
Heterogeneity-Based Management Restores Diversity and Alters Vegetation Structure without Decreasing Invasive Grasses in Working Mixed-Grass Prairie


 ,
, 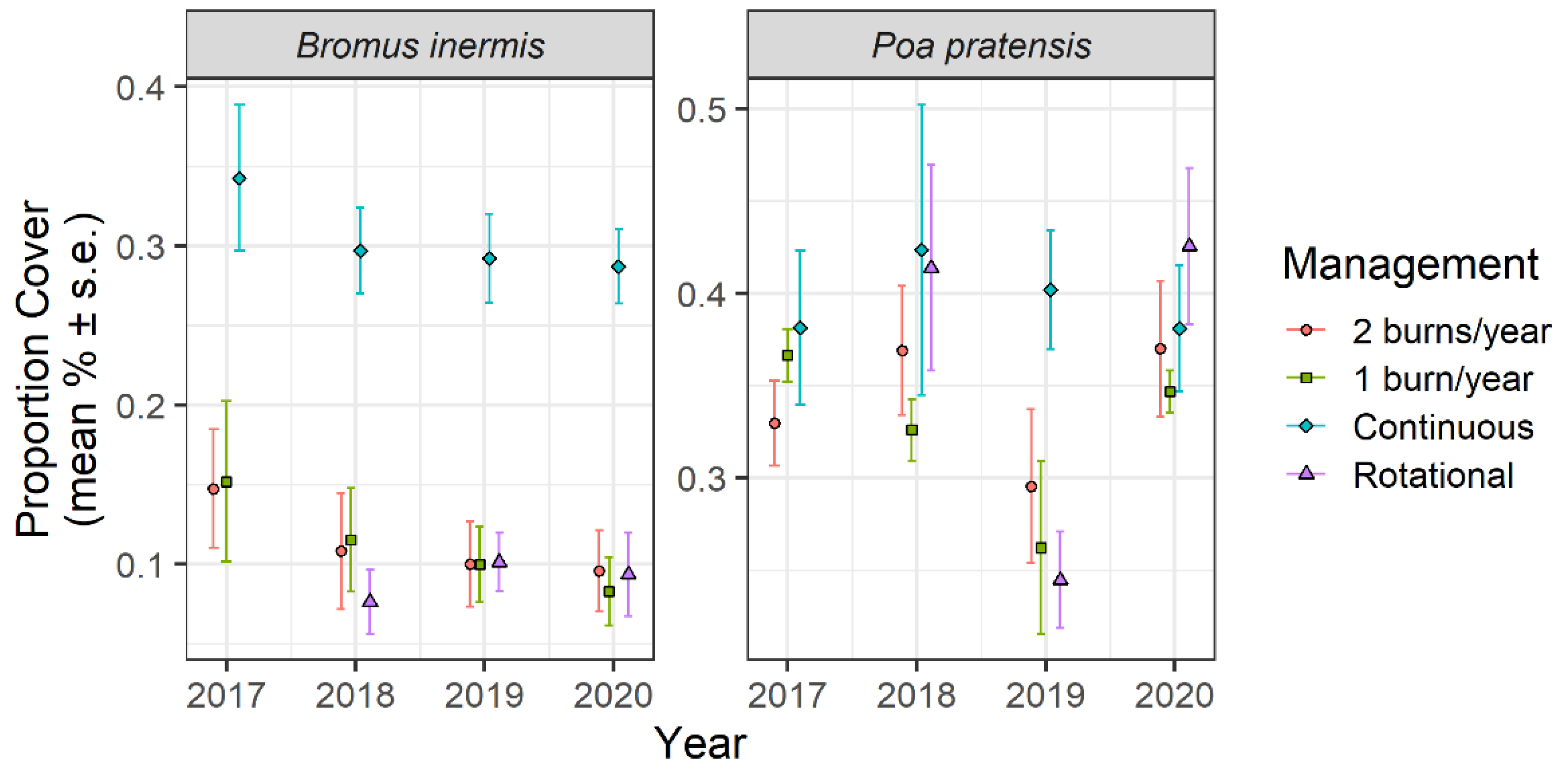{kind=link}
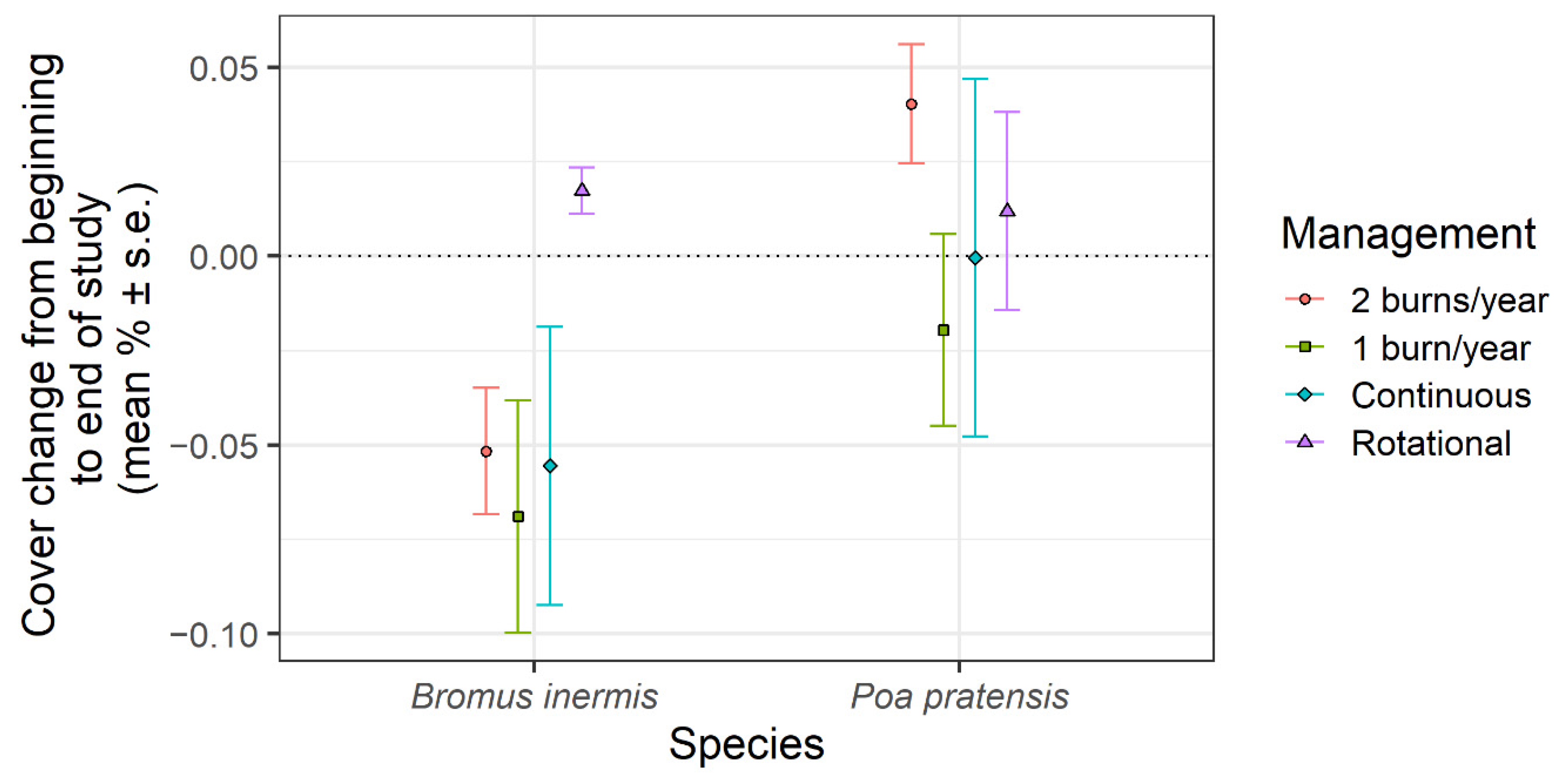{kind=link}
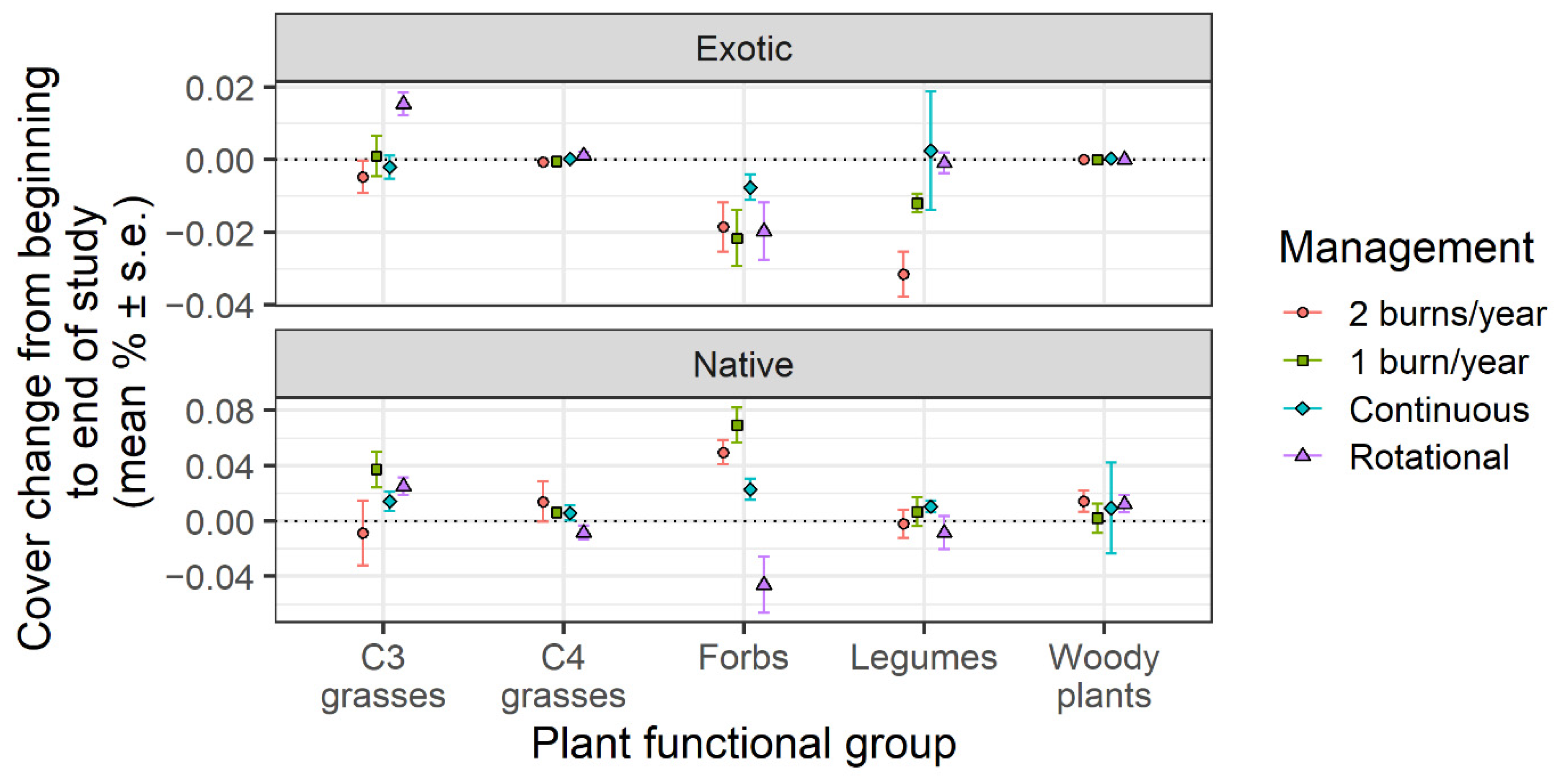{kind=link}
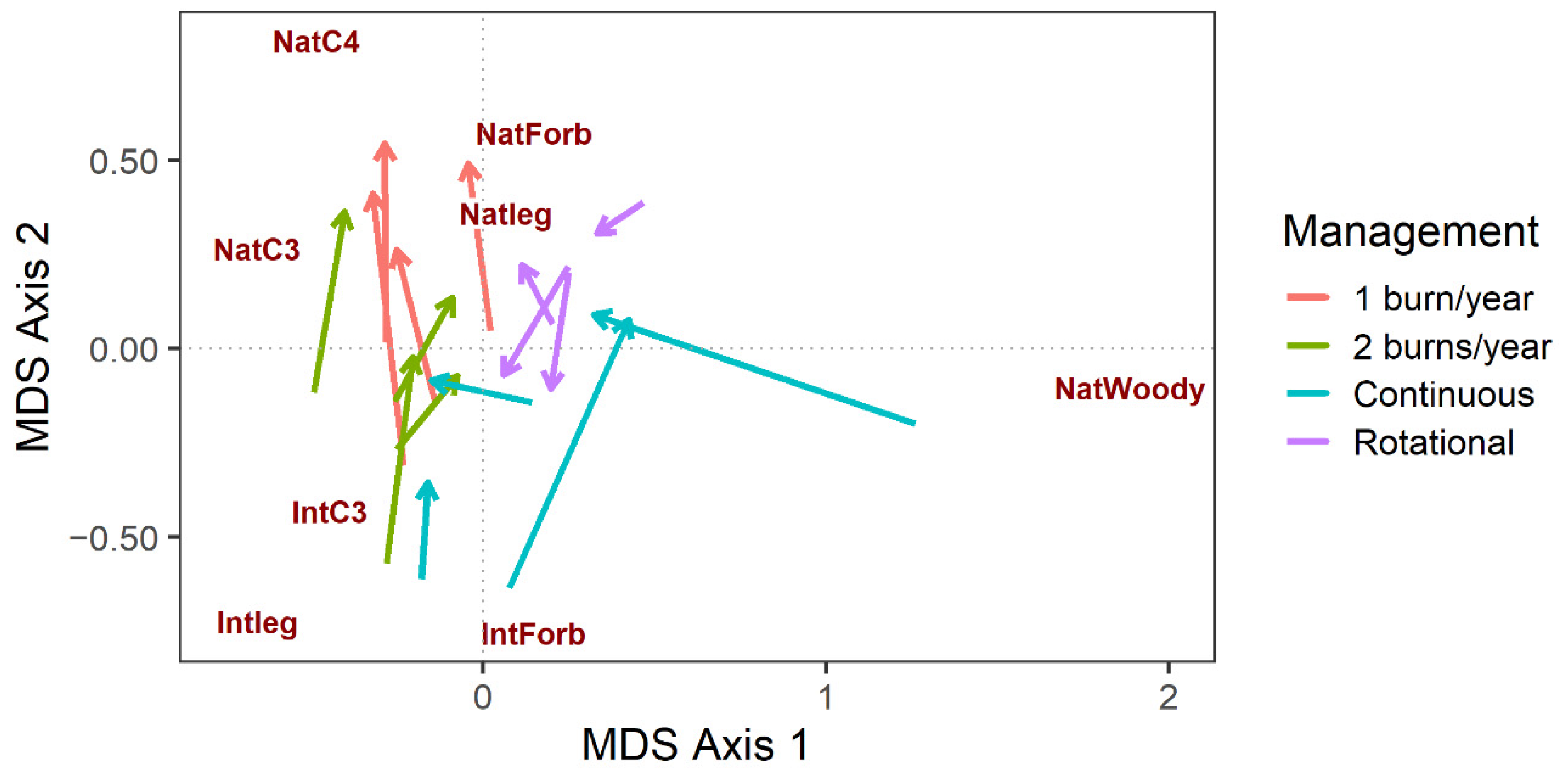{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
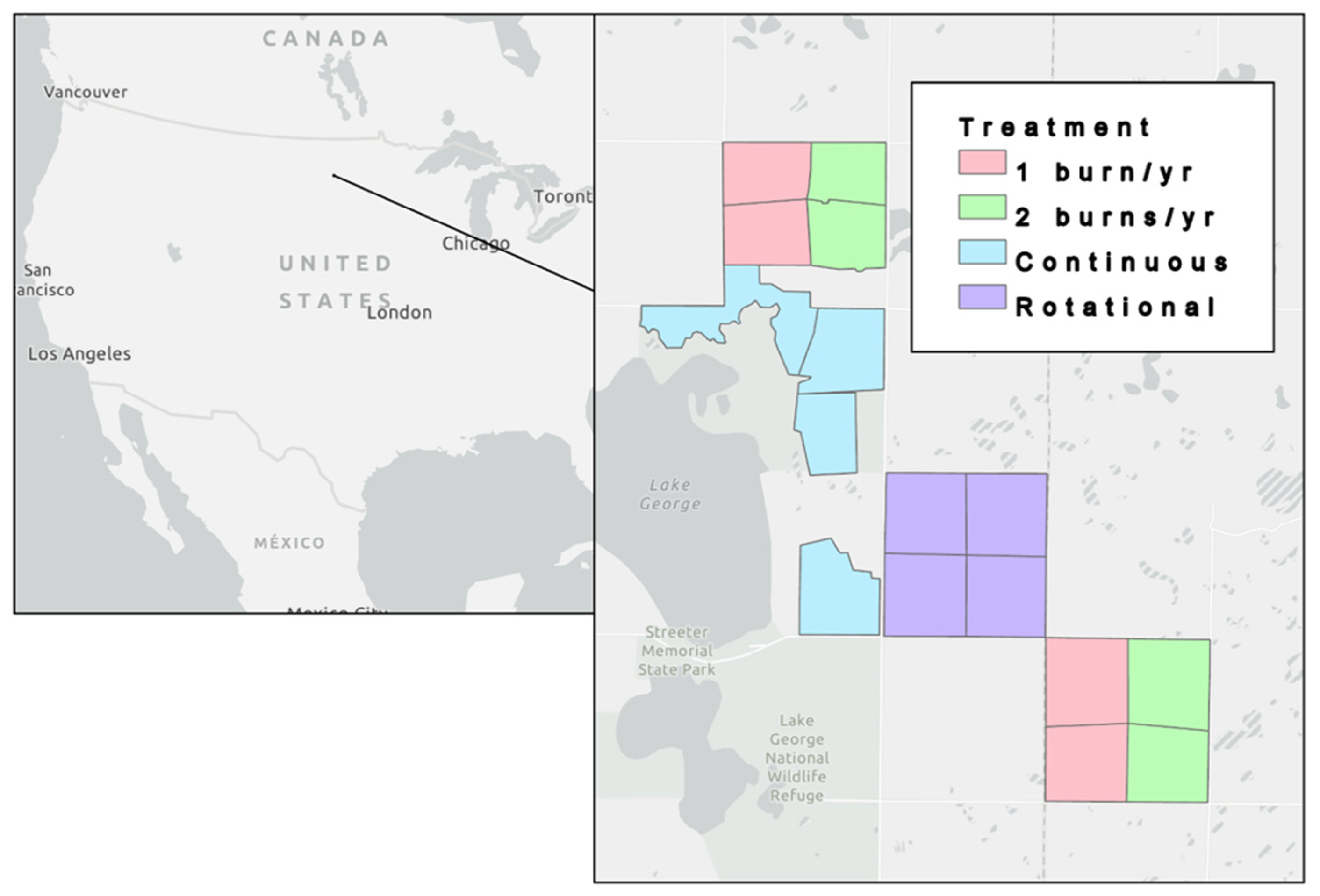
2.1. Study Site
2.2. Data Collection
2.3. Data Analysis
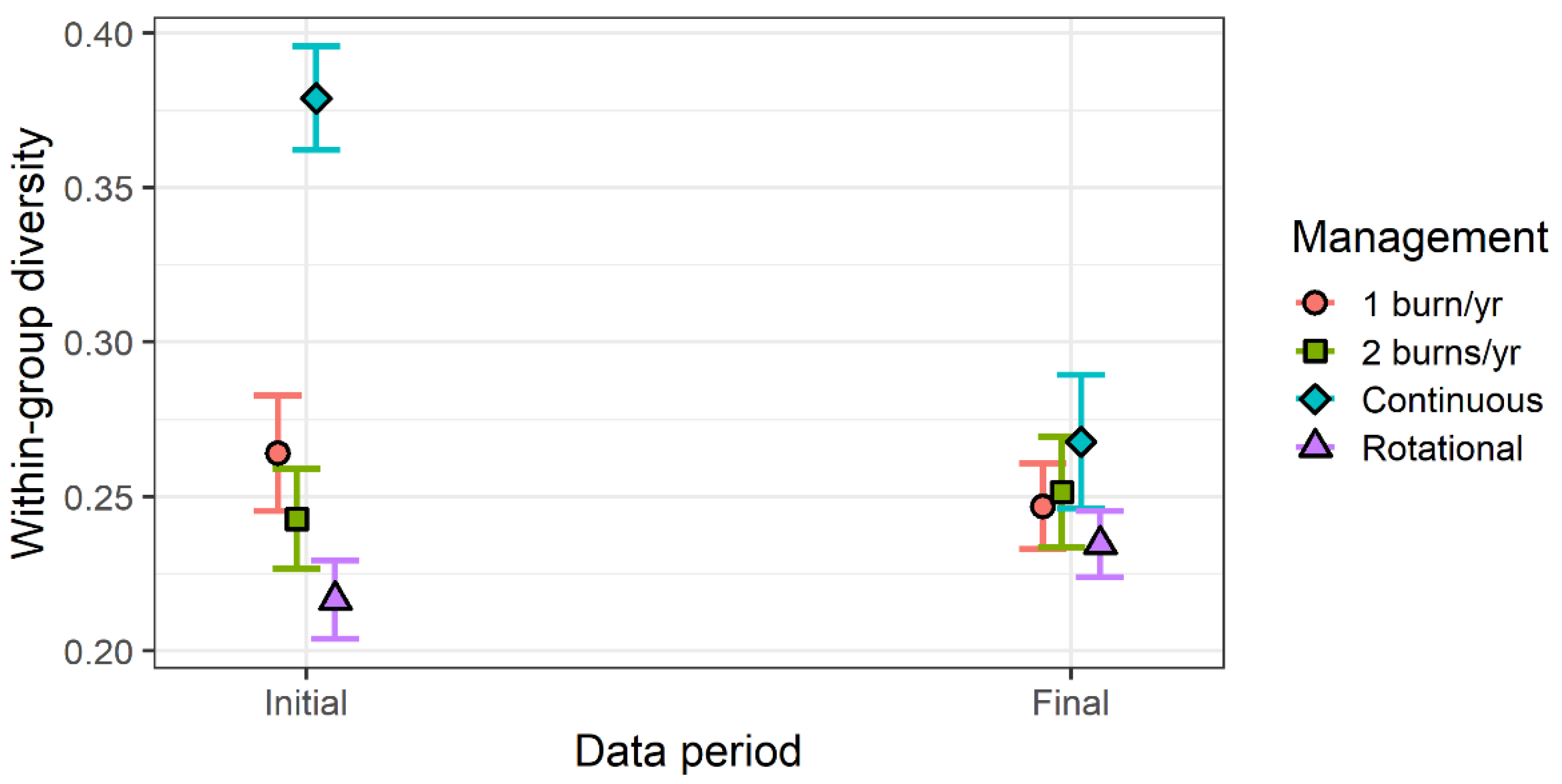
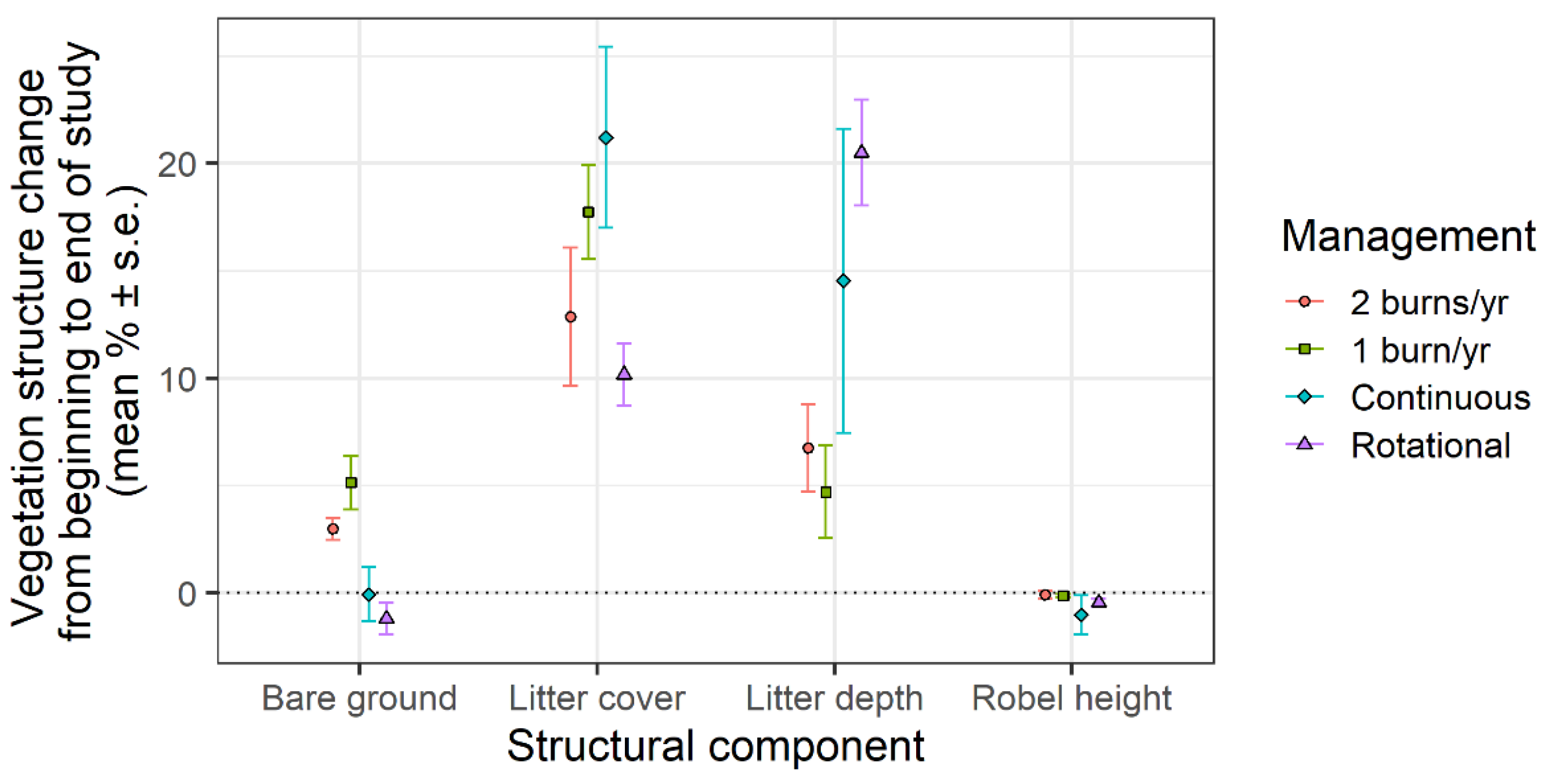
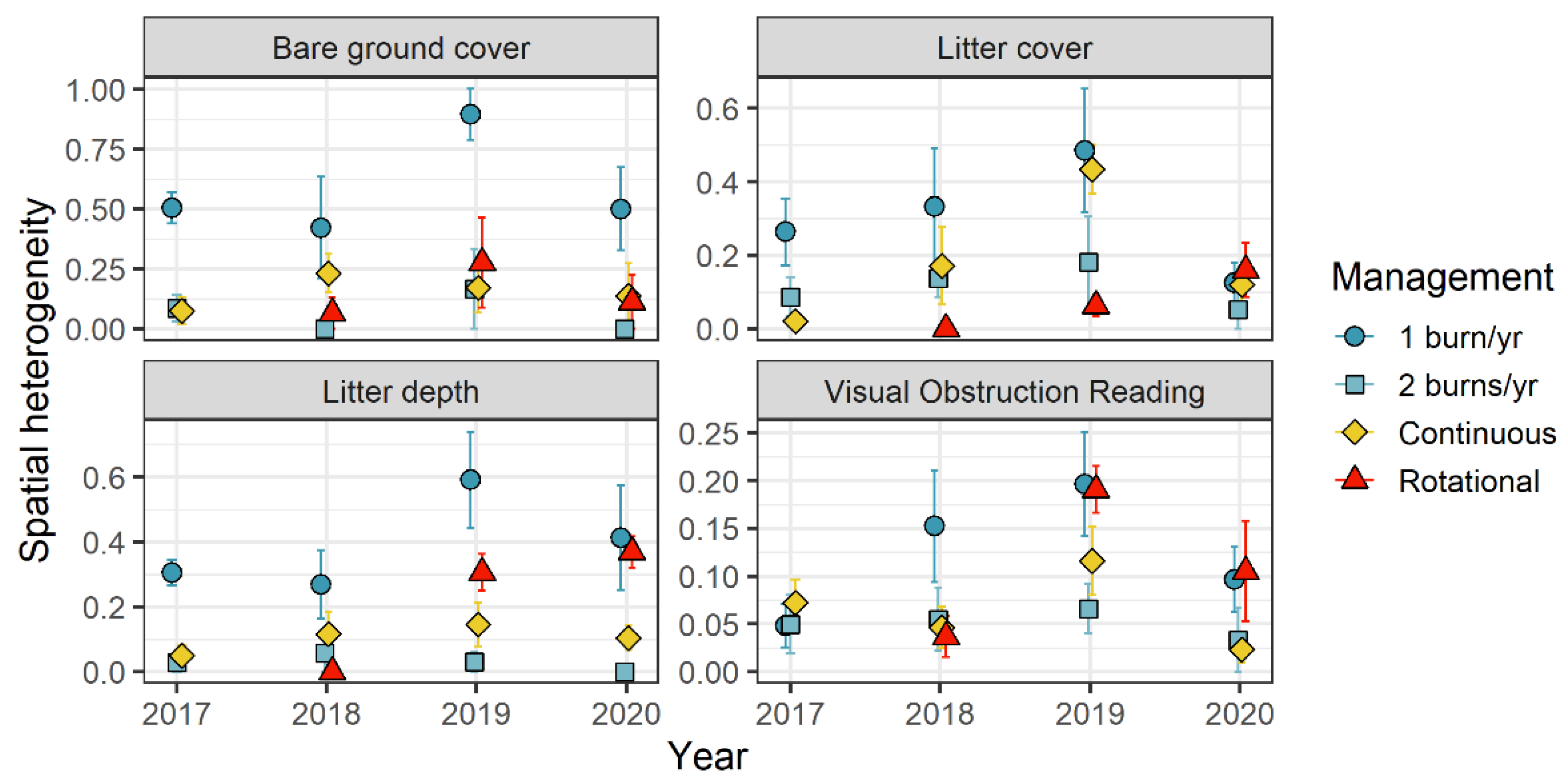
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Flanders, A.A.; Kuvlesky, W.P.; Ruthven, D.C.; Zaiglin, R.E.; Bingham, R.L.; Fulbright, T.E.; Hernández, F.; Brennan, L.A. Effects of Invasive Exotic Grasses on South Texas Rangeland Breeding Birds. Auk 2006, 123, 171–182. [Google Scholar] [CrossRef]
- Pearson, D.E. Invasive Plant Architecture Alters Trophic Interactions by Changing Predator Abundance and Behavior. Oecologia 2009, 159, 549–558. [Google Scholar] [CrossRef]
- Gasch, C.K.; Toledo, D.; Kral-O’Brien, K.; Baldwin, C.; Bendel, C.; Fick, W.; Gerhard, L.; Harmon, J.; Hendrickson, J.; Hovick, T.; et al. Kentucky Bluegrass Invaded Rangeland: Ecosystem Implications and Adaptive Management Approaches. Rangelands 2020, 42, 106–116. [Google Scholar] [CrossRef]
- Sexton, A.N.; Emery, S.M. Grassland Restorations Improve Pollinator Communities: A Meta-Analysis. J. Insect. Conserv. 2020, 24, 719–726. [Google Scholar] [CrossRef]
