
Microplastic Accumulation and Degradation in Environment via Biotechnological Approaches



Abstract
:1. Introduction
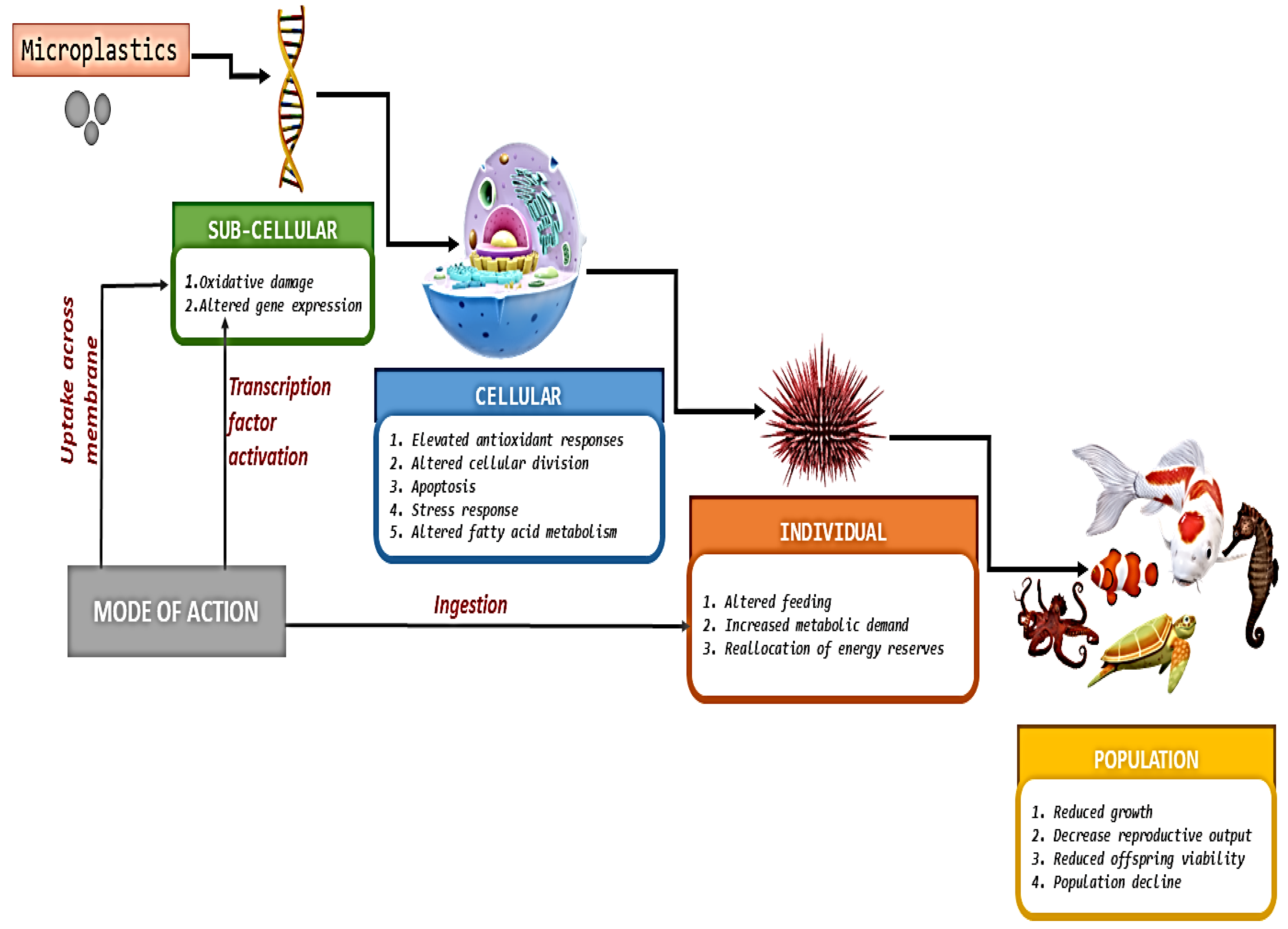
2. Effect of Microplastics on Environment
3. Health Risks Associated with the Use of Microplastics
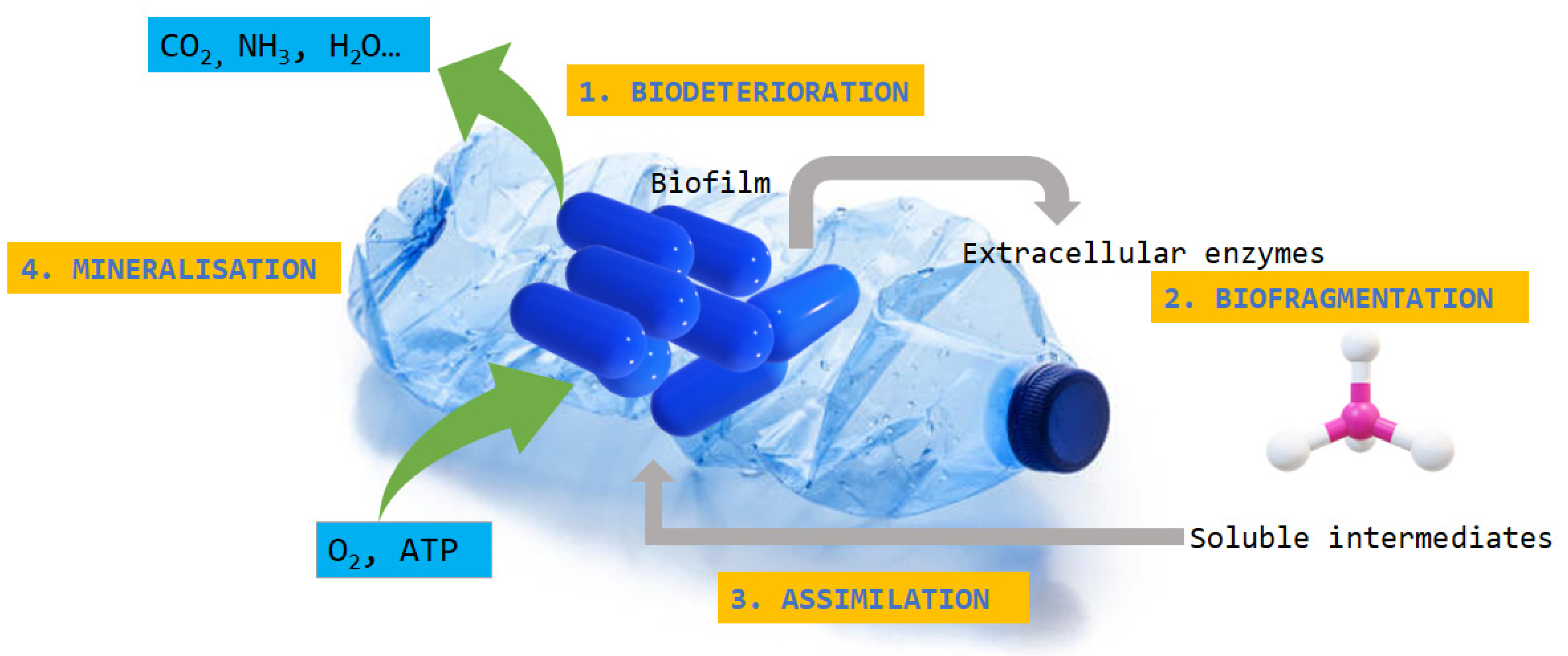
4. Role of Microorganisms in Plastic Degradation
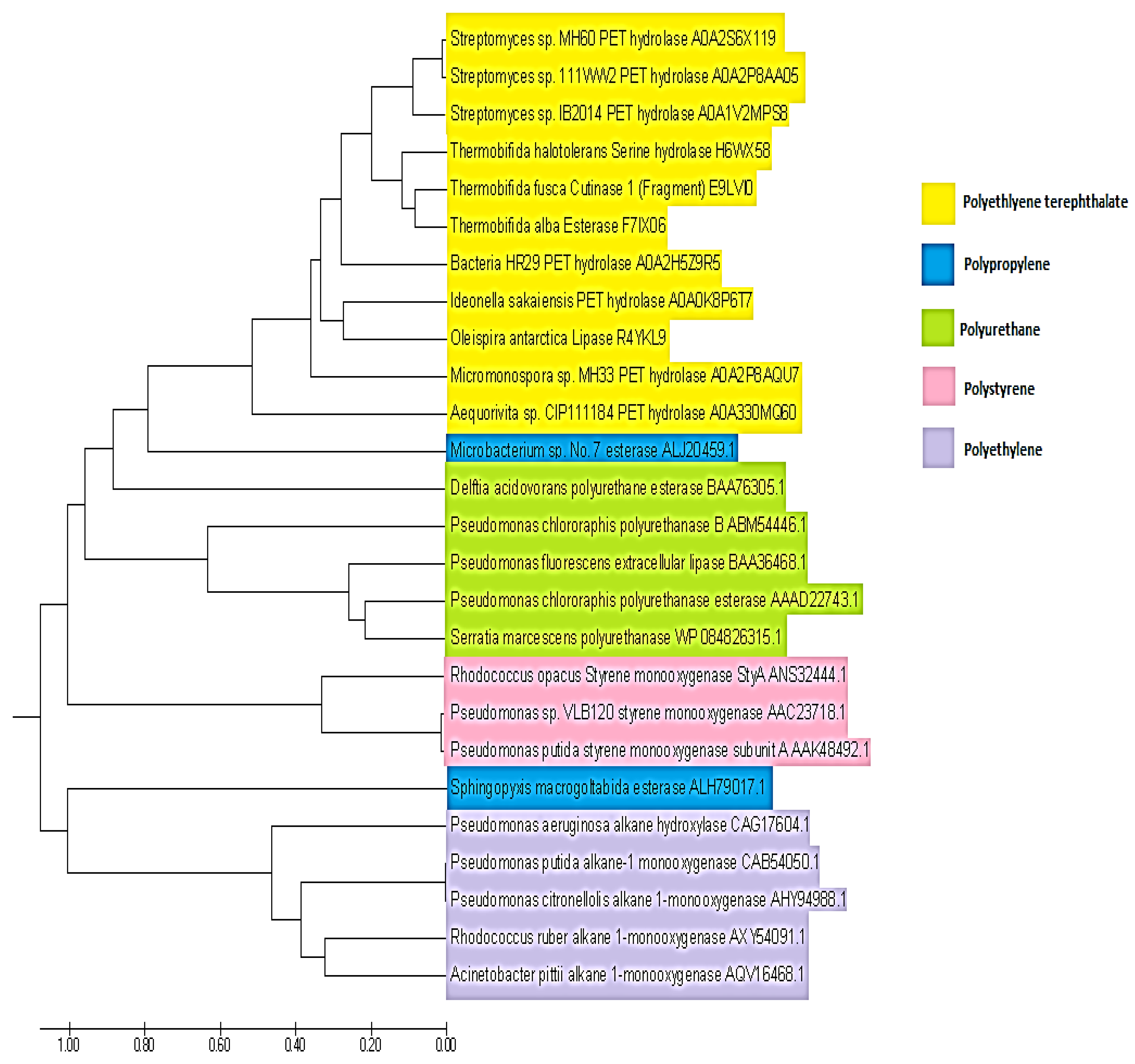
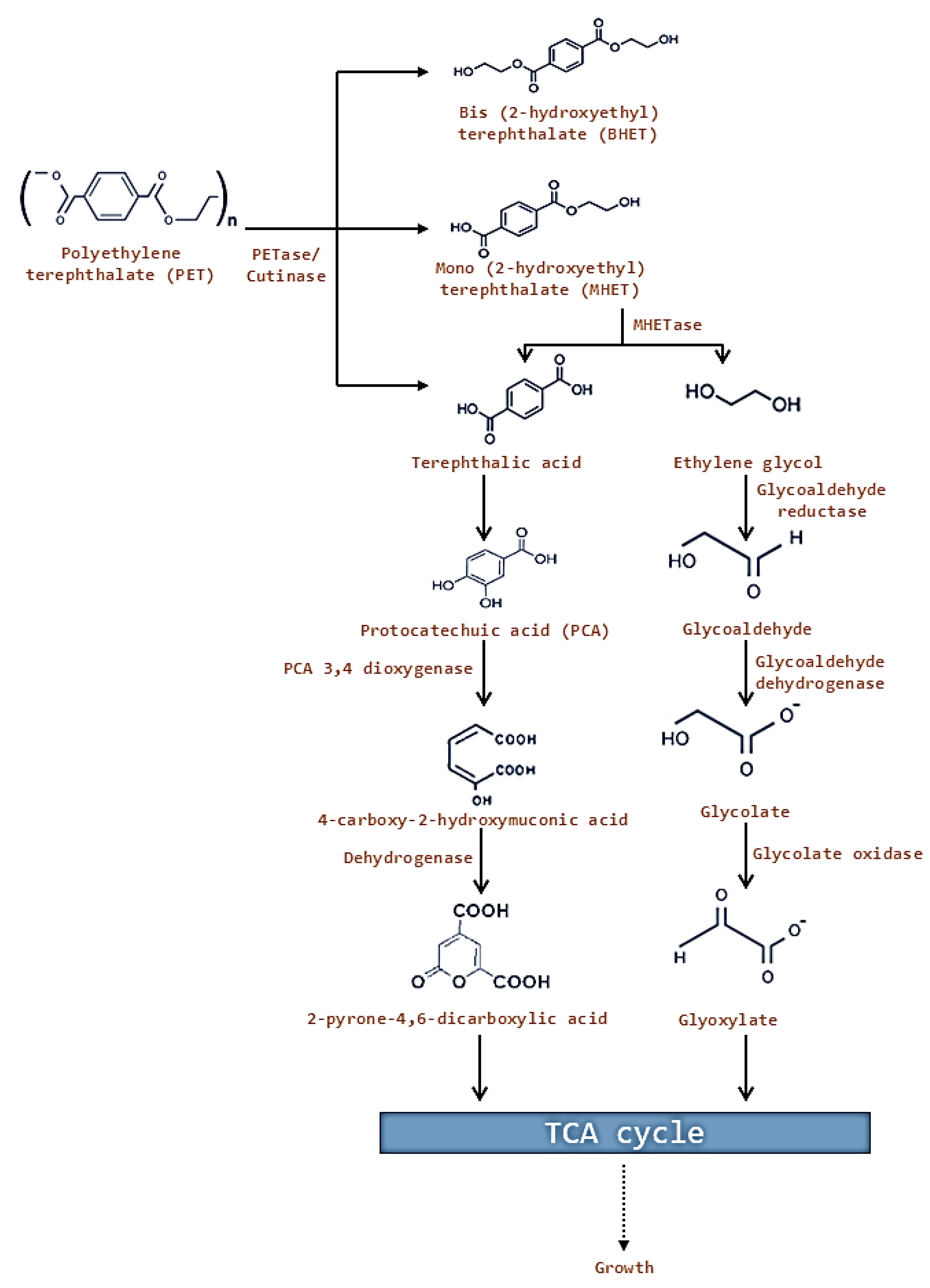
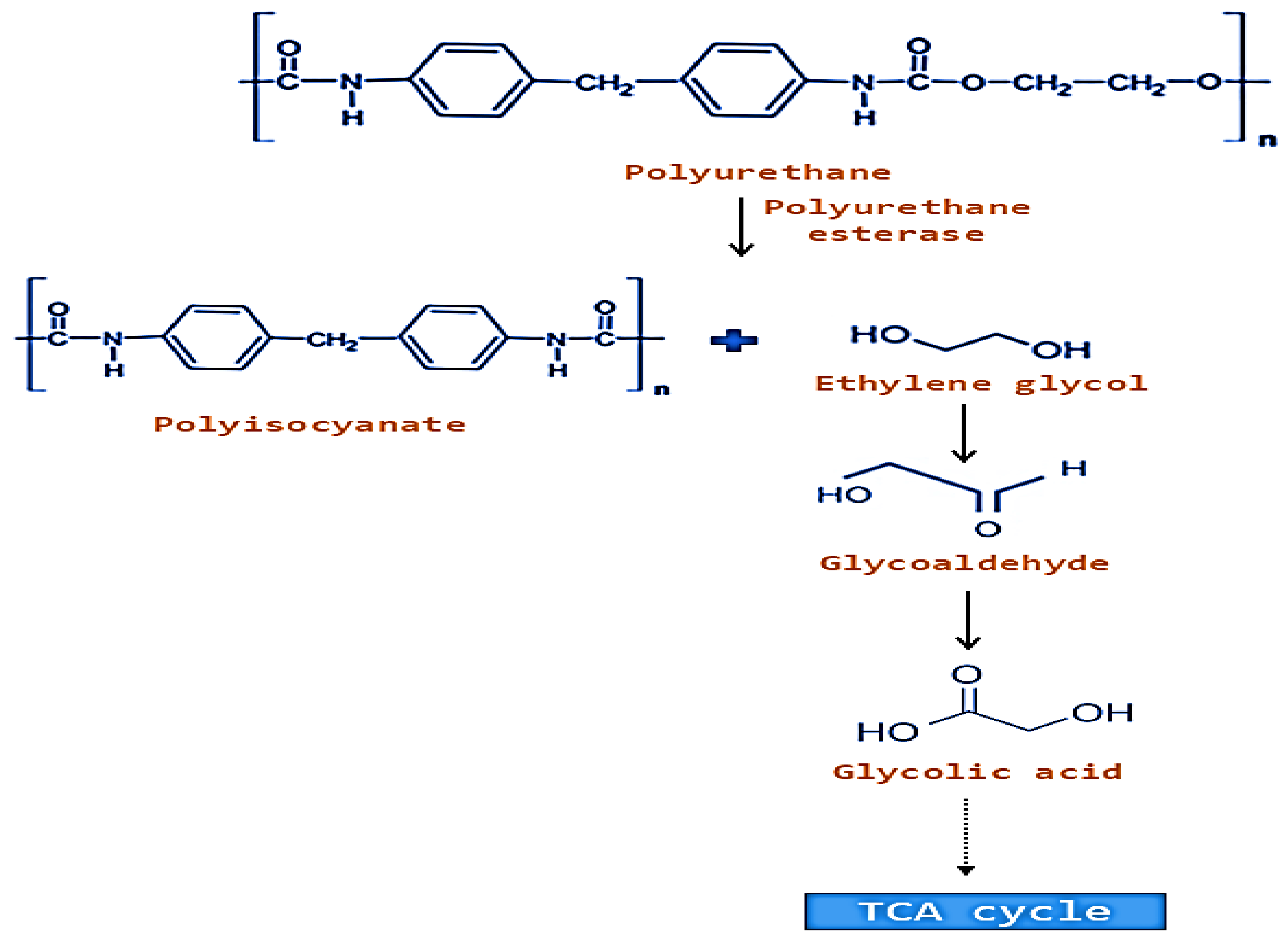
5. Enzymes Involved in Biodegradation of Plastic Polymers
6. Genetic Engineering Approaches to Enhance Plastic Biodegradation
7. Gene Editing Tools for Plastic Waste Degradation
8. Immobilized Enzymes for Bioremediation of Plastics
9. Designing Synthetic Microbial Consortium for Plastic Degradation
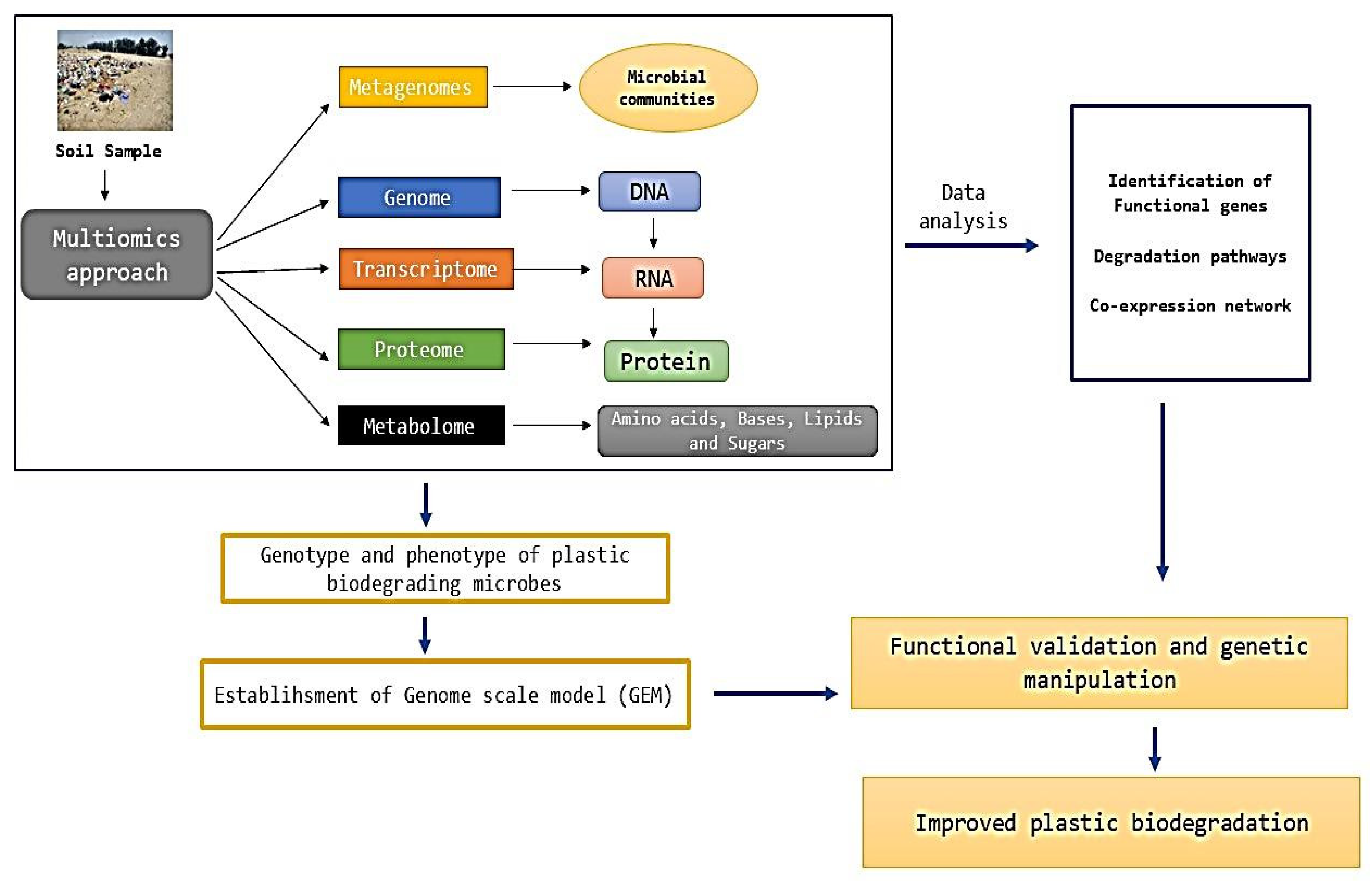
10. System Biology Approaches for Plastic Degradation
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muhamad, W.N.A.W.; Othman, R.; Shaharuddin, R.I.; Hasni, M.S.I. Microorganism as Plastic Biodegradation Agent towards Sustainable Environment. Adv. Environ. Biol. 2015, 9, 8–14. [Google Scholar]
- Yoshida, S.; Hiraga, K.; Takehana, T.; Taniguchi, I.; Yamaji, H.; Maeda, Y.; Toyohara, K.; Miyamoto, K.; Kimura, Y.; Oda, K. A Bacterium That Degrades and Assimilates Poly(Ethylene Terephthalate). Science 2016, 351, 1196–1199. [Google Scholar] [CrossRef]
- Baekeland, L.H. The Synthesis, Constitution, and Uses of Bakelite. J. Ind. Eng. Chem. 1909, 1, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Worm, B.; Lotze, H.K.; Jubinville, I.; Wilcox, C.; Jambeck, J. Plastic as a Persistent Marine Pollutant. Annu. Rev. Environ. Resour. 2017, 42, 1–26. [Google Scholar] [CrossRef]
- Chen, Z.; Wei, W.; Ni, B.-J.; Chen, H. Plastic Wastes Derived Carbon Materials for Green Energy and Sustainable Environmental Applications. Environ. Funct. Mater. 2022, 1, 34–48. [Google Scholar] [CrossRef]
- Shen, M.; Huang, W.; Chen, M.; Song, B.; Zeng, G.; Zhang, Y. (Micro)Plastic Crisis: Un-Ignorable Contribution to Global Greenhouse Gas Emissions and Climate Change. J. Clean. Prod. 2020, 254, 120138. [Google Scholar] [CrossRef]
- Andrady, A.L.; Neal, M.A. Applications and Societal Benefits of Plastics. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1977–1984. [Google Scholar] [CrossRef] [Green Version]
- Hahladakis, J.N.; Velis, C.A.; Weber, R.; Iacovidou, E.; Purnell, P. An Overview of Chemical Additives Present in Plastics: Migration, Release, Fate and Environmental Impact during Their Use, Disposal and Recycling. J. Hazard. Mater. 2018, 344, 179–199. [Google Scholar] [CrossRef]
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, Use, and Fate of All Plastics Ever Made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [Green Version]
- Vethaak, A.D.; Legler, J. Microplastics and Human Health. Science 2021, 371, 672–674. [Google Scholar] [CrossRef]
- da Costa, J.P.; Santos, P.S.M.; Duarte, A.C.; Rocha-Santos, T. (Nano)Plastics in the Environment—Sources, Fates and Effects. Sci. Total Environ. 2016, 566–567, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, M.; Lebreton, L.C.M.; Carson, H.S.; Thiel, M.; Moore, C.J.; Borerro, J.C.; Galgani, F.; Ryan, P.G.; Reisser, J. Plastic Pollution in the World’s Oceans: More than 5 Trillion Plastic Pieces Weighing over 250,000 Tons Afloat at Sea. PLoS ONE 2014, 9, e111913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galloway, T.S.; Lewis, C.N. Marine Microplastics Spell Big Problems for Future Generations. Proc. Natl. Acad. Sci. USA 2016, 113, 2331–2333. [Google Scholar] [CrossRef] [Green Version]
- Woodall, L.C.; Sanchez-Vidal, A.; Canals, M.; Paterson, G.L.J.; Coppock, R.; Sleight, V.; Calafat, A.; Rogers, A.D.; Narayanaswamy, B.E.; Thompson, R.C. The Deep Sea Is a Major Sink for Microplastic Debris. R. Soc. Open Sci. 2014, 1, 140317. [Google Scholar] [CrossRef] [Green Version]
- Rhein, F.; Nirschl, H.; Kaegi, R. Separation of Microplastic Particles from Sewage Sludge Extracts Using Magnetic Seeded Filtration. Water Res. X 2022, 17, 100155. [Google Scholar] [CrossRef]
- Cole, M.; Galloway, T.S. Ingestion of Nanoplastics and Microplastics by Pacific Oyster Larvae. Environ. Sci. Technol. 2015, 49, 14625–14632. [Google Scholar] [CrossRef] [Green Version]
