
Effects of Grazer Exclusion on Carbon Cycling in Created Freshwater Wetlands


Abstract
:1. Introduction
2. Methods
2.1. Site Description
2.2. Grazer Abundance
2.3. Vegetation Cover, Biomass and Elemental Composition
2.4. Soil Characteristics and Elemental Composition
2.5. Decomposition
2.6. Gas Fluxes
2.7. Statistical Analyses
3. Results
3.1. Grazing Pressure
3.2. Hydrologic Conditions
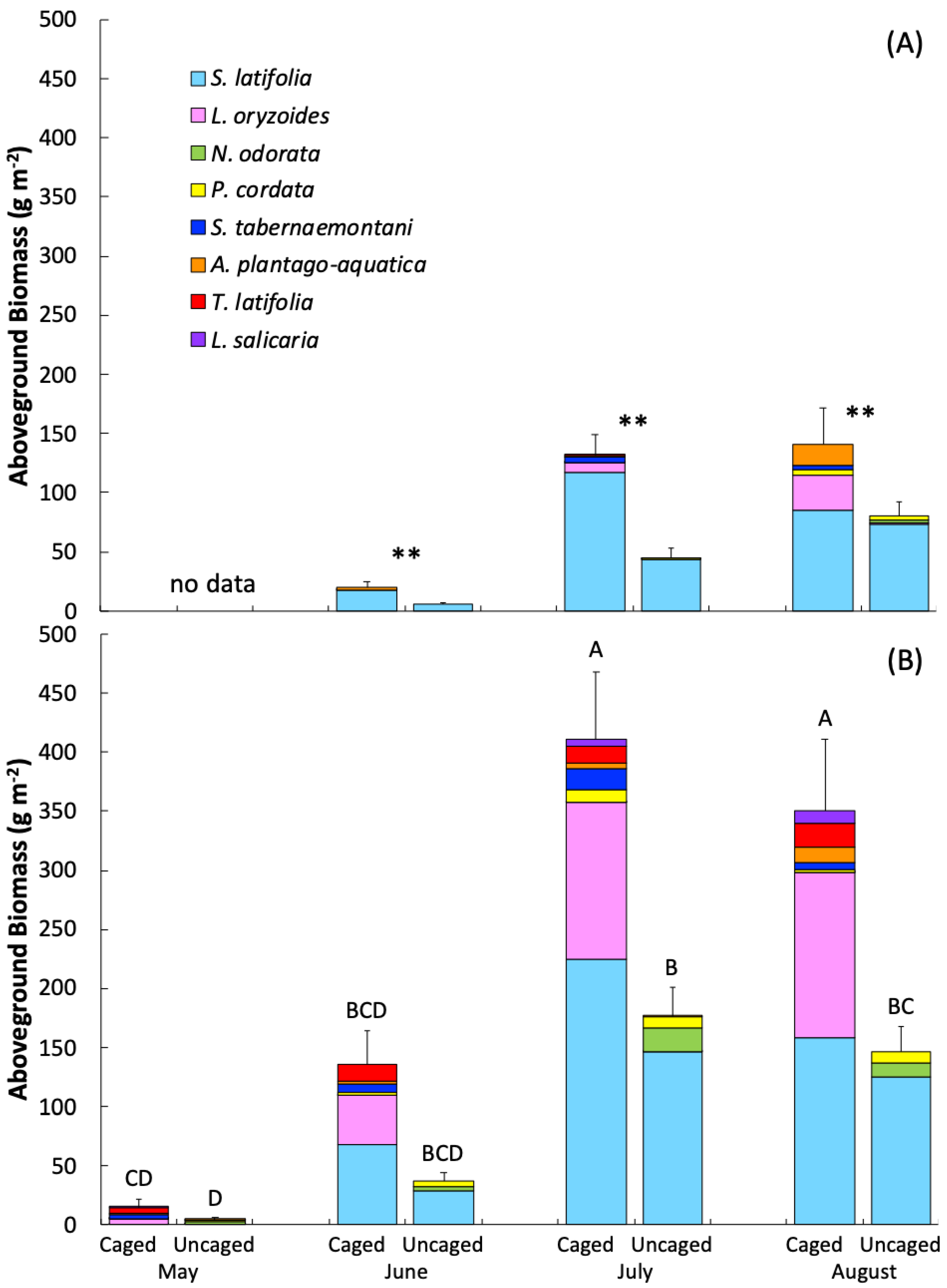
3.3. Vegetation
3.4. Soil and Belowground Biomass
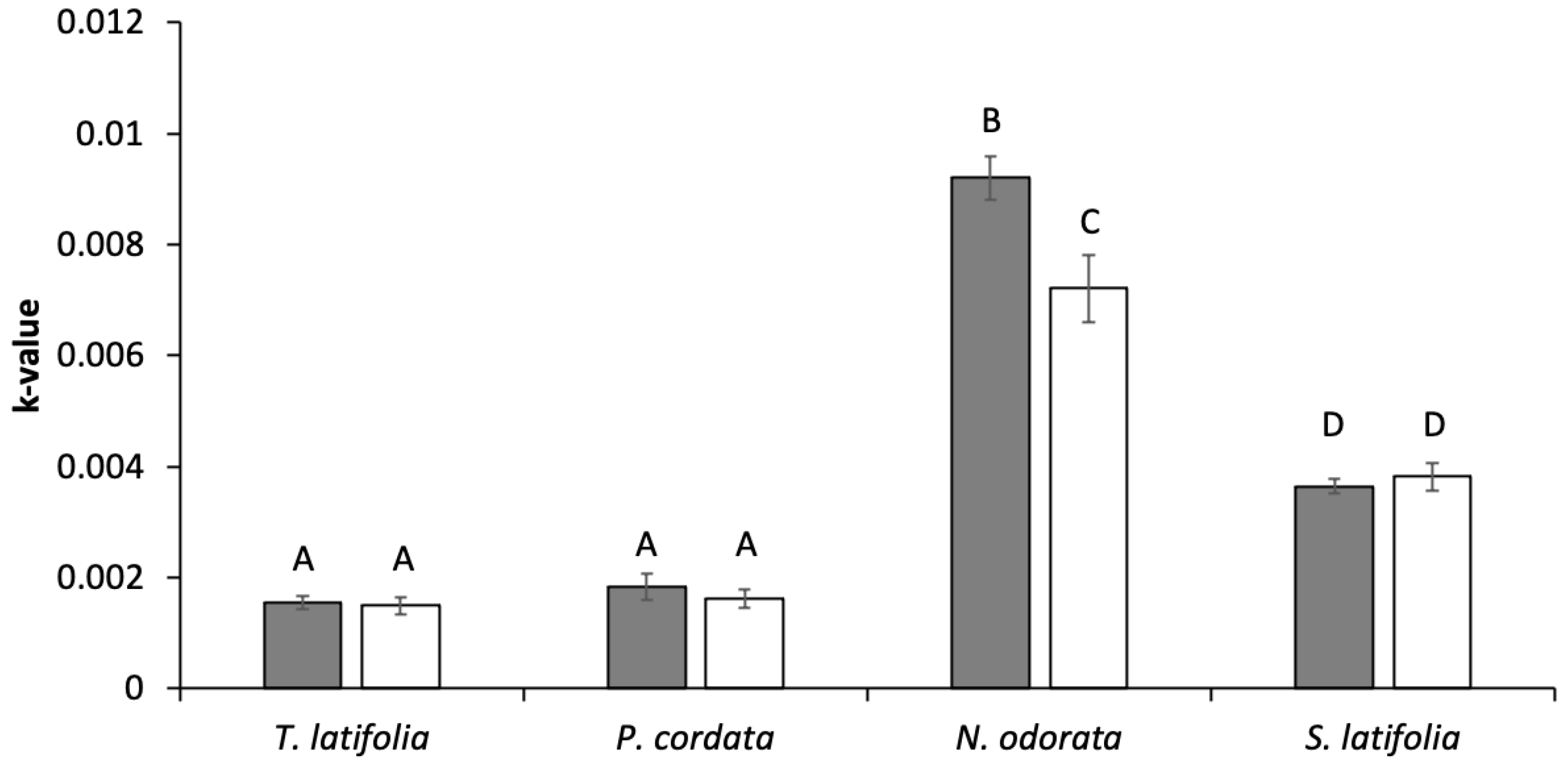
3.5. Decomposition Rates
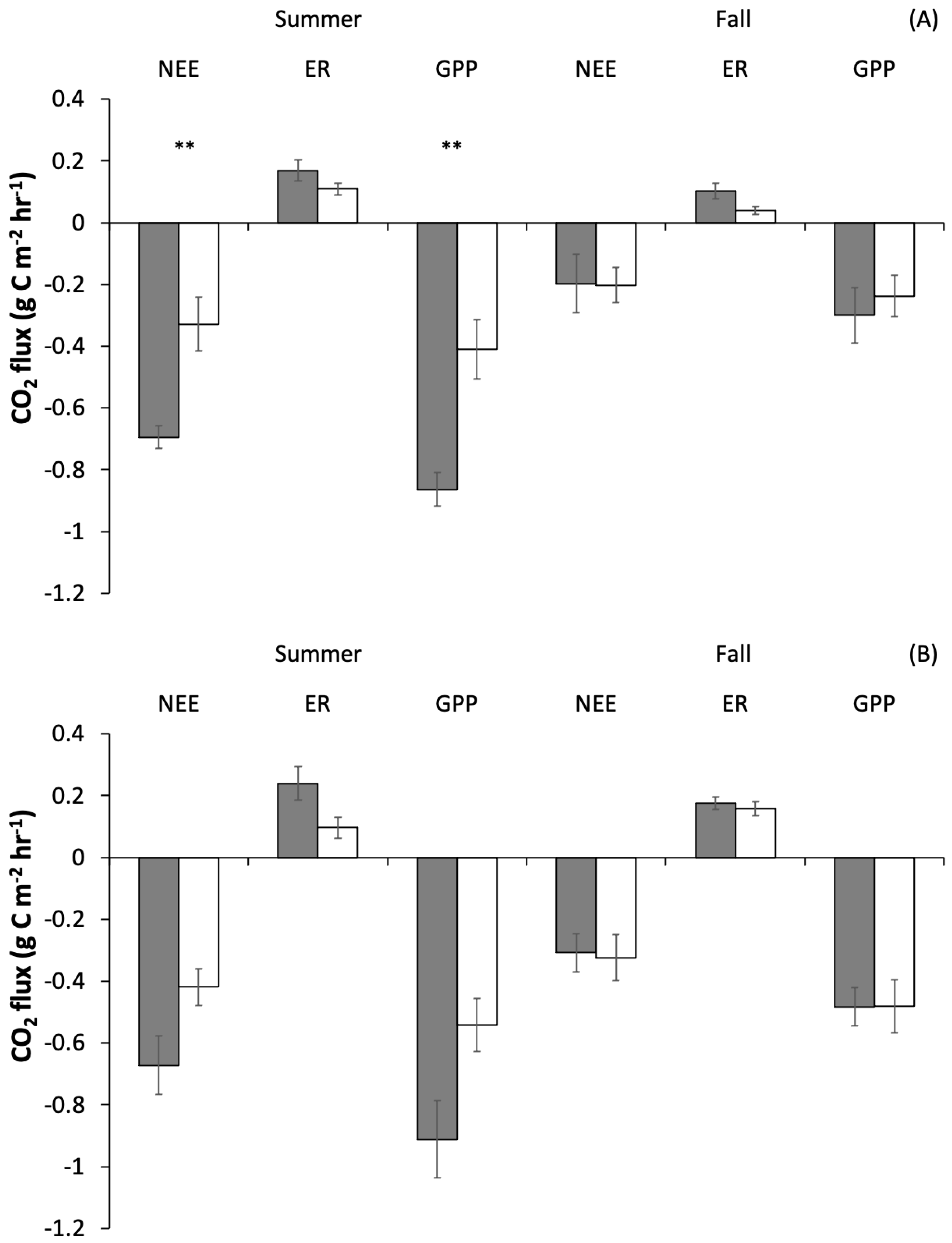
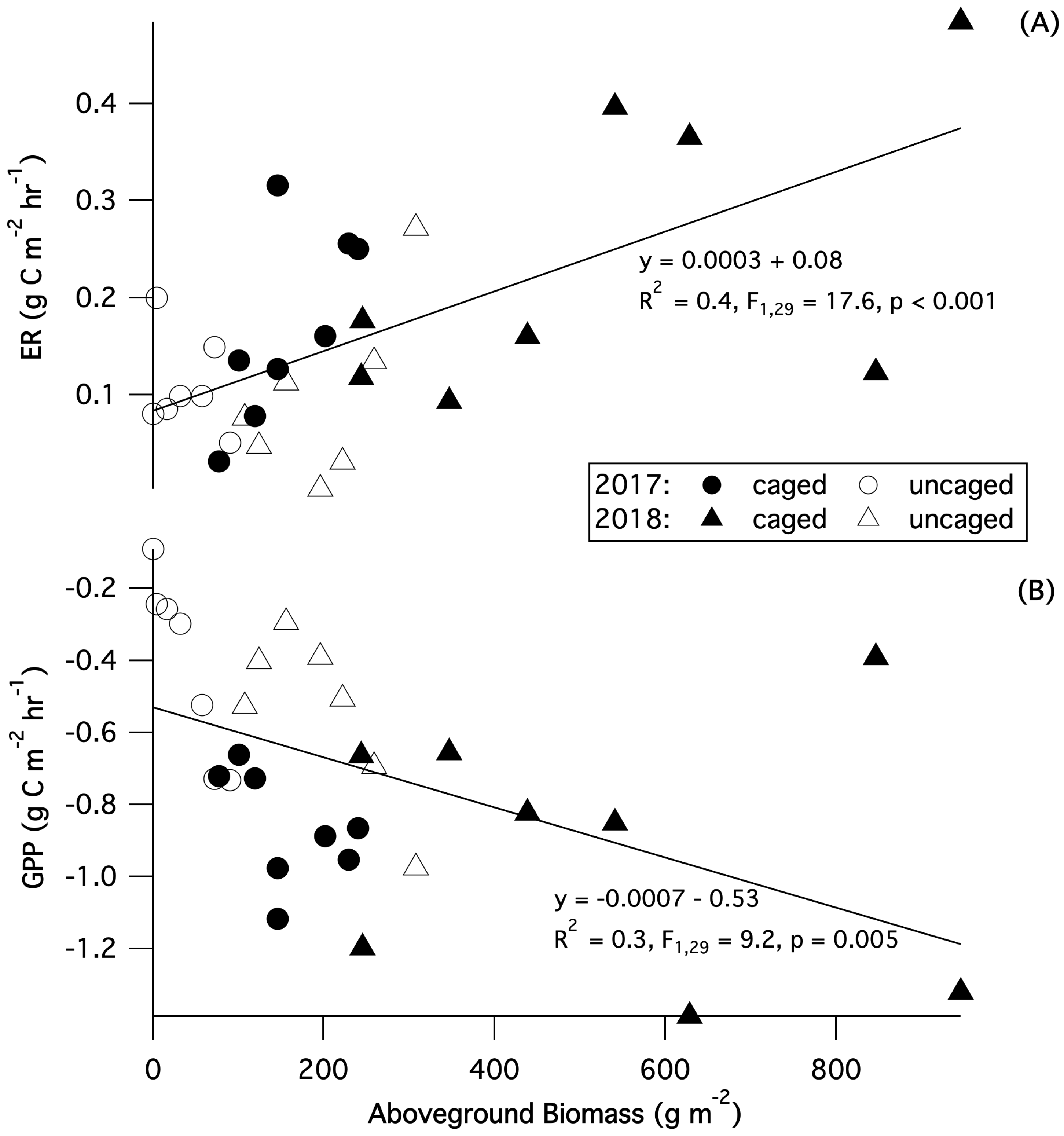
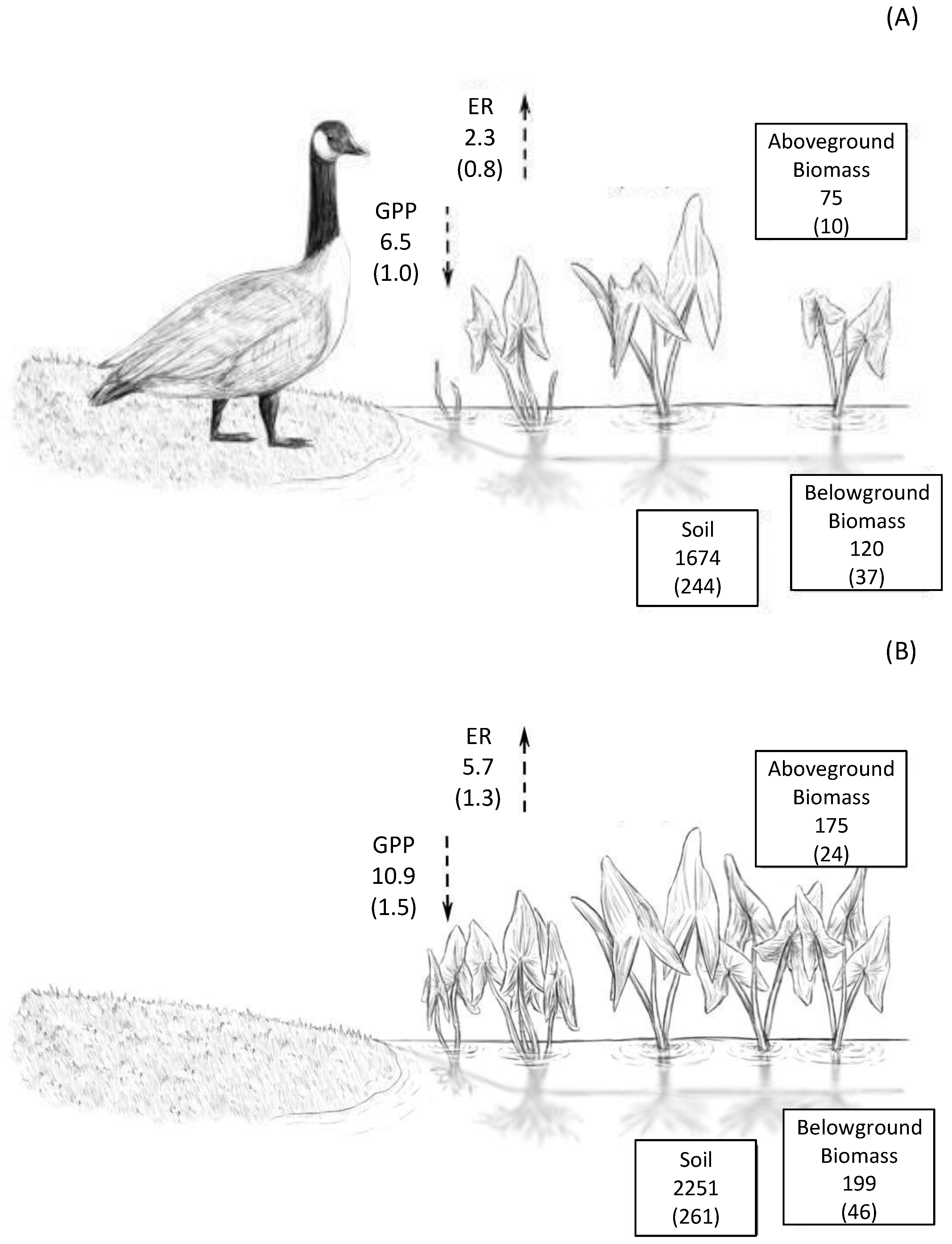
3.6. Gas Fluxes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zedler, J.B.; Kercher, S. WETLAND RESOURCES: Status, Trends, Ecosystem Services, and Restorability. Annu. Rev. Environ. Resour. 2005, 30, 39–74. [Google Scholar] [CrossRef] [Green Version]
- Aerts, R.; Verhoeven, J.T.A.; Whigham, D.F. Plant-Mediated Controls On Nutrient Cycling in Temperate Fens and Bogs. Ecology 1999, 70, 2170–2181. [Google Scholar] [CrossRef]
- DeAngelis, D.L.; Bartell, S.M.; Brenkert, A.L. Effects of Nutrient Recycling and Food-Chain Length on Resilience. Am. Nat. 1989, 134, 778–805. [Google Scholar] [CrossRef]
- Costanza, R.; D’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Chmura, G.L.; Anisfeld, S.C.; Cahoon, D.R.; Lynch, J.C. Global carbon sequestration in tidal, saline wetland soils. Glob. Biogeochem. Cycles 2003, 17, 1111. [Google Scholar] [CrossRef]
- Kayranli, B.; Scholz, M.; Mustafa, A.; Hedmark, Å. Carbon Storage and Fluxes within Freshwater Wetlands: A Critical Review. Wetlands 2010, 30, 111–124. [Google Scholar] [CrossRef]
- Campbell, D.A.; Cole, C.A.; Brooks, R.P. A comparison of created and natural wetlands in Pennsylvania, USA. Wetl. Ecol. Manag. 2002, 10, 41–49. [Google Scholar] [CrossRef]
- Fennessy, M.S.; Rokosch, A.; Mack, J.J. Patterns of plant decomposition and nutrient cycling in natural and created wetlands. Wetlands 2008, 28, 300–310. [Google Scholar] [CrossRef]
