
Optimal Irrigation Regime for Woody Species Potentially Suitable for Effective and Sustainable Afforestation in the Desert Region of Mongolia

 ,
,  ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
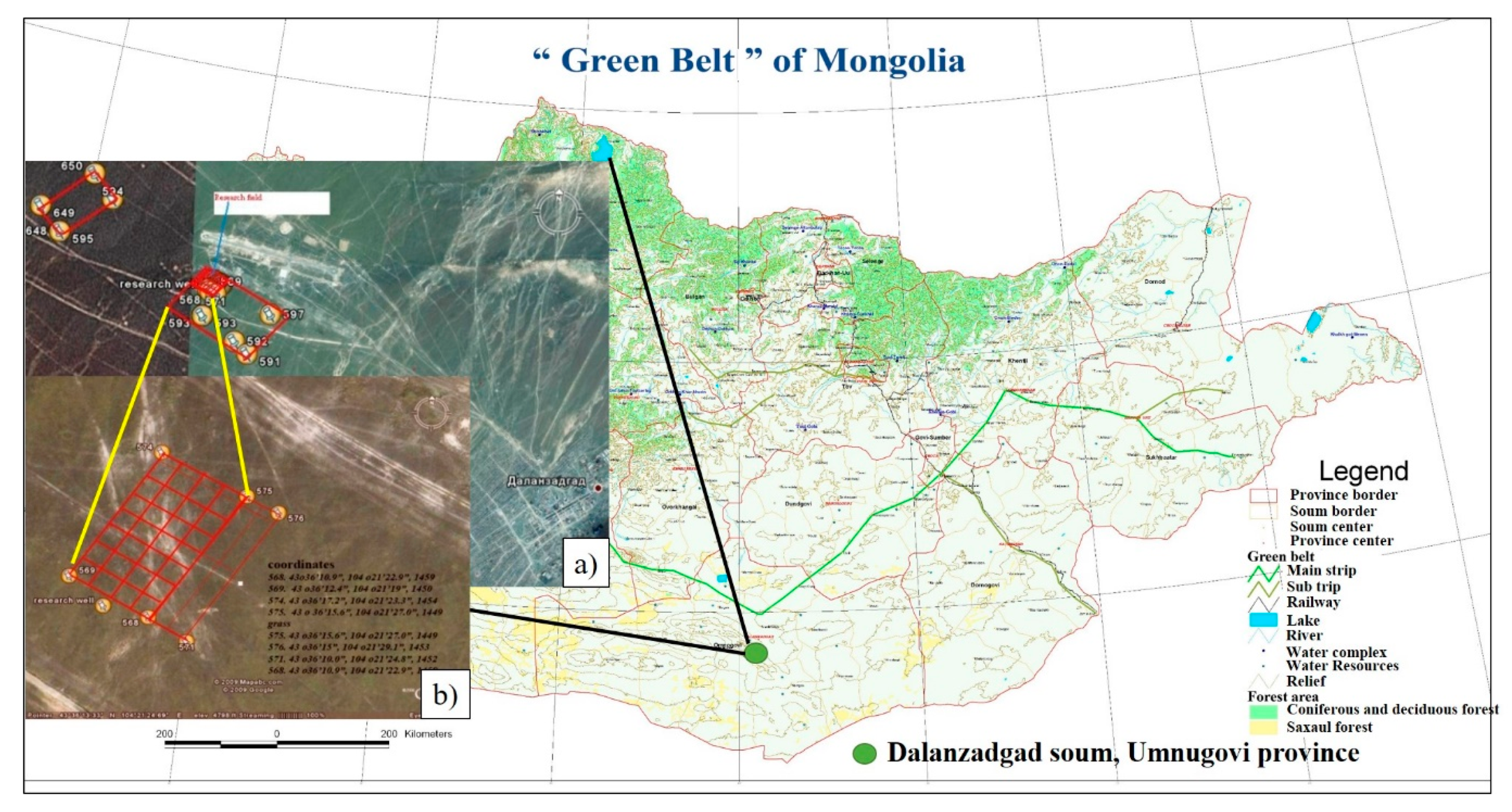
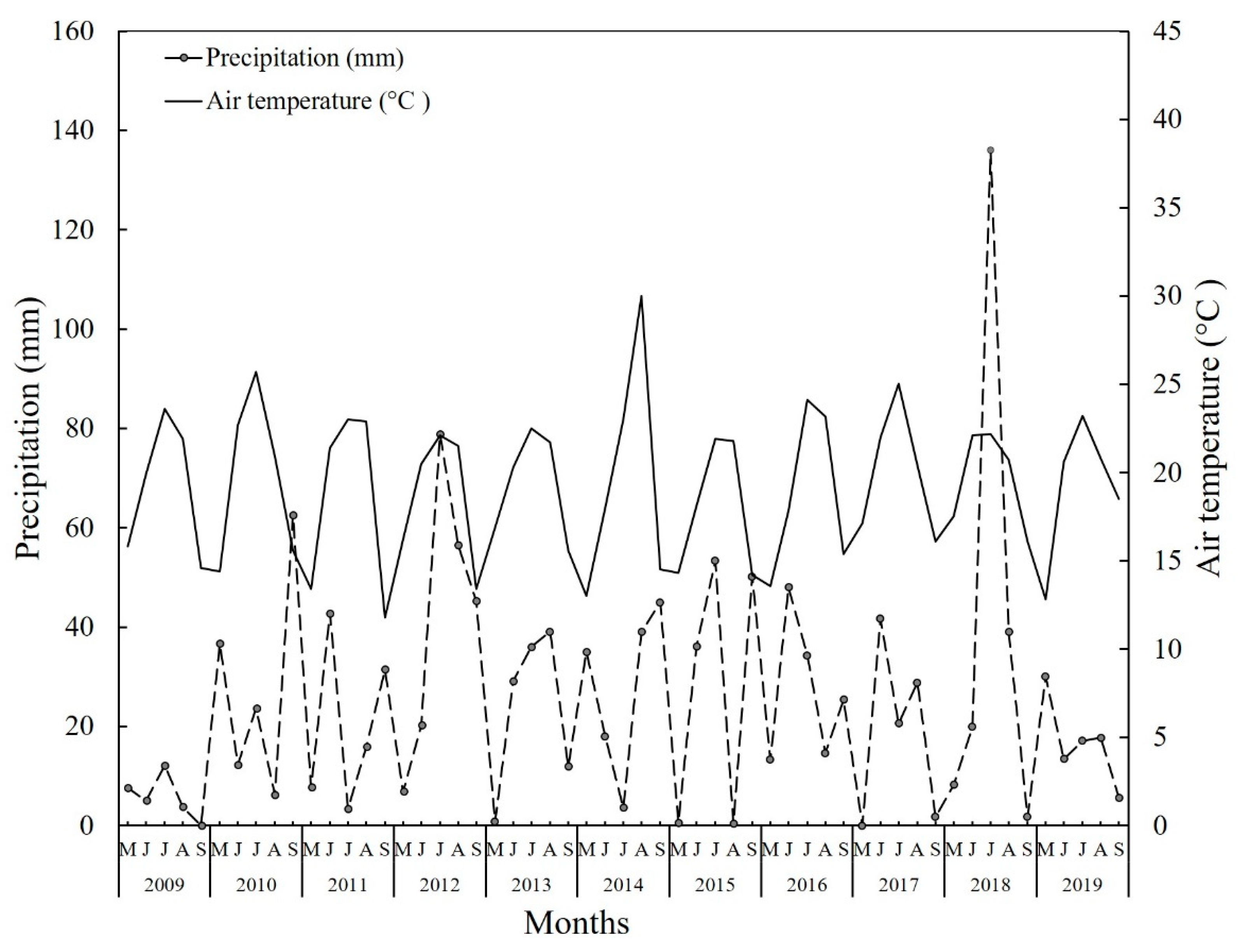
2.1. Experimental Site
2.2. Experimental Design
2.3. Growth Measurements
2.4. Measurements of Morpho-Physiological Responses
Chlorophyll b (Cb, µg/mL) = 20.13 A646 − 5.03 A663
Total chlorophyll (Ca+b, µg/mL) = Ca + Cb
Chlorophyll a/b ratio = Ca/Cb
2.5. Statistical Analysis
3. Results
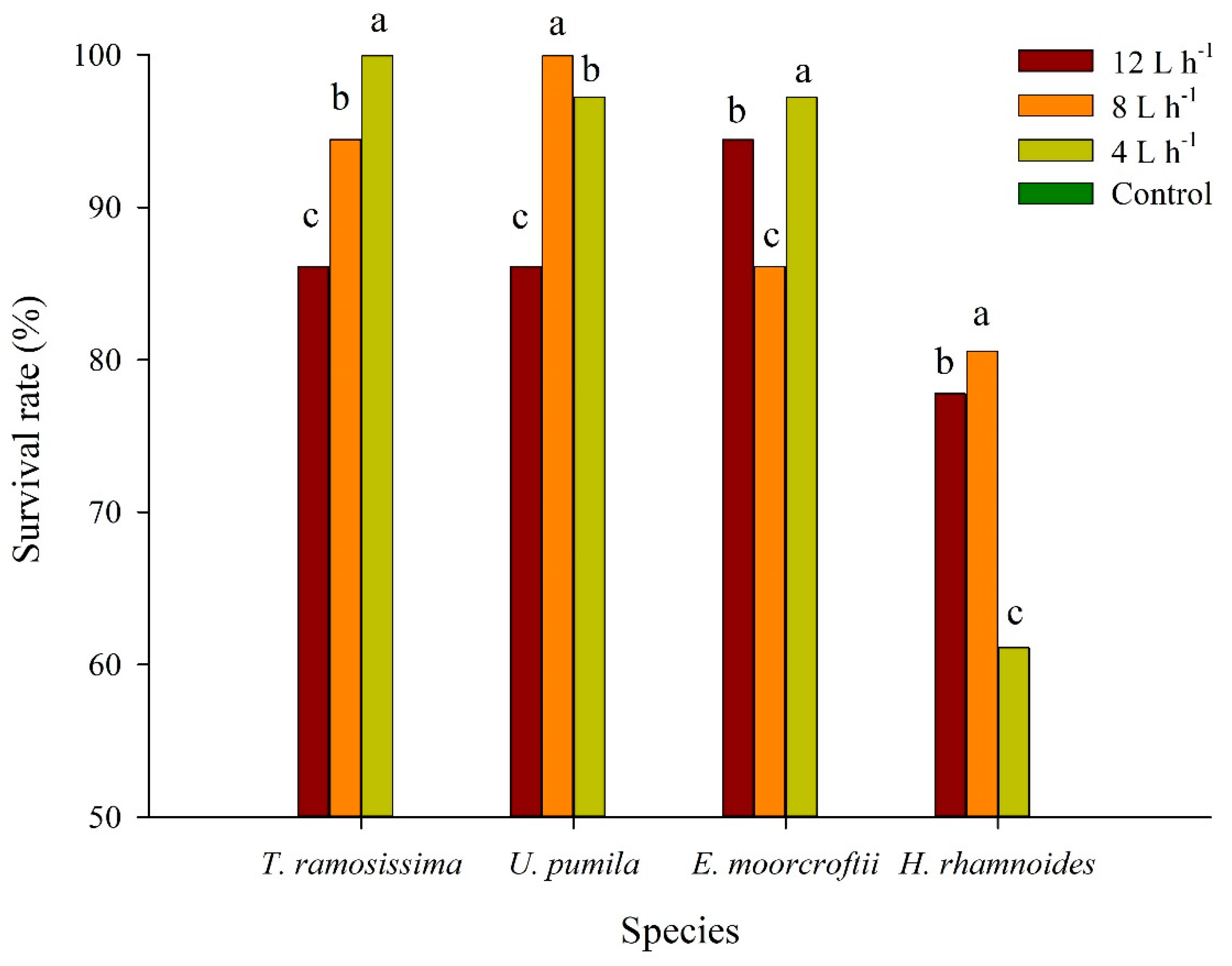
3.1. Survival Rate across Irrigation Treatments
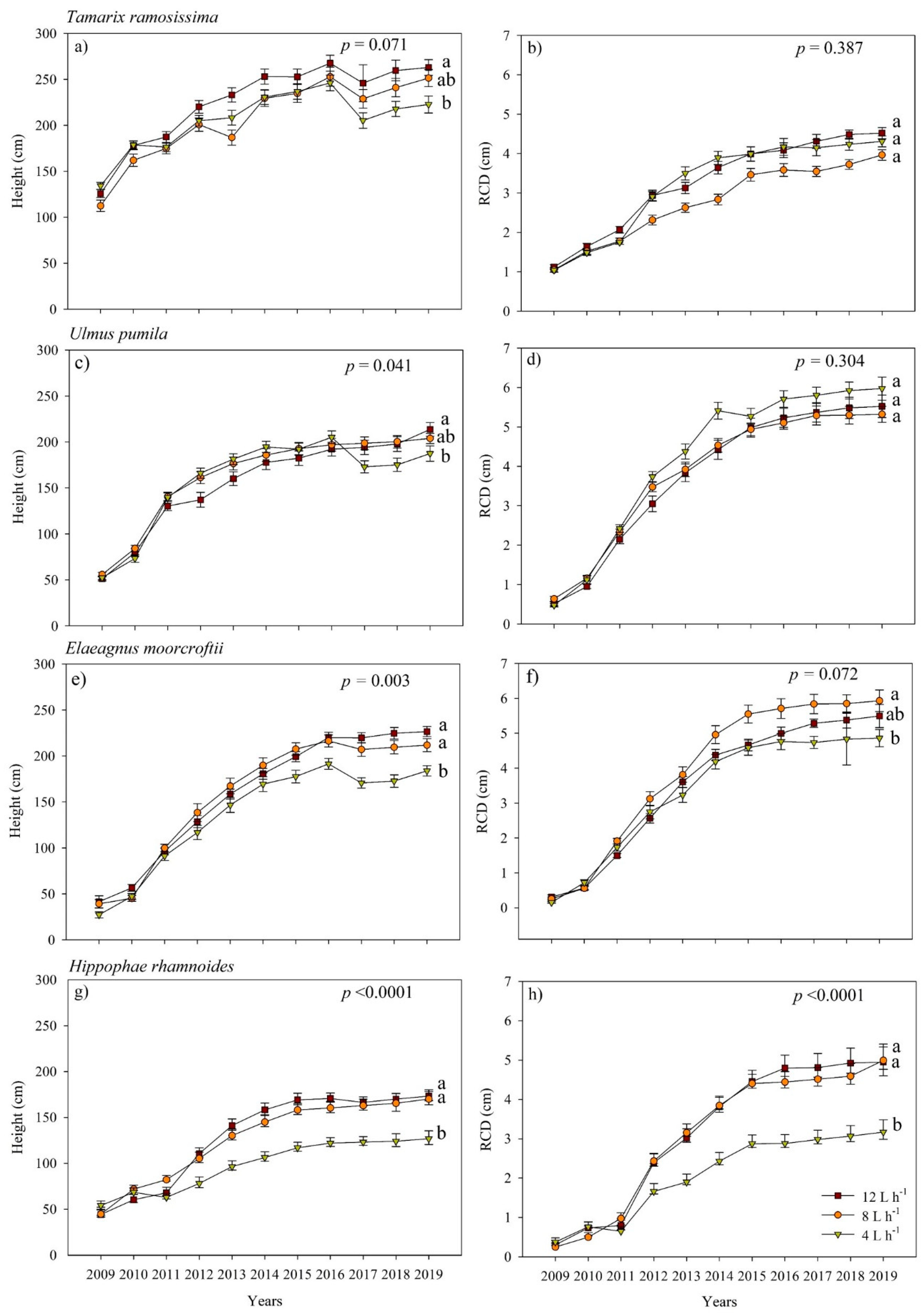
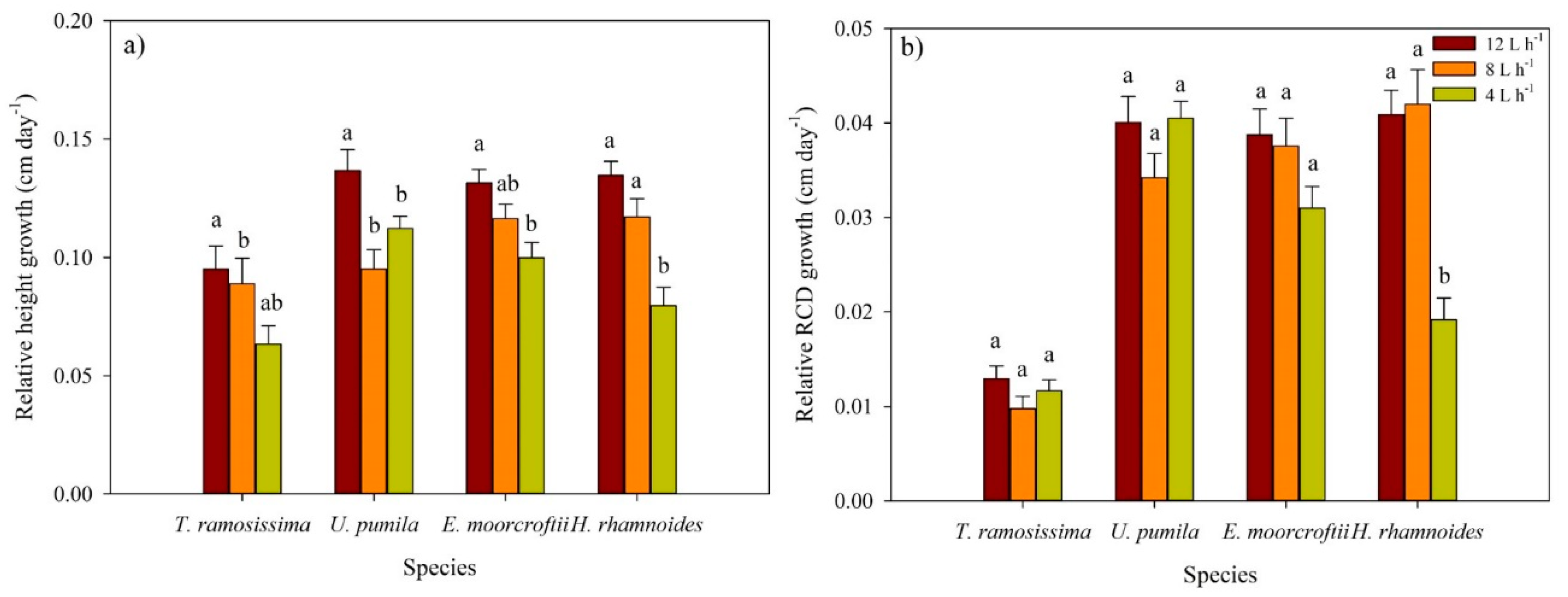
3.2. Root Collar Diameter (RCD) and Height Growth
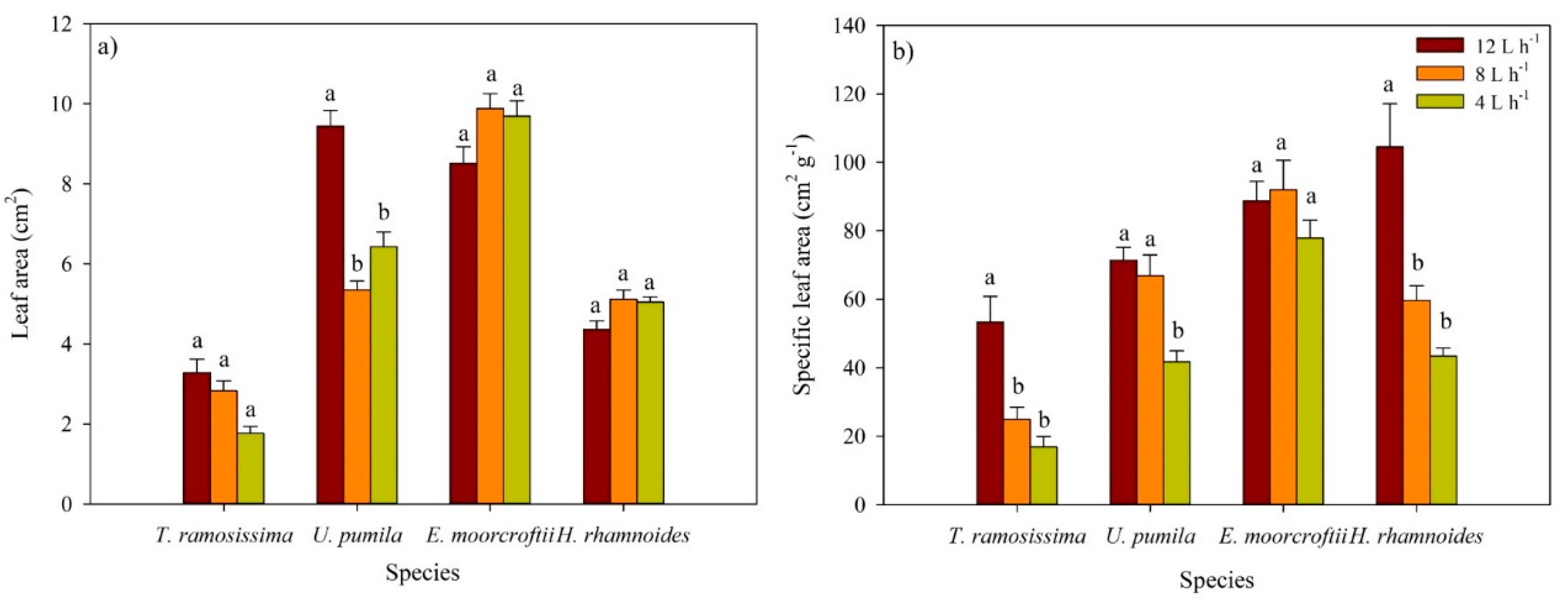
3.3. Leaf Area and Biomass and Specific Leaf Area across Irrigation Treatments
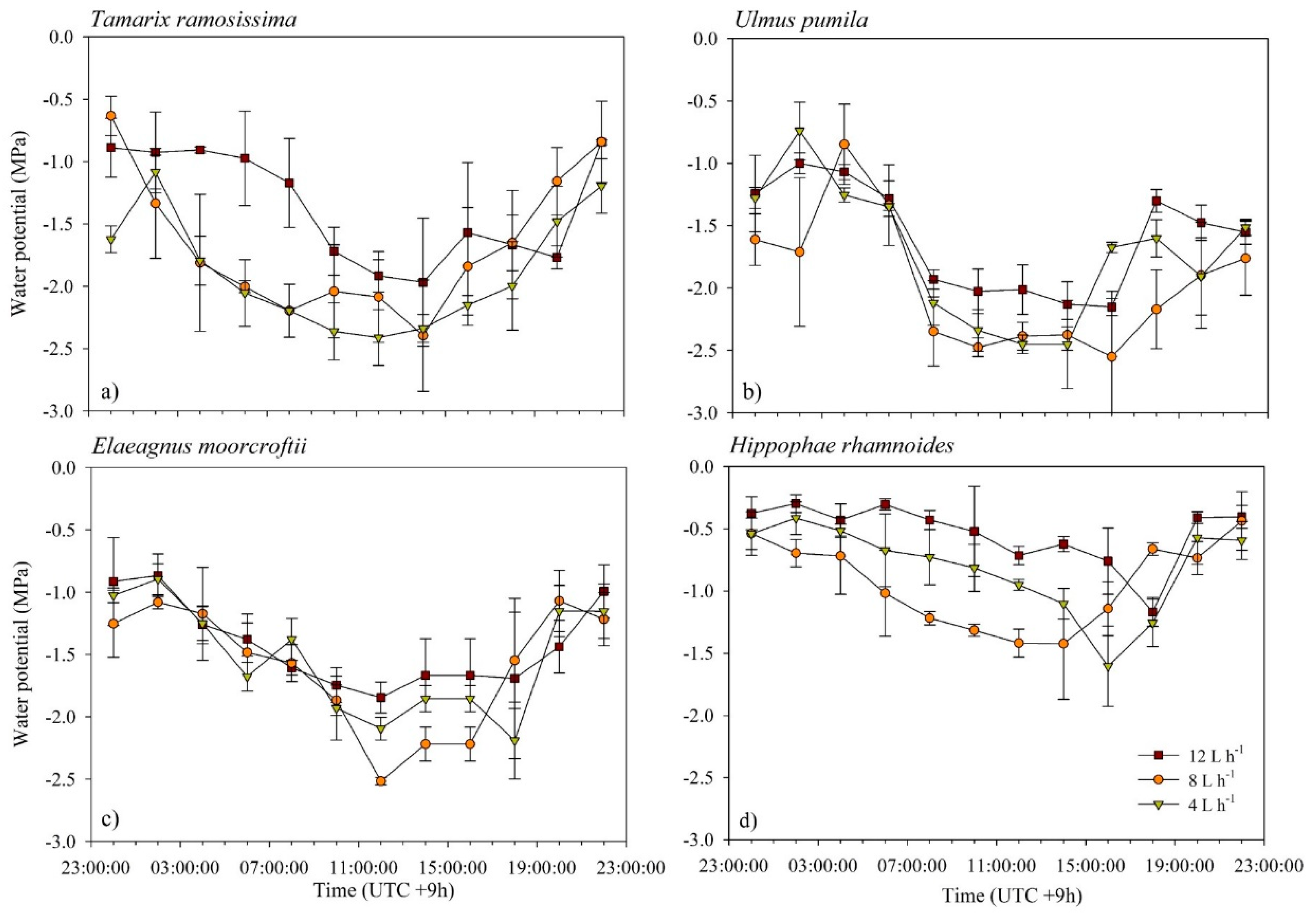
3.4. Changes in Leaf Water Potential across Irrigation Treatments
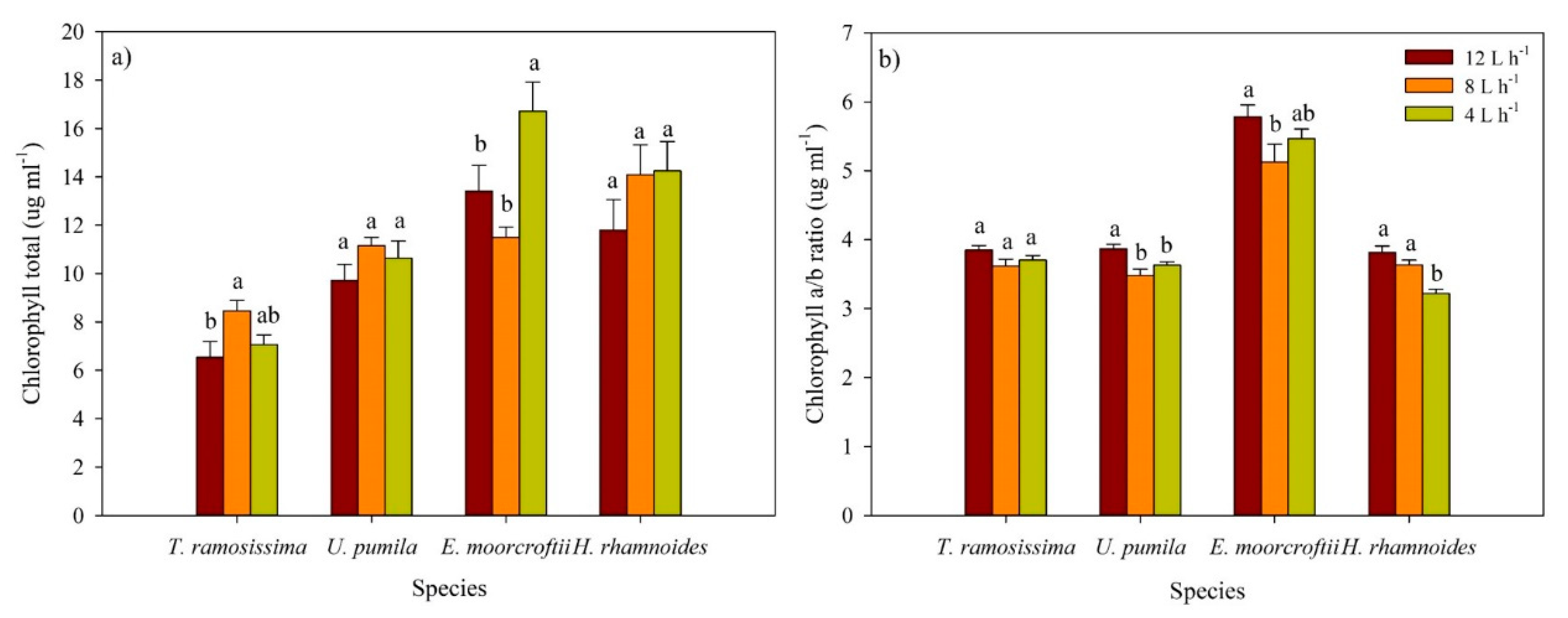
3.5. Variation in Chlorophyll Content
4. Discussion
4.1. Effects of Different Levels of Irrigation on Plant Survivability and Growth of Four Contrasting Species
4.2. Responses of Morpho-Physiological Traits of Four Contrasting Species to Different Irrigation Regimes
4.3. Optimal Irrigation Regime for Each Woody Species Potentially Suitable for Effective and Sustainable Afforestation in Mongolia
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ye, G.; Ma, Y.; Feng, Z.; Zhang, X. Transcriptomic analysis of drought stress responses of sea buckthorn (Hippophae rhamnoides subsp. sinensis) by RNA-Seq. PLoS ONE 2018, 13, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Xu, D.; Wang, Z.; Zhang, Y. Balance of water supply and consumption during ecological restoration in arid regions of Inner Mongolia, China. J. Arid Environ. 2021, 186, 104406. [Google Scholar] [CrossRef]
