
Topology, Vegetation and Stratigraphy of Far Eastern Aapa Mires (Khabarovsk Region, Russia)


Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Area
2.2. Climate
2.3. Study Sites
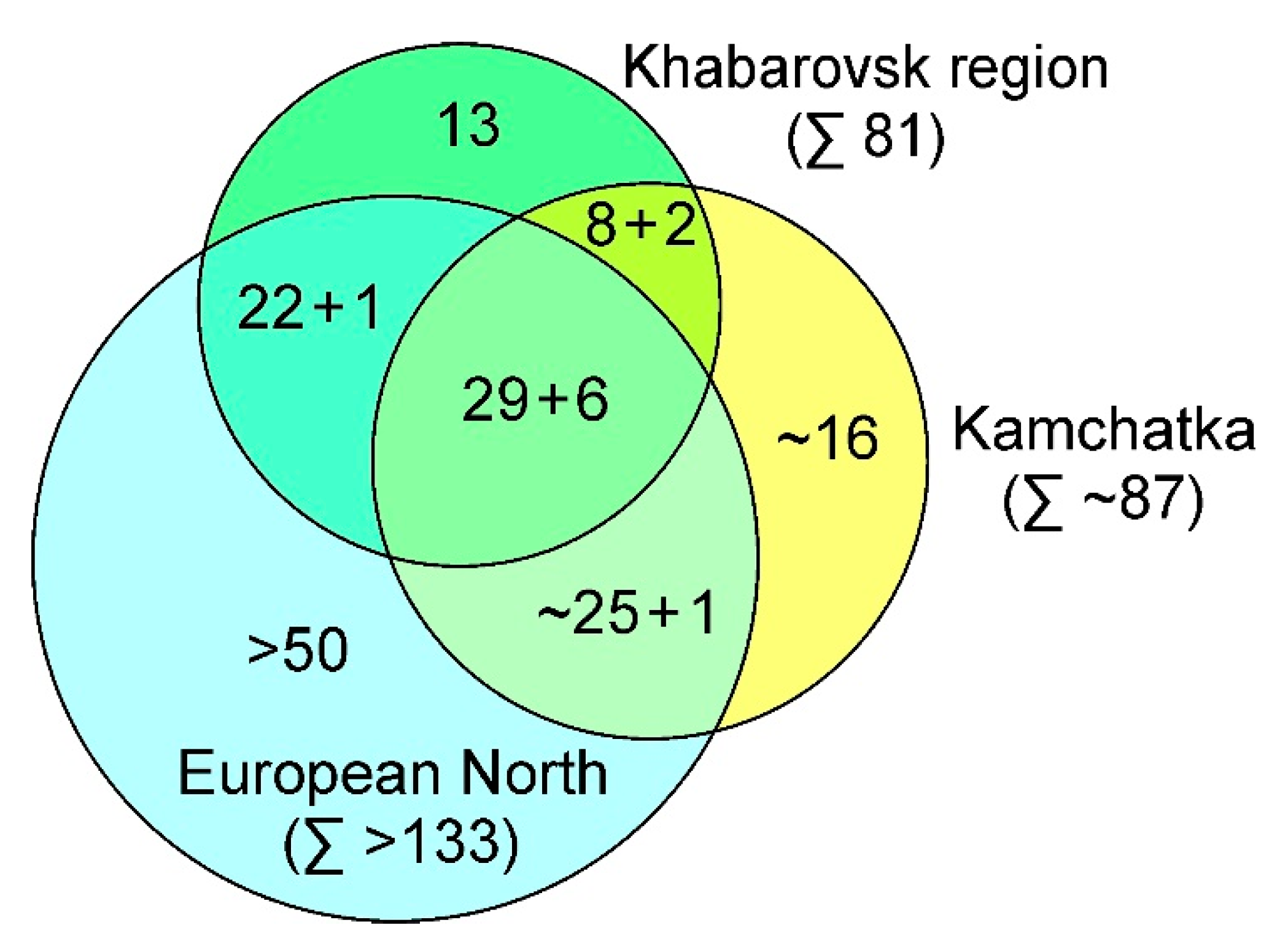
3. Results
3.1. Topology
3.2. Vegetation
3.2.1. Strings
3.2.2. Lawns
3.2.3. Flarks
3.3. Peat Deposit Stratigraphy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Cajander, A.K. Studien über die Moore Finnlands. Acta For. Fenn. 1913, 2, 7530. [Google Scholar] [CrossRef]
- Ruuhijärvi, R. Über die regionale Einteilung der nordfinnischen Moore. Ann. Bot. Soc. Vanamo 1960, 31, 1–360. (In German) [Google Scholar]
- Eurola, S.; Kaakinen, E. Suotyyppiopas. [Guide to Mire Site Types]; WSOY: Porvoo, Finland, 1978; pp. 1–87. [Google Scholar]
- Eurola, S.; Huttunen, A.; Kukko-oja, K. Suokasvillisuusopas. [Mire vegetation guide]. In Oulanka Reports 14; Oulanka Biological Station: Kuusamo, Finland; University of Oulu: Oulu, Finland, 1995; pp. 1–85. [Google Scholar]
- Laitinen, J.; Rehell, S.; Huttunen, A. Vegetation-related hydrotopographic and hydrologic classification for aapa mires (Hirvisuo, Finland). Ann. Bot. Fenn. 2005, 42, 107–121. [Google Scholar]
- Laitinen, J.; Rehell, S.; Huttunen, A.; Tahvanainen, T.; Heikkilä, R.; Lindholm, T. Mire systems of Finland with special view to aapa mires and their water flow pattern. Suo 2007, 58, 1–26. [Google Scholar]
- Yurkovskaya, T.K. Peatlands of the taiga north-east of European Russia. Russ. J. Bot. 2021, 106, 211–228. (In Russian) [Google Scholar]
- Yurkovskaya, T.K. About the geography of mires of Russia. In XI Meeting in Memoriam of E. A. Galkina, Proceedings of the Scientific Conference, Saint-Petersburg, Russia, 21 April 2021; Komarov Botanical Institute RAS: Saint Petersburg, Russia, 2021; pp. 3–4. [Google Scholar]
- Yurkovskaya, T.K. Geography and cartography of mire vegetation of the European Russia and neighboring territories. In Transactions of Komarov Botanical Institute; Proceeding of Komarov Botanical Institute RAS: Saint Petersburg, Russia, 1992; Volume 4, pp. 1–256. (In Russian) [Google Scholar]
- Yurkovskaya, T.K. Ribbed fens. In Mires of Northern Europe: Biodiversity, Dynamics, Management: Abstracts of International Symposium, Petrozavodsk, Russia, 2–5 September 2015; Kuznetsov, O.L., Znamenskiy, S.R., Kantserova, L.V., Eds.; Karelian Research Centre of RAS: Petrozavodsk, Russia, 2015; pp. 103–104. [Google Scholar]
- Moen, A.; Joosten, H.; Tanneberger, F. Mire diversity in Europe: Mire regionality. In Mires and Peatlands of Europe: Status, Distribution and Conservation; Joosten, H., Tanneberger, F., Moen, A., Eds.; Schweizerbart Science Publishers: Stuttgart, Germany, 2017; pp. 97–149. [Google Scholar]
- Kutenkov, S.A.; Philippov, D.A. Aapa mire on the southern limit: A case study of the Vologda Region (north-western Russia). Mires Peat 2019, 24, 1–20. [Google Scholar] [CrossRef]