- Rodhouse, T.J.; Lonneker, J.; Bowersock, L.; Popp, D.; Thompson, J.C.; Dicus, G.H.; Irvine, K.M. Resilience to Fire and Resistance to Annual Grass Invasion in Sagebrush Ecosystems of US National Parks. Glob. Ecol. Conserv. 2021, 28, e01689. [Google Scholar] [CrossRef]
- Ratcliffe, H.; Ahlering, M.; Carlson, D.; Vacek, S.; Allstadt, A.; Dee, L.E. Invasive Species Do Not Exploit Early Growing Seasons in Burned Tallgrass Prairies. Ecol. Appl. 2022, e2641. [Google Scholar] [CrossRef] [PubMed]
- Terry, T.J.; Madsen, M.D.; Gill, R.A.; Anderson, V.J.; St. Clair, S.B. Selective Herbicide Control: Using Furrows and Carbon Seed Coatings to Establish a Native Bunchgrass While Reducing Cheatgrass Cover. Restor. Ecol. 2021, 29, e13351. [Google Scholar] [CrossRef]
- Fogarty, D.T.; Roberts, C.P.; Uden, D.R.; Donovan, V.M.; Allen, C.R.; Naugle, D.E.; Jones, M.O.; Allred, B.W.; Twidwell, D. Woody Plant Encroachment and the Sustainability of Priority Conservation Areas. Sustainability 2020, 12, 8321. [Google Scholar] [CrossRef]
- Thomas, M.; Reid, A. Are Exotic Natural Enemies an Effective Way of Controlling Invasive Plants? Trends Ecol. Evol. 2007, 22, 447–453. [Google Scholar] [CrossRef]
- Rouget, M.; Richardson, D.M. Inferring Process from Pattern in Plant Invasions: A Semimechanistic Model Incorporating Propagule Pressure and Environmental Factors. Am. Nat. 2003, 162, 713–724. [Google Scholar] [CrossRef]
- Boyd, C.S. Managing for Resilient Sagebrush Plant Communities in the Modern Era: We’re Not in 1850 Anymore. Rangelands 2022, 44, 167–172. [Google Scholar] [CrossRef]