- Chasek, P.; Akhtar-Schuster, M.; Orr, B.J.; Luise, A.; Rakoto Ratsimba, H.; Safriel, U. Land degradation neutrality: The science-policy interface from the UNCCD to national implementation. Environ. Sci. Policy 2019, 92, 182–190. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, M.; Zhang, H.; Du, Z. Vegetation and soil wind erosion dynamics of sandstorm control programs in the agro-pastoral transitional zone of northern China. Front. Earth Sci. 2019, 13, 430–443. [Google Scholar] [CrossRef]
- Asbjornsen, H.; Goldsmith, G.R.; Alvarado-Barrientos, M.S.; Rebel, K.; Van Osch, F.P.; Rietkerk, M.; Chen, J.; Gotsch, S.; Tobón, C.; Geissert, D.R.; et al. Ecohydrological advances and applications in plant-water relations research: A review. J. Plant Ecol. 2011, 4, 3–22. [Google Scholar] [CrossRef]
- Farooq, M.; Gogoi, N.; Barthakur, S.; Baroowa, B.; Bharadwaj, N.; Alghamdi, S.S.; Siddique, K.H.M. Drought Stress in Grain Legumes during Reproduction and Grain Filling. J. Agron. Crop Sci. 2017, 203, 81–102. [Google Scholar] [CrossRef]
- Padilla, F.M.; Pugnaire, F.I. Rooting depth and soil moisture control Mediterranean woody seedling survival during drought. Funct. Ecol. 2007, 21, 489–495. [Google Scholar] [CrossRef]
- Sukhbaatar, G.; Ganbaatar, B.; Jamsran, T.; Purevragchaa, B.; Nachin, B.; Gradel, A. Assessment of early survival and growth of planted Scots pine (Pinus sylvestris) seedlings under extreme continental climate conditions of northern Mongolia. J. For. Res. 2019. [Google Scholar] [CrossRef] [Green Version]
