


Floristic Composition, Diversity, Palatability, and Forage Availability of Forest Rangelands in the Southern Mediterranean Region of Northern Morocco


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
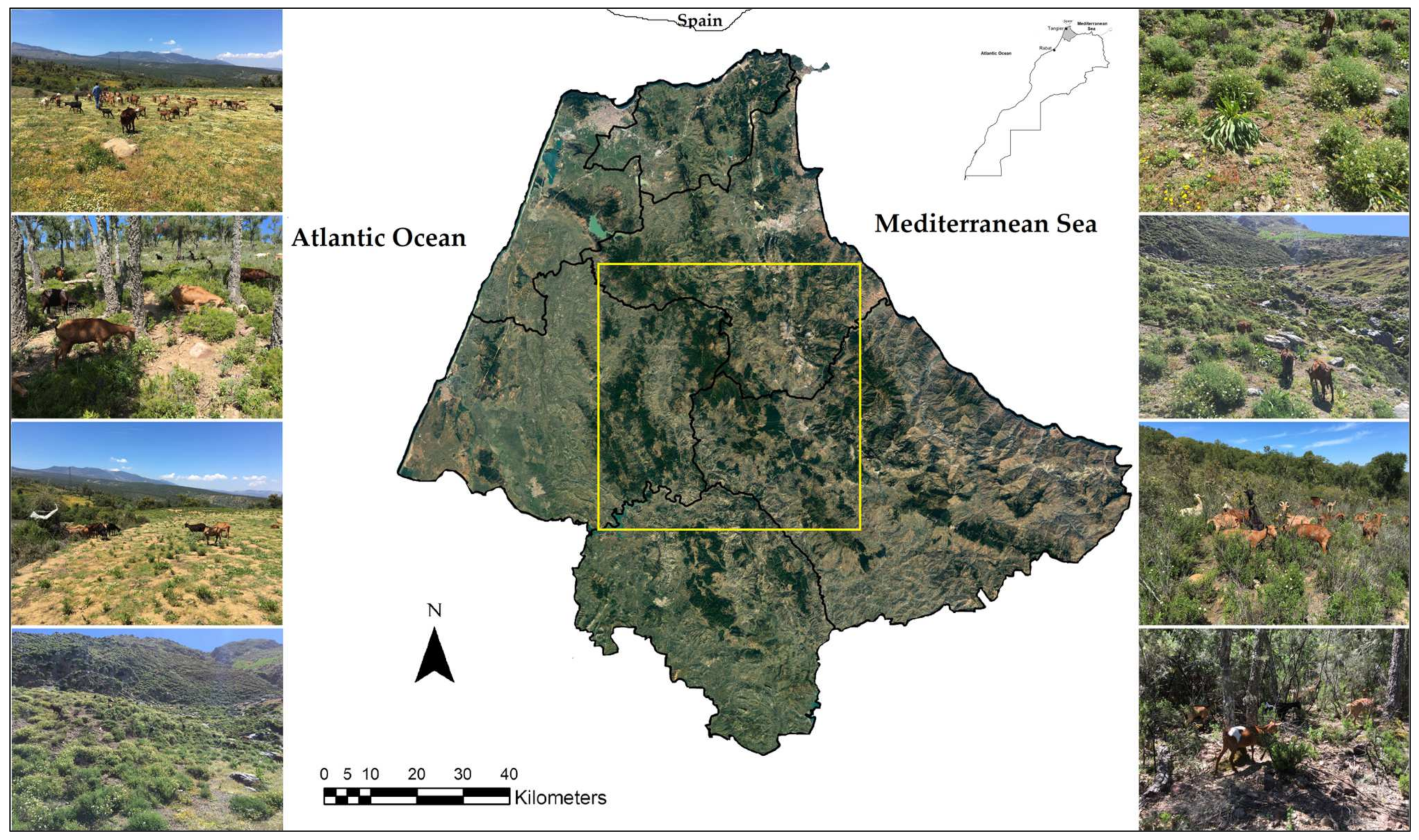
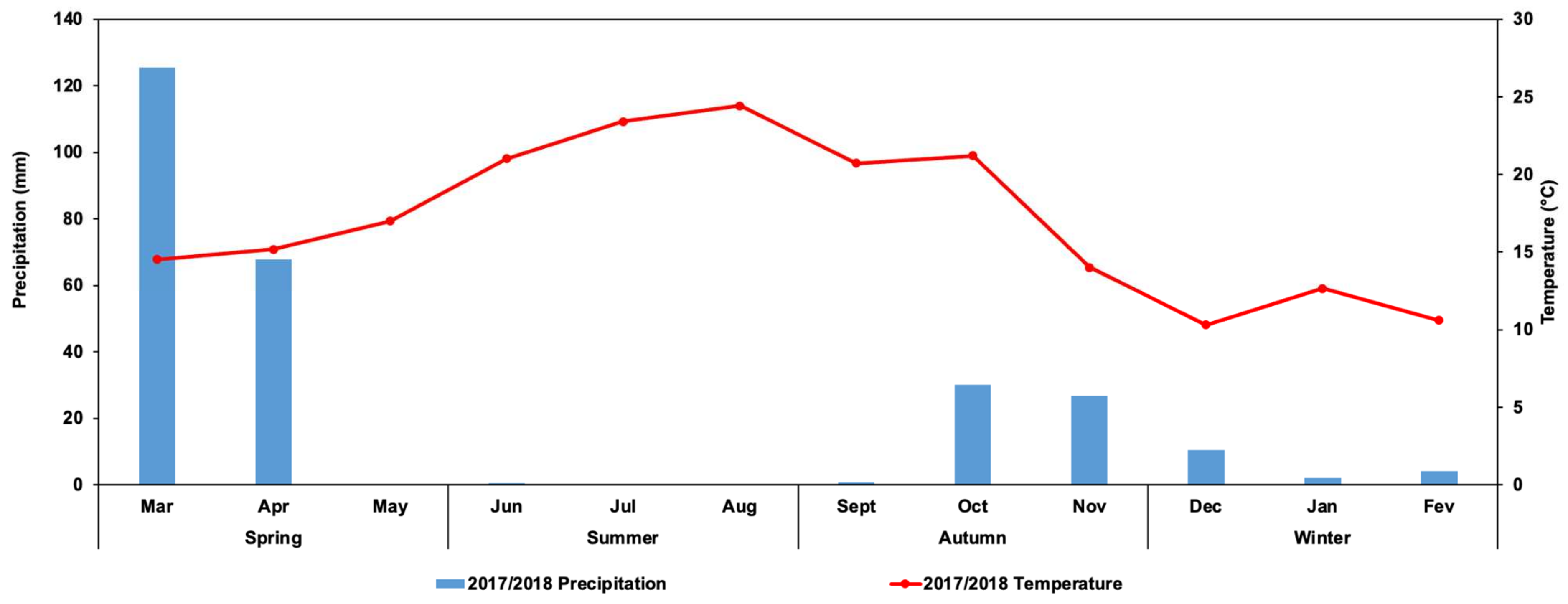