- Mai, L.; Bao, L.-J.; Wong, C.S.; Zeng, E.Y. Chapter 12—Microplastics in the Terrestrial Environment. In Microplastic Contamination in Aquatic Environments; Zeng, E.Y., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 365–378. [Google Scholar] [CrossRef]
- Meijer, L.J.J.; van Emmerik, T.; van der Ent, R.; Schmidt, C.; Lebreton, L. More than 1000 Rivers Account for 80% of Global Riverine Plastic Emissions into the Ocean. Sci. Adv. 2021, 7, eaaz5803. [Google Scholar] [CrossRef]
- Steer, M.; Cole, M.; Thompson, R.C.; Lindeque, P.K. Microplastic Ingestion in Fish Larvae in the Western English Channel. Environ. Pollut. 2017, 226, 250–259. [Google Scholar] [CrossRef]
- Gregory, M.R. Environmental Implications of Plastic Debris in Marine Settings—Entanglement, Ingestion, Smothering, Hangers-on, Hitch-Hiking and Alien Invasions. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2013–2025. [Google Scholar] [CrossRef] [Green Version]
- Sussarellu, R.; Suquet, M.; Thomas, Y.; Lambert, C.; Fabioux, C.; Pernet, M.E.J.; Goïc, N.L.; Quillien, V.; Mingant, C.; Epelboin, Y.; et al. Oyster Reproduction Is Affected by Exposure to Polystyrene Microplastics. Proc. Natl. Acad. Sci. USA 2016, 113, 2430–2435. [Google Scholar] [CrossRef]
- Farrell, P.; Nelson, K. Trophic Level Transfer of Microplastic: Mytilus edulis (L.) to Carcinus maenas (L.). Environ. Pollut. 2013, 177, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Mathalon, A.; Hill, P. Microplastic Fibers in the Intertidal Ecosystem Surrounding Halifax Harbor, Nova Scotia. Mar. Pollut. Bull. 2014, 81, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, C.; Burton, H. Origins and Biological Accumulation of Small Plastic Particles in Fur Seals from Macquarie Island. Ambio 2003, 32, 380–384. [Google Scholar] [CrossRef]
- Lusher, A.L.; Hernandez-Milian, G.; O’Brien, J.; Berrow, S.; O’Connor, I.; Officer, R. Microplastic and Macroplastic Ingestion by a Deep Diving, Oceanic Cetacean: The True’s Beaked Whale Mesoplodon mirus. Environ. Pollut. 2015, 199, 185–191. [Google Scholar] [CrossRef]
- Jang, M.; Shim, W.J.; Han, G.M.; Rani, M.; Song, Y.K.; Hong, S.H. Styrofoam Debris as a Source of Hazardous Additives for Marine Organisms. Environ. Sci. Technol. 2016, 50, 4951–4960. [Google Scholar] [CrossRef] [PubMed]
- Rochman, C.M. The Complex Mixture, Fate and Toxicity of Chemicals Associated with Plastic Debris in the Marine Environment. In Marine Anthropogenic Litter; Bergmann, M., Gutow, L., Klages, M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 117–140. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Takada, H.; Yamashita, R.; Mizukawa, K.; Fukuwaka, M.; Watanuki, Y. Accumulation of Plastic-Derived Chemicals in Tissues of Seabirds Ingesting Marine Plastics. Mar. Pollut. Bull. 2013, 69, 219–222. [Google Scholar] [CrossRef]
- Hodson, M.E.; Duffus-Hodson, C.A.; Clark, A.; Prendergast-Miller, M.T.; Thorpe, K.L. Plastic Bag Derived-Microplastics as a Vector for Metal Exposure in Terrestrial Invertebrates. Environ. Sci. Technol. 2017, 51, 4714–4721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huerta Lwanga, E.; Gertsen, H.; Gooren, H.; Peters, P.; Salánki, T.; van der Ploeg, M.; Besseling, E.; Koelmans, A.A.; Geissen, V. Microplastics in the Terrestrial Ecosystem: Implications for Lumbricus Terrestris (Oligochaeta, Lumbricidae). Environ. Sci. Technol. 2016, 50, 2685–2691. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.P.; Schratzberger, M.; Sapp, M.; Osborn, A.M. Rapid Bacterial Colonization of Low-Density Polyethylene Microplastics in Coastal Sediment Microcosms. BMC Microbiol. 2014, 14, 232. [Google Scholar] [CrossRef]
- Zhao, S.; Zhu, L.; Li, D. Microscopic Anthropogenic Litter in Terrestrial Birds from Shanghai, China: Not Only Plastics but Also Natural Fibers. Sci. Total Environ. 2016, 550, 1110–1115. [Google Scholar] [CrossRef]
- Maaß, S.; Daphi, D.; Lehmann, A.; Rillig, M.C. Transport of Microplastics by Two Collembolan Species. Environ. Pollut. 2017, 225, 456–459. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, X.; Wei, W.; Chen, H.; Ni, B.-J. Removal of Microplastics and Nanoplastics from Urban Waters: Separation and Degradation. Water Res. 2022, 221, 118820. [Google Scholar] [CrossRef] [PubMed]