- Sussmilch, F.C.; McAdam, S.A.M. Surviving a dry future: Abscisic acid (ABA)-mediated plant mechanisms for conserving water under low humidity. Plants 2017, 6, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wairiu, M. Land degradation and sustainable land management practices in Pacific Island Countries. Reg. Environ. Chang. 2017, 17, 1053–1064. [Google Scholar] [CrossRef]
- Cao, S.; Zhang, J.; Chen, L.; Zhao, T. Ecosystem water imbalances created during ecological restoration by afforestation in China, and lessons for other developing countries. J. Environ. Manage. 2016, 183, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Buckley, T.N.; He, X.; Zhang, X.; Zhang, C.; Luo, Z.; Wang, H.; Sterling, N.; Guan, H. Improvement of a simplified process-based model for estimating transpiration under water-limited conditions. Hydrol. Process. 2019, 33, 1670–1685. [Google Scholar] [CrossRef]
- Stropp, J.; dos Santos, I.M.; Correia, R.A.; dos Santos, J.G.; Silva, T.L.P.; dos Santos, J.W.; Ladle, R.J.; Malhado, A.C.M. Drier climate shifts leaf morphology in Amazonian trees. Oecologia 2017, 185, 525–531. [Google Scholar] [CrossRef]
- Yan, Y.; Liu, Q.; Zhang, Q.; Ding, Y.; Li, Y. Adaptation of dominant species to drought in the inner mongolia grassland – species level and functional type level analysis. Front. Plant Sci. 2019, 10, 231. [Google Scholar] [CrossRef]
- Karamouz, M.; Mohammadpour, P.; Mahmoodzadeh, D. Assessment of Sustainability in Water Supply-Demand Considering Uncertainties. Water Resour. Manag. 2017, 31, 3761–3778. [Google Scholar] [CrossRef]
- Hernandez, J.O.; Quimado, M.O.; Fernando, E.S.; Pulan, D.E.; Malabrigo, P.L.; Maldia, L.S.J. Functional traits of stem and leaf of Wrightia candollei S. Vidal. Philipp. J. Sci. 2019, 148, 307–314. [Google Scholar]