- Moreno-Mateos, D.; Power, M.E.; Comín, F.A.; Yockteng, R. Structural and functional loss in restored wetland ecosystems. PLoS Biol. 2012, 10, e1001247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Mateos, D.; Alberdi, A.; Morriën, E.; van der Putten, W.H.; Rodríguez-Uña, A.; Montoya, D. The long-term restoration of ecosystem complexity. Nat. Ecol. Evol. 2020, 4, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.L.; Fujii, R. Plant community, primary productivity, and environmental conditions following wetland re-establishment in the Sacramento-San Joaquin Delta, California. Wetl. Ecol. Manag. 2010, 18, 1–16. [Google Scholar] [CrossRef]
- Rothman, E.; Bouchard, V. Regulation of carbon processes by macrophyte species in a Great Lakes coastal wetland. Wetlands 2007, 27, 1134–1143. [Google Scholar] [CrossRef]
- Collins, M.E.; Kuehl, R. Organic Matter Accumulation and Organic Soils; Lewis Publishers: Boca Raton, FL, USA, 2000. [Google Scholar]
- Mitsch, W.; Gosselink, J. Wetlands; Wiley: New York, NY, USA, 2007. [Google Scholar]
- De Deyn, G.B.; Cornelissen, J.H.C.; Bardgett, R.D. Plant functional traits and soil carbon sequestration in contrasting biomes. Ecol. Lett. 2008, 11, 516–531. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Bernal, B.; Nahlik, A.M.; Mander, Ü.; Zhang, L.; Anderson, C.J.; Jørgensen, S.E.; Brix, H. Wetlands, carbon, and climate change. Landsc. Ecol. 2013, 28, 583–597. [Google Scholar] [CrossRef]
- Yu, L.; Huang, Y.; Sun, F.; Sun, W. A synthesis of soil carbon and nitrogen recovery after wetland restoration and creation in the United States. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossler, K.; Bouchard, V. Soil development and establishment of carbon-based properties in created freshwater marshes. Ecol. Appl. 2010, 20, 539–553. [Google Scholar] [CrossRef] [PubMed]
- Murkin, H.R.; Murkin, E.J.; Ball, J.P. Avian Habitat Selection and Prairie Wetland Dynamics: A 10-Year Experiment. Ecol. Appl. 1997, 7, 1144–1159. [Google Scholar] [CrossRef]
- Lor, S.; Malecki, R.A. Breeding Ecology and Nesting Habitat Associations of Five Marsh Bird Species in Western New York. Waterbirds 2006, 29, 427–436. [Google Scholar] [CrossRef]
- Lodge, K.A.; Tyler, A.C. Divergent impact of grazing on plant communities of created wetlands with varying hydrology and antecedent land use. Wetl. Ecol. Manag. 2020, 28, 797–813. [Google Scholar] [CrossRef]