- Emery, S.M.; Doran, P.J. Presence and Management of the Invasive Plant Gypsophila Paniculata (Baby’s Breath) on Sand Dunes Alters Arthropod Abundance and Community Structure. Biol. Conserv. 2013, 161, 174–181. [Google Scholar] [CrossRef]
- Davis, M.A.; Chew, M.K.; Hobbs, R.J.; Lugo, A.E.; Ewel, J.J.; Vermeij, G.J.; Brown, J.H.; Rosenzweig, M.L.; Gardener, M.R.; Carroll, S.P.; et al. Don’t Judge Species on Their Origins. Nature 2011, 474, 153–154. [Google Scholar] [CrossRef] [PubMed]
- Rinella, M.J.; Maxwell, B.D.; Fay, P.K.; Weaver, T.; Sheley, R.L. Control Effort Exacerbates Invasive-Species Problem. Ecol. Appl. 2009, 19, 155–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assis, G.B.; Pilon, N.A.L.; Siqueira, M.F.; Durigan, G. Effectiveness and Costs of Invasive Species Control Using Different Techniques to Restore Cerrado Grasslands. Restor. Ecol. 2021, 29, e13219. [Google Scholar] [CrossRef]
- Fuhlendorf, S.D.; Fynn, R.W.; McGranahan, D.A.; Twidwell, D. Heterogeneity as the Basis for Rangeland Management. In Rangeland Systems; Springer: New York, NY, USA, 2017; pp. 169–196. [Google Scholar]
- Fuhlendorf, S.D.; Engle, D.M. Restoring Heterogeneity on Rangelands: Ecosystem Management Based on Evolutionary Grazing Patterns. BioScience 2001, 51, 625. [Google Scholar] [CrossRef] [Green Version]
- Fuhlendorf, S.D.; Harrell, W.C.; Engle, D.M.; Hamilton, R.G.; Davis, C.A.; Leslie, D.M. Should Heterogeneity Be the Basis for Conservation? Grassland Bird Response to Fire and Grazing. Ecol. Appl. 2006, 16, 1706–1716. [Google Scholar] [CrossRef] [Green Version]
