
Microbial Respiration and Enzyme Activity Downstream from a Phosphorus Source in the Everglades, Florida, USA



Abstract
:1. Introduction
2. Materials and Methods
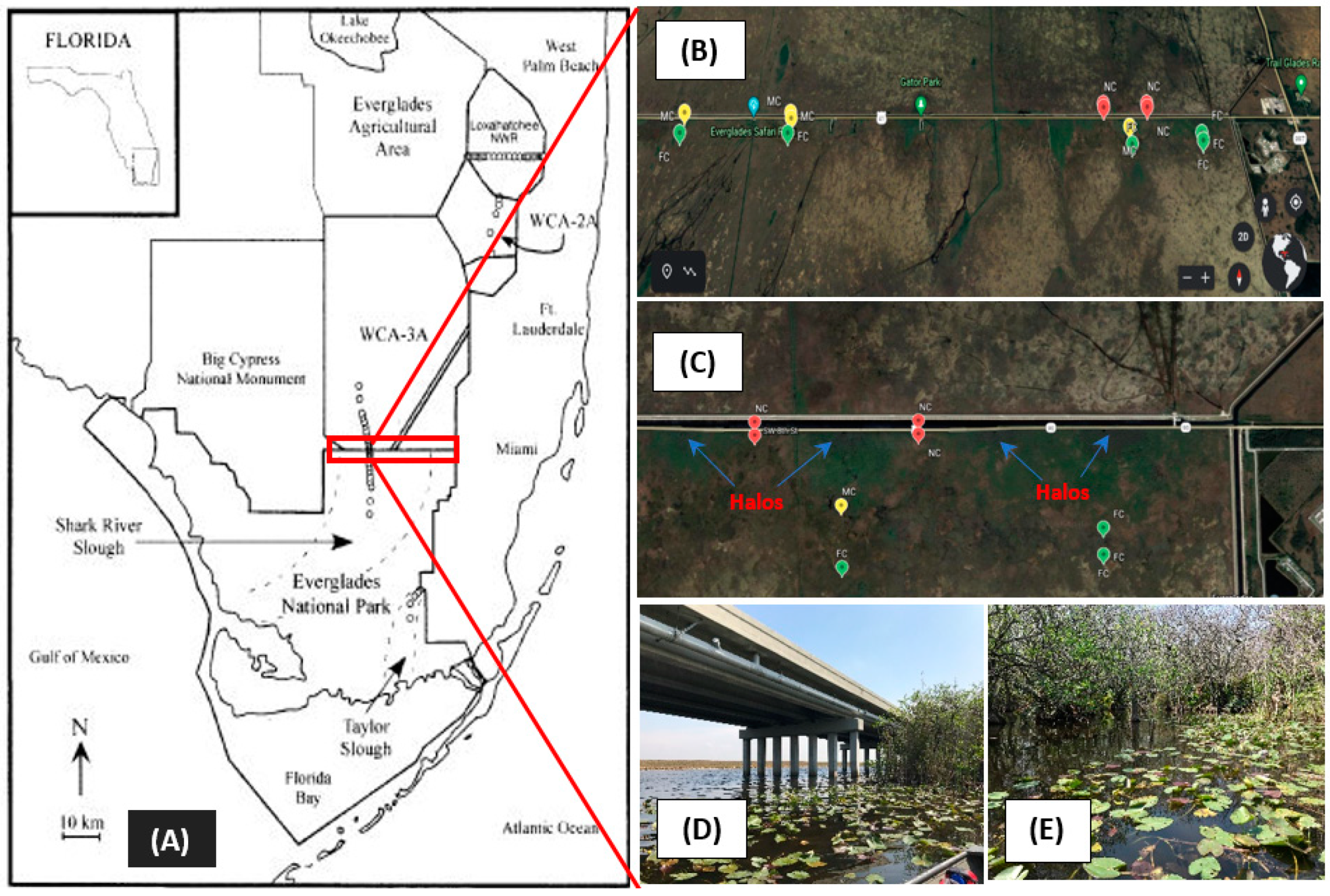
2.1. Site Location and Sample Collection
2.2. Laboratory Analyses
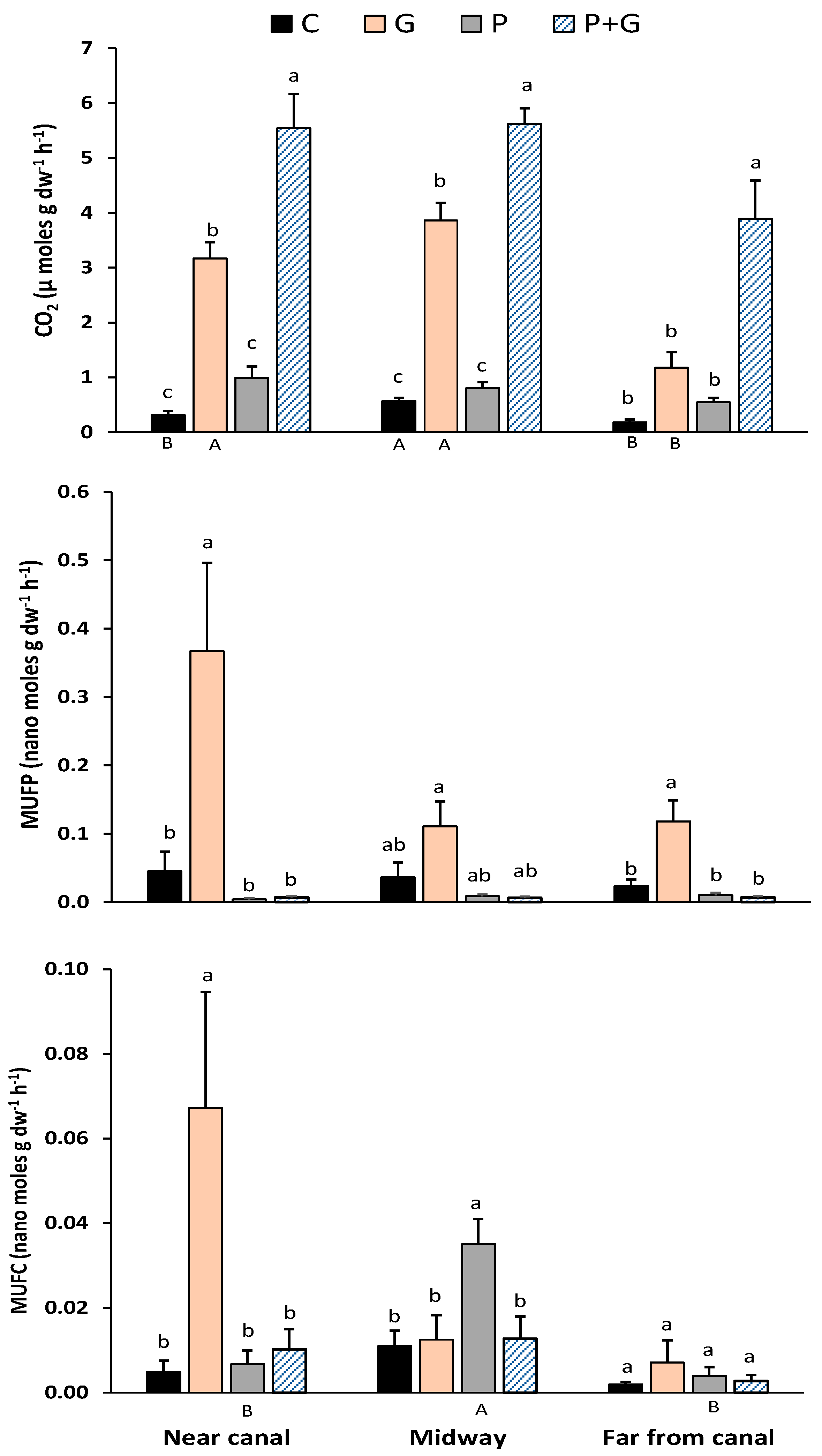
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lal, R. Carbon sequestration. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 815–830. [Google Scholar] [CrossRef]
- Nahlik, A.M.; Fennessy, M.S. Carbon storage in US wetlands. Nat. Commun. 2016, 7, 13835. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, M.N.; Winding, A.; Binnerup, S. Microorganisms as Indicators of Soil Health. 2002. Available online: https://www.dmu.dk/1_Viden/2_Publikationer/3_Fagrapporter/rapporter/FR388.pdf (accessed on 30 June 2021).