- Isola, C.R.; Colwell, M.A.; Taft, O.W.; Safran, R.J. Interspecific Differences in Habitat Use of Shorebirds and Waterfowl Foraging in Managed Wetlands of California’s San Joaquin Valley. Waterbirds 2000, 23, 196–203. [Google Scholar]
- Ankney, C.D. An embarrassment of riches: Too many geese. J. Wildl. Manag. 1996, 60, 217–223. [Google Scholar] [CrossRef]
- Lauridsen, T.L.; Jeppesen, E.; Andersen, F.Ø. Colonization of submerged macrophytes in shallow fish manipulated Lake Væng: Impact of sediment composition and waterfowl grazing. Aquat. Bot. 1993, 46, 1–15. [Google Scholar] [CrossRef]
- Smith III, T.J.; Odum, W.E. The effects of grazing by snow geese on coastal salt marshes. Ecology 1981, 62, 98–106. [Google Scholar] [CrossRef]
- Silliman, B.R.; Zieman, J.C. Top-down control of Spartina alterniflora production by periwinkle grazing in a Virginia salt marsh. Ecology 2001, 82, 2830–2845. [Google Scholar] [CrossRef]
- Silliman, B.R.; Bertness, M.D. A trophic cascade regulates salt marsh primary production. Proc. Natl. Acad. Sci. USA 2002, 99, 10500–10505. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, D.S.; Jefferies, R. A positive feedback: Herbivory, plant growth, salinity, and the desertification of an Arctic salt-marsh. J. Ecol. 1996, 84, 31–42. [Google Scholar] [CrossRef]
- Smith, L.M.; Kadlec, J.A. Fire and herbivory in a Great Salt Lake marsh. Ecology 1985, 66, 259–265. [Google Scholar] [CrossRef]
- Jefferies, R.L.; Jano, A.P.; Abraham, K.F. A biotic agent promotes large-scale catastrophic change in the coastal marshes of Hudson Bay. J. Ecol. 2006, 94, 234–242. [Google Scholar] [CrossRef]
- Jefferies, R.L.; Rockwell, R.F. Foraging geese, vegetation loss and soil degradation in an Arctic salt marsh. Appl. Veg. Sci. 2002, 5, 7–16. [Google Scholar] [CrossRef]
- Ström, L.; Mastepanov, M.; Christensen, T.R. Species-specific Effects of Vascular Plants on Carbon Turnover and Methane Emissions from Wetlands. Biogeochemistry 2005, 75, 65–82. [Google Scholar] [CrossRef]
- Dingemans, B.J.J.; Bakker, E.S.; Bodelier, P.L.E. Aquatic herbivores facilitate the emission of methane from wetlands. Ecology 2011, 92, 1166–1173. [Google Scholar] [CrossRef]