- Ruuhijärvi, R. The Finnish mire types and their regional distribution. In Mires: Swamp, Bog, Fen and Moor. Ecosystems of the World; Gore, A.J.P., Ed.; Elsevier Publishing: Amsterdam, The Netherlands, 1983; pp. 47–67. [Google Scholar]
- Zinserling Yu., D. Plant cover geography of the north-west of the European part of the USSR. In Trudy Geomorfologicheskogo Instituta. Ser. Fiziko-Geograficheskaya; Academy of Sciences of the USSR: Leningrad, Russia, 1934; Volume 4, pp. 1–377. (In Russian) [Google Scholar]
- Sirin, A.; Minayeva, T.; Yurkovskaya, T.; Kuznetsov, O.; Smagin, V.; Fedotov, Y. Russian Federation (European Part). In Mires and Peatlands of Europe: Status, Distribution and Conservation; Joosten, H., Tanneberger, F., Moen, A., Eds.; Schweizerbart Science Publishers: Stuttgart, Germany, 2017; pp. 589–616. [Google Scholar]
- Liss, O.; Abramova, L.; Avetov, N.; Berezina, N.; Inisheva, L.; Kurnishkova, T.; Sluka, Z.; Tolpysheva, T.; Shvedchikova, N. Mire Systems of West Siberia and Its Nature Conservation Importance; Grif and Co: Tula, Russia, 2001; pp. 1–584. (In Russian) [Google Scholar]
- Terentieva, I.E.; Glagolev, M.V.; Lapshina, E.D.; Sabrekov, A.F.; Maksyutov, S. Mapping of West Siberian taiga wetland complexes using Landsat imagery: Implications for methane emissions. Biogeosciences 2016, 13, 4615–4626. [Google Scholar] [CrossRef] [Green Version]
- Lapshina, E.D. Zonal-geographical mire types of West Siberia. In XI Meeting in Memoriam of E. A. Galkina, Proceedings of the Scientific Conference, Saint-Petersburg, Russia, 21 April 2021; Komarov Botanical Institute RAS: Saint Petersburg, Russia, 2021; pp. 31–34. [Google Scholar]
- Lashchinsky, N.N.; Pisarenko, O.Y. Ribbon mires in Kija-Yaya interfluve (Western Siberia). Plant Life Asian Russ. 2010, 1, 42–48. (In Russian) [Google Scholar]
- Neshatayev, V.Y.; Neshataeva, V.Y.; Kirichenko, V.E. Mire types of the North of the Koryak Okrug. In IX Meeting in Memoriam of E. A. Galkina, Proceedings of the Scientific Conference, Saint-Petersburg, Russia, 5–7 February 2018; Yurkovskaya, T.K., Ed.; Publishing House “LETI”: Saint-Petersburg, Russia, 2018; pp. 168–170. (In Russian) [Google Scholar]
- Neshatayeva, V.Y.; Neshataev, V.Y. The geographical distribution of mires of the Koryak Okrug (Kamchatka Area) In X Meeting in Memoriam of E. A. Galkina, Proceedings of the Scientific Conference, Saint-Petersburg, Russia, 4–6 February 2019; Publishing House “LETI”: Saint-Petersburg, Russia, 2019; pp. 144–148. (In Russian) [Google Scholar]
- Botch, M.S. North Kamchatka Province of aapa-palsa mires. In Conference Abstracts of the VII Delegate Congress of the All-Russian Botanical Society, Donetsk, Russia, 11–14 May 1983; Vasilevich, V., Ed.; Nauka: Leningrad, Russia; pp. 129–130. (In Russian)