- Cabugao, K.G.; Timm, C.M.; Carrell, A.A.; Childs, J.; Lu, T.Y.S.; Pelletier, D.A.; Weston, D.J.; Norby, R.J. Root and rhizosphere bacterial phosphatase activity varies with tree species and soil phosphorus availability in Puerto Rico tropical forest. Front. Plant. Sci. 2017, 8, 1834. [Google Scholar] [CrossRef] [Green Version]
- Margalef, O.; Sardans, J.; Fernández-Martínez, M.; Molowny-Horas, R.; Janssens, I.A.; Ciais, P.; Goll, D.; Richter, A.; Obersteiner, M.; Asensio, D.; et al. Global patterns of phosphatase activity in natural soils. Sci. Rep. 2017, 7, 1337. [Google Scholar] [CrossRef] [Green Version]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Pisani, O.; Scinto, L.J.; Munyon, J.W.; Jaffé, R. The respiration of flocculent detrital organic matter (floc) is driven by phosphorus limitation and substrate quality in a subtropical wetland. Geoderma 2015, 241, 272–278. [Google Scholar] [CrossRef]
- Childers, D.L.; Doren, R.F.; Jones, R.; Noe, G.B.; Rugge, M.; Scinto, L.J. Decadal change in vegetation and soil phosphorus pattern across the Everglades landscape. J. Environ. Qual. 2003, 32, 344–362. [Google Scholar] [CrossRef] [PubMed]
- Gaiser, E. Periphyton as an indicator of restoration in the Florida Everglades. Ecol. Indic. 2009, 9, S37–S45. [Google Scholar] [CrossRef]
- Malone, S.L.; Starr, G.; Staudhammer, C.L.; Ryan, M.G. Effects of simulated drought on the carbon balance of Everglades short-hydroperiod marsh. Glob. Chang. Biol. 2013, 19, 2511–2523. [Google Scholar] [CrossRef]
- Brown, P.; Wright, A.L. The Role of Periphyton in the Everglades. (2013, February). Available online: http://edis.ifas.ufl.edu/pdffiles/SS/SS52200.pdf (accessed on 30 June 2021).
- Noe, G.B.; Childers, D.L.; Jones, R.D. Phosphorus biogeochemistry and the impact of phosphorus enrichment: Why is the Everglades so unique? Ecosystems 2001, 4, 603–624. [Google Scholar] [CrossRef]
- Dattamudi, S.; Wang, J.J.; Dodla, S.K.; Viator, H.P.; DeLaune, R.; Hiscox, A.; Darapuneni, M.; Jeong, C.; Colyer, P. Greenhouse gas emissions as influenced by nitrogen fertilization and harvest residue management in sugarcane production. Agrosyst. Geosci. Environ. 2019, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis: Part 3 Chemical Methods; Sparks, D.L., Ed.; SSSA Book Series 5; SSSA: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Solorzano, L.; Sharp, J.H. Determination of total dissolved phosphorus and particulate phosphorus in natural waters. Limnol. Oceanogr. 1980, 25, 754–758. [Google Scholar] [CrossRef]
- O’Dell, J.W. Method 365.1—Determination of Phosphorus by Semi-Automated Colorimetry; USEPA, Environmental Monitoring Systems Laboratory, Office of Research and Development: Cincinnati, OH, USA, 1993. [Google Scholar]
- Rivero, R.G.; Grunwald, S.; Osborne, T.Z.; Reddy, K.R.; Newman, S. Characterization of the spatial distribution of soil properties in Water Conservation Area 2A, Everglades, Florida. Soil Sci. 2007, 172, 149–166. [Google Scholar] [CrossRef] [Green Version]
- Steinmuller, H.E.; Stoffella, S.L.; Vidales, R.; Ross, M.S.; Dattamudi, S.; Scinto, L.J. Characterizing hydrologic effects on soil physicochemical variation within tree islands and marshes in the coastal florida everglades. Soil Sci. Soc. Am. J. 2021. [Google Scholar] [CrossRef]
- Irick, D.L.; Li, Y.C.; Inglett, P.W.; Harris, W.G.; Gu, B.; Ross, M.S.; Wright, A.L.; Migliaccio, K.W. Characteristics of soil phosphorus in tree island hardwood hammocks of the southern Florida Everglades. Soil Sci. Soc. Am. J. 2013, 77, 1048–1056. [Google Scholar] [CrossRef]