- Carr, S.A.; Liu, J.; Tesoro, A.G. Transport and Fate of Microplastic Particles in Wastewater Treatment Plants. Water Res. 2016, 91, 174–182. [Google Scholar] [CrossRef]
- Fendall, L.S.; Sewell, M.A. Contributing to Marine Pollution by Washing Your Face: Microplastics in Facial Cleansers. Mar. Pollut. Bull. 2009, 58, 1225–1228. [Google Scholar] [CrossRef] [PubMed]
- Nizzetto, L.; Langaas, S.; Futter, M. Pollution: Do Microplastics Spill on to Farm Soils? Nature 2016, 537, 488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rillig, M.C. Microplastic in Terrestrial Ecosystems and the Soil? Environ. Sci. Technol. 2012, 46, 6453–6454. [Google Scholar] [CrossRef]
- Conley, K.; Clum, A.; Deepe, J.; Lane, H.; Beckingham, B. Wastewater Treatment Plants as a Source of Microplastics to an Urban Estuary: Removal Efficiencies and Loading per Capita over One Year. Water Res. X 2019, 3, 100030. [Google Scholar] [CrossRef]
- Elkhatib, D.; Oyanedel-Craver, V. A Critical Review of Extraction and Identification Methods of Microplastics in Wastewater and Drinking Water. Environ. Sci. Technol. 2020, 54, 7037–7049. [Google Scholar] [CrossRef]
- Na, S.-H.; Kim, M.-J.; Kim, J.-T.; Jeong, S.; Lee, S.; Chung, J.; Kim, E.-J. Microplastic Removal in Conventional Drinking Water Treatment Processes: Performance, Mechanism, and Potential Risk. Water Res. 2021, 202, 117417. [Google Scholar] [CrossRef]
- Lithner, D.; Larsson, Å.; Dave, G. Environmental and Health Hazard Ranking and Assessment of Plastic Polymers Based on Chemical Composition. Sci. Total Environ. 2011, 409, 3309–3324. [Google Scholar] [CrossRef]
- Koch, H.M.; Calafat, A.M. Human Body Burdens of Chemicals Used in Plastic Manufacture. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2063–2078. [Google Scholar] [CrossRef] [PubMed]
- Van Cauwenberghe, L.; Janssen, C.R. Microplastics in Bivalves Cultured for Human Consumption. Environ. Pollut. 2014, 193, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Leslie, H.A.; van Velzen, M.J.M.; Brandsma, S.H.; Vethaak, A.D.; Garcia-Vallejo, J.J.; Lamoree, M.H. Discovery and Quantification of Plastic Particle Pollution in Human Blood. Environ. Int. 2022, 163, 107199. [Google Scholar] [CrossRef]
- Yan, Z.; Liu, Y.; Zhang, T.; Zhang, F.; Ren, H.; Zhang, Y. Analysis of Microplastics in Human Feces Reveals a Correlation between Fecal Microplastics and Inflammatory Bowel Disease Status. Environ. Sci. Technol. 2022, 56, 414–421. [Google Scholar] [CrossRef]
- Deng, Y.; Zhang, Y.; Lemos, B.; Ren, H. Tissue Accumulation of Microplastics in Mice and Biomarker Responses Suggest Widespread Health Risks of Exposure. Sci. Rep. 2017, 7, 46687. [Google Scholar] [CrossRef] [Green Version]
- John, R.C.; Essien, J.P.; Akpan, S.B.; Okpokwasili, G.C. Polycyclic Aromatic Hydrocarbon-Degrading Bacteria from Aviation Fuel Spill Site at Ibeno, Nigeria. Bull. Environ. Contam. Toxicol. 2012, 88, 1014–1019. [Google Scholar] [CrossRef] [Green Version]
- Kyaw, B.M.; Champakalakshmi, R.; Sakharkar, M.K.; Lim, C.S.; Sakharkar, K.R. Biodegradation of Low Density Polythene (LDPE) by Pseudomonas Species. Indian J. Microbiol. 2012, 52, 411–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanda, S.; Sahu, S.S.; Abraham, J. Studies on the Biodegradation of Natural and Synthetic Polyethylene by Pseudomonas Spp. J. Appl. Sci. Environ. Manag. 2010, 14. [Google Scholar] [CrossRef] [Green Version]
- Tribedi, P.; Sarkar, S.; Mukherjee, K.; Sil, A.K. Isolation of a Novel Pseudomonas Sp from Soil That Can Efficiently Degrade Polyethylene Succinate. Environ. Sci. Pollut. Res. 2012, 19, 2115–2124. [Google Scholar] [CrossRef]
- Usha, R.; Sangeetha, T.; Palaniswamy, M. Screening of polyethylene degrading microorganisms from garbage soi. Libyan Agric. Res. Cent. J. Int. 2011, 2, 200–204. [Google Scholar]
- Orr, I.G.; Hadar, Y.; Sivan, A. Colonization, Biofilm Formation and Biodegradation of Polyethylene by a Strain of Rhodococcus Ruber. Appl. Microbiol. Biotechnol. 2004, 65, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.A.; Hasan, F.; Hameed, A.; Ahmed, S. Biological Degradation of Plastics: A Comprehensive Review. Biotechnol. Adv. 2008, 26, 246–265. [Google Scholar] [CrossRef] [PubMed]