- Capozzelli, J.F.; Miller, J.R.; Debinski, D.M.; Schacht, W.H. Restoring the Fire–Grazing Interaction Promotes Tree–Grass Coexistence by Controlling Woody Encroachment. Ecosphere 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Hovick, T.J.; Elmore, R.D.; Fuhlendorf, S.D.; Engle, D.M.; Hamilton, R.G. Spatial Heterogeneity Increases Diversity and Stability in Grassland Bird Communities. Ecol. Appl. 2015, 25, 662–672. [Google Scholar] [CrossRef]
- Ricketts, A.M.; Sandercock, B.K. Patch-burn Grazing Increases Habitat Heterogeneity and Biodiversity of Small Mammals in Managed Rangelands. Ecosphere 2016, 7, e01431. [Google Scholar] [CrossRef] [Green Version]
- Lituma, C.M.; Buckley, B.R.; Keyser, P.D.; Holcomb, E.; Smith, R.; Morgan, J.; Applegate, R.D. Effects of Patch-Burn Grazing and Rotational Grazing on Grassland Bird Abundance, Species Richness, and Diversity in Native Grassland Pastures of the Midsouth USA. Agric. Ecosyst. Environ. 2022, 324, 107710. [Google Scholar] [CrossRef]
- Sherrill, C.W.; Fuhlendorf, S.D.; Goodman, L.E.; Elmore, R.D.; Hamilton, R.G. Managing an Invasive Species While Simultaneously Conserving Native Plant Diversity. Rangel. Ecol. Manag. 2022, 80, 87–95. [Google Scholar] [CrossRef]
- Dixon, C.; Vacek, S.; Grant, T. Evolving Management Paradigms on U.S. Fish and Wildlife Service Lands in the Prairie Pothole Region. Rangelands 2019, 41, 36–43. [Google Scholar] [CrossRef]
- Palit, R.; Gramig, G.; DeKeyser, E.S. Kentucky Bluegrass Invasion in the Northern Great Plains and Prospective Management Approaches to Mitigate Its Spread. Plants 2021, 10, 817. [Google Scholar] [CrossRef]