- Winton, R.S.; Richardson, C.J. Top-down control of methane emission and nitrogen cycling by waterfowl. Ecology 2017, 98, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Lubchenco, J. Littornia and Fucus: Effects of Herbivores, Substratum Heterogeneity, and Plant Escapes During Succession. Ecology 1983, 64, 1116–1123. [Google Scholar] [CrossRef]
- Elaine Evers, D.; Sasser, C.E.; Gosselink, J.G.; Fuller, D.A.; Visser, J.M. The impact of vertebrate herbivores on wetland vegetation in Atchafalaya Bay, Louisiana. Estuaries 1998, 21, 1–13. [Google Scholar] [CrossRef]
- Kennedy, M.A.; Heck, K.L.; Michot, T.C. Impacts of wintering redhead ducks (Aythya americana) on seagrasses in the northern Gulf of Mexico. J. Exp. Mar. Biol. Ecol. 2018, 506, 42–48. [Google Scholar] [CrossRef]
- Bagchi, S.; Ritchie, M.E. Introduced grazers can restrict potential soil carbon sequestration through impacts on plant community composition. Ecol. Lett. 2010, 13, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Speed, J.D.; Woodin, S.; Tømmervik, H.; Van der Wal, R. Extrapolating herbivore-induced carbon loss across an arctic landscape. Polar Biol. 2010, 33, 789–797. [Google Scholar] [CrossRef]
- Sjögersten, S.; van der Wal, R.; Loonen, M.J.; Woodin, S.J. Recovery of ecosystem carbon fluxes and storage from herbivory. Biogeochemistry 2011, 106, 357–370. [Google Scholar] [CrossRef] [Green Version]
- Hirota, M.; Tang, Y.; Hu, Q.; Kato, T.; Hirata, S.; Mo, W.; Cao, G.; Mariko, S. The potential importance of grazing to the fluxes of carbon dioxide and methane in an alpine wetland on the Qinghai-Tibetan Plateau. Atmos. Environ. 2005, 39, 5255–5259. [Google Scholar] [CrossRef]
- Wang, Z.; Yuan, X.; Wang, D.; Zhang, Y.; Zhong, Z.; Guo, Q.; Feng, C. Large herbivores influence plant litter decomposition by altering soil properties and plant quality in a meadow steppe. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Colmer, T. Long-distance transport of gases in plants: A perspective on internal aeration and radial oxygen loss from roots. Plant Cell Environ. 2003, 26, 17–36. [Google Scholar] [CrossRef] [Green Version]
- Wolf, A.A.; Drake, B.G.; Erickson, J.E.; Megonigal, J.P. An oxygen-mediated positive feedback between elevated carbon dioxide and soil organic matter decomposition in a simulated anaerobic wetland. Glob. Chang. Biol. 2007, 13, 2036–2044. [Google Scholar] [CrossRef]