- Ojha, N.; Pradhan, N.; Singh, S.; Barla, A.; Shrivastava, A.; Khatua, P.; Rai, V.; Bose, S. Evaluation of HDPE and LDPE Degradation by Fungus, Implemented by Statistical Optimization. Sci. Rep. 2017, 7, 39515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raaman, N.; Rajitha, N.; Jayshree, A.; Jegadeesh, R. Biodegradation of Plastic by Aspergillus Spp. Isolated from Polythene Polluted Sites around Chennai. J. Acad. Ind. Res. 2012, 1, 4. [Google Scholar]
- Mohan, S.K.; Srivastava, T. Microbial Deterioration and Degradation of Polymeric Materials. J. Biochem. Technol. 2010, 2, 210–215. [Google Scholar]
- Siracusa, V.; Rocculi, P.; Romani, S.; Rosa, M.D. Biodegradable Polymers for Food Packaging: A Review. Trends Food Sci. Technol. 2008, 19, 634–643. [Google Scholar] [CrossRef]
- Slor, G.; Papo, N.; Hananel, U.; Amir, R.J. Tuning the Molecular Weight of Polymeric Amphiphiles as a Tool to Access Micelles with a Wide Range of Enzymatic Degradation Rates. Chem. Commun. 2018, 54, 6875–6878. [Google Scholar] [CrossRef]
- Syranidou, E.; Karkanorachaki, K.; Amorotti, F.; Franchini, M.; Repouskou, E.; Kaliva, M.; Vamvakaki, M.; Kolvenbach, B.; Fava, F.; Corvini, P.F.-X.; et al. Biodegradation of Weathered Polystyrene Films in Seawater Microcosms. Sci. Rep. 2017, 7, 17991. [Google Scholar] [CrossRef] [Green Version]
- Brodhagen, M.; Peyron, M.; Miles, C.; Inglis, D.A. Biodegradable Plastic Agricultural Mulches and Key Features of Microbial Degradation. Appl. Microbiol. Biotechnol. 2015, 99, 1039–1056. [Google Scholar] [CrossRef]
- Mohan, A.J.; Sekhar, V.C.; Bhaskar, T.; Nampoothiri, K.M. Microbial Assisted High Impact Polystyrene (HIPS) Degradation. Bioresour. Technol. 2016, 213, 204–207. [Google Scholar] [CrossRef]
- Novotný, Č.; Erbanová, P.; Sezimová, H.; Malachová, K.; Rybková, Z.; Malinová, L.; Prokopová, I.; Brožek, J. Biodegradation of Aromatic-Aliphatic Copolyesters and Polyesteramides by Esterase Activity-Producing Microorganisms. Int. Biodeterior. Biodegrad. 2015, 97, 25–30. [Google Scholar] [CrossRef]
- Ruiz, C.; Main, T.; Hilliard, N.P.; Howard, G.T. Purification and Characterization of Twopolyurethanase Enzymes from Pseudomonas Chlororaphis. Int. Biodeterior. Biodegrad. 1999, 43, 43–47. [Google Scholar] [CrossRef]
- Sangale, M.K. A Review on Biodegradation of Polythene: The Microbial Approach. J. Bioremediation Biodegrad. 2012, 3, 1–9. [Google Scholar] [CrossRef]
- Müller, R.-J.; Schrader, H.; Profe, J.; Dresler, K.; Deckwer, W.-D. Enzymatic Degradation of Poly(Ethylene Terephthalate): Rapid Hydrolyse Using a Hydrolase from T. Fusca. Macromol. Rapid Commun. 2005, 26, 1400–1405. [Google Scholar] [CrossRef]
- Sulaiman, S.; Yamato, S.; Kanaya, E.; Kim, J.-J.; Koga, Y.; Takano, K.; Kanaya, S. Isolation of a Novel Cutinase Homolog with Polyethylene Terephthalate-Degrading Activity from Leaf-Branch Compost by Using a Metagenomic Approach. Appl. Environ. Microbiol. 2012, 78, 1556–1562. [Google Scholar] [CrossRef] [Green Version]
- Li, W.-J.; Jayakody, L.N.; Franden, M.A.; Wehrmann, M.; Daun, T.; Hauer, B.; Blank, L.M.; Beckham, G.T.; Klebensberger, J.; Wierckx, N. Laboratory Evolution Reveals the Metabolic and Regulatory Basis of Ethylene Glycol Metabolism by Pseudomonas Putida KT2440. Environ. Microbiol. 2019, 21, 3669–3682. [Google Scholar] [CrossRef] [PubMed]
- Santo, M.; Weitsman, R.; Sivan, A. The Role of the Copper-Binding Enzyme—Laccase—In the Biodegradation of Polyethylene by the Actinomycete Rhodococcus Ruber. Int. Biodeterior. Biodegrad. 2013, 84, 204–210. [Google Scholar] [CrossRef]
- Jeon, H.J.; Kim, M.N. Comparison of the Functional Characterization between Alkane Monooxygenases for Low-Molecular-Weight Polyethylene Biodegradation. Int. Biodeterior. Biodegrad. 2016, 114, 202–208. [Google Scholar] [CrossRef]
- Nakajima-Kambe, T.; Onuma, F.; Kimpara, N.; Nakahara, T. Isolation and Characterization of a Bacterium Which Utilizes Polyester Polyurethane as a Sole Carbon and Nitrogen Source. FEMS Microbiol. Lett. 1995, 129, 39–42. [Google Scholar] [CrossRef]