- Schade-Poole, K.; Möller, G. Impact and mitigation of nutrient pollution and overland water flow change on the Florida Everglades, USA. Sustainability 2016, 8, 940. [Google Scholar] [CrossRef] [Green Version]
- DeBusk, W.F.; Reddy, K.R. Nutrient and hydrology effects on soil respiration in a northern Everglades marsh. J. Environ. Qual. 2003, 32, 702–710. [Google Scholar] [CrossRef]
- DeBusk, W.F.; Reddy, K.R. Litter decomposition and nutrient dynamics in a phosphorus enriched everglades marsh. Biogeochemistry 2005, 75, 217–240. [Google Scholar] [CrossRef]
- Pisani, O.; Gao, M.; Maie, N.; Miyoshi, T.; Childers, D.L.; Jaffé, R. Compositional aspects of herbaceous litter decomposition in the freshwater marshes of the Florida Everglades. Plant. Soil 2018, 423, 87–98. [Google Scholar] [CrossRef]
- Amador, J.A.; Jones, R.D. Response of carbon mineralization to combined changes in soil moisture and carbon-phosphorus ratio in a low phosphorus histosol. Soil Sci. 1997, 162, 275–282. [Google Scholar] [CrossRef]
- Wetzel, P.R.; Van Der Valk, A.G.; Newman, S.; Coronado, C.A.; Troxler-Gann, T.G.; Childers, D.L.; Orem, W.H.; Sklar, F.H. Heterogeneity of phosphorus distribution in a patterned landscape, the Florida Everglades. Plant. Ecol. 2009, 200, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Medvedeff, C.A.; Inglett, K.S.; Inglett, P.W. Patterns and controls of anaerobic soil respiration and methanogenesis following extreme restoration of calcareous subtropical wetlands. Geoderma 2015, 245, 74–82. [Google Scholar] [CrossRef]
- Lane, R.R.; Mack, S.K.; Day, J.W.; DeLaune, R.D.; Madison, M.J.; Precht, P.R. Fate of soil organic carbon during wetland loss. Wetlands 2016, 36, 1167–1181. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sites | n | Water Depth | BD | SOM | TP | TN | TC |
|---|---|---|---|---|---|---|---|
| cm | g dw cm−3 | % | ug g−1 dw | mg g−1 dw | |||
| Near canal | 28 | 19 ± 8 a | 0.140 ± 0.035 a | 54 ± 19 b | 1143 ± 273 a | 2.2 ± 0.9 b | 32 ± 6 c |
| Midway | 24 | 12 ± 4 a | 0.129 ± 0.029 a | 84 ± 17 a | 712 ± 151 b | 3.4 ± 0.3 a | 46 ± 8 a |
| Far from canal | 24 | 8 ± 4 a | 0.149 ± 0.052 a | 67 ± 11 b | 467 ± 87 c | 2.8 ± 0.4 a | 40 ± 9 b |
| Parameters | TP | Soil Moisture | TN | TC | SOM |
|---|---|---|---|---|---|
| BD | −0.33 | −0.60 * | −0.35 | −0.50 | −0.51 |
| TP | 0.25 | −0.28 | −0.29 | −0.21 | |
| Soil moisture | 0.39 * | 0.44 * | 0.53 * | ||
| TN | 0.95 * | 0.95 * | |||
| TC | 0.99 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dattamudi, S.; Chanda, S.; Scinto, L.J. Microbial Respiration and Enzyme Activity Downstream from a Phosphorus Source in the Everglades, Florida, USA. Land 2021, 10, 696. https://doi.org/10.3390/land10070696
Dattamudi S, Chanda S, Scinto LJ. Microbial Respiration and Enzyme Activity Downstream from a Phosphorus Source in the Everglades, Florida, USA. Land. 2021; 10(7):696. https://doi.org/10.3390/land10070696
Chicago/Turabian StyleDattamudi, Sanku, Saoli Chanda, and Leonard J. Scinto. 2021. "Microbial Respiration and Enzyme Activity Downstream from a Phosphorus Source in the Everglades, Florida, USA" Land 10, no. 7: 696. https://doi.org/10.3390/land10070696
APA StyleDattamudi, S., Chanda, S., & Scinto, L. J. (2021). Microbial Respiration and Enzyme Activity Downstream from a Phosphorus Source in the Everglades, Florida, USA. Land, 10(7), 696. https://doi.org/10.3390/land10070696