- Eziz, A.; Yan, Z.; Tian, D.; Han, W.; Tang, Z.; Fang, J. Drought effect on plant biomass allocation: A meta-analysis. Ecol. Evol. 2017, 7, 11002–11010. [Google Scholar] [CrossRef]
- Cutler, J.M.; Rains, D.W.; Loomis, R.S. The Importance of Cell Size in the Water Relations of Plants. Physiol. Plant. 1977, 40, 255–260. [Google Scholar] [CrossRef]
- Dale, J.E. The Control of Leaf Expansion. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1988, 39, 267–295. [Google Scholar] [CrossRef]
- Tezara, W.; Marín, O.; Rengifo, E.; Martínez, D.; Herrera, A. Photosynthesis and photoinhibition in two xerophytic shrubs during drought. Photosynthetica 2005, 43, 37–45. [Google Scholar] [CrossRef]
- Maréchaux, I.; Bartlett, M.K.; Sack, L.; Baraloto, C.; Engel, J.; Joetzjer, E.; Chave, J. Drought tolerance as predicted by leaf water potential at turgor loss point varies strongly across species within an Amazonian forest. Funct. Ecol. 2015, 29, 1268–1277. [Google Scholar] [CrossRef]
- Scoffoni, C.; McKown, A.D.; Rawls, M.; Sack, L. Dynamics of leaf hydraulic conductance with water status: Quantification and analysis of species differences under steady state. J. Exp. Bot. 2012, 63, 643–658. [Google Scholar] [CrossRef] [PubMed]
- Leuschner, C.; Wedde, P.; Lübbe, T. The relation between pressure–volume curve traits and stomatal regulation of water potential in five temperate broadleaf tree species. Ann. For. Sci. 2019, 76. [Google Scholar] [CrossRef]
- Mclean, E.H.; Prober, S.M.; Stock, W.D.; Steane, D.A.; Potts, B.M.; Vaillancourt, R.E.; Byrne, M. Plasticity of functional traits varies clinally along a rainfall gradient in Eucalyptus tricarpa. Plant Cell Environ. 2014, 37, 1440–1451. [Google Scholar] [CrossRef] [Green Version]