- Schmidt, J.; Wei, R.; Oeser, T.; Dedavid e Silva, L.A.; Breite, D.; Schulze, A.; Zimmermann, W. Degradation of Polyester Polyurethane by Bacterial Polyester Hydrolases. Polymers 2017, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Ho, B.T.; Roberts, T.K.; Lucas, S. An Overview on Biodegradation of Polystyrene and Modified Polystyrene: The Microbial Approach. Crit. Rev. Biotechnol. 2018, 38, 308–320. [Google Scholar] [CrossRef] [PubMed]
- Hussain, I.; Aleti, G.; Naidu, R.; Puschenreiter, M.; Mahmood, Q.; Rahman, M.M.; Wang, F.; Shaheen, S.; Syed, J.H.; Reichenauer, T.G. Microbe and Plant Assisted-Remediation of Organic Xenobiotics and Its Enhancement by Genetically Modified Organisms and Recombinant Technology: A Review. Sci. Total Environ. 2018, 628–629, 1582–1599. [Google Scholar] [CrossRef] [PubMed]
- AW, T. Application of Recombinant DNA Technology (Genetically Modified Organisms) to the Advancement of Agriculture, Medicine, Bioremediation and Biotechnology Industries. J. Appl. Biotechnol. Bioeng. 2016, 1, 1–4. [Google Scholar] [CrossRef]
- Kumar, N.M.; Muthukumaran, C.; Sharmila, G.; Gurunathan, B. Genetically Modified Organisms and Its Impact on the Enhancement of Bioremediation. In Bioremediation: Applications for Environmental Protection and Management; Energy, Environment, and Sustainability; Varjani, S.J., Agarwal, A.K., Gnansounou, E., Gurunathan, B., Eds.; Springer: Singapore, 2018; pp. 53–76. [Google Scholar] [CrossRef]
- Shirke, A.N.; White, C.; Englaender, J.A.; Zwarycz, A.; Butterfoss, G.L.; Linhardt, R.J.; Gross, R.A. Stabilizing Leaf and Branch Compost Cutinase (LCC) with Glycosylation: Mechanism and Effect on PET Hydrolysis. Biochemistry 2018, 57, 1190–1200. [Google Scholar] [CrossRef] [PubMed]
- Oda, M.; Yamagami, Y.; Inaba, S.; Oida, T.; Yamamoto, M.; Kitajima, S.; Kawai, F. Enzymatic Hydrolysis of PET: Functional Roles of Three Ca2+ Ions Bound to a Cutinase-like Enzyme, Cut190*, and Its Engineering for Improved Activity. Appl. Microbiol. Biotechnol. 2018, 102, 10067–10077. [Google Scholar] [CrossRef]
- Austin, H.P.; Allen, M.D.; Donohoe, B.S.; Rorrer, N.A.; Kearns, F.L.; Silveira, R.L.; Pollard, B.C.; Dominick, G.; Duman, R.; El Omari, K.; et al. Characterization and Engineering of a Plastic-Degrading Aromatic Polyesterase. Proc. Natl. Acad. Sci. USA 2018, 115, E4350–E4357. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Wei, R.; Cui, Q.; Bornscheuer, U.T.; Liu, Y.-J. Thermophilic Whole-Cell Degradation of Polyethylene Terephthalate Using Engineered Clostridium Thermocellum. Microb. Biotechnol. 2021, 14, 374–385. [Google Scholar] [CrossRef]
- Gyung Yoon, M.; Jeong Jeon, H.; Nam Kim, M. Biodegradation of Polyethylene by a Soil Bacterium and AlkB Cloned Recombinant Cell. J. Bioremediation Biodegrad. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Sadler, J.C.; Wallace, S. Microbial Synthesis of Vanillin from Waste Poly(Ethylene Terephthalate). Green Chem. 2021, 23, 4665–4672. [Google Scholar] [CrossRef]
- Perpetuo, E.A.; Souza, C.B.; Nascimento, C.A.O. Engineering Bacteria for Bioremediation; IntechOpen: London, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- Arazoe, T.; Kondo, A.; Nishida, K. Targeted Nucleotide Editing Technologies for Microbial Metabolic Engineering. Biotechnol. J. 2018, 13, e1700596. [Google Scholar] [CrossRef]
- Yadav, R.; Kumar, V.; Baweja, M.; Shukla, P. Gene Editing and Genetic Engineering Approaches for Advanced Probiotics: A Review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1735–1746. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.; Shukla, P. Alternative Strategies for Microbial Remediation of Pollutants via Synthetic Biology. Front. Microbiol. 2020, 11, 808. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome Engineering Using the CRISPR-Cas9 System. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, B.; Habibi, M.; Esvand, M. Biodegradation of Naphthalene Using Pseudomonas Aeruginosa by up Flow Anoxic–Aerobic Continuous Flow Combined Bioreactor. J. Environ. Health Sci. Eng. 2015, 13, 26. [Google Scholar] [CrossRef] [Green Version]