- Palit, R.; DeKeyser, E.S. Impacts and Drivers of Smooth Brome (Bromus inermis Leyss.) Invasion in Native Ecosystems. Plants 2022, 11, 1340. [Google Scholar] [CrossRef]
- Toledo, D.; Sanderson, M.; Spaeth, K.; Hendrickson, J.; Printz, J. Extent of Kentucky Bluegrass and Its Effect on Native Plant Species Diversity and Ecosystem Services in the Northern Great Plains of the United States. Invasive Plant Sci. Manag. 2014, 7, 543–552. [Google Scholar] [CrossRef]
- Kral-O’Brien, K.C.; Limb, R.F.; Hovick, T.J.; Harmon, J.P. Compositional Shifts in Forb and Butterfly Communities Associated with Kentucky Bluegrass Invasions. Rangel. Ecol. Manag. 2019, 72, 301–309. [Google Scholar] [CrossRef]
- Waller, S.S.; Schmidt, D.K. Improvement of Eastern Nebraska Tallgrass Range Using Atrazine or Glyphosate. J. Range Manag. 1983, 36, 87. [Google Scholar] [CrossRef]
- Hendrickson, J.R.; Sedivec, K.K.; Toledo, D.; Printz, J. Challenges Facing Grasslands in the Northern Great Plains and North Central Region. Rangelands 2019, 41, 23–29. [Google Scholar] [CrossRef]
- McGranahan, D.A.; Hovick, T.J.; Elmore, R.D.; Engle, D.M.; Fuhlendorf, S.D. Moderate Patchiness Optimizes Heterogeneity, Stability, and Beta Diversity in Mesic Grassland. Ecol. Evol. 2018, 8, 5008–5015. [Google Scholar] [CrossRef] [Green Version]
- Patton, B.D.; Dong, X.; Nyren, P.E.; Nyren, A. Effects of Grazing Intensity, Precipitation, and Temperature on Forage Production. Rangel. Ecol. Manag. 2007, 60, 656–665. [Google Scholar] [CrossRef]