- Lodge, K.A. Hydrology, Nutrient Availability, and Herbivory Interacting to Control Ecosystem Functions and Services in Created Emergent Freshwater Wetlands. Master’s Thesis, Rochester Institute of Technology, ProQuest Dissertations Publishing, Rochester, NY, USA, 2017. [Google Scholar]
- Bakker, J.P. The impact of grazing on plant communities, plant populations and soil conditions on salt marshes. Vegetatio 1985, 62, 391–398. [Google Scholar] [CrossRef]
- Koh, H.S.; Ochs, C.A.; Yu, K. Hydrologic gradient and vegetation controls on CH4 and CO2 fluxes in a spring-fed forested wetland. Hydrobiologia 2009, 630, 271–286. [Google Scholar] [CrossRef]
- Brinson, M.M.; Lugo, A.E.; Brown, S. Primary Productivity, Decomposition and Consumer Activity in Freshwater Wetlands. Annu. Rev. Ecol. Syst. 1981, 12, 123–161. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Z.X.; Zhang, X.Y.; Zhu, S.X.; Ge, Y.; Chang, S.X.; Zhang, C.B.; Huang, C.C.; Chang, J. Plant Species Richness Increased Belowground Plant Biomass and Substrate Nitrogen Removal in a Constructed Wetland. CLEAN Soil Air Water 2013, 41, 657–664. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Q.; Li, M.; Liu, F.; Li, W. Does the different photosynthetic pathway of plants affect soil respiration in a subtropical wetland? Ecol. Evol. 2016, 6, 8010–8017. [Google Scholar] [CrossRef] [PubMed]
- Deghi, G.S.; Ewel, K.C.; Mitsch, W.J. Effects of Sewage Effluent Application on Litter Fall and Litter Decomposition in Cypress Swamps. J. Appl. Ecol. 1980, 17, 397–408. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Currie, W.S.; Rastetter, E.B.; Parton, W.J.; Harmon, M.E. Climate and litter quality controls on decomposition: An analysis of modeling approaches. Glob. Biogeochem. Cycles 1999, 13, 575–589. [Google Scholar] [CrossRef] [Green Version]
- Carroll, P.; Crill, P. Carbon balance of a temperate poor fen. Glob. Biogeochem. Cycles 1997, 11, 349–356. [Google Scholar] [CrossRef]
- Hossler, K.; Bouchard, V.; Fennessy, M.S.; Frey, S.D.; Anemaet, E.; Herbert, E. No-net-loss not met for nutrient function in freshwater marshes: Recommendations for wetland mitigation policies. Ecosphere 2011, 2, 1–36. [Google Scholar] [CrossRef]