2.1. Study Area
2.2. Data Collection
2.2.1. Botanical Study
2.2.2. Palatability Index
2.2.3. Forage Availability Measurement
2.3. Statistical Analysis
3. Results and Discussion
3.1. Brief Paleoecological Analysis
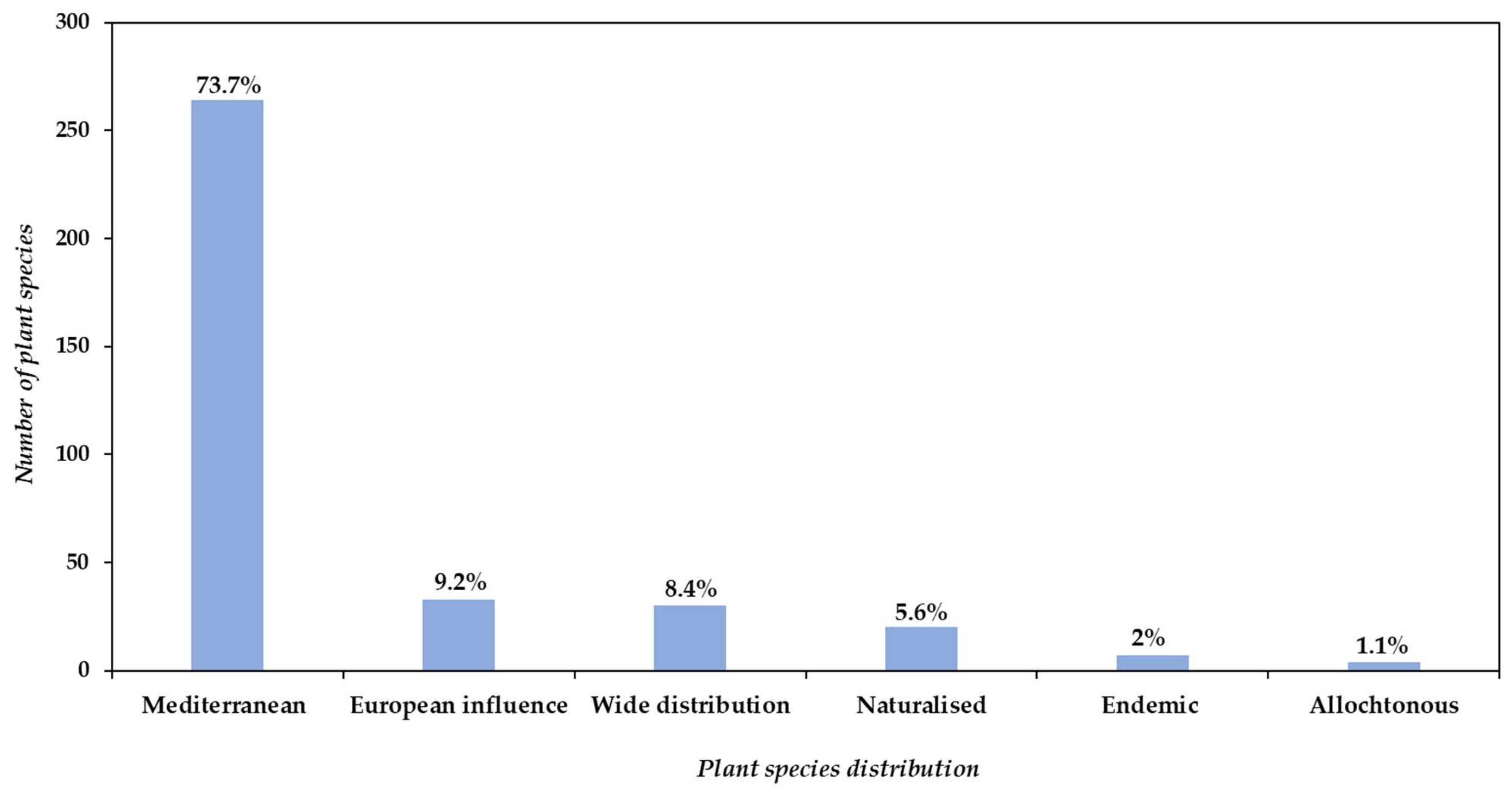