- Duquette, C.A.; Hovick, T.J.; Geaumont, B.A.; Harmon, J.P.; Limb, R.F.; Sedivec, K.K. Spatially Discrete Disturbance Processes Enhance Grassland Floral Resources. J. Appl. Ecol. 2022, 59, 1700–1708. [Google Scholar] [CrossRef]
- Duquette, C.A.; Hovick, T.J.; Geaumont, B.A.; Harmon, J.P.; Limb, R.F.; Sedivec, K.K. Embracing inherent and imposed sources of heterogeneity in rangeland bird management. In preprint.
- Robel, R.J.; Briggs, J.N.; Dayton, A.D.; Hulbert, L.C. Relationships between Visual Obstruction Measurements and Weight of Grassland Vegetation. J. Range Manag. 1970, 23, 295. [Google Scholar] [CrossRef] [Green Version]
- Harrell, W.C.; Fuhlendorf, S.D. Evaluation of Habitat Structural Measures in a Shrubland Community. Rangel. Ecol. Manag. 2002, 55, 488–493. [Google Scholar] [CrossRef]
- Daubenmire, R.F. Canopy Coverage Method of Vegetation Analysis. Northwest Sci. 1959, 33, 43–64. [Google Scholar]
- Kruskal, J.B. Multidimensional Scaling by Optimizing Goodness of Fit to a Nonmetric Hypothesis. Psychometrika 1964, 29, 1–27. [Google Scholar] [CrossRef]
- Oksanen, J.; Kindt, R.; Legendre, P.; O’Hara, B.; Stevens, M.H.H.; Oksanen, M.J.; Suggests, M.A.S.S. The Vegan Package. Community Ecol. Package 2007, 10, 631–637. [Google Scholar]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate Dispersion as a Measure of Beta Diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef]
- Raynor, E.J.; McGranahan, D.A.; Miller, J.R.; Debinski, D.M.; Schacht, W.H.; Engle, D.M. Moderate Grazer Density Stabilizes Forage Availability More Than Patch Burning in Low-Stature Grassland. Land 2021, 10, 395. [Google Scholar] [CrossRef]