- Veen, G.; Sarneel, J.M.; Ravensbergen, L.; Huig, N.; van Paassen, J.; Rip, W.; Bakker, E.S. Aquatic grazers reduce the establishment and growth of riparian plants along an environmental gradient. Freshw. Biol. 2013, 58, 1794–1803. [Google Scholar] [CrossRef]
- Mulder, C.P.H.; Ruess, R.W. Effects of Herbivory on Arrowgrass: Interactions Between Geese, Neighboring Plants, and Abiotic Factors. Ecol. Monogr. 1998, 68, 275–293. [Google Scholar] [CrossRef]
- Myers, R.S.; Shaffer, G.P.; Llewellyn, D.W. Baldcypress (Taxodium distichum (L.) Rich.) restoration in southeast Louisiana: The relative effects of herbivory, flooding, competition, and macronutrients. Wetlands 1995, 15, 141–148. [Google Scholar] [CrossRef]
- Goranson, C.E.; Ho, C.K.; Pennings, S.C. Environmental gradients and herbivore feeding preferences in coastal salt marshes. Oecologia 2004, 140, 591–600. [Google Scholar] [CrossRef]
- Buij, R.; Melman, T.C.; Loonen, M.J.; Fox, A.D. Balancing ecosystem function, services and disservices resulting from expanding goose populations. Ambio 2017, 46, 301–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, F.E.; Bergamaschi, B.; Sturtevant, C.; Knox, S.; Hastings, L.; Windham-Myers, L.; Detto, M.; Hestir, E.L.; Drexler, J.; Miller, R.L.; et al. Variation of energy and carbon fluxes from a restored temperate freshwater wetland and implications for carbon market verification protocols. J. Geophys. Res. Biogeosci. 2016, 121, 777–795. [Google Scholar] [CrossRef]
- Altor, A.E.; Mitsch, W.J. Pulsing hydrology, methane emissions and carbon dioxide fluxes in created marshes: A 2-year ecosystem study. Wetlands 2008, 28, 423–438. [Google Scholar] [CrossRef]
- Valach, A.C.; Kasak, K.; Hemes, K.S.; Anthony, T.L.; Dronova, I.; Taddeo, S.; Silver, W.L.; Szutu, D.; Verfaillie, J.; Baldocchi, D.D. Productive wetlands restored for carbon sequestration quickly become net CO2 sinks with site-level factors driving uptake variability. PLoS ONE 2021, 16, e0248398. [Google Scholar] [CrossRef] [PubMed]