3.2. Floristic Composition of the Study Area
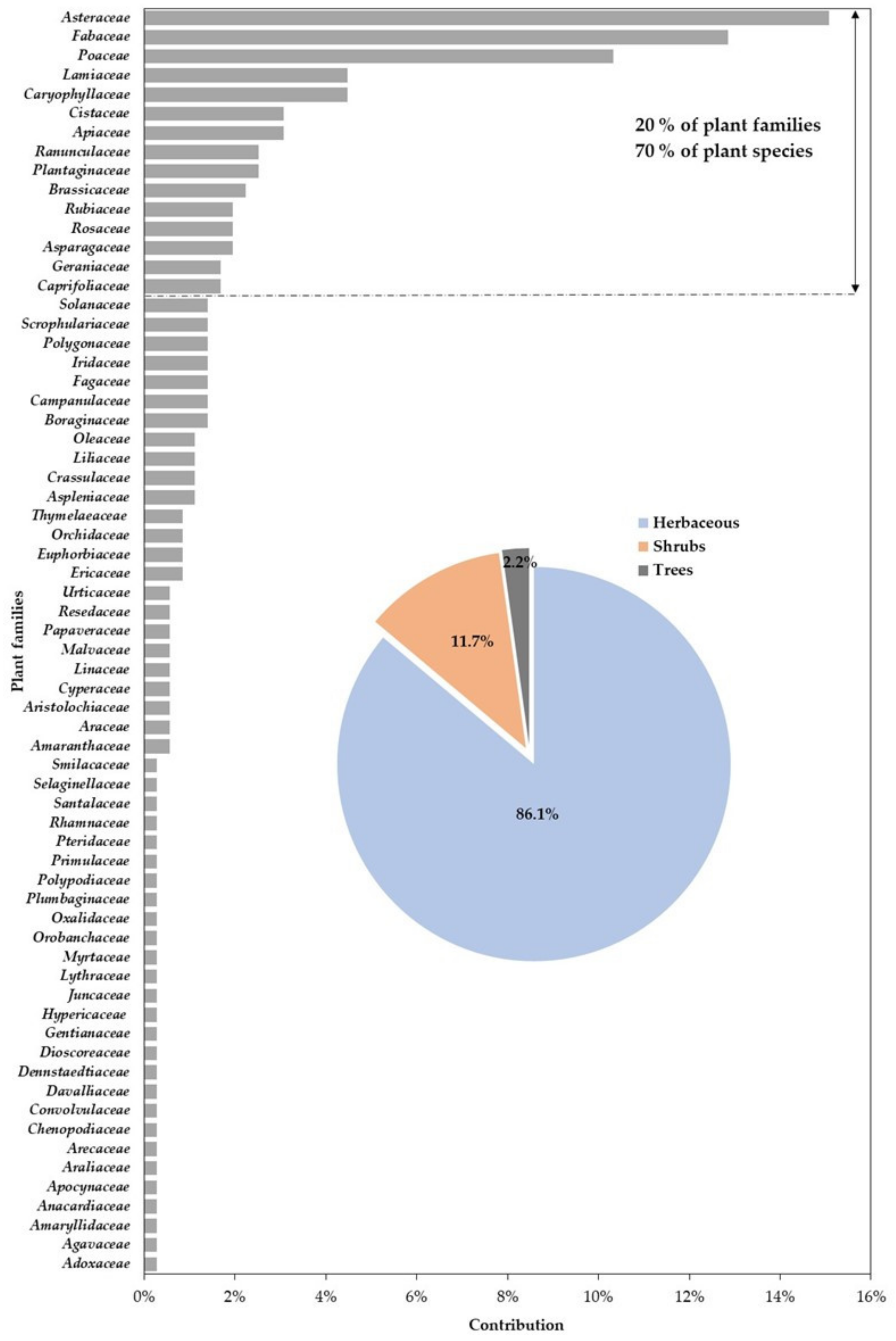
3.2.1. Taxonomic Diversity
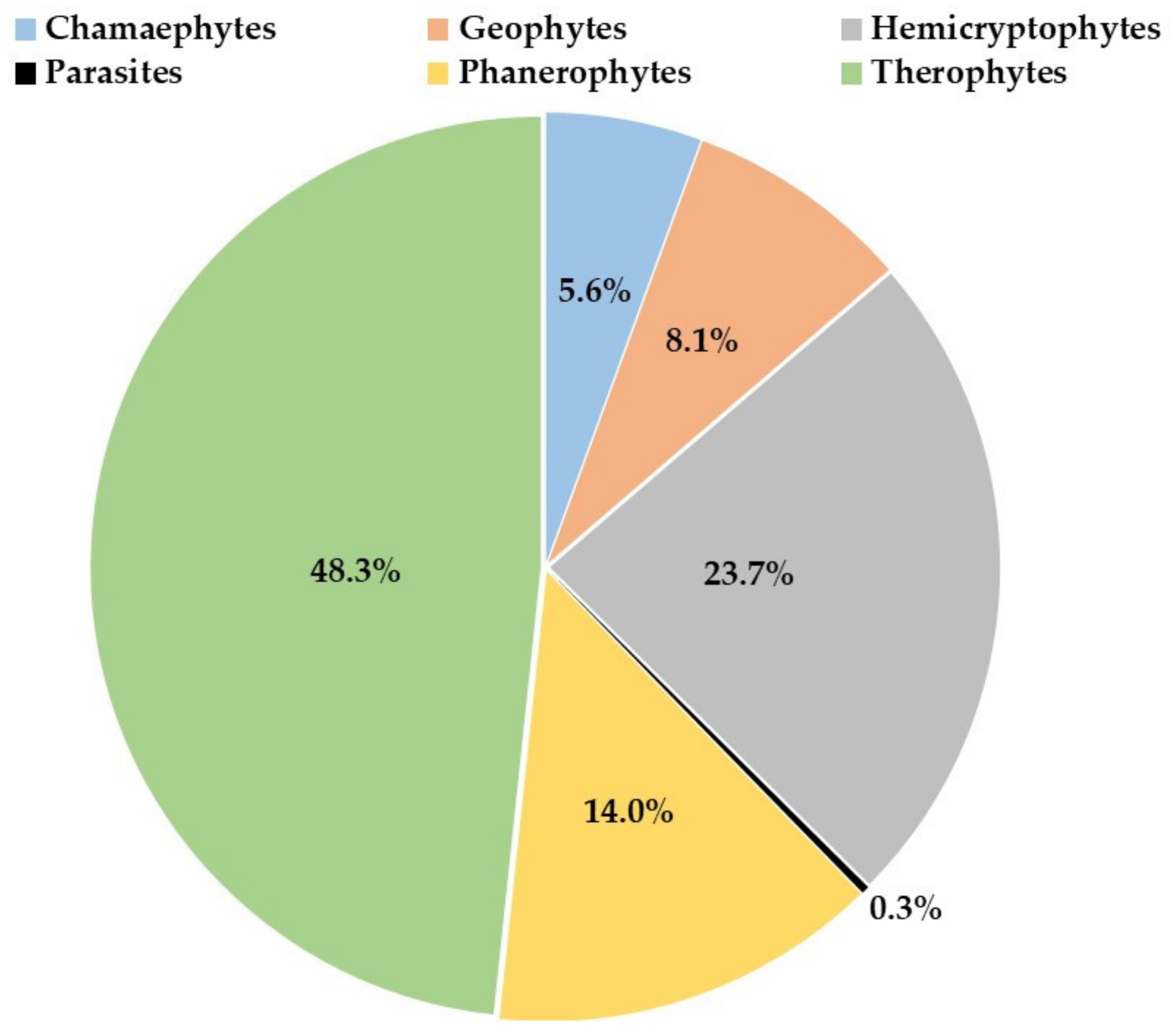
3.2.2. Biological Spectrum and Perturbation Index
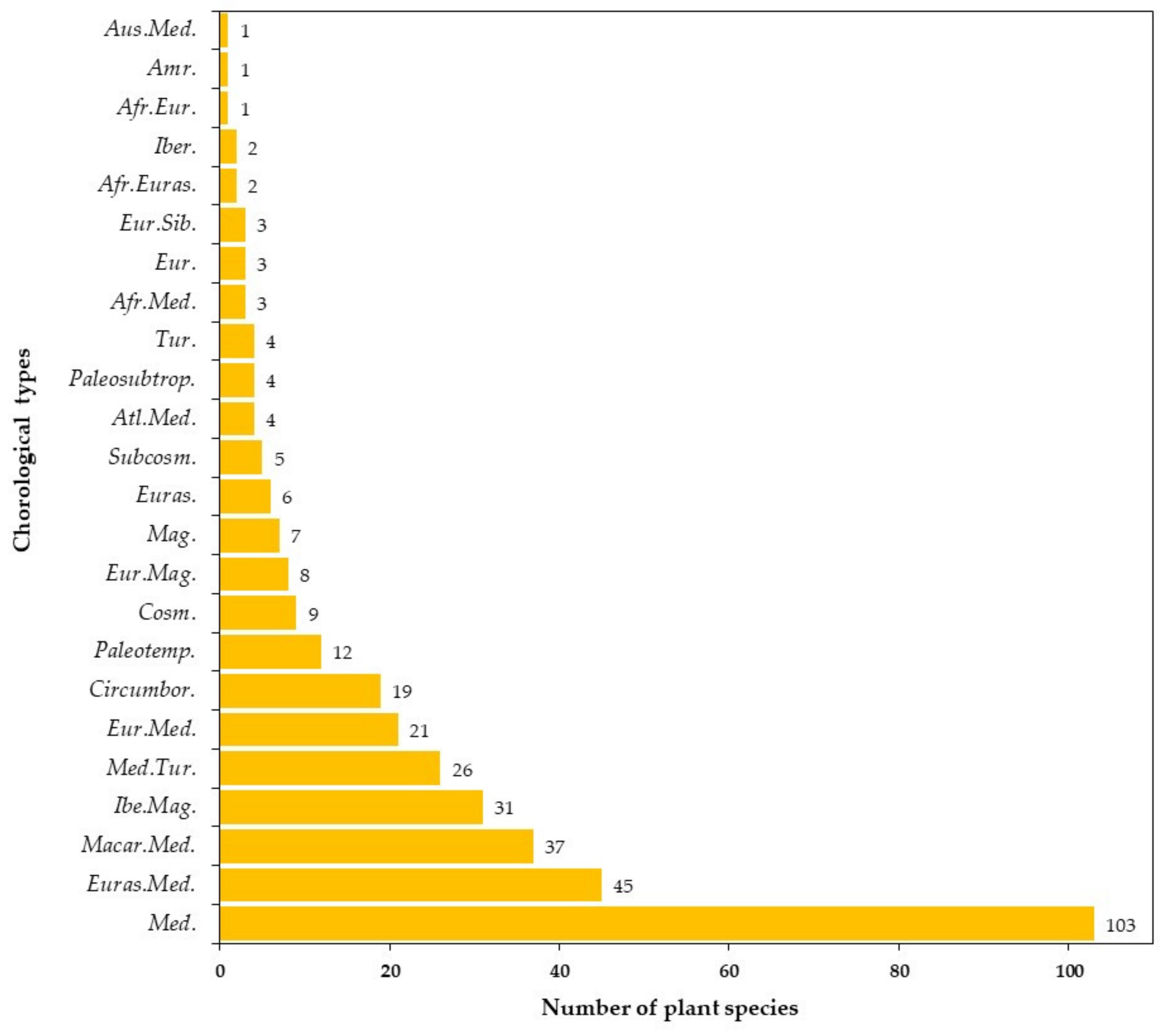
3.2.3. Phytogeographical and Chorological Analysis