- McGranahan, D.A.; Hovick, T.J.; Elmore, R.D.; Engle, D.M.; Fuhlendorf, S.D.; Winter, S.L.; Miller, J.R.; Debinski, D.M. Temporal Variability in aboveground plant biomass decreases as spatial variability increases. Ecology 2016, 97, 555–560. [Google Scholar] [CrossRef]
- McGranahan, D.A.; Engle, D.M.; Fuhlendorf, S.D.; Winter, S.L.; Miller, J.R.; Debinski, D.M. Inconsistent Outcomes of Heterogeneity-Based Management Underscore Importance of Matching Evaluation to Conservation Objectives. Environ. Sci. Policy 2013, 31, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Link, A.; Kobiela, B.; DeKeyser, S.; Huffington, M. Effectiveness of Burning, Herbicide, and Seeding Toward Restoring Rangelands in Southeastern North Dakota. Rangel. Ecol. Manag. 2017, 70, 599–603. [Google Scholar] [CrossRef]
- Ruehmann, M.B.; Desmond, M.J.; Gould, W.R. Effects of Smooth Brome on Brewer’s Sparrow Nest Survival in Sagebrush Steppe. Condor 2011, 113, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Kral, K.; Limb, R.; Ganguli, A.; Hovick, T.; Sedivec, K. Seasonal Prescribed Fire Variation Decreases Inhibitory Ability of Poa Pratensis, L. and Promotes Native Plant Diversity. J. Environ. Manag. 2018, 223, 908–916. [Google Scholar] [CrossRef] [PubMed]
- Hovick, T.J.; Elmore, R.D.; Fuhlendorf, S.D. Structural Heterogeneity Increases Diversity of Non-Breeding Grassland Birds. Ecosphere 2014, 5, art62. [Google Scholar] [CrossRef]
- Duquette, C.A.; Hovick, T.J.; Geaumont, B.A.; Harmon, J.P.; Limb, R.F.; Sedivec, K.K. Patch-burn grazing benefits nesting of short-vegetation specialists in the northern Great Plains. In preprint.