- Mueller, P.; Jensen, K.; Megonigal, J.P. Plants mediate soil organic matter decomposition in response to sea level rise. Glob. Chang. Biol. 2016, 22, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Blagodatskaya, E.; Kuzyakov, Y. Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: Critical review. Biol. Fertil. Soils 2008, 45, 115–131. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Wardle, D.A.; Yeates, G.W. Linking above-ground and below-ground interactions: How plant responses to foliar herbivory influence soil organisms. Soil Biol. Biochem. 1998, 30, 1867–1878. [Google Scholar] [CrossRef]
- He, M.; Zhou, G.; Yuan, T.; van Groenigen, K.J.; Shao, J.; Zhou, X. Grazing intensity significantly changes the C: N: P stoichiometry in grassland ecosystems. Glob. Ecol. Biogeogr. 2020, 29, 355–369. [Google Scholar] [CrossRef]
- Tanentzap, A.J.; Coomes, D.A. Carbon storage in terrestrial ecosystems: Do browsing and grazing herbivores matter? Biol. Rev. 2012, 87, 72–94. [Google Scholar] [CrossRef] [PubMed]
- Sasser, C.E.; Holm, G.O.; Evers-Hebert, E.; Shaffer, G.P. The nutria in Louisiana: A current and historical perspective. In Mississippi Delta Restoration; Springer: Berlin, Germany, 2018; pp. 39–60. [Google Scholar]
- Yu, O.; Chmura, G. Soil carbon may be maintained under grazing in a St Lawrence Estuary tidal marsh. Environ. Conserv. 2009, 36, 312–320. [Google Scholar] [CrossRef]
- Van Der WAL, R.; Sjögersten, S.; Woodin, S.J.; Cooper, E.J.; Jónsdóttir, I.S.; Kuijper, D.; Fox, T.A.; Huiskes, A. Spring feeding by pink-footed geese reduces carbon stocks and sink strength in tundra ecosystems. Glob. Chang. Biol. 2007, 13, 539–545. [Google Scholar] [CrossRef]
- Frank, A.B.; Tanaka, D.L.; Hofmann, L.; Follett, R.F. Soil carbon and nitrogen of Northern Great Plains grasslands as influenced by long-term grazing. Rangel. Ecol. Manag. J. Range Manag. Arch. 1995, 48, 470–474. [Google Scholar] [CrossRef]
- Cui, X.; Wang, Y.; Niu, H.; Wu, J.; Wang, S.; Schnug, E.; Rogasik, J.; Fleckenstein, J.; Tang, Y. Effect of long-term grazing on soil organic carbon content in semiarid steppes in Inner Mongolia. Ecol. Res. 2005, 20, 519–527. [Google Scholar] [CrossRef]
- Elschot, K.; Bakker, J.P.; Temmerman, S.; van de Koppel, J.; Bouma, T.J. Ecosystem engineering by large grazers enhances carbon stocks in a tidal salt marsh. Mar. Ecol. Prog. Ser. 2015, 537, 9–21. [Google Scholar] [CrossRef]