- Warren, C.R.; Tausz, M.; Adams, M.A. Does rainfall explain variation in leaf morphology and physiology among populations of red ironbark (Eucalyptus sideroxylon subsp. tricarpa) grown in a common garden? Tree Physiol. 2005, 25, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- Park, G.E.; Lee, D.K.; Kim, K.W.; Batkhuu, N.O.; Tsogtbaatar, J.; Zhu, J.J.; Jin, Y.; Park, P.S.; Hyun, J.O.; Kim, H.S. Morphological characteristics and water-use efficiency of siberian elm trees (Ulmus pumila L.) within arid regions of northeast asia. Forests 2016, 7, 280. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.; Ser-Oddamba, B.; Batkhuu, N.-O.; Seok Kim, H. Comparison of water use efficiency and biomass production in 10-year-old Populus sibirica and Ulmus pumila plantations in Lun soum, Mongolia. Forest Sci. Technol. 2019, 15, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Byambadorj, S.O.; Chiatante, D.; Akhmadi, K.; Lunten, J.; Ochirbat, B.; Park, B.B.; Scippa, G.S.; Montagnoli, A.; Nyam-Osor, B. The effect of different watering regimes and fertilizer addition on the growth of tree species used to afforest the semi-arid steppe of Mongolia. Plant Biosyst. 2020, 154, 1–12. [Google Scholar] [CrossRef]
- Ser-Oddamba, B.; Khaulenbek, A.; Janchivdorj, L.; Batkhishig, O.; Park, B.B.; Batkhuu, N.-O. The effect of different watering regimes on the growth of trees and shrubs in desert regions of Mongolia. Mong. J. Biol. Sci. 2020, 18, 55–64. [Google Scholar] [CrossRef]
- Yang, B.; Wang, R.; Xiao, H.; Cao, Q.; Liu, T. Spatio-temporal variations of soil water content and salinity around individual Tamarix ramosissima in a semi-arid saline region of the upper Yellow River, northwest China. J. Arid Land 2018, 10, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhang, Q.; Li, A.; Liang, C. Historical landscape dynamics of Inner Mongolia: Patterns, drivers, and impacts. Landsc. Ecol. 2015, 30, 1579–1598. [Google Scholar] [CrossRef]