3.3. Palatability and Biomass Production
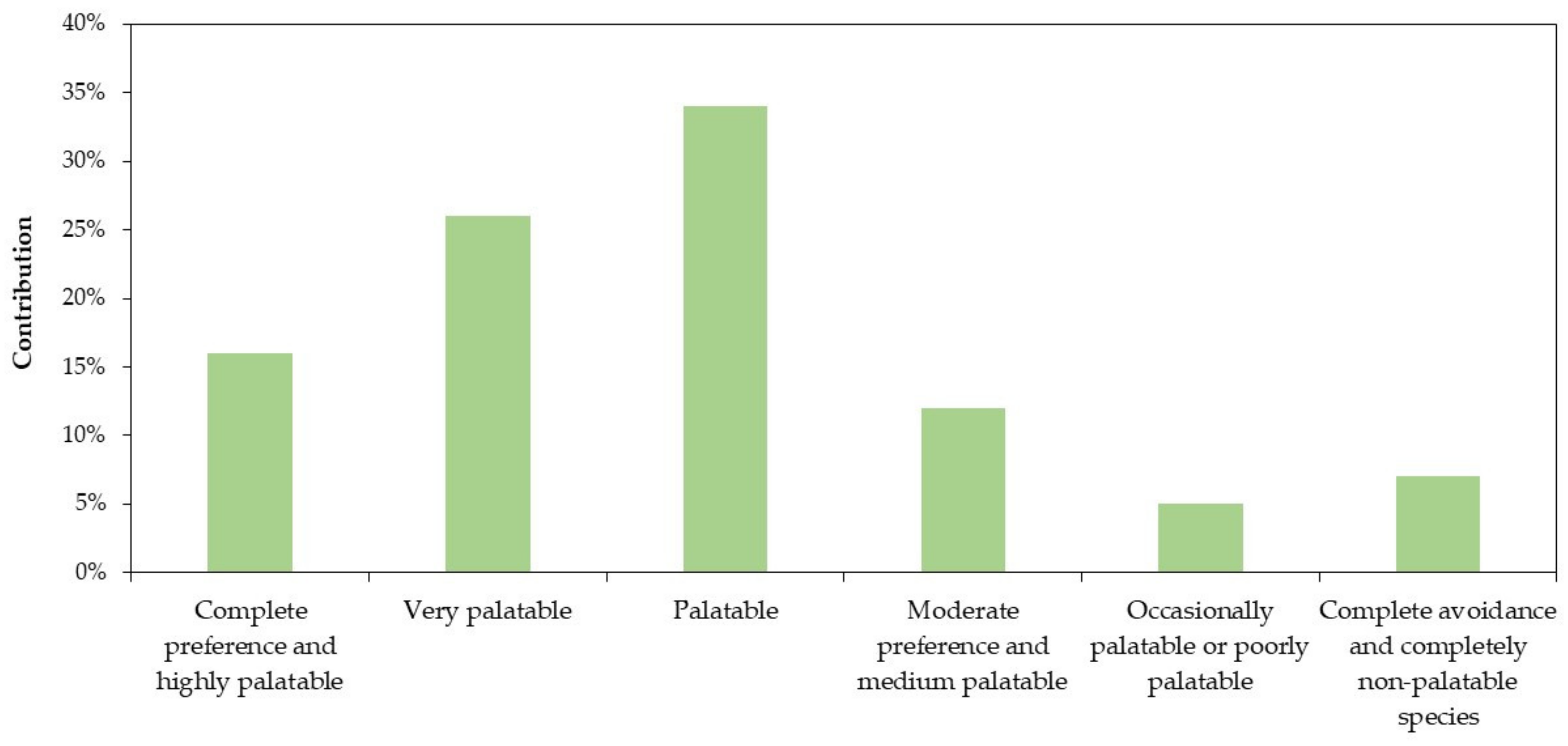
3.3.1. Plant Species Palatability
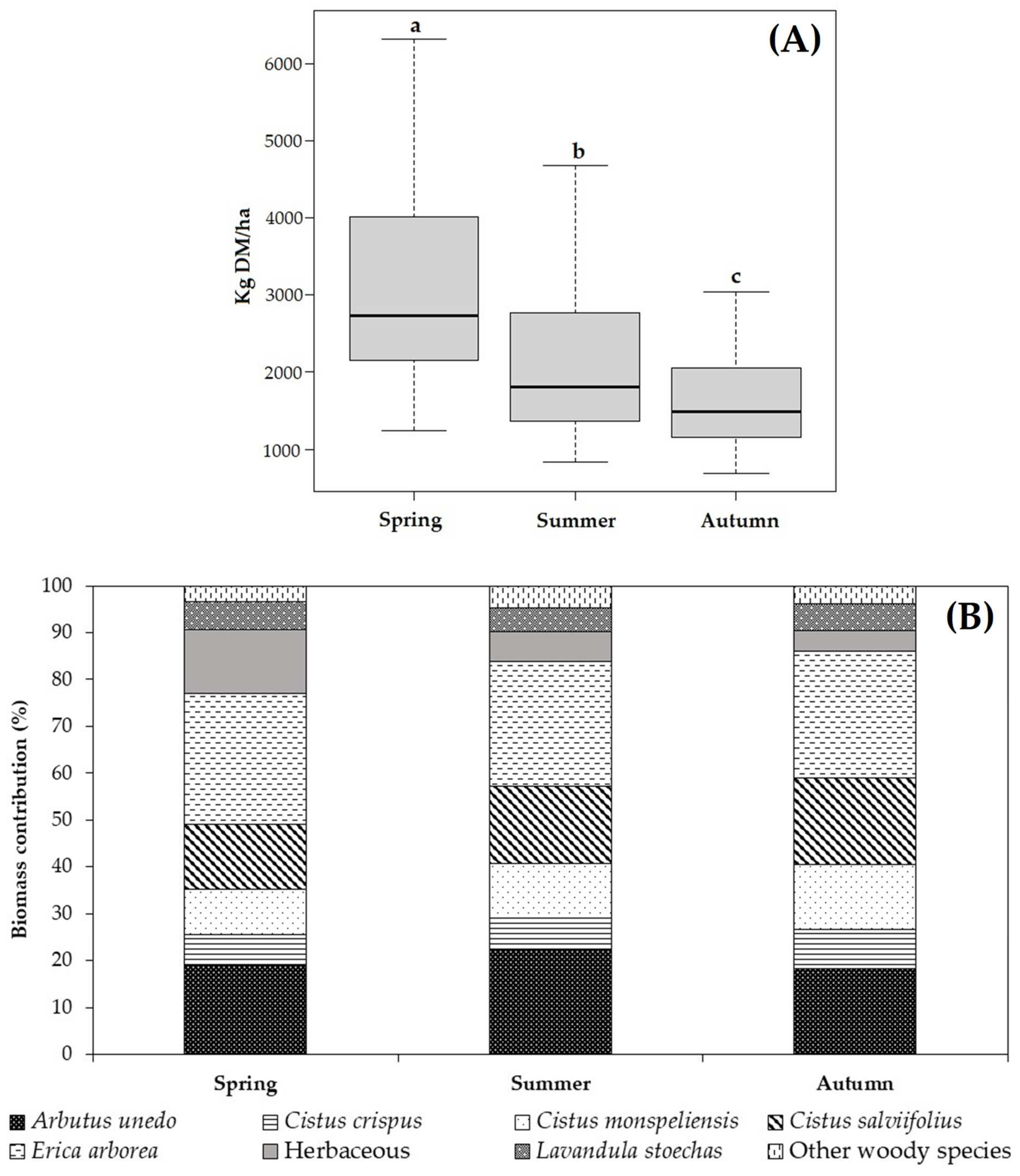
3.3.2. Forage Availability
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MAPMDREF (Ministère de L’Agriculture, de la Pêche Maritime, du Développement Rural et des Eaux et Forêts). Forêts en Chiffre. Département des Eaux et Forêts. 2018. Available online: http://www.eauxetforets.gov.ma/ForetsMarocaines/ForetsChiffres/Pages/Forets-En-Chiffres.aspx (accessed on 20 April 2021).