- Neshatayeva, V.Y.; Neshataev, V.Y. Mire vegetation of the Southern Kamchatka state nature reserve. Veg. Russ. 2001, 2, 58–70. (In Russian) [Google Scholar] [CrossRef]
- Neshataev, V.Y.; Neshataeva, V.Y.; Khabarova, N.N. Vegetation of the mires. In Vegetation of the Kronotsky Nature Reserve (Eastern Kamchatka); Neshataev, Y.N., Neshataeva, V.Y., Naumenko, A.T., Eds.; Proceeding of Komarov Botanical Institute RAS: Saint-Petersburg, Russia, 1994; pp. 167–196. (In Russian) [Google Scholar]
- Neshatayev, V.Y.; Neshataeva, V.Y. Diversity of mires of Kamchatka Krai. In XI Meeting in Memoriam of E. A. Galkina, Proceedings of the Scientific Conference, Saint-Petersburg, Russia, 21 April 2021; Komarov Botanical Institute RAS: Saint Petersburg, Russia, 2021; pp. 35–42. (In Russian) [Google Scholar]
- Neshatayev, V.Y.; Cherniagina, O.A.; Neshatayeva, V.Y.; Korablev, A.P.; Kuzmina, E.Y. Flora and vegetation of the planned salmon reserve territory “Utholok”. In Conservation of Biodiversity of Kamchatka and Adjacent Seas, Proceedings of the VIII International Scientific Conference Dedicated to the 275th Anniversary of the Beginning of the Second Kamchatka Expedition (1732–1733), Petropavlovsk-Kamchatkiy, Russia, 27–28 November 2007; Tokranov, A.M., Ed.; Publishing House Kamchatpress: Petropavlovsk-Kamchatsky, Russia, 2008; pp. 141–166. (In Russian) [Google Scholar]
- Prozorov, Y.S. Natural features of the Amur bogs, their study and the tasks of further research. In Natural Features of the Amur Mires; Science: Novosibirsk, Russia, 1973; pp. 2–42. (In Russian) [Google Scholar]
- Lindberg, G.U. Large planetary fluctuations in ocean level and paleogeography of the Quaternary period. In Basic Problems of Studying the Quaternary Period; Science: Moscow, Russia, 1965; pp. 135–142. [Google Scholar]
- Geibert, W.; Matthiessen, J.; Stimac, I.; Wollenburg, J.; Stein, R. Glacial episodes of a freshwater Arctic Ocean covered by a thick ice shelf. Nature 2021, 590, 97–102. [Google Scholar] [CrossRef]
- Prozorov, Y.S. Correlation of mire formation with fluctuations in the basis of erosion on the Nimelen-Chukchagir lowland. In Questions of the Geography of the Far East. Volume 12. Questions of the Evolution of Landscapes in the South of the Far East; KhCRI FESC of AN SSSR and PB GS USSR: Khabarovsk, Russia, 1973; pp. 208–231. [Google Scholar]
- Prozorov, Y.S. Regularities of Development, Classification and Use of Mire Biogeocenoses; Science: Moscow, Russia, 1985; p. 195. (In Russian) [Google Scholar]
- Tahvanainen, T. Abrupt ombrotrophication of a boreal aapa mire triggered by hydrological disturbance in the catchment. J. Ecol. 2011, 99, 404–415. [Google Scholar] [CrossRef]
- Väliranta, M.; Salojärvi, N.; Vuorsalo, A.; Juutinen, S.; Korhola, A.; Luoto, M.; Tuittila, E.S. Holocene fen–bog transitions, current status in Finland and future perspectives. Holocene 2016, 27, 752–764. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, K.E. Fundamentals of Peatland Hydrology in the Forest Zone; Gydrometeoizdat: Leningrad, Russia, 1957; pp. 1–499. (In Russian) [Google Scholar]