- Ykhanbai, H. Mongolia Forestry Outlook Study. 2010. Available online: http://www.fao.org/3/am616e/am616e.pdf (accessed on 2 October 2019).
- Wesche, K.; Miehe, S.; Miehe, G. Plant communities of the Gobi Gurvan Sayhan National Park (South Gobi Aymak, Mongolia). Candollea 2005, 60, 149–205. [Google Scholar]
- Batkhuu, N.-O.; Khaulenbek, A.; Bayasgalan, D.; Batkhishig, O.; Janchivdorj, L.; Tsagaantsooj, N.; Ser-Oddamba, B. Mongolia-Korea Joint “Green Belt” Project Research Report; Art soft LLC: Ulaanbaatar, Mongolia, 2012.
- NAMEM The National Agency for Meteorology and Environmental Monitoring of Mongolia. Weather Data. Available online: http://namem.gov.mn/eng/?p=56 (accessed on 2 October 2019).
- Thompson, J.R.; Schultz, R.C. Root system morphology of Quercus rubra L. planting stock and 3-year field performance in Iowa. New For. 1995, 9, 225–236. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Li, Y.; Johnson, D.A.; Su, Y.; Cui, J.; Zhang, T. Specific leaf area and leaf dry matter content of plants growing in sand dunes. Bot. Bull. Acad. Sin. 2005, 46, 127–134. [Google Scholar]
- Kousar, M.; Suresh, G.B.; Lavanya, G.R.; Grard, A. Studies of chlorophyll content by different methods in Black Gram. Int. J. Agric. Res. 2007, 2, 651–654. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Scholander, P.F.; Hammel, H.T.; Hemmingsen, E.A.; Bradstreet, E.D. Hydrostatic Pressure and Osmotic Potential in Leaves of Mangroves and Some Other Plants. Proc. Natl. Acad. Sci. USA 1964, 52, 119–125. [Google Scholar] [CrossRef] [Green Version]
- SAS Institute Inc SAS Software 9.4; SAS Inst. Inc.: Cary, NC, USA, 2014; pp. 1–25.