- Chebli, Y.; El Otmani, S.; Chentouf, M.; Hornick, J.L.; Cabaraux, J.F. Temporal variations in chemical composition, in vitro digestibility, and metabolizable energy of plant species browsed by goats in southern Mediterranean forest rangeland. Animals 2021, 11, 1441. [Google Scholar] [CrossRef] [PubMed]
- Chebli, Y.; El Otmani, S.; Hornick, J.L.; Bindelle, J.; Cabaraux, J.F.; Chentouf, M. Estimation of Grazing Activity of Dairy Goats Using Accelerometers and Global Positioning System. Sensors 2022, 22, 5629. [Google Scholar] [CrossRef] [PubMed]
- Direction Régionale de l’Agriculture de Tanger-Tétouan (DRATT). Monographie de la Région du Nord du Maroc. Rapports et Données Statistiques sur l’Agriculture et l’Elevage. 2017. Available online: https://www.agriculture.gov.ma/fr/region/tanger-tetouan-al-hoceima/ (accessed on 8 July 2019).
- Chebli, Y.; El Otmani, S.; Chentouf, M.; Hornick, J.-L.; Bindelle, J.; Cabaraux, J.F. Foraging behavior of goats browsing in southern Mediterranean forest rangeland. Animals 2020, 10, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chentouf, M.; Zantar, S.; Doukkali, M.R.; Farahat, L.B.; Jouamaa, A.; Aden, H. Performances techniques et économiques des élevages caprins dans le nord du Maroc. In Economic, Social and Environmental Sustainability in Sheep and Goat Production Systems; Bernués, A., Boutonnet, J., Casasús, I., Chentouf, M., Gabiña, D., Joy, M., Eds.; CIHEAM: Zaragoza, Spain, 2011; pp. 151–156. [Google Scholar]
- Cinar, S.; Abdullayev, A.; Esenov, N.; Karadag, Y. Determination of botanical composition, hay yield and forage quality of some natural rangelands in Kyrgyzstan’s chuy region. Appl. Ecol. Environ. Res. 2020, 18, 401–416. [Google Scholar] [CrossRef]
- Harris, S.E. Cyprus as a degraded landscape or resilient environment in the wake of colonial intrusion. Proc. Natl. Acad. Sci. USA 2012, 109, 3670–3675. [Google Scholar] [CrossRef] [Green Version]
- Lovreglio, R.; Meddour-Sahar, O.; Leone, V. Goat grazing as a wildfire prevention tool: A basic review. IForest 2014, 7, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Chebli, Y.; Chentouf, M.; Ozer, P.; Hornick, J.L.; Cabaraux, J.F. Forest and silvopastoral cover changes and its drivers in northern Morocco. Appl. Geogr. 2018, 101, 23–35. [Google Scholar] [CrossRef]
- Louhaichi, M.; Gamoun, M.; Hassan, S.; Abdallah, M.A.B. Characterizing biomass yield and nutritional value of selected indigenous range species from arid Tunisia. Plants 2021, 10, 2031. [Google Scholar] [CrossRef]
- Palahi, M.; Mavsar, R.; Gracia, C.; Birot, Y. Mediterranean forests under focus. Int. For. Rev. 2008, 10, 676–688. [Google Scholar] [CrossRef]
- Quézel, P.; Médail, F.; Loisel, R.; Barbero, M.; Quézel, P.; Médail, F.; Loisel, R.; Barbero, M. Biodiversity and conservation of forest species in the Mediterranean basin Biodiversity and conservation of forest species in the Mediterranean basin. Unasylva 1999, 50, 21–28. Available online: http://www.fao.org/docrep/x1880e/x1880e05.htm (accessed on 14 April 2022).