- Ivanov, K.E. Water Movement in Mirelands; Academic Press: London, Britain, 1981; pp. 1–278. [Google Scholar]
- Kolesnikov, B.P. An Outline of the Vegetation of the Far East; Khabarovsk Book Publishing House: Khabarovsk, Russia, 1955; pp. 1–104. [Google Scholar]
- Kryukova, M.V. Vascular Plants of the Lower Amur Region; Dalnauka: Vladivostok, Russia, 2013; pp. 1–354. (In Russian) [Google Scholar]
- Prozorov, Y.S. Mires of the Mari Landscape of the Middle Amur Lowland; Publishing House of the Institute of Forest and Wood of Siberian Branch of AS USSR: Moscow, Russia, 1961; pp. 1–121. (In Russian) [Google Scholar]
- Anisimov, V.M.; Savchenko, I.F.; Chakov, V.V. Mires of the northwestern part of the Middle Amur lowland and their genesis. Quest. Geogr. Far East 1973, 12, 196–207. (In Russian) [Google Scholar]
- Chakov, V.V. Features of the formation of complex ridge-hollow mires in the Middle Amur Lowland. Sov. J. Ecol. 1983, 2, 13–20. [Google Scholar]
- Chemekov, Y.F. The problem of the Quaternary glaciation of the North-East and the Far East of the USSR. In The Problem of the Quaternary Glaciation of the Siberia and the Far East; New seria, Proceedings of the All-Union Scientific Research Geological Institute (VSEGEI); Gosgeoizdat: Leningrad, Russia, 1961; Volume 64. [Google Scholar]
- Gorbarenko, S.A.; Southon, J.R.; Keigwinc, L.D.; Cherepanova, M.V.; Gvozdevaa, I.G. Late Pleistocene–Holocene oceanographic variability in the Okhotsk Sea: Geochemical, lithological and paleontological evidence. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 209, 281–301. [Google Scholar] [CrossRef]
- Chakov, V.V.; Parkhomchuk, E.V.; Zakharchenko, E.N. Dynamics of peat accumulation processes in sections of the periglacial zone of the basin watershed of the Tugur-Nimelen. In Regions of New Development: The Current State of Natural Complexes and Their Protection, Proceedings of International Conference, Khabarovsk, Russia, 5–7 October 2021; IWEP FEB RAS: Khabarovsk, Russia; pp. 99–104. (In Russian)
- Petrov, E.S.; Novorotskiy, P.V.; Lenshin, V.T. The Climate of the Khabarovsk Territory and the Jewish Autonomous Region; Dalnauka: Vladivostok-Khabarovsk, Russia, 2000; pp. 1–174. (In Russian) [Google Scholar]
- Razuvaev, V.N.; Bulygina, O.N.; Korshunova, N.N.; Kleshchenko, L.K.; Kuznetsova, V.N.; Trofimenko, L.T.; Sherstyukov, A.B.; Shvets, N.V.; Davletshin, S.G.; Zvereva, G.N. Climate of Russia: Scientific and Applied Reference Book; All-Russian Research Institute of Hydrometeorological Information—World Data Center: Obninsk, Russia, 2018; Available online: http://meteo.ru/climate/197-nauchno-prikladnoj-spravochnik-klimat-rossii (accessed on 20 October 2021).