- Systat Software, I. Sigma Plot Version 12.5; Systat Softw. Inc.: Madison, WI, USA, 2013. [Google Scholar]
- Wang, G.; Liang, X.; Feng, B. The regeneration niches of sand desert plants I. The studies of seed germination Artemisia ordosica, Caragana korshinskii, Hedysarum scoparivm. Acta Bot. Boreali-Occidentalia Sin. 1995, 15, 102–105. [Google Scholar]
- Alizadeh, A.; Alizade, V.; Nassery, L.; Eivazi, A. Effect of Drought Stress on apple dwarf rootstocks. Tech. J. Eng. Appl. Sci. 2011, 1, 86–94. [Google Scholar]
- Ibrahim, Y.M.; Saeed, A.B.; Widaa, A.; Elamin, M. Effect of Irrigation Water Management on Growth of Date Palm offshoots (Phoenix dactylifera) under the River Nile State Conditions. Univ. Khartoum J. Agric. Sci. 2012, 20, 275–285. [Google Scholar]
- Wahba, S.A.; Rahman, S.I.A.; Tayel, M.Y.; Matyn, M.A. Soil moisture, salinity, water use efficiency and sunflower growth as influenced by irrigation, bitumen mulch and plant density. Soil Technol. 1990, 3, 33–44. [Google Scholar] [CrossRef]
- Hérault, B.; Bachelot, B.; Poorter, L.; Rossi, V.; Bongers, F.; Chave, J.; Paine, C.E.T.; Wagner, F.; Baraloto, C. Functional traits shape ontogenetic growth trajectories of rain forest tree species. J. Ecol. 2011, 99, 1431–1440. [Google Scholar] [CrossRef]
- Ryan, M.G.; Yoder, B.J. Hydraulic limits to tree height and tree growth: What keeps trees from growing beyond a certain height? Bioscience 1997, 47, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, W.N.; De Lacerda, C.F.; Da Costa, R.C.; Filho, S.M. Effect of water stress on seedling growth in two species with different abundances: The importance of Stress Resistance Syndrome in seasonally dry tropical forest. Acta Bot. Brasilica 2015, 29, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.J.; Wang, Y.R.; Zhang, J.M. Is reduced seed germination due to water limitation a special survival strategy used by xerophytes in arid dunes? J. Arid Environ. 2010. [Google Scholar] [CrossRef]
- Hernandez, J.O.; Quimado, M.O.; Fernando, E.S.; Province, B. Xerophytic characteristics of Tectona philippinensis Benth. & Hook.f. Philipp. J. Sci. 2016, 145, 259–269. [Google Scholar]
- Liu, Z.; Dickmann, D.I. Abscisic acid accumulation in leaves of two contrasting hybrid poplar clones affected by nitrogen fertilization plus cyclic flooding and soil drying. Tree Physiol. 1992, 11, 109–122. [Google Scholar] [CrossRef]
- Lázaro-Nogal, A.; Matesanz, S.; Godoy, A.; Pérez-Trautman, F.; Gianoli, E.; Valladares, F. Environmental heterogeneity leads to higher plasticity in dry-edge populations of a semi-arid Chilean shrub: Insights into climate change responses. J. Ecol. 2015, 103, 338–350. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Plant-soil interactions in Mediterranean forest and shrublands: Impacts of climatic change. Plant Soil 2013, 365, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Wierzchos, J.; de los Ríos, A.; Ascaso, C. Microorganisms in desert rocks: The edge of life on Earth. Int. Microbiol. 2012, 15, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Urban, L.; Restrepo-Diaz, H. Influences of Abiotic Factors in Growth and Development. Ref. Modul. Life Sci. 2009, 29, 1330–1340. [Google Scholar] [CrossRef]
- Niinemets, Ü. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology 2001, 82, 453–469. [Google Scholar] [CrossRef]
- Ordoñez, J.C.; Van Bodegom, P.M.; Witte, J.P.M.; Bartholomeus, R.P.; Van Dobben, H.F.; Aerts, R. Leaf habit and woodiness regulate different leaf economy traits at a given nutrient supply. Ecology 2010, 91, 3218–3228. [Google Scholar] [CrossRef]
- Veneklaas, E.J.; Santos Silva, M.P.R.M.; Den Ouden, F. Determinants of growth rate in Ficus benjamina L. compared to related faster-growing woody and herbaceous species. Sci. Hortic. (Amst.) 2002, 93, 75–84. [Google Scholar] [CrossRef]
- De La Riva, E.G.; Olmo, M.; Poorter, H.; Ubera, J.L.; Villar, R. Leaf mass per area (LMA) and its relationship with leaf structure and anatomy in 34 mediterranean woody species along a water availability gradient. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weraduwage, S.M.; Chen, J.; Anozie, F.C.; Morales, A.; Weise, S.E.; Sharkey, T.D. The relationship between leaf area growth and biomass accumulation in Arabidopsis thaliana. Front. Plant Sci. 2015, 6, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Onoda, Y.; Westoby, M.; Adler, P.B.; Choong, A.M.F.; Clissold, F.J.; Cornelissen, J.H.C.; Díaz, S.; Dominy, N.J.; Elgart, A.; Enrico, L.; et al. Global patterns of leaf mechanical properties. Ecol. Lett. 2011, 14, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Read, J.; Sanson, G.D.; De Garine-Wichatitsky, M.; Jaffré, T. Sclerophylly in two contrasting tropical environments: Low nutrients vs. low rainfall. Am. J. Bot. 2006, 93, 1601–1614. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Alonzo, R.; Ewers, F.W.; Sack, L. Ecological variation in leaf biomechanics and its scaling with tissue structure across three mediterranean-climate plant communities. Funct. Ecol. 2013, 27, 544–554. [Google Scholar] [CrossRef] [Green Version]
- Jinwen, L.; Jingping, Y.; Dongsheng, L.; Pinpin, F.; Tiantai, G.; Changshui, G.; Wenyue, C. Chlorophyll meter’s estimate of weight-based nitrogen concentration in rice leaf is influenced by leaf thickness. Plant Prod. Sci. 2011, 14, 177–183. [Google Scholar] [CrossRef]
- Peng, S. Single-leaf and canopy photosynthesis of rice. Stud. Plant Sci. 2000, 7, 213–228. [Google Scholar] [CrossRef]
