
Does Water Brownification Affect Duckweeds in Freshwaters? Results from a Laboratory Experiment



Abstract
:1. Introduction
2. Material and Methods
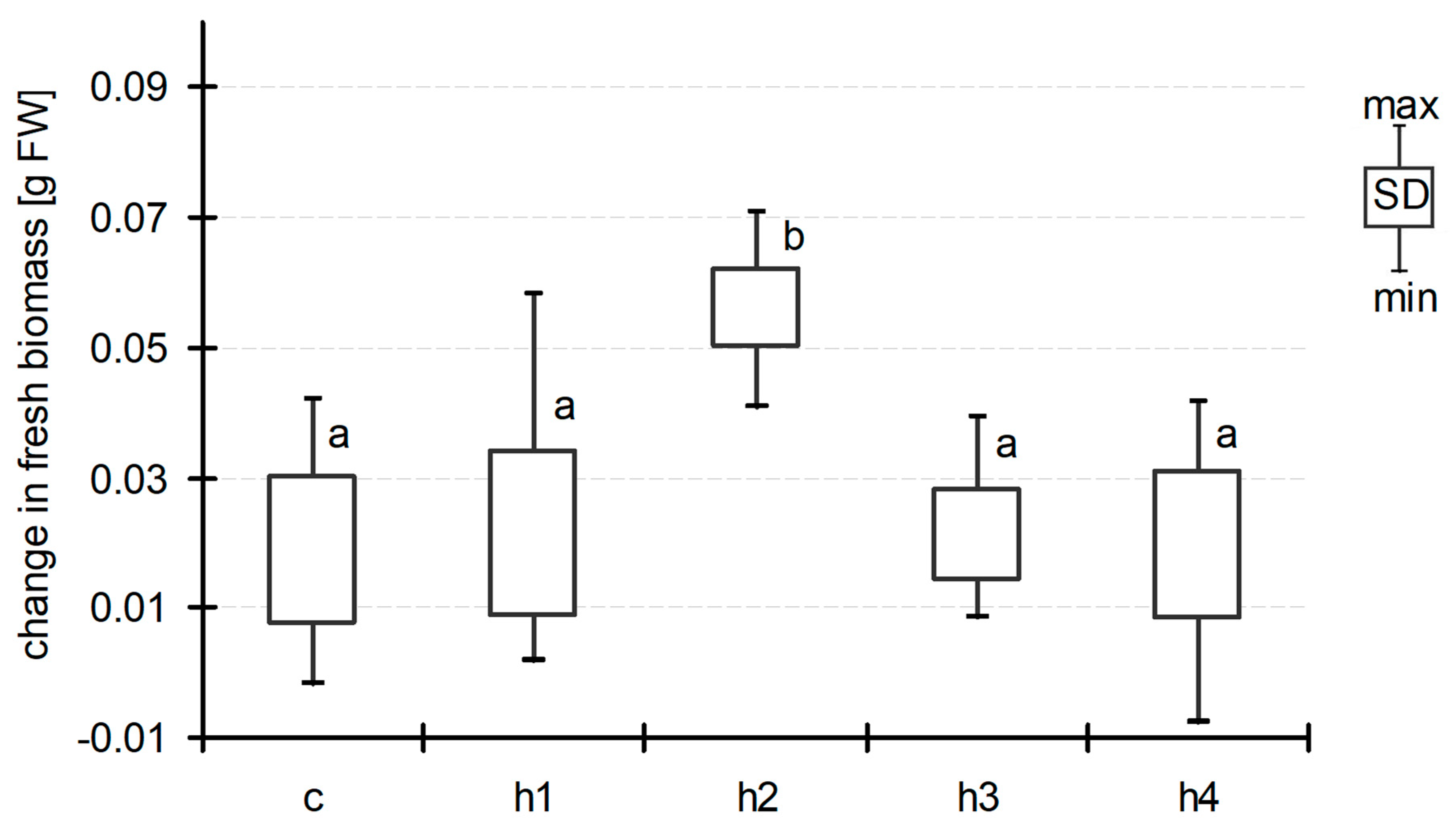
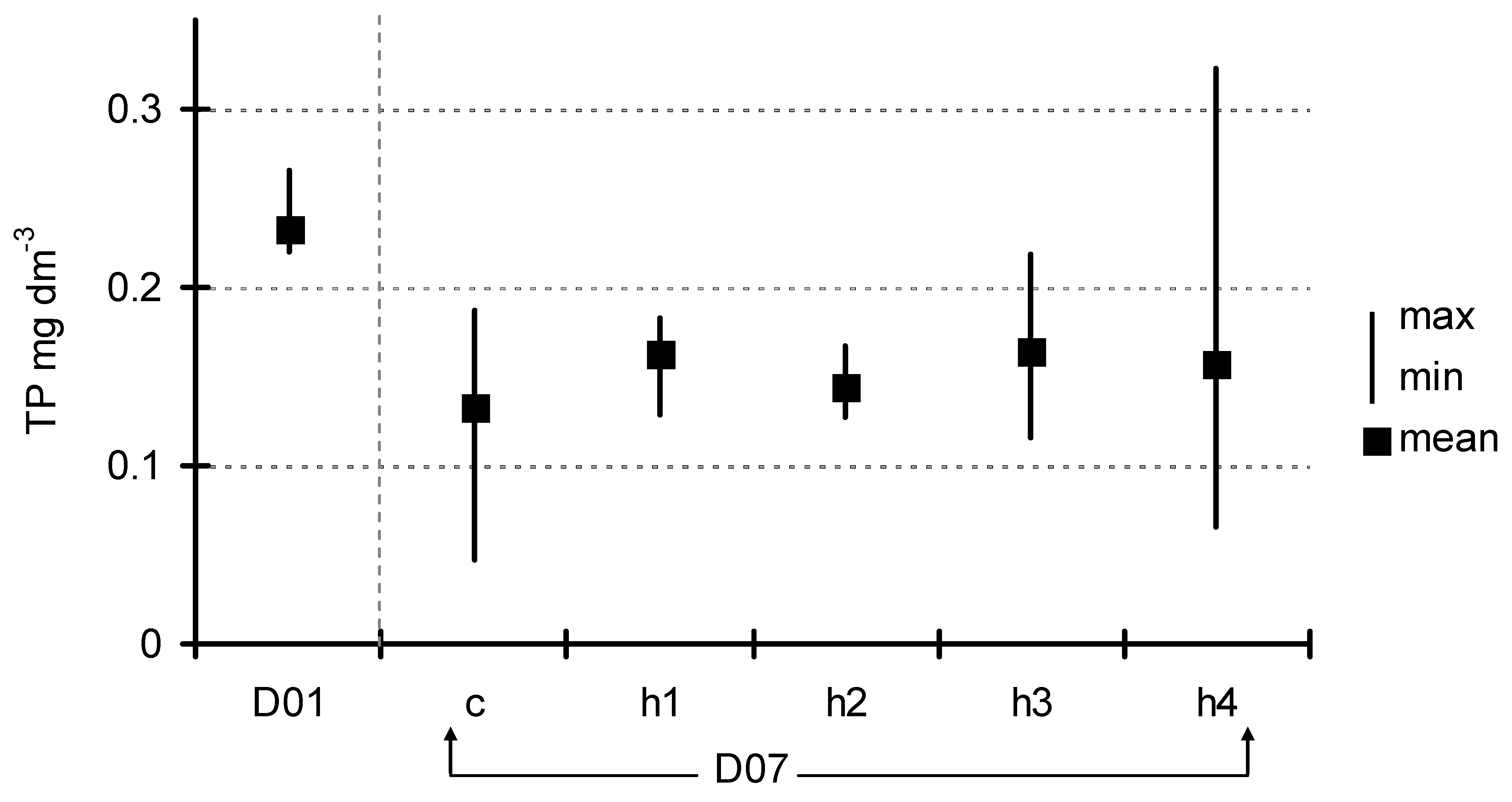
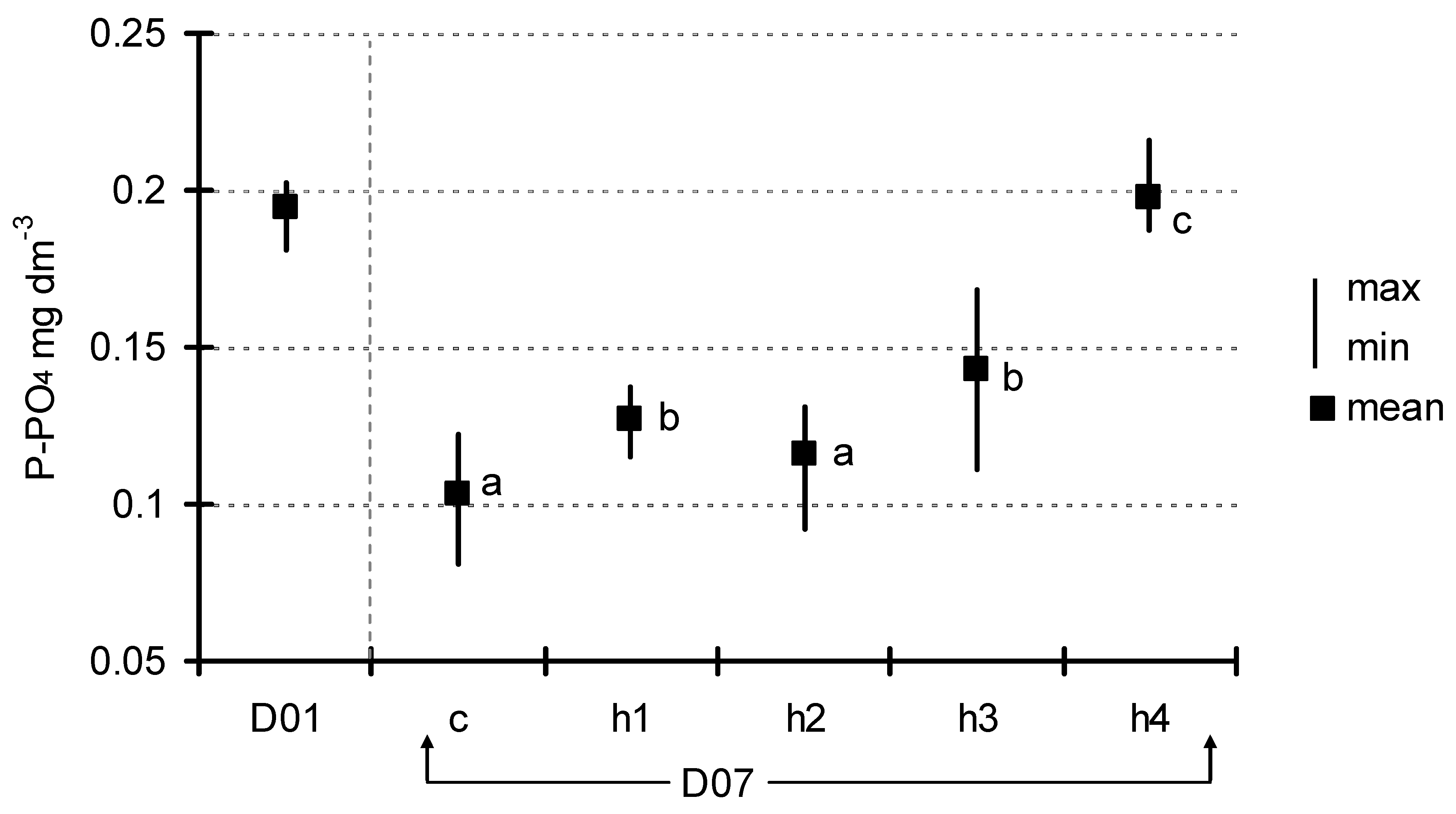
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Diffenbaugh, N.S.; Singh, D.; Mankin, J.S. Unprecedented climate events: Historical changes, aspirational targets, and national commitments. Sci. Adv. 2018, 4, eaao3354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Falconi, L. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Urban, M.C.; Bocedi, G.; Hendry, A.P.; Mihoub, J.B.; Pe’er, G.; Singer, A.; Gonzalez, A. Improving the forecast for biodiversity under climate change. Science 2016, 353, aad8466. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, D.P.; Gianoli, E.; Morris, W.F.; Bozinovic, F. Ecological and evolutionary impacts of changing climatic variability. Biol. Rev. 2017, 92, 22–42. [Google Scholar] [CrossRef] [Green Version]
- Kunkel, K.E. North American trends in extreme precipitation. Nat. Hazards 2003, 29, 291–305. [Google Scholar] [CrossRef]
- Gallego, M.C.; García, J.A.; Vaquero, J.M.; Mateos, V.L. Changes in frequency and intensity of daily precipitation over the Iberian Peninsula. J. Geophys. Res. Atmos. 2006, 111, D24105. [Google Scholar] [CrossRef]
- Chou, C.; Chen, C.A.; Tan, P.H.; Chen, K.T. Mechanisms for global warming impacts on precipitation frequency and intensity. J. Clim. 2012, 25, 3291–3306. [Google Scholar] [CrossRef]
- Ban, N.; Schmidli, J.; Schär, C. Heavy precipitation in a changing climate: Does short-term summer precipitation increase faster? Geophys. Res. Lett. 2015, 42, 1165–1172. [Google Scholar] [CrossRef]