- Gauquelin, T.; Michon, G.; Joffre, R.; Duponnois, R.; Génin, D.; Fady, B.; Bou Dagher-Kharrat, M.; Derridj, A.; Slimani, S.; Badri, W.; et al. Mediterranean forests, land use and climate change: A social-ecological perspective. Reg. Environ. Chang. 2018, 18, 623–636. [Google Scholar] [CrossRef]
- Walas, Ł.; Taib, A. Environmental regionalization and endemic plant distribution in the Maghreb. Environ. Monit. Assess. 2022, 194, 100. [Google Scholar] [CrossRef]
- Meddour, R.; Sahar, O. Floristic diversity of Quercus suber woodlands in Kabylia (north eastern Algeria). Checklist and brief analysis. Bot. Complut. 2022, 46, 1–21. [Google Scholar] [CrossRef]
- Moriondo, M.; Good, P.; Durao, R.; Bindi, M.; Giannakopoulos, C.; CorteReal, J. Potential impact of climate change on fire risk in the Mediterranean area. Clim. Res. 2006, 31, 85–95. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; GarciaGonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolstrom, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Niirou, N.; Gupta, A. Life forms classification and biological spectrum in natural and human impacted ecosystems of senapati district, manipur, india. Ecol. Environ. Conserv. 2020, 26, S307–S313. [Google Scholar]
- Emberger, L.; Maire, R. Catalogue des Plantes du Maroc; Minerva: Alger, Algeria, 1941; Volume IV. [Google Scholar]
- Maire, R. Flore de l’Afrique du Nord; 16 Tomes; Lechevalier: Paris, France, 1952–1987. [Google Scholar]
- Fennane, M.; Ibn Tattou, M. Observations sur la flore vasculaire endémique, rare ou menacée du Maroc. Flora Mediterr. 1999, 9, 113–124. [Google Scholar]
- Benabid, A. Flore et diversité des écosystèmes du Maroc. In Évaluation et Préservation de la Biodiversité; Édition Ibis Press: Paris, France, 2000; p. 236. [Google Scholar]
- Hebrard, J.P.; Loisel, R.; Roux, C.; Goumila, H.; Bouni, G. Incidence of clearing on phanerogamic and cryptogamic vegetation in South Eastern France disturbance indices. In Ext. de Technique et Documentation; Intercept Ltd.: Bambous, Mauritius, 1995; pp. 747–758. [Google Scholar]
- Le Houérou, H.N. Les Pâturages Naturels de la Tunisie Aride et Désertique; Institut des Sciences Economiques Appliquées: Tunis, Tunisia, 1962. [Google Scholar]
- Le Houérou, H.N. Improvements of Natural Pastures and Fodder Resources. In Report to the Government of Libya; EPTA Rep.; FAO: Rome, Italy, 1979. [Google Scholar]
- Le Houérou, H.N.; Ionesco, T. Appétabilité des Espèces Végétales de la Tunisie Steppique; AG-TUN 71/ 525; FAO: Rome, Italy, 1973. [Google Scholar]
- Sarson, M.; Salmon, P. Appétabilité de Certaines Espèces Vivaces Spontanées ou Introduites au Maroc et en Afrique du Nord; FAO: Rabat, Maroc; Rome, Italy, 1976. [Google Scholar]
- Benabid, A. Les écosystèmes forestiers, préforestiers et presteppiques du Maroc: Diversité, répartition biogéographique et problèmes posés par leur aménagement. Forêt Méditerranéenne 1985, 7, 53–64. [Google Scholar]
- Grovel, R. La préservation des forêts du Rif centro-occidental: Un enjeu de développement de la montagne rifaine/Preservation of forests in the central western Rif massif: A key element in the development of the Rif Mountains. Rev. Géographie Alp. 1996, 84, 75–94. [Google Scholar] [CrossRef]
- SBF (Bulletin de la Socieéteé Botanique de Franc); Études Sur La Végétation Du Maroc. Comptes-Rendus Herborisations Mémoires Sur Maroc 1921, 68, 9–152. [CrossRef] [Green Version]
- Cheddadi, R.; Bouaissa, O.; Rhoujjati, A.; Dezileau, L. Environmental changes in the Moroccan western Rif mountains over the last 9,000 years. Rev. L’association Française Pour L’étude Quat. 2016, 27, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Moukrim, S.; Lahssini, S.; Rifai, N.; Menzou, K.; Mharzi-Alaoui, H.; Labbaci, A.; Rhazi, M.; Wahby, I.; El Madihi, M.; Rhazi, L. Modélisation de la distribution potentielle de Cedrus atlantica Manetti au Maroc et impacts du changement climatique. Bois Trop. 2020, 344, 3–16. [Google Scholar] [CrossRef]
- Montalembert, M.R. L’économie des Produits Forestiers au Maroc: Bilan et Pespectives. Ph.D. Thesis, Faculté de droits et des sciences économiques, Université de Montpellier, Montpellier, France, 1975; p. 341. [Google Scholar]
- Taiqui, L. La dégradation écologique au Rif marocain: Nécessités d’une nouvelle approche. Mediterranea 1997, 16, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Zhao, W.; Wang, J.; Ferreira, C.S.S. Precipitation drives the floristic composition and diversity of temperate grasslands in China. Glob. Ecol. Conserv. 2021, 32, e01933. [Google Scholar] [CrossRef]
- Bertiller, M.B.; Beeskow, A.M.; Coronato, F. Seasonal environmental variation, and plant phenology in arid Patagonia (Argentina). J. Arid Environ. 1991, 21, 1–11. [Google Scholar] [CrossRef]
- Seghieri, J.; Galle, S. Run-on contribution to a Sahelian two-phase mosaic system: Soil water regime and vegetation life cycles. Acta Oecol. 1999, 20, 209–217. [Google Scholar] [CrossRef]
- Ni, J. Plant functional types and climate along a precipitation gradient in temperate grasslands, north-east China and south-east Mongolia. J. Arid Environ. 2003, 53, 501–516. [Google Scholar] [CrossRef] [Green Version]
- Loeser, M.R.R.; Sisk, T.D.; Crews, T.E. Impact of grazing intensity during drought in an Arizona grassland. Conserv. Biol. 2007, 21, 87–97. Available online: http://www.jstor.org/stable/4124645 (accessed on 14 April 2022). [CrossRef]
- Cheng, Y.; Tsubo, M.; Ito, T.Y.; Nishihara, E.; Shinoda, M. Impact of rainfall variability and grazing pressure on plant diversity in Mongolian grasslands. J. Arid Environ. 2011, 75, 471–476. [Google Scholar] [CrossRef]