- Vitvitsky, G.N. Climate. In Southern Part of the Far East; Science: Moscow, Russia, 1969; p. 419. (In Russian) [Google Scholar]
- Philippov, D.A.; Prokin, A.A.; Przhiboro, A.A. Methods and Methodology of Hydrobiological Study of Mires: Tutorial; Publishing house of Tyumen State University: Tyumen, Russia, 2017; pp. 1–207. (In Russian) [Google Scholar]
- Maksimov, A.I.; Ignatova, E.A. Sphagnum alaskense (Sphagnaceae, Bryophyta), a new species for Russia. Arctoa 2008, 17, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Andrus, R.E.; Janssens, J.A. Sphagnum alaskense, a new species from western North America. Bryologist 2003, 106, 435–438. [Google Scholar] [CrossRef]
- Hassel, K.; Kyrkjeeide, M.; Yousefi, N.; Prestø, T.; Stenøien, H.; Shaw, J.; Flatberg, K. Sphagnum divinum (sp. nov.) and S. medium Limpr. and their relationship to S. magellanicum Brid. J. Bryol. 2018, 40, 197–222. [Google Scholar] [CrossRef]
- Ellis, L.T.; Afonina, O.M.; Czernyadjeva, I.V.; Konoreva, L.A.; Potemkin, A.D.; Kotkova, V.M.; Alataş, M.; Blom, H.H.; Boiko, M.; Cabral, R.A.; et al. New national and regional bryophyte records. J. Bryol. 2020, 42, 281–296. [Google Scholar] [CrossRef]
- Prozorov, Y.S. Mires of the Lower Amur Lowlands; Science: Novosibirsk, Russia, 1974; p. 210. (In Russian) [Google Scholar]
- Botch, M.S.; Smagin, V.A. Flora and Vegetation of Mires in the North-West Russia and Principles of Their Protection; Transactions of Komarov Botanical Institute: Saint Petersburg, Russia, 1993; pp. 1–225. (In Russian) [Google Scholar]
- Smagin, V.A. Syntaxonomy of ridge dwarf shrub-grass-peatmoss communities in aapa-mires and fens of European Russia). Russ. J. Bot. 2012, 97, 939–960. (In Russian) [Google Scholar]
- Kuznetsov, O.L. Topological-ecological classification of mire vegetation of Karelia (ombrotrophic and oligotrophic communities). Transactions of Karelian Research Centre of RAS. 2005, 8, 15–46. (In Russian) [Google Scholar]
- Lapshina, E.D. Mire Vegetation of South-East of West Siberia; Yugorsk State University: Khanty-Mansijk-Novosibirsk, Russia, 2010; pp. 1–186. (In Russian) [Google Scholar]
- Neshatayeva, V.Y. Vegetation of the Kamchatka Peninsula; KMK Scientific Publishing Association: Moscow, Russia, 2009; pp. 1–537. (In Russian) [Google Scholar]
- Smagin, V.A.; Noskova, M.G.; Denisenkov, V.P. Localities of aapa mires in Arkhangelsk Region. Russ. J. Bot. 2013, 98, 1095–1108. (In Russian) [Google Scholar]
- Braslavskaya, T.Y.; Goryachkin, S.V.; Kutenkov, S.A.; Mamontov, V.N.; Popov, S.Y.; Puchnina, L.V.; Sidorova, O.V.; Torkhov, S.V.; Fedchenko, I.; Churakova, E.Y. Flora and Vegetation of the Belomorsko-Kulojskoe Plateau; North (Arctic) Federal University: Arkhangelsk, Russia, 2017; pp. 1–302. (In Russian) [Google Scholar]
- Smagin, V.A. Vegetation of fen, aapa-mires and palsa-bogs (pools) on the north of European Russia. Russ. J. Bot. 1999, 84, 80–96. (In Russian) [Google Scholar]