- Hongve, D.; Riise, G.; Kristiansen, J.F. Increased colour and organic acid concentrations in Norwegian forest lakes and drinking water–a result of increased precipitation? Aquat. Sci. 2004, 66, 231–238. [Google Scholar] [CrossRef]
- Weyhenmeyer, G.A.; Willén, E.; Sonesten, L. Effects of an extreme precipitation event on water chemistry and phytoplankton in the Swedish Lake Mälaren. Boreal Environ. Res. 2004, 9, 409–420. [Google Scholar]
- Arvola, L.; Rask, M.; Ruuhijärvi, J.; Tulonen, T.; Vuorenmaa, J.; Ruoho-Airola, T.; Tulonen, J. Long-term patterns in pH and colour in small acidic boreal lakes of varying hydrological and landscape settings. Biogeochemistry 2010, 101, 269–279. [Google Scholar] [CrossRef]
- Pęczuła, W. Variability of coloured dissolved organic carbon in seepage humic lakes as related to lake morphometry, catchment characteristics and precipitation. Ecohydrology 2015, 8, 1229–1238. [Google Scholar] [CrossRef]
- de Wit, H.A.; Valinia, S.; Weyhenmeyer, G.A.; Futter, M.N.; Kortelainen, P.; Austnes, K.; Vuorenmaa, J. Current browning of surface waters will be further promoted by wetter climate. Environ. Sci. Technol. Lett. 2016, 3, 430–435. [Google Scholar] [CrossRef] [Green Version]
- Delpla, I.; Jung, A.V.; Baures, E.; Clement, M.; Thomas, O. Impacts of climate change on surface water quality in relation to drinking water production. Environ. Int. 2009, 35, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.D.; Freeman, C.; Cork, L.G.; Thomas, D.N.; Reynolds, B.; Billett, M.F.; Garnett, M.H.; Norris, D. Evidence against recent climate-induced destabilisation of soil carbon from 14C analysis of riverine dissolved organic matter. Geophys. Res. Lett. 2007, 34, L07407. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.D.; Jones, T.G.; Burden, A.; Ostle, N.; Zieliński, P.; Cooper, M.D.; Peacock, M.; Clark, J.M.; Oulehle, F.; Cooper, D.; et al. Acidity controls on dissolved organic carbon mobility in organic soils. Glob. Chang. Biol. 2012, 18, 3317–3331. [Google Scholar] [CrossRef] [Green Version]
- Creed, I.F.; Bergström, A.K.; Trick, C.G.; Grimm, N.B.; Hessen, D.O.; Karlsson, J.; Senar, O.E. Global change-driven effects on dissolved organic matter composition: Implications for food webs of northern lakes. Glob. Chang. Biol. 2018, 24, 3692–3714. [Google Scholar] [CrossRef]
- Evans, C.D.; Monteith, D.T.; Cooper, D.M. Long-term increases in surface water dissolved organic carbon, Observations, possible causes and environmental impacts. Environ. Pollut. 2005, 137, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Monteith, D.T.; Stoddard, J.L.; Evans, C.D.; de Wit, H.A.; Forsius, M.; Hogasen, T.; Wilander, A.; Skjelkvale, B.L.; Jeffries, D.S.; Vuorenmaa, J.; et al. Dissolved organic carbon trends resulting from changes in atmospheric deposition chemistry. Nature 2007, 450, 537–540. [Google Scholar] [CrossRef]
- Worrall, F.; Harriman, R.; Evans, C.D.; Watts, C.D.; Adamson, J.; Neal, C.; Stevens, P. Trends in dissolved organic carbon in UK rivers and lakes. Biogeochemistry 2004, 70, 369–402. [Google Scholar] [CrossRef]
- Hejzlar, J.; Dubrovský, M.; Buchtele, J.; Růžička, M. The apparent and potential effects of climate change on the inferred concentration of dissolved organic matter in a temperate stream (the Malše River, South Bohemia). Sci. Total Environ. 2003, 310, 143–152. [Google Scholar] [CrossRef]
- Williamson, C.E.; Morris, D.P.; Pace, M.L.; Olson, O.G. Dissolved organic carbon and nutrients as regulators of lake ecosystems, Resurrection of a more integrated paradigm. Limnol. Oceanogr. 1999, 44, 795–803. [Google Scholar] [CrossRef] [Green Version]
- Hessen, D.O.; Andersen, T.; Lyehe, A. Carbon metabolism in a humic lake: Pool sires and cycling through zooplankton. Limnol. Oceanogr. 1990, 35, 84–99. [Google Scholar] [CrossRef]
- Ask, J.; Karlsson, J.; Persson, L.; Ask, P.; Byström, P.; Jansson, M. Terrestrial organic matter and light penetration: Effects on bacterial and primary production in lakes. Limnol. Oceanogr. 2009, 54, 2034–2040. [Google Scholar] [CrossRef]
- Solomon, C.T.; Jones, S.E.; Weidel, B.C.; Buffam, I.; Fork, M.L.; Karlsson, J.; Saros, J.E. Ecosystem consequences of changing inputs of terrestrial dissolved organic matter to lakes: Current knowledge and future challenges. Ecosystems 2015, 18, 376–389. [Google Scholar] [CrossRef]
- Sřndergaard, M.; Phillips, G.; Hellsten, S.; Kolada, A.; Ecke, F.; Mäemets, H.; Oggioni, A. Maximum growing depth of submerged macrophytes in European lakes. Hydrobiologia 2013, 704, 165–177. [Google Scholar] [CrossRef]
- Landolt, E.; Kandeler, R. Phytochemistry, Physiology, Application, Bibliography. In The Family of Lemnaceae—A Monographic Study; Veroeffentlichungen des Geobotanischen Instituts der ETH Stiftung Ruebel: Zürich, Switzerland, 1988; Volume 2. [Google Scholar]
- Portielje, R.; Roijackers, R.M.M. Primary succession of aquatic macrophytes in experimental ditches in relation to nutrient input. Aquat. Bot. 1995, 50, 127–140. [Google Scholar] [CrossRef]
- Szabó, S.; Roijackers, R.; Scheffer, M.; Borics, G. The strength of limiting factors for duckweed during algal competition. Arch. Hydrobiol. 2005, 164, 127–140. [Google Scholar] [CrossRef]
- Kufel, L.; Strzałek, M.; Konieczna, A.; Izdebska, K. The effect of Stratiotes aloides L. and nutrients on the growth rate of Lemna minor L. Aquat. Bot. 2010, 92, 168–172. [Google Scholar] [CrossRef]
- Vermaat, J.E.; Hanif, M.K. Performance of common duckweed species (Lemnaceae) and the waterfern Azolla filiculoides on different types of waste water. Water Res. 1998, 32, 2569–2576. [Google Scholar] [CrossRef]
- Frederic, M.; Samir, L.; Louise, M.; Abdelkrim, A. Comprehensive modeling of mat density effect on duckweed (Lemna minor) growth under controlled eutrophication. Water Res. 2006, 40, 2901–2910. [Google Scholar] [CrossRef] [PubMed]
- Mehra, A.; Farago, M.E.; Banerjee, D.K. A study of eichhornia crassipes growing in the overbank and floodplain soils of the River Yamuna in Delhi, India. Environ. Monit. Assess. 2000, 60, 25–45. [Google Scholar] [CrossRef]
- Brendonck, L.; Maes, J.; Rommens, W.; Dekeza, N.; Nhiwatiwa, T.; Barson, M.; Marshall, B. The impact of water hyacinth (Eichhornia crassipes) in a eutrophic subtropical impoundment (Lake Chivero, Zimbabwe). II. Species diversity. Arch. Hydrobiol. 2003, 158, 389–405. [Google Scholar] [CrossRef]
- Kufel, L.; Strzałek, M.; Przetakiewicz, A. Plant response to overcrowding–Lemna minor example. Acta Oecol. 2018, 91, 73–80. [Google Scholar] [CrossRef]
- Driever, S.M.; van Nes, E.H.; Roijackers, R.M. Growth limitation of Lemna minor due to high plant density. Aquat. Bot. 2005, 81, 245–251. [Google Scholar] [CrossRef]
- Scheffer, M.; van Nes, E. Shallow lakes theory revisited: Various alternative regimes driven by climate, nutrients, depth and lake size. Hydrobiologia 2007, 584, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Mihajlović, V.; Tomić, T.; Tubić, A.; Molnar Jazić, J.; Ivančev Tumbas, I.; Šunjka, D.; Teodorović, I. The impact of humic acid on toxicity of individual herbicides and their mixtures to aquatic macrophytes. Environ. Sci. Pollut. Res. 2019, 26, 23571–23582. [Google Scholar] [CrossRef]
- Easlon, H.M.; Bloom, A.J. Easy Leaf Area: Automated digital image analysis for rapid and accurate measurement of leaf area. Appl. Plant Sci. 2014, 2, 1400033. [Google Scholar] [CrossRef]
- PN-EN ISO 6878; Jakość Wody—Oznaczanie Fosforu—Metoda Spektrometryczna z Molibdenianem Amonu. Polski Komitet Normalizacyjny: Warszawa, Poland, 2006.
- DIN EN ISO 13 395; Water Quality—Determination of Nitrite Nitrogen and Nitrate Nitrogen and the Sum of both by Flow Analysis (CFA and FIA) and Spectrometric Detection. Beuth Verlag GmbH: Berlin, Germany, 1996.
- Reitsema, R.E.; Meire, P.; Schoelynck, J. The future of freshwater macrophytes in a changing world: Dissolved organic carbon quantity and quality and its interactions with macrophytes. Front. Plant Sci. 2018, 9, 629. [Google Scholar] [CrossRef]
- Wetzel, R.G. Detrital dissolved and particulate organic carbon functions in aquatic ecosystems. Bull. Mar. Sci. 1984, 35, 503–509. [Google Scholar]
- Sachse, A.; Henrion, R.; Gelbrecht, J.; Steinberg, C.E.W. Classification of dissolved organic carbon (DOC) in river systems: Influence of catchment characteristics and autochthonous processes. Org. Geochem. 2005, 36, 923–935. [Google Scholar] [CrossRef]
- Boavida, M.J.; Wetzel, R.G. Inhibition of phosphatase activity by dissolved humic substances and hydrolytic reactivation by natural ultraviolet light. Freshw. Biol. 1998, 40, 285–293. [Google Scholar] [CrossRef]
- Sobek, S.; Tranvik, L.J.; Prairie, Y.T.; Kortelainen, P.; Cole, J.J. Patterns and regulation of dissolved organic carbon: An analysis of 7,500 widely distributed lakes. Limnol. Oceanogr. 2007, 52, 1208–1219. [Google Scholar] [CrossRef] [Green Version]
- Pflugmacher, S.; Tidwell, L.F.; Steinberg, C.E. Dissolved humic substances can directly affect freshwater organisms. Acta Hydrochim. Hydrobiol. 2001, 29, 34–40. [Google Scholar] [CrossRef]
- Wetzel, R.G. Gradient-dominated ecosystems: Sources and regulatory functions of dissolved organic matter in freshwater ecosystems. Hydrobiologia 1992, 229, 181–198. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Z.; Cheng, S.; Wu, Z. Influence of humic acids on the accumulation of copper and cadmium in Vallisneria spiralis L. from sediment. Environ. Earth Sci. 2010, 61, 1207–1213. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Pietsch, C.; Rieger, W.; Steinberg, C.E. Dissolved natural organic matter (NOM) impacts photosynthetic oxygen production and electron transport in coontail Ceratophyllum demersum. Sci. Total Environ. 2006, 357, 169–175. [Google Scholar] [CrossRef]
- Steinberg, C.E.; Kamara, S.; Prokhotskaya, V.Y.; Manusadžianas, L.; Karasyova, T.A.; Timofeyev, M.A.; Menzel, R. Dissolved humic substances–ecological driving forces from the individual to the ecosystem level? Freshw. Biol. 2006, 51, 1189–1210. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Spangenberg, M.; Steinberg, C.E.W. Dissolved Organic Matter (DOM) and effects on the aquatic macrophyte Ceratophyllum demersum in relation to photosynthesis, pigmentpattern and activity of detoxication enzymes. J. Appl. Bot. 1999, 73, 184–190. [Google Scholar]
- Grigutytė, R.; Nimptsch, J.; Manusadžianas, L.; Pflugmacher, S. Response of oxidative stress enzymes in charophyte Nitellopsis obtusa exposed to allochthonous leaf extracts from beech Fagus sylvatica. Biologija 2009, 55, 3–4. [Google Scholar] [CrossRef]
- Reid, M.S.; Bieleski, R.L. Response of Spirodela oligorrhiza to phosphorus deficiency. Plant Physiol. 1970, 46, 609–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Bucio, J.; Hernández-Abreu, E.; Sánchez-Calderón, L.; Nieto-Jacobo, M.F.; Simpson, J.; Herrera-Estrella, L. Phosphate availability alters architecture and causes changes in hormone sensitivity in the Arabidopsis root system. Plant Physiol. 2002, 129, 244–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linkohr, B.I.; Williamson, L.C.; Fitter, A.H.; Leyser, H.M.O. Nitrate and phosphate availability and distribution have different effects on root system architecture of Arabidopsis. Plant J. 2002, 29, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Roijackers, R.; Szabo, S.; Scheffer, M. Experimental analysis of the competition between algae and duckweed. Arch. Hydrobiol. 2004, 160, 401–412. [Google Scholar] [CrossRef]
- Juhel, G.; Batisse, E.; Hugues, Q.; Daly, D.; van Pelt, F.N.; O’Halloran, J.; Jansen, M.A. Alumina nanoparticles enhance growth of Lemna minor. Aquat. Toxicol. 2011, 105, 328–336. [Google Scholar] [CrossRef]
- Steinberg, C.E.W.; Meinelt, T.; Timofeyev, M.A.; Bittner, M.; Menzel, R. Humic substances (review series). Part 2: Interactions with Organisms. Environ. Sci. Pollut. Res. 2008, 15, 128–135. [Google Scholar] [CrossRef] [Green Version]
- Toming, K.; Kotta, J.; Uuemaa, E.; Sobek, S.; Kutser, T.; Tranvik, L.J. Predicting lake dissolved organic carbon at a global scale. Sci. Rep. 2020, 10, 8471. [Google Scholar] [CrossRef]
- Alles, E. Fließgewässerversauerung im Schwarzwald. In Ökologische Bewertung Auf Der Basis Benthischer Diatomeen; Oberirdische Gewässer, Gewässerökologie (Vol. 51); Landesanstalt für Umweltschutz Baden-Württemberg: Karlsruhe, Germany, 1996. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | c | h1 | h2 | h3 | h4 |
|---|---|---|---|---|---|
| mean | 0.086 | 0.086 | 0.086 | 0.085 | 0.085 |
| SD | 0.004 | 0.001 | 0.003 | 0.003 | 0.002 |
| D01 | D07 | |||||
|---|---|---|---|---|---|---|
| c | h1 | h2 | h3 | h4 | ||
| EC (μS cm−1) | 364 | 371.5 ± 15.3 | 355 ± 7.6 | 358.7 ± 9.1 | 368.7 ± 25.6 | 357.5 ± 7.0 |
| pH | 8.7 | 8.8–8.9 | 8.8–8.9 | 8.7–8.8 | 8.6–8.7 | 8.5–8.6 |
| TOC (mg dm−3) | 4.0 ± 0.1–36.5 ± 0.4 | 4.9 ± 0.1 | 6.7 ± 0.4 | 10.1 ± 0.3 | 22.9 ± 5.0 | 38.0 ± 9.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pęczuła, W.; Tarkowska-Kukuryk, M.; Niedźwiecki, M. Does Water Brownification Affect Duckweeds in Freshwaters? Results from a Laboratory Experiment. Water 2022, 14, 3826. https://doi.org/10.3390/w14233826
Pęczuła W, Tarkowska-Kukuryk M, Niedźwiecki M. Does Water Brownification Affect Duckweeds in Freshwaters? Results from a Laboratory Experiment. Water. 2022; 14(23):3826. https://doi.org/10.3390/w14233826
Chicago/Turabian StylePęczuła, Wojciech, Monika Tarkowska-Kukuryk, and Michał Niedźwiecki. 2022. "Does Water Brownification Affect Duckweeds in Freshwaters? Results from a Laboratory Experiment" Water 14, no. 23: 3826. https://doi.org/10.3390/w14233826
APA StylePęczuła, W., Tarkowska-Kukuryk, M., & Niedźwiecki, M. (2022). Does Water Brownification Affect Duckweeds in Freshwaters? Results from a Laboratory Experiment. Water, 14(23), 3826. https://doi.org/10.3390/w14233826