- Evans, J.R.; Poorter, H. Photosynthetic acclimation of plants to growth irradiance: The relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Plant Cell Environ. 2001, 24, 755–767. [Google Scholar] [CrossRef]
- Vesala, T.; Sevanto, S.; Grönholm, T.; Salmon, Y.; Nikinmaa, E.; Hari, P.; Hölttä, T. Effect of leaf water potential on internal humidity and CO2 dissolution: Reverse transpiration and improved water use efficiency under negative pressure. Front. Plant Sci. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Wu, W.; Zhou, X.; Zhang, D.; Yu, Y.; Wang, H.; Wang, Q. The ecosystem effects of sand-binding shrub Hippophae rhamnoides in alpine semi-arid desert in the Northeastern Qinghai-Tibet plateau. Land 2019, 8, 183. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Yu, B.; Zhao, C.; Nowak, R.S.; Zhao, Z.; Sheng, Y.; Li, J. Physiological and morphological responses of Tamarix ramosissima and Populus euphratica to altered groundwater availability. Tree Physiol. 2013, 33, 57–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swaminathan, P.; Ohrtman, M.; Carinder, A.; Deuja, A.; Wang, C.; Gaskin, J.; Fennell, A.; Clay, S. Water deficit transcriptomic responses differ in the invasive Tamarix chinensis and T. ramosissima established in the southern and northern United States. Plants 2020, 9, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babushkina, E.A.; Zhirnova, D.F.; Belokopytova, L.V.; Tychkov, I.I.; Vaganov, E.A.; Krutovsky, K.V. Response of Four Tree Species to Changing Climate in a Moisture-Limited Area of South Siberia. Forests 2019, 10, 999. [Google Scholar] [CrossRef] [Green Version]
- Dulamsuren, C.; Hauck, M.; Bader, M.; Osokhjargal, D.; Oyungerel, S.; Nyambayar, S.; Runge, M.; Leuschner, C. Water relations and photosynthetic performance in Larix sibirica growing in the forest-steppe ecotone of northern Mongolia. Tree Physiol. 2009, 29, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Xiongwen, C. Ecophysiological and Growth Responses of Elm, Ulmus pumila, to Different Water Tables. J. Biol. Sci. 2005, 5, 813–819. [Google Scholar]
- Sun, J.K.; Zhang, W.H.; Lu, Z.H.; Liu, X.C. Effects of drought stress on gas exchange characteristics and protective enzyme activities in Elaeagnus angustifolia and Grewia. biloba G. Don var. parviflora seedlings. Shengtai Xuebao/Acta Ecol. Sin. 2009, 29, 1330–1340. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Age (Year) | Height (cm) | RCD (mm) | Root Length (cm) |
|---|---|---|---|---|
| Tamarix ramosissima | 2 | 83.6 | 7.7 | 34.5 |
| Ulmus pumila | 2 | 51.4 | 4.4 | 37.2 |
| Elaeagnus moorcroftii | 2 | 52.1 | 5.3 | 33.5 |
| Hippophae rhamnoides | 2 | 39.4 | 3.8 | 21.9 |
| Species | Treatment | June | July | August | September | |
|---|---|---|---|---|---|---|
| Tamarix ramosissima | ψp | 12 L h−1 | −0.38 a | −0.91 a | −0.77 a | −0.98 a |
| 8 L h−1 | −0.45 ab | −1.81 a | −0.84 a | −0.94 a | ||
| 4 L h−1 | −0.67 b | −1.79 a | −1.12 a | −0.67 a | ||
| ψm | 12 L h−1 | −1.05 a | −1.91 a | −1.28 a | −2.38 a | |
| 8 L h−1 | −1.23 a | −2.08 a | −1.45 a | −1.84 a | ||
| 4 L h−1 | −1.76 a | −2.33 a | −2.59 b | −1.84 a | ||
| Ulmus pumila | ψp | 12 L h−1 | −0.31 a | −1.07 a | −0.39 a | −0.61 a |
| 8 L h−1 | −0.39 a | −0.84 a | −1.01 a | −0.96 ab | ||
| 4 L h−1 | −0.51 a | −1.25 a | −0.56 a | −1.06 b | ||
| ψm | 12 L h−1 | −1.62 ab | −2.13 a | −2.52 a | −1.88 a | |
| 8 L h−1 | −1.89 b | −2.37 ab | −1.97 a | −1.86 a | ||
| 4 L h−1 | −1.29 a | −2.45 b | −2.13 a | −2.64 b | ||
| Elaeagnus moorcroftii | ψp | 12 L h−1 | −0.27 a | −1.26 a | −0.27 a | −0.31 a |
| 8 L h−1 | −0.37 a | −1.17 a | −0.37 a | −0.79 a | ||
| 4 L h−1 | −0.40 a | −1.25 a | −0.40 a | −0.71 a | ||
| ψm | 12 L h−1 | −1.20 a | −1.84 a | −0.99 a | −1.91 a | |
| 8 L h−1 | −1.35 a | −2.51 a | −1.11 a | −2.23 a | ||
| 4 L h−1 | −1.09 a | −2.09 a | −1.09 a | −2.46 a | ||
| Hippophae rhamnoides | ψp | 12 L h−1 | −0.24 a | −0.42 a | −0.49 a | −0.22 a |
| 8 L h−1 | −0.36 a | −0.71 a | −0.71 a | −0.35 a | ||
| 4 L h−1 | −0.51 a | −0.51 a | −0.46 a | −0.54 a | ||
| ψm | 12 L h−1 | −0.97 a | −0.71 a | −1.18 a | −1.68 a | |
| 8 L h−1 | −0.80 a | −1.41 b | −1.36 a | −2.31 a | ||
| 4 L h−1 | −1.64 a | −0.95 a | −1.57 a | −2.37 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byambadorj, S.-O.; Park, B.B.; Hernandez, J.O.; Dulamsuren, N.; Sainbuyan, Z.; Altantugs, O.; Sharavdorj, K.; Seong, I.K.; Batkhuu, N.-O. Optimal Irrigation Regime for Woody Species Potentially Suitable for Effective and Sustainable Afforestation in the Desert Region of Mongolia. Land 2021, 10, 212. https://doi.org/10.3390/land10020212
Byambadorj S-O, Park BB, Hernandez JO, Dulamsuren N, Sainbuyan Z, Altantugs O, Sharavdorj K, Seong IK, Batkhuu N-O. Optimal Irrigation Regime for Woody Species Potentially Suitable for Effective and Sustainable Afforestation in the Desert Region of Mongolia. Land. 2021; 10(2):212. https://doi.org/10.3390/land10020212
Chicago/Turabian StyleByambadorj, Ser-Oddamba, Byung Bae Park, Jonathan O. Hernandez, Narantugs Dulamsuren, Zoljargal Sainbuyan, Oyuntugs Altantugs, Khulan Sharavdorj, In Kyeong Seong, and Nyam-Osor Batkhuu. 2021. "Optimal Irrigation Regime for Woody Species Potentially Suitable for Effective and Sustainable Afforestation in the Desert Region of Mongolia" Land 10, no. 2: 212. https://doi.org/10.3390/land10020212
APA StyleByambadorj, S. -O., Park, B. B., Hernandez, J. O., Dulamsuren, N., Sainbuyan, Z., Altantugs, O., Sharavdorj, K., Seong, I. K., & Batkhuu, N. -O. (2021). Optimal Irrigation Regime for Woody Species Potentially Suitable for Effective and Sustainable Afforestation in the Desert Region of Mongolia. Land, 10(2), 212. https://doi.org/10.3390/land10020212