- Nogales, J.; Mueller, J.; Gudmundsson, S.; Canalejo, F.J.; Duque, E.; Monk, J.; Feist, A.M.; Ramos, J.L.; Niu, W.; Palsson, B.O. High-Quality Genome-Scale Metabolic Modelling of Pseudomonas Putida Highlights Its Broad Metabolic Capabilities. Environ. Microbiol. 2020, 22, 255–269. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Zhang, Y.; Zhang, Y.; Pi, Y.; Gu, T.; Song, L.; Wang, Y.; Ji, Q. CRISPR/Cas9-Based Genome Editing in Pseudomonas Aeruginosa and Cytidine Deaminase-Mediated Base Editing in Pseudomonas Species. iScience 2018, 6, 222–231. [Google Scholar] [CrossRef] [Green Version]
- Marshall, R.; Maxwell, C.S.; Collins, S.P.; Jacobsen, T.; Luo, M.L.; Begemann, M.B.; Gray, B.N.; January, E.; Singer, A.; He, Y.; et al. Rapid and Scalable Characterization of CRISPR Technologies Using an E. Coli Cell-Free Transcription-Translation System. Mol. Cell 2018, 69, 146–157.e3. [Google Scholar] [CrossRef] [Green Version]
- Dash, H.R.; Mangwani, N.; Chakraborty, J.; Kumari, S.; Das, S. Marine Bacteria: Potential Candidates for Enhanced Bioremediation. Appl. Microbiol. Biotechnol. 2013, 97, 561–571. [Google Scholar] [CrossRef]
- Khan, F.; Sajid, M.; Cameotra, S. In Silico Approach for the Bioremediation of Toxic Pollutants. J. Pet. Environ. Biotechnol. 2013, 4, 2157–7463. [Google Scholar] [CrossRef] [Green Version]
- Quecholac-Piña, X.; García-Rivera, M.A.; Espinosa-Valdemar, R.M.; Vázquez-Morillas, A.; Beltrán-Villavicencio, M.; Cisneros-Ramos, A.D.L.L. Biodegradation of Compostable and Oxodegradable Plastic Films by Backyard Composting and Bioaugmentation. Environ. Sci. Pollut. Res. 2017, 24, 25725–25730. [Google Scholar] [CrossRef]
- Rayu, S.; Karpouzas, D.G.; Singh, B.K. Emerging Technologies in Bioremediation: Constraints and Opportunities. Biodegradation 2012, 23, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Christena, L.R.; Rajaram, Y.R.S. Enzyme Immobilization: An Overview on Techniques and Support Materials. 3 Biotech 2013, 3, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, M.; Honak, A.; Oeser, T.; Wei, R.; Belisário-Ferrari, M.R.; Then, J.; Schmidt, J.; Zimmermann, W. A Dual Enzyme System Composed of a Polyester Hydrolase and a Carboxylesterase Enhances the Biocatalytic Degradation of Polyethylene Terephthalate Films. Biotechnol. J. 2016, 11, 1082–1087. [Google Scholar] [CrossRef]
- Anwar, M.S.; Kapri, A.; Chaudhry, V.; Mishra, A.; Ansari, M.W.; Souche, Y.; Nautiyal, C.S.; Zaidi, M.G.H.; Goel, R. Response of Indigenously Developed Bacterial Consortia in Progressive Degradation of Polyvinyl Chloride. Protoplasma 2016, 253, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Skariyachan, S.; Manjunatha, V.; Sultana, S.; Jois, C.; Bai, V.; Vasist, K.S. Novel Bacterial Consortia Isolated from Plastic Garbage Processing Areas Demonstrated Enhanced Degradation for Low Density Polyethylene. Environ. Sci. Pollut. Res. Int. 2016, 23, 18307–18319. [Google Scholar] [CrossRef]
- García-Jiménez, B.; García, J.L.; Nogales, J. FLYCOP: Metabolic Modeling-Based Analysis and Engineering Microbial Communities. Bioinformatics 2018, 34, i954–i963. [Google Scholar] [CrossRef]
- Wanapaisan, P.; Laothamteep, N.; Vejarano, F.; Chakraborty, J.; Shintani, M.; Muangchinda, C.; Morita, T.; Suzuki-Minakuchi, C.; Inoue, K.; Nojiri, H.; et al. Synergistic Degradation of Pyrene by Five Culturable Bacteria in a Mangrove Sediment-Derived Bacterial Consortium. J. Hazard. Mater. 2018, 342, 561–570. [Google Scholar] [CrossRef]
- Polti, M.A.; Aparicio, J.D.; Benimeli, C.S.; Amoroso, M.J. Simultaneous Bioremediation of Cr(VI) and Lindane in Soil by Actinobacteria. Int. Biodeterior. Amp Biodegrad. 2014, 88, 48–55. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, B.; Jiang, Z.; Dong, X.; Hu, M.; Wang, Z. Metabolic Ability and Individual Characteristics of an Atrazine-Degrading Consortium DNC5. J. Hazard. Mater. 2012, 237–238, 376–381. [Google Scholar] [CrossRef]
- Johns, N.I.; Blazejewski, T.; Gomes, A.L.C.; Wang, H.H. Principles for Designing Synthetic Microbial Communities. Curr. Opin. Microbiol. 2016, 31, 146–153. [Google Scholar] [CrossRef]
- Carbajosa, G.; Trigo, A.; Valencia, A.; Cases, I. Bionemo: Molecular Information on Biodegradation Metabolism. Nucleic Acids Res. 2009, 37, D598–D602. [Google Scholar] [CrossRef]