- Kinugasa, T.; Tsunekawa, A.; Shinoda, M. Increasing nitrogen deposition enhances post-drought recovery of grassland productivity in the Mongolian steppe. Oecologia 2012, 170, 857–865. [Google Scholar] [CrossRef]
- Zemmar, N.; M’hammedi Bouzina, M.; Ababou, A.; Hedidi, D. Analysis of the floristic diversity in a southern Mediterranean ecosystem. Case of Bissa forest, Chlef (Algeria). Bot. Complut. 2020, 44, 19–28. [Google Scholar] [CrossRef]
- Gamoun, M.; Louhaichi, M. Botanical Composition and Species Diversity of Arid and Desert Rangelands in Tataouine, Tunisia. Land 2021, 10, 313. [Google Scholar] [CrossRef]
- Al-Turki, T.A. A prelude to the study of the flora of Jabal Fayfa in Saudi Arabia. Kuwait J. Sci. Eng. 2004, 31, 77–145. [Google Scholar]
- Sohair, T.H.; Sheded, M.G.; Badry, M.O. Floristic Composition of Some Riverian Islands at Qena Governorate-Egypt. Egypt. J. Bot. 2012, 299–322. [Google Scholar]
- Abbas, A.M.; Al-Kahtani, M.A.; Alfaifi, M.Y.; Elbehairi, S.E.I.; Badry, M.O. Floristic Diversity and Phytogeography of JABAL Fayfa: A Subtropical Dry Zone, South-West Saudi Arabia. Diversity 2020, 12, 345. [Google Scholar] [CrossRef]
- Vrahnakis, M.; Fotiadis, G.; Pantera, A.; Papadopoulos, A.; Papanastasis, V. Floristic diversity of Valonia oak silvopastoral woodlands in Greece. Agrofor. Syst. 2014, 88, 877–893. [Google Scholar] [CrossRef]
- Rodríguez-Guitián, M.A.; Amigo, J.; Real, C.; Romero Franco, R. Cork oak forests in the NW Iberian Peninsula: Phytosociological reassessment and new proposals. Plant Biosyst. 2021, 155, 944–961. [Google Scholar] [CrossRef]
- Chawla, A.; Rajkumar, S.; Singh, K.N.; Lal, B.; Singh, R.D.; Thukral, A.K. Plant species diversity along an altitudinal gradient of Bhabha valley in Western Himalaya. J. Mater. Sci. 2008, 5, 157–177. [Google Scholar] [CrossRef]
- Nogués-Bravo, D.; Araújo, M.B.; Romdal, T.; Rahbek, C. Scale effects and human impact on the elevational species richness gradients. Nature 2008, 453, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Kulaib, J.F. Natural Grazing Vegetation in Two Areas of the Egyptian North West Coast Range Lands. Ph.D. Thesis, Faculty of Agriculture, Cairo University, Cairo, Egypt, 2008; pp. 29–113. [Google Scholar]
- Abusaief, H.M.A. Life forms and rangeland for many habitats of Jarjar oma in Al-Jabal Al-Akhdar on Mediterranean Sea. J. Am. Sci. 2013, 9, 236–249. [Google Scholar]
- Burju, T.; Hundera, K.; Kelbessa, E. Floristic composition and structural analysis of Jibat humid afromontane forest, West Shewa zone, Oromia national regional state, Ethiopia. Ethiop. J. Educ. Sci. 2013, 8, 11–33. Available online: https://ejhs.ju.edu.et/index.php/ejes/article/view/681 (accessed on 15 April 2022).
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; with 47 Tables, 3rd ed.; Springer Science and Business Media: Cham, Switzerland, 2003; pp. 1–349. [Google Scholar]
- Ghafari, S.; Ghorbani, A.; Moameri, M.; Mostafazadeh, R.; Bidarlord, M. Composition and structure of species along altitude gradient in Moghan-Sabalan rangelands, Iran. J. Mt. Sci. 2018, 15, 1209–1228. [Google Scholar] [CrossRef]
- Arévalo, J.R.; Delgado, J.D.; Otto, R.; Naranjoc, A.; Salasd José, M.; Fernández-Palaciosa, M. Distribution of alien vs. native plant species in roadside communities along an altitudinal gradient in Tenerife and Gran Canaria (Canary Islands). Perspect. Plant Ecol. Evol. Syst. 2005, 7, 185–202. [Google Scholar] [CrossRef]
- Arila, K.E.; Gupta, A. Life-forms and biological spectrum along the altitudinal gradient in Montane Forests of Senapati district of Manipur in North-East India. Pleione 2016, 10, 80–89. [Google Scholar]
- El-Barasi, Y.M.; Barrani, M.W.; El-Amrouni, A.O.; Mohamad, N.F. Check list of flora and vegetation on South El-Marj zone: South El-Jabal El-Akhadar Libya. Ann. Fac. Eng. Hunedoara 2011, 9, 141. [Google Scholar]
- Raunkiaer, C. The Life Form of Plants and Statistical Plant Geography; Collected Papers; Clarendon Press: Oxford, UK, 1934. [Google Scholar]
- Aafi, A.; Achhal El Kadmiri, A.; Benabid, A.; Rochdi, M. Richesse et diversité floristique de la suberaie de la Mamora (Maroc). Acta Bot. Malacit. 2005, 30, 127–138. [Google Scholar] [CrossRef]
- Ouelmouhoub, S.; Benhouhou, S. Evolution floristique des suberaies incendiées dans la région d’El Kala (nord-est Algérie). Ecol. Mediterr. 2007, 33, 85–94. [Google Scholar] [CrossRef]
- Sarmoum, M.; Feddag, F.; Masloub, A.; Belkaid, B. Diagnostic de l’état actuel de la suberaie du Parc National de Theniet El Had (wilaya Tissemsilt). In Proceedings of the Journées d’étude sur la réhabilitation des subéraies incendiées et reboisements, Recueil des résumes, Tlemcen, Algeria, 17–18 January 2013. [Google Scholar]
- Benkhettou, A.; Azouzi, B.; Djili, K.; Benkhettou, M.; Zedek, M.; Saadi, R. Diversité floristique du massif du Nador en zone steppique (Tiaret, Algérie). Europ. Sci. J. 2015, 11, 401–419. Available online: https://eujournal.org/index.php/esj/article/download/6008/5791 (accessed on 15 April 2022).
- Le Houerou, H.N. L’impact de l’homme et des animaux sur la forêt méditerranéenne. Forêt Medit. 1980, 2, 155–174. [Google Scholar]
- Le Houerou, H.N. Bioclimatologie et Biogéographie des steppes arides du Nord de l’Afrique: Diversité biologique, développement durable et désertisation. Options Mediterr. 1995, 15–31. [Google Scholar]
- Quezel, P. Réflexion sur L’évolution de la Flore et de la Végétation au Maghreb Méditerranéen; Ibis Press: Paris, France, 2000. [Google Scholar]
- