 ± 11.2 | 0.240 ± 0.04 | 16.5 ± 2.3 |
| Fertilizer Quantity (g L−1) | Initial Concentration of NH4+-N (mg L−1) | % Removal Rate of NH4+-N | Initial Biomass Concentration (mg L−1) | Maximum Biomass Productivity (mg L−1 d−1) | Specific Growth Rate (d−1) | % Maximum Lipid Content |
|---|---|---|---|---|---|---|
| 0.1 | 6.3 ± 0.2 | 97.1 ± 0.2 | 92.5 ± 10.6 | 81.8 ± 6.8 | 0.236 ± 0.09 | 15.9 ± 2.5 |
| 0.2 | 10.0 ± 0.6 | 97.7 ± 0.1 | 112.1 ± 17.1 | 84.2 ± 3.2 | 0.240 ± 0.05 | 16.3 ± 2.1 |
| 0.4 | 16.0 ± 2.8 | 88.7± 0.3 | 115.0 ± 57.8 | 62.8 ± 4.3 | 0.212 ± 0.07 | 16.1 ± 1.2 |
| 0.6 | 25.8 ± 2.0 | 90.0 ± 0.3 | 115.0 ± 45.4 | 60.7 ± 3.3 | 0.127 ± 0.02 | 14.7 ± 0.7 |
| 0.8 | 33.0 ± 3.5 | 41.6 ± 0.6 | 108.5 ± 12.0 | 66.7 ± 2.8 | 0.213 ± 0.04 | 12.7 ± 5.1 |
| 1.0 | 43.1 ± 2.7 | 41.0 ± 0.5 | 109.8 ± 0.3 | 59.7 ± 3.1 | 0.183 ± 0.02 | 9.8 ± 0.7 |
| Initial Concentrations (mg L−1) | ||||||||
|---|---|---|---|---|---|---|---|---|
| pH Value | NH4+-N | TN | PO43− | Total Sugars | d-COD | Biomass | C:N | N:P |
| 8 | 24.2 ± 0.6 | 93.9 ± 4.0 | 12.3 ± 0.2 | 5.8 ± 1.5 | 115.3 ± 5.6 | 87.0 ± 26.8 | 1.23 | 7.64 |
| 7 | 23.1 ± 0.1 | 82.1 ± 0.3 | 12.1 ± 0.1 | 6.0 ± 0.1 | 94.6 ± 16.9 | 60.0 ± 2.8 | 1.15 | 6.78 |
| % Removal Rate | Maximum Biomass Productivity (mg L−1 d−1) | Specific Growth Rate (d−1) | |||||
|---|---|---|---|---|---|---|---|
| pH Value | NH4+-N | TN | PO43− | Total sugars | d-COD | ||
| 8 | 98.7 ± 0.1 | 79.1± 2.5 | 99 ± 0.2 | 0.0 | 94.8 ± 2.8 | 79.8 ± 1.4 | 0.266 ± 0.06 |
| 7 | 94.6 ± 0.4 | 76.4 ± 0.1 | 97.8 ± 0.4 | 0.0 | 56.3 ± 9.1 | 60.1 ± 5.5 | 0.248 ± 0.05 |
| % d.w. Content | |||||
|---|---|---|---|---|---|
| pH Value | Proteins | Lipids | Carbohydrates | Total Chlorophylls | Total Carotenoids |
| 8 | 51.2 ± 2.0 | 27.8 ± 1.0 | 14.6 ± 1.5 | 5.1 ± 0.1 | 0.20 ± 0.01 |
| 7 | 47.7 ± 0.5 | 26.4 ± 0.4 | 14.7 ± 0.8 | 4.6 ± 0.1 | 0.88 ± 0.10 |
| Initial Concentrations (mg L−1) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Temperature | NH4+-N | TN | PO43− | Total Sugars | d-COD | Biomass | C:N | N:P |
| 19 ± 1 °C | 30.4 ± 0.4 | 84.9 ± 3.0 | 9.9 ± 0.1 | 2.6 ± 1.1 | 76.7 ± 4.7 | 48.8 ± 18.2 | 0.90 | 8.51 |
| 25 ± 1 °C | 34.3 ± 2.8 | 92.9 ± 2.9 | 11.5 ± 0.1 | 2.3 ± 5.2 | 92.0 ± 0.2 | 42.9 ± 6.9 | 1.00 | 8.10 |
| 28 ± 1 °C | 31.7 ± 3.3 | 89.8 ± 3.6 | 11.7 ± 0.2 | 2.5 ± 0.01 | 22.0 ± 0.3 | 69 ± 26.8 | 0.24 | 7.71 |
| % Removal Rate | Maximum Biomass Productivity (mg L−1 d−1) | Specific Growth Rate (d−1) | |||||
|---|---|---|---|---|---|---|---|
| Temperature | NH4+-N | TN | PO43− | Total Sugars | d-COD | ||
| 19 ± 1 °C | 99.8 ± 0.1 | 99.1 ± 0.1 | 99.7 ± 0.2 | 0.0 | 53.0 ± 2.3 | 69.3 ± 3.9 | 0.237 ±0.07 |
| 25 ± 1 °C | 99.2 ± 0.1 | 95.5 ± 1.6 | 100.0 ± 0.1 | 0.0 | 72.5 ± 0.9 | 93.7 ± 11.5 | 0.283 ±0.09 |
| 28 ± 1 °C | 94.5 ±0.1 | 95.4 ± 2.1 | 99.6 ± 0.1 | 0.0 | 0.0 | 61.5 ± 1.3 | 0.223 ±0.06 |
| % d.w. Content | |||||
|---|---|---|---|---|---|
| Temperature | Proteins | Lipids | Carbohydrates | Total Chlorophylls | Total Carotenoids |
| 19 ± 1 °C | 34.8 ± 1.0 | 26.7 ± 1.7 | 16.3 ± 1.3 | 3.4 ± 0.10 | 0.42 ± 0.03 |
| 25 ± 1 °C | 38.7 ± 1.2 | 30.2 ± 1.5 | 20.4 ± 2.2 | 5.1 ± 1.20 | 0.51 ± 0.10 |
| 28 ± 1 °C | 41.6 ± 2.1 | 26.2 ± 1.3 | 13.5 ± 0.4 | 2.7 ± 0.05 | 0.27 ± 0.05 |
| Initial Concentrations (mg L−1) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Photoperiod | NH4+-N | TN | PO43− | Total Sugars | d-COD | Biomass | C:N | N:P |
| 20:4 h L:D | 21.6 ± 1.2 | 80.4 ± 1.0 | 9.9 ± 0.1 | 1.9 ± 0.5 | 26.0 ± 4.7 | 70.3 ± 10.6 | 0.32 | 8.07 |
| 18:6 h L:D | 31.7 ± 2.8 | 84.7 ± 4.2 | 10.7± 0.1 | 1.2 ± 0.7 | 84.7 ± 0.9 | 48.3 ± 7.1 | 1.00 | 7.85 |
| 12:12 h L:D | 15.6 ± 1.5 | 77.2 ± 6.1 | 9.7 ± 0.2 | 2.1 ± 0.1 | 77.7 ± 6.1 | 46.4 ± 4.4 | 1.00 | 7.94 |
| % Removal Rate | Maximum Biomass Productivity (mg L−1 d−1) | Specific Growth Rate (d−1) | |||||
|---|---|---|---|---|---|---|---|
| Photoperiod | NH4+-N | TN | PO43− | Total Sugars | d-COD | ||
| 20:4 h L:D | 62.6 ± 1.8 | 74.00 ± 2.7 | 98.7 ± 0.10 | 0.0 | 0.0 | 46.3 ± 3.1 | 0.136 ± 0.04 |
| 18:6 h L:D | 52.0 ± 3.8 | 76.80 ± 6.1 | 97.7 ± 0.03 | 0.0 | 47.2 ± 8.0 | 36.3 ± 4.2 | 0.208 ± 0.08 |
| 12:12 h L:D | 47.8 ± 0.5 | 61.01 ± 2.5 | 96.8 ± 0.01 | 0.0 | 68.8 ± 2.3 | 30.0 ± 2.6 | 0.118 ± 0.03 |
| Control set | |||||||
| 24:0 h L:D | 99.2 ± 0.1 | 95.5 ± 1.6 | 100.0 ± 0.1 | 0.0 | 72.5 ± 0.9 | 93.7 ± 11.5 | 0.283 ± 0.09 |
| % d.w. Content | |||||
|---|---|---|---|---|---|
| Photoperiod | Proteins | Lipids | Carbohydrates | Total Chlorophylls | Total Carotenoids |
| 20:4 h L:D | 53.2 ± 0.5 | 28.5 ± 3.8 | 15.6 ± 1.4 | 4.2 ± 0.1 | 0.60 ± 0.10 |
| 18:6 h L:D | 50.4 ± 1.2 | 25.8 ± 1.1 | 14.4 ± 1.6 | 2.8 ± 0.6 | 0.53 ± 0.01 |
| 12:12 h L:D | 54.0 ± 1.8 | 24.7 ± 2.5 | 13.8 ± 0.5 | 3.8 ± 0.4 | 0.59 ± 0.20 |
| Control set | |||||
| 24:0 h L:D | 38.7 ± 1.2 | 30.2 ± 1.5 | 20.4 ± 2.2 | 5.1 ± 1.2 | 0.51 ± 0.10 |
| Initial Concentrations (mg L−1) | ||||||||
|---|---|---|---|---|---|---|---|---|
| CO2 Flow Rate | NH4+-N | TN | PO43− | Total Sugars | d-COD | Biomass | C:N | N:P |
| 10 mL min−1 | 20.2 ± 1.9 | 87.5 ± 1.3 | 10.9 ± 0.1 | 2.4 ± 0.2 | 28.4 ± 5.2 | 83.3 ± 7.8 | 0.32 | 8.03 |
| 20 mL min−1 | 17.8 ± 2.3 | 78.4 ± 3.2 | 9.5 ± 1.4 | 3.4 ± 0.1 | 88.9 ± 0.2 | 45.0 ± 15.0 | 1.13 | 8.21 |
| 90 mL min−1 | 14.5 ± 0.4 | 80.8 ± 2.5 | 10.7 ± 1.7 | 2.8 ± 0.7 | 43.3 ± 4.2 | 84.0 ± 5.6 | 0.54 | 7.60 |
| % Removal Rate | Maximum Biomass Productivity (mg L−1 d−1) | Specific Growth Rate (d−1) | |||||
|---|---|---|---|---|---|---|---|
| CO2 Flow Rate | NH4+-N | TN | PO43− | Total Sugars | d-COD | ||
| 10 mL min−1 | 99.6 ± 0.1 | 91.8 ± 0.4 | 68.0 ± 0.8 | 0.0 | 21.0 ± 3.5 | 87.5 ± 16.4 | 0.262 ± 0.01 |
| 20 mL min−1 | 82.8 ± 1.9 | 78.1 ± 1.2 | 52.8 ± 0.7 | 0.0 | 0.0 | 73.8 ± 10.1 | 0.216 ± 0.07 |
| 90 mL min−1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 20.3 ± 2.2 | 0.050 ± 0.01 |
| % d.w. Content | |||||
|---|---|---|---|---|---|
| CO2 Flow Rate | Proteins | Lipids | Carbohydrates | Total Chlorophylls | Total Carotenoids |
| 10 mL min−1 | 44.6 ± 0.8 | 27.3 ± 1.5 | 10.3 ± 2.6 | 5.2 ± 0.3 | 1.13 ± 0.3 |
| 20 mL min−1 | 44.5 ± 0.7 | 22.5 ± 4.5 | 11.5 ± 1.0 | 3.5 ± 0.4 | 0.78 ± 0.1 |
| 90 mL min−1 | 25.4 ± 0.2 | 19.0 ± 2.6 | 11.5 ± 0.7 | 1.4 ± 0.07 | 0.19 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patrinou, V.; Daskalaki, A.; Kampantais, D.; Kanakis, D.C.; Economou, C.N.; Bokas, D.; Kotzamanis, Y.; Aggelis, G.; Vayenas, D.V.; Tekerlekopoulou, A.G. Optimization of Cultivation Conditions for Tetraselmis striata and Biomass Quality Evaluation for Fish Feed Production. Water 2022, 14, 3162. https://doi.org/10.3390/w14193162
Patrinou V, Daskalaki A, Kampantais D, Kanakis DC, Economou CN, Bokas D, Kotzamanis Y, Aggelis G, Vayenas DV, Tekerlekopoulou AG. Optimization of Cultivation Conditions for Tetraselmis striata and Biomass Quality Evaluation for Fish Feed Production. Water. 2022; 14(19):3162. https://doi.org/10.3390/w14193162
Chicago/Turabian StylePatrinou, Vasiliki, Alexandra Daskalaki, Dimitris Kampantais, Dimitris C. Kanakis, Christina N. Economou, Dimitris Bokas, Yannis Kotzamanis, George Aggelis, Dimitris V. Vayenas, and Athanasia G. Tekerlekopoulou. 2022. "Optimization of Cultivation Conditions for Tetraselmis striata and Biomass Quality Evaluation for Fish Feed Production" Water 14, no. 19: 3162. https://doi.org/10.3390/w14193162
APA StylePatrinou, V., Daskalaki, A., Kampantais, D., Kanakis, D. C., Economou, C. N., Bokas, D., Kotzamanis, Y., Aggelis, G., Vayenas, D. V., & Tekerlekopoulou, A. G. (2022). Optimization of Cultivation Conditions for Tetraselmis striata and Biomass Quality Evaluation for Fish Feed Production. Water, 14(19), 3162. https://doi.org/10.3390/w14193162