- Gao, J.; Ellis, L.B.M.; Wackett, L.P. The University of Minnesota Biocatalysis/Biodegradation Database: Improving Public Access. Nucleic Acids Res. 2010, 38, D488–D491. [Google Scholar] [CrossRef] [Green Version]
- Caspi, R.; Billington, R.; Ferrer, L.; Foerster, H.; Fulcher, C.A.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Latendresse, M.; Mueller, L.A.; et al. The MetaCyc Database of Metabolic Pathways and Enzymes and the BioCyc Collection of Pathway/Genome Databases. Nucleic Acids Res. 2016, 44, D471–D480. [Google Scholar] [CrossRef] [PubMed]
- Caspi, R.; Altman, T.; Billington, R.; Dreher, K.; Foerster, H.; Fulcher, C.A.; Holland, T.A.; Keseler, I.M.; Kothari, A.; Kubo, A.; et al. The MetaCyc Database of Metabolic Pathways and Enzymes and the BioCyc Collection of Pathway/Genome Databases. Nucleic Acids Res. 2014, 42, D459–D471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muto, A.; Kotera, M.; Tokimatsu, T.; Nakagawa, Z.; Goto, S.; Kanehisa, M. Modular Architecture of Metabolic Pathways Revealed by Conserved Sequences of Reactions. J. Chem. Inf. Model. 2013, 53, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.; Yamada, T.; Hattori, M.; Goto, S.; Kanehisa, M. Systematic Analysis of Enzyme-Catalyzed Reaction Patterns and Prediction of Microbial Biodegradation Pathways. J. Chem. Inf. Model. 2007, 47, 1702–1712. [Google Scholar] [CrossRef]
- Finley, S.D.; Broadbelt, L.J.; Hatzimanikatis, V. Computational Framework for Predictive Biodegradation. Biotechnol. Bioeng. 2009, 104, 1086–1097. [Google Scholar] [CrossRef] [Green Version]
- Chou, C.-H.; Chang, W.-C.; Chiu, C.-M.; Huang, C.-C.; Huang, H.-D. FMM: A Web Server for Metabolic Pathway Reconstruction and Comparative Analysis. Nucleic Acids Res. 2009, 37, W129–W134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClymont, K.; Soyer, O.S. Metabolic Tinker: An Online Tool for Guiding the Design of Synthetic Metabolic Pathways. Nucleic Acids Res. 2013, 41, e113. [Google Scholar] [CrossRef] [Green Version]
- Pazos, F.; Guijas, D.; Valencia, A.; Lorenzo, V.D. MetaRouter: Bioinformatics for Bioremediation. Nucleic Acids Res. 2005, 33, D588. [Google Scholar] [CrossRef]
- Borja, A. Testing the Efficiency of a Bacterial Community-Based Index (MicrogAMBI) to Assess Distinct Impact Sources in Six Locations around the World. Ecol. Indic. 2018, 85, 594–602. [Google Scholar] [CrossRef]
- Kong, W.; Meldgin, D.R.; Collins, J.J.; Lu, T. Designing Microbial Consortia with Defined Social Interactions. Nat. Chem. Biol. 2018, 14, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Arora, P.K.; Bae, H. Integration of Bioinformatics to Biodegradation. Biol. Proced. Online 2014, 16, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulekar, M.; Sharma, J. Bioinformatics Applied in Bioremediation. Innov. Rom. Food Biotechnol. 2008, 3, 28–36. [Google Scholar]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra Precision Glide: Docking and Scoring Incorporating a Model of Hydrophobic Enclosure for Protein-Ligand Complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [Green Version]
- Grosdidier, A.; Zoete, V.; Michielin, O. SwissDock, a Protein-Small Molecule Docking Web Service Based on EADock DSS. Nucleic Acids Res. 2011, 39, W270–W277. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Skariyachan, S.; Megha, M.; Kini, M.N.; Mukund, K.M.; Rizvi, A.; Vasist, K. Selection and Screening of Microbial Consortia for Efficient and Ecofriendly Degradation of Plastic Garbage Collected from Urban and Rural Areas of Bangalore, India. Environ. Monit. Assess. 2015, 187, 4174. [Google Scholar] [CrossRef]
- Lindorff-Larsen, K.; Piana, S.; Palmo, K.; Maragakis, P.; Klepeis, J.L.; Dror, R.O.; Shaw, D.E. Improved Side-Chain Torsion Potentials for the Amber Ff99SB Protein Force Field. Proteins 2010, 78, 1950–1958. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Cai, L.; Yang, Y.; Ju, F.; Li, X.; Yu, Y.; Zhang, T. Metagenomic Analysis Reveals Potential Biodegradation Pathways of Persistent Pesticides in Freshwater and Marine Sediments. Sci. Total Environ. 2014, 470–471, 983–992. [Google Scholar] [CrossRef]