- Clermont, J.; Réale, D.; Giroux, J.F. Plasticity in laying dates of Canada Geese in response to spring phenology. Ibis 2018, 160, 597–607. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Date | Treatment | Dt × Tr | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| Aboveground biomass 2017 | F = 20.5 | p < 0.0001 | F = 17.0 | p < 0.0001 | F = 2.6 | p = 0.08 |
| Aboveground biomass 2018 | F = 34.2 | p< 0.0001 | F = 35.7 | p< 0.0001 | F = 4.9 | p= 0.003 |
| GPP 2017 | F = 23.2 | p< 0.0001 | F = 11.6 | p= 0.002 | F = 6.5 | p= 0.02 |
| GPP 2018 | F = 7.7 | p= 0.01 | F = 4.0 | p = 0.06 | F = 3.9 | p = 0.06 |
| ER 2017 | F = 7.7 | p= 0.01 | F = 6.4 | p= 0.02 | F = 0.01 | p = 0.9 |
| ER 2018 | F = 0.003 | p = 0.95 | F = 5.1 | p= 0.03 | F = 3.1 | p = 0.09 |
| NEE 2017 | F = 17.4 | p= 0.0003 | F = 7.5 | p= 0.01 | F = 7.5 | p= 0.01 |
| NEE 2018 | F = 10.4 | p= 0.003 | F = 2.7 | p = 0.1 | F = 3.6 | p = 0.07 |
| Variable | %C | %N | C:N |
|---|---|---|---|
| Vegetation | |||
| S. latifolia | 42.0 ± 0.01 | 2.0 ± 0.05 | 24.9 ± 0.6 |
| N. odorata | 41.9 ± 0.23 | 2.5 ± 0.01 | 19.8 ± 0.2 |
| T. latifolia | 44.4 ± 0.14 | 2.1 ± 0.08 | 24.6 ± 0.8 |
| P. cordata | 41.2 ±0.07 | 1.3 ± 0.02 | 37.3 ± 0.6 |
| L. oryzoides | 43.1 ± 0.03 | 1.4 ± 0.13 | 35.9 ± 3.3 |
| Belowground biomass | |||
| Caged | 35.9 ± 1.66 | 1.6 ± 0.07 | 26.0 ± 1.3 |
| Uncaged | 35.5 ± 2.00 | 1.6 ± 0.14 | 25.7 ± 1.7 |
| Soil | |||
| Caged | 6.5 ± 0.33 | 0.5 ± 0.13 | 19.7 ± 0.7 |
| Uncaged | 5.1 ± 0.36 | 0.3 ± 0.02 | 20.8 ± 0.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spangler, D.M.; Tyler, A.C.; McCalley, C.K. Effects of Grazer Exclusion on Carbon Cycling in Created Freshwater Wetlands. Land 2021, 10, 805. https://doi.org/10.3390/land10080805
Spangler DM, Tyler AC, McCalley CK. Effects of Grazer Exclusion on Carbon Cycling in Created Freshwater Wetlands. Land. 2021; 10(8):805. https://doi.org/10.3390/land10080805
Chicago/Turabian StyleSpangler, Delanie M., Anna Christina Tyler, and Carmody K. McCalley. 2021. "Effects of Grazer Exclusion on Carbon Cycling in Created Freshwater Wetlands" Land 10, no. 8: 805. https://doi.org/10.3390/land10080805
APA StyleSpangler, D. M., Tyler, A. C., & McCalley, C. K. (2021). Effects of Grazer Exclusion on Carbon Cycling in Created Freshwater Wetlands. Land, 10(8), 805. https://doi.org/10.3390/land10080805