- Londe, D.W.; Elmore, R.D.; Davis, C.A.; Fuhlendorf, S.D.; Hovick, T.J.; Luttbeg, B.; Rutledge, J. Fine-Scale Habitat Selection Limits Trade-Offs between Foraging and Temperature in a Grassland Bird. Behav. Ecol. 2021, 32, 625–637. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Beck, J.L.; Duchardt, C.J.; Rachlow, J.L.; Veblen, K.E. Leveraging Rangeland Monitoring Data for Wildlife: From Concept to Practice. Rangelands 2022, 44, 87–98. [Google Scholar] [CrossRef]
- Bailey, D.W.; Brown, J.R. Rotational Grazing Systems and Livestock Grazing Behavior in Shrub-Dominated Semi-Arid and Arid Rangelands. Rangel. Ecol. Manag. 2011, 64, 1–9. [Google Scholar] [CrossRef]
- Vermeire, L.T.; Mitchell, R.B.; Fuhlendorf, S.D.; Gillen, R.L. Patch Burning Effects on Grazing Distribution. Rangel. Ecol. Manag. 2004, 57, 248–252. [Google Scholar] [CrossRef]
- Fuhlendorf, S.D.; Engle, D.M.; Elmore, R.D.; Limb, R.F.; Bidwell, T.G. Conservation of Pattern and Process: Developing an Alternative Paradigm of Rangeland Management. Rangel. Ecol. Manag. 2012, 65, 579–589. [Google Scholar] [CrossRef] [Green Version]
- Printz, J.L.; Hendrickson, J.R. Impacts of Kentucky Bluegrass Invasion (Poa pratensis L.) on Ecological Processes in the Northern Great Plains. Rangelands 2015, 37, 226–232. [Google Scholar] [CrossRef] [Green Version]
- Bosy, J.L.; Reader, R.J. Mechanisms Underlying the Suppression of Forb Seedling Emergence by Grass (Poa pratensis) Litter. Funct. Ecol. 1995, 9, 635. [Google Scholar] [CrossRef]
- Cutter, J.; Geaumont, B.; McGranahan, D.; Harmon, J.; Limb, R.; Schauer, C.; Hovick, T. Cattle and Sheep Differentially Alter Floral Resources and the Native Bee Communities in Working Landscapes. Ecol. Appl. 2021, 31, e02406. [Google Scholar] [CrossRef] [PubMed]
- Simanonok, S.C.; Otto, C.R.V.; Buhl, D.A. Floral Resource Selection by Wild Bees and Honey Bees in the Midwest United States: Implications for Designing Pollinator Habitat. Restor. Ecol. 2021, 29, e13456. [Google Scholar] [CrossRef]
- Duchardt, C.J.; Miller, J.R.; Debinski, D.M.; Engle, D.M. Adapting the Fire-Grazing Interaction to Small Pastures in a Fragmented Landscape for Grassland Bird Conservation. Rangel. Ecol. Manag. 2016, 69, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Limb, R.F.; Hickman, K.R.; Engle, D.M.; Norland, J.E.; Fuhlendorf, S.D. Digital photography: Reduced investigator variation in visual obstruction measurements for southern tallgrass prairie. Rangel. Ecol. Manag. 2007, 60, 548–552. [Google Scholar] [CrossRef]
- Jansen, V.S.; Kolden, C.A.; Greaves, H.E.; Eitel, J.U.H. Lidar provides novel insights into the effect of pixel size and grazing intensity on measures of spatial heterogeneity in a native bunchgrass ecosystem. Remote Sens. Environ. 2019, 235, 111432. [Google Scholar] [CrossRef]
- Angeler, D.G.; Chaffin, B.C.; Sundstrom, S.M.; Garmestani, A.; Pope, K.L.; Uden, D.R.; Twidwell, D.; Allen, C.R. Coerced regimes: Management challenges in the Anthropocene. Ecol. Soc. 2020, 25, 25. [Google Scholar] [CrossRef] [Green Version]
- McGranahan, D.A.; Henderson, C.B.; Hill, J.S.; Raicovich, G.M.; Wilson, W.N.; Smith, C.K. Patch burning improves forage quality and creates grass-bank in an old-field pasture: Results of a demonstration trial. Southeast Nat. 2014, 13, 200–207. [Google Scholar] [CrossRef]
- Wanchuk, M.R.; McGranahan, D.A.; Sedivec, K.K.; Swanson, K.C.; Hovick, T.J.; Limb, R.F. Contrasts in Forage Mineral Concentration with Patch-Burn Grazing: A Preliminary Analysis. Transl. Anim. Sci. 2021, 5, S75–S79. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duquette, C.; McGranahan, D.A.; Wanchuk, M.; Hovick, T.; Limb, R.; Sedivec, K. Heterogeneity-Based Management Restores Diversity and Alters Vegetation Structure without Decreasing Invasive Grasses in Working Mixed-Grass Prairie. Land 2022, 11, 1135. https://doi.org/10.3390/land11081135
Duquette C, McGranahan DA, Wanchuk M, Hovick T, Limb R, Sedivec K. Heterogeneity-Based Management Restores Diversity and Alters Vegetation Structure without Decreasing Invasive Grasses in Working Mixed-Grass Prairie. Land. 2022; 11(8):1135. https://doi.org/10.3390/land11081135
Chicago/Turabian StyleDuquette, Cameron, Devan Allen McGranahan, Megan Wanchuk, Torre Hovick, Ryan Limb, and Kevin Sedivec. 2022. "Heterogeneity-Based Management Restores Diversity and Alters Vegetation Structure without Decreasing Invasive Grasses in Working Mixed-Grass Prairie" Land 11, no. 8: 1135. https://doi.org/10.3390/land11081135
APA StyleDuquette, C., McGranahan, D. A., Wanchuk, M., Hovick, T., Limb, R., & Sedivec, K. (2022). Heterogeneity-Based Management Restores Diversity and Alters Vegetation Structure without Decreasing Invasive Grasses in Working Mixed-Grass Prairie. Land, 11(8), 1135. https://doi.org/10.3390/land11081135