- Kumari, A.; Bano, N.; Bag, S.K.; Chaudhary, D.R.; Jha, B. Transcriptome-Guided Insights Into Plastic Degradation by the Marine Bacterium. Front. Microbiol. 2021, 12, 2761. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Liu, R.; Zuo, Z.; Che, Y.; Yu, H.; Song, C.; Yang, C. Metabolic Engineering of Pseudomonas Putida KT2440 for Complete Mineralization of Methyl Parathion and γ-Hexachlorocyclohexane. ACS Synth. Biol. 2016, 5, 434–442. [Google Scholar] [CrossRef]
- Liu, S.Y.; Leung, M.M.-L.; Fang, J.K.-H.; Chua, S.L. Engineering a Microbial ‘Trap and Release’ Mechanism for Microplastics Removal. Chem. Eng. J. 2021, 404, 127079. [Google Scholar] [CrossRef]
- Feng, L.-A.; Liang, B.; Zeng, X.; Shi, C.; Yin, H.; Feng, Y.; Chen, Y.; Yu, Q. Engineered Bacterium-Binding Protein Promotes Root Recruitment of Functional Bacteria for Enhanced Cadmium Removal from Wastewater by Phytoremediation. Water Res. 2022, 221, 118746. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Mismanaged Waste Emitted to the Ocean (Metric tons Year−1) | Mismanaged Plastic Waste to Ocean per Capita (kg per Year) |
|---|---|---|
| Philippines | 356,371 | 3.296 |
| India | 126,513 | 0.093 |
| Malaysia | 73,098 | 2.288 |
| China | 70,707 | 0.049 |
| Indonesia | 56,333 | 0.208 |
| Brazil | 37,799 | 0.179 |
| Vietnam | 28,221 | 0.293 |
| Bangladesh | 24,640 | 0.151 |
| Thailand | 22,806 | 0.328 |
| Nigeria | 18,640 | 0.093 |
| Sr. No. | Database | Link | Importance in Plastic Degradation | Reference |
|---|---|---|---|---|
| 1 | Biodegradation Network-Molecular Biology Database (Bionemo) | https://bionemo.bioinfo.cnio.es | Manually curated database providing information about genes and proteins involved in biodegradation metabolism | [105] |
| 2 | University of Minnesota Biocatalysis/Biodegradation Database (UM-BBD) | http://eawag-bbd.ethz.ch/ | Provides information pertaining to metabolic pathways along with microbial biocatalytic reactions, enzymes, and genes in the process of biodegradation | [106] |
| 3 | Metacyc | https://metacyc.org/ | Highly curated database providing information about experimentally elucidated metabolic pathways and enzymes for all domains of life | [107] |
| 4 | Biocyc | https://biocyc.org/ | It gives information pertaining to entire genomes and has predicted a metabolic network of an organism | [108] |
| 5 | PathPred | http://www.genome.jp/tools/pathpred/ | Retrieves information from KEGG reaction and KEGG RPAIR databases of query compound and predicts possible enzyme-catalyzed reaction pathways | [109,110] |
| 6 | Biochemical Network Integrated Computational Explorer (BNICE) | http://minedatabase.mcs.anl.gov | Using the reaction rules based on the Enzyme Commission classification system, it designs novel chemical structures and pathways | [111] |
| 7 | From Metabolite to Metabolite | http://FMM.mbc.nctu.edu.tw/ | Proposes metabolic pathways including the information about enzymes involved and genes and organisms which can be compared between different species and is based on KEGG databases and other integrated databases | [112] |
| 8 | Metabolic Tinker | http://osslab.ex.ac.uk/tinker.aspx | Used to design metabolic pathways between source compounds and end-products synthetically | [113] |
| 9 | MetaRouter | http://pdg.cnb.uam.es/biodeg_net/MetaRouter | Maintains diverse information related to bioremediation and biodegradation pathways stored in an integrated framework | [114] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thakur, S.; Mathur, S.; Patel, S.; Paital, B. Microplastic Accumulation and Degradation in Environment via Biotechnological Approaches. Water 2022, 14, 4053. https://doi.org/10.3390/w14244053
Thakur S, Mathur S, Patel S, Paital B. Microplastic Accumulation and Degradation in Environment via Biotechnological Approaches. Water. 2022; 14(24):4053. https://doi.org/10.3390/w14244053
Chicago/Turabian StyleThakur, Sonal, Shivangi Mathur, Saumya Patel, and Biswaranjan Paital. 2022. "Microplastic Accumulation and Degradation in Environment via Biotechnological Approaches" Water 14, no. 24: 4053. https://doi.org/10.3390/w14244053
APA StyleThakur, S., Mathur, S., Patel, S., & Paital, B. (2022). Microplastic Accumulation and Degradation in Environment via Biotechnological Approaches. Water, 14(24), 4053. https://doi.org/10.3390/w14244053