- Smagin, V.A. Vegetation of hollows, flarks and pools oligotrophic and mesotrophic mires of European Russia. Bull. Samara Sci. Cent. RAS 2012, 14, 1125–1129. (In Russian) [Google Scholar]
- Kuznetsov, O.L. The structure and dynamics of aapa bogs in Northern Karelia. Russ. Bot. J. 1982, 67, 1394–1400. (In Russian) [Google Scholar]
- Mironov, V.L.; Kuznetsov, O.L.; Maksimov, A.I.; Antipin, V.K.; Heikkilä, R.; Lindholm, T.; Kutenkov, S.A. On the flora of the Ypäyssuo hydrological (mire) nature reserve, Karelia (Russia). Trans. Karelian Res. Cent. RAS 2017, 1, 18–31. (In Russian) [Google Scholar]
- Borovichev, E.A.; Kozhin, M.N.; Ignashov, P.A.; Kirillova, N.R.; Kopeina, E.I.; Kravchenko, A.V.; Kuznetsov, O.L.; Kutenkov, S.A.; Melekhin, A.V.; Popova, K.B.; et al. Noteworthy records of plants, lichens and fungi in Murmansk region. II Proc. Karelian Sci. Cent. Russ. Acad. Sci. 2020, 1, 17–33. [Google Scholar] [CrossRef] [Green Version]
- Antonova, L.A.; Baburin, A.A.; Barkalov, V.Y.; Bezdeleva, T.A.; Van, V.M.; Vernoslova, M.I.; Dobrovolnaya, S.V.; Dulin, A.F.; Ermoshkin, A.V.; Kozhevnikova, A.E.; et al. Red Data Book of the Khabarovsk Territory: Rare and Endangered Species of Plants, Fungi and Animals: Official Publication; Ministry of Natural Resources of the Khabarovsk Territory: Khabarovsk, Russia; Institute of Water and Environmental Problems: Khabarovsk, Russia; FEB RAS. LLC “MIR”: Voronezh, Russia, 2019; p. 604. (In Russian) [Google Scholar]
- Chernyuk, A.V. The Main Stages of Vegetation Development in the Lower Amur Region in the Pleistocene and Holocene According to Palynological Data: Author’s Abstract of Cand. (Geogr.) Diss.; Lomonosov Moscow State University: Moscow, Russia, 1976; pp. 1–24. (In Russian) [Google Scholar]
- Bazarova, V.B.; Klimin, M.A.; Kopoteva, T.A. Holocene dynamics of the East Asian monsoon in the Lower Amur region (south of the Far East). Geogr. Nat. Resour. 2018, 3, 124–133. (In Russian) [Google Scholar]
- Razzhigaeva, N.G.; Grebennikova, T.A.; Ganzei, L.A.; Chakov, V.V.; Klimin, M.A.; Mokhova, L.M.; Zakharchenko, E.N. Stratigraphy of the watershed peatland and the development of the natural environment of Bolshoy Shantar Island in the Late Glacial-Holocene. Pac. Geol. 2021, 40, 85–102. [Google Scholar]
- Bogdanovskaia-Guihéneuf, I.D. Formation and evolution of strings and hollows in mires. Sov. Bot. 1936, 6, 35–52. (In Russian) [Google Scholar]
- Kuznetsov, O.L.; Kutenkov, S.A.; Churakova, E.Y. Aapa mires of the Arkhangelsk region: Diversity and genesis. In Wetlands and Flyways in the Barents/Euro-Arctic Region and along Green Belt of Fennoscandia, Proceedings of International Conference, 13–15 September 2011, Murmansk, Russia; State Nature Reserve “Pasvik”: Petrozavodsk, Russia, 2013; pp. 122–131. [Google Scholar]
- Rehell, S. Land uplift phenomenon and its effects on mire vegetation. In Finland—Land of Mires. Finnish Environment; Lindholm, T., Heikkilä, R., Eds.; Finnish Environment Institute: Helsinki, Finland, 2006; pp. 145–154. [Google Scholar]
- Kolari, T.H.M.; Sallinen, A.; Wolff, F.; Kumpula, T.; Tolonen, K.; Tahvanainen, T. Ongoing fen–bog transition in a boreal aapa mire inferred from repeated field sampling, aerial images, and Landsat data. Ecosystems 2021, 1–23. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | String, Height * of 25–50 cm | Low Hillock, Lawn, Height of 5–25 cm | Flark | |
|---|---|---|---|---|
| Number of relevé | 12 | 9 | 11 | |
| Vascular Plants | ||||
| 1. | Larix cajanderi Mayr | V1 ** | I+ | |
| 2. | Betula divaricata Ledeb. | V10 | IV+ | I1 |
| 3. | Andromeda polifolia L. | V5 | V3 | V+ |
| 4. | Calamagrostis langsdorfii (Lik.) Trin. | IV+ | II+ | I+ |
| 5. | Carex cespitosa var. minuta (Franch.) Kük. | I+ | II+ | IV2 |
| 6. | Carex chordoriza L. | II+ | II+ | II+ |
| 7. | Carex gynocrates Warnst. | I+ | ||
| 8. | Carex lasiocarpa Ehrh. | IV5 | V10 | V2 |
| 9. | Carex laxa Wahleb. | III+ | ||
| 10. | Carex limosa L. | I+ | IV+ | V+ |
| 11. | Carex middendorffii F.Schmidt | III2 | I+ | I+ |
| 12. | Carex tenuifolia Wahlehb. | I+ | I+ | |
| 13. | Chamaedaphne calyculata (L.) Moench | V5 | III+ | |
| 14. | Drosera anglica Huds. | III+ | V+ | |
| 15. | Drosera rotundifolia L. | IV+ | IV+ | |
| 16. | Eleocharis wichurae Boech. | I+ | ||
| 17. | Equisetum fluviatile L. | IV+ | IV+ | V2 |
| 18. | Eriocaulon schischkinii Tzvelev | II+ | IV+ | |
| 19. | Eriophorum gracile Koch | I+ | ||
| 20. | Eriophorum russeolum Fr. | I+ | I+ | |
| 21. | Glyceria spiculosa (F. Schmidt) Roshev. | IV+ | III+ | |
| 22. | Habenaria linearifolia Maxim. | I+ | III+ | |
| 23. | Hammarbia paludosa (L.) O. Kuntze | II+ | ||
| 24. | Iris laevigata Fishet Mey. | III+ | V+ | V+ |
| 25. | Juncus stygius L. | III+ | IV+ | |
| 26. | Ledum palustre L | III1 | ||
| 27. | Lobelia sessilifolia Lamb. | II+ | IV+ | V+ |
| 28. | Menyanthes trifoliata L. | IV+ | V+ | V5 |
| 29. | Osmundastrum asiaticum (Fernald) Tagawa | I+ | ||
| 30. | Ostericum maximowiczii (F. Schmidt ex Maxim.) Kitag. | I+ | ||
| 31. | Oxycoccus microcarpus Turcz. ex Rupr. | III+ | I+ | |
| 32. | Oxycoccus palustris Pers. | V4 | IV+ | I1 |
| 33. | Parnassia palustris L. | II+ | III+ | I+ |
| 34. | Pedicularis grandiflora Fisch. | I+ | ||
| 35. | Pedicularis resupinata L. | I+ | I+ | |
| 36. | Pogonia japonica Rchb. f. | II+ | + | |
| 37. | Rhynchospora alba (L.) Vahl | I+ | V2 | V2 |
| 38. | Salix myrtilloides L. | III+ | III+ | |
| 39. | Sanguisorba parviflora (Maxim.) Takeda | V1 | IV1 | I+ |
| 40. | Saussurea amurensis Turcz. ex DC. | III+ | II+ | |
| 41. | Scheuchzeria palustris L. | IV+ | V1 | |
| 42. | Scutellaria regeliana Nakai. | II+ | III+ | |
| 43. | Smilacina trifolia (L.) Desf. | IV+ | II+ | |
| 44. | Sparganium hyperboreum Laest. ex Beurl. | I+ | ||
| 45. | Spiranthes sinensis (Pers.) Ames | I+ | II+ | |
| 46. | Triadenum japonicum (Blume) Makino | I+ | ||
| 47. | Trichophorum alpinum (L.) Pers. | III+ | ||
| 48. | Utricularia intermedia Hayne | V11 | ||
| 49. | Utricularia macrozhiza Le Conte | III8 | ||
| 50. | Utricularia minor L. | III+ | ||
| Bryophytes | ||||
| 51. | Aulacomnium palustre (Hedw.) Schwägr. | III+ | II+ | I+ |
| 52. | Campylium stellatum (Hedw.) CEOJensen | II+ | II+ | |
| 53. | Dicranum bergerii Bland. Ex Hoppe | I+ | ||
| 54. | Dicranum bonjeanii De Not. | I+ | ||
| 55. | Dicranum polysetum Sw. | I+ | ||
| 56. | Drepanocladus polygamus (Bruch. et al.) Hedenäs | I+ | ||
| 57. | Pleurozium schreberi (Brid.) Mitt. | II+ | ||
| 58. | Polytrichum commune Hedw | I+ | ||
| 59. | Polytrichum strictum Brid. | I+ | ||
| 60. | Sphagnum magellanicum s.l. *** | V65 | V5 | I+ |
| 61. | Sphagnum angustifolium (Warnst.) CEO Jensen | IV3 | I+ | |
| 62. | Sphagnum annulatum Warnstorf, Carl (Friedrich E.) | I+ | ||
| 63. | Sphagnum aongstroemii C.Hartman | I+ | ||
| 64. | Sphagnum capilifolium (Ehrh.) Hedw. | I+ | ||
| 65. | Sphagnum fallax (Klinggr.) Klinggr. | I+ | III+ | |
| 66. | Sphagnum fuscum (Schimp.) H.Klinggr. | III3 | ||
| 67. | Sphagnum imbricatum Hornsch. ex Russow | II+ | II+ | I+ |
| 68. | Sphagnum jensenii H. Lindb. | II3 | ||
| 69. | Sphagnum lenense H. Lindb | I+ | ||
| 70. | Sphagnum obtusum Warnst. | I+ | II+ | V+ |
| 71. | Sphagnum orientale L.I. Savicz | I+ | V9 | IV1 |
| 72. | Sphagnum papillosum Lindb. | II2 | V70 | V4 |
| 73. | Sphagnum rubellum Wils. | V3 | I+ | |
| 74. | Sphagnum squarrosum Crome | I+ | ||
| 75. | Sphagnum subfulvum Sjors | I+ | III1 | I+ |
| 76. | Sphagnum subsecundum Nees | I+ | I+ | |
| 77. | Sphagnum warnstorfii Russ. | I+ | I+ | |
| 78. | Straminergon stramineum (Dicks. ex Brid.) Hedenäs | I+ | ||
| 79. | Tomentypnum nitens (Hedw.) Loeske | I+ | ||
| 80. | Warnstorfia exannulata (Schimp.) Loeske | I+ | ||
| 81. | Warnstorfia fluitans (Hedw.) Loeske | I+ | ||
| Lichens | ||||
| 82. | Cetraria islandica (L.) Ach. | I+ | ||
| 83. | Cladonia arbuscula (Wallr.) Flot. | I+ | ||
| 84. | Cladonia furcata (Huds.) Schrad. | I+ | ||
| 85. | Cladonia rangiferina (L.) F. H. Wigg | I+ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kutenkov, S.; Chakov, V.; Kuptsova, V. Topology, Vegetation and Stratigraphy of Far Eastern Aapa Mires (Khabarovsk Region, Russia). Land 2022, 11, 96. https://doi.org/10.3390/land11010096
Kutenkov S, Chakov V, Kuptsova V. Topology, Vegetation and Stratigraphy of Far Eastern Aapa Mires (Khabarovsk Region, Russia). Land. 2022; 11(1):96. https://doi.org/10.3390/land11010096
Chicago/Turabian StyleKutenkov, Stanislav, Vladimir Chakov, and Viktoriya Kuptsova. 2022. "Topology, Vegetation and Stratigraphy of Far Eastern Aapa Mires (Khabarovsk Region, Russia)" Land 11, no. 1: 96. https://doi.org/10.3390/land11010096
APA StyleKutenkov, S., Chakov, V., & Kuptsova, V. (2022). Topology, Vegetation and Stratigraphy of Far Eastern Aapa Mires (Khabarovsk Region, Russia). Land, 11(1), 96. https://doi.org/10.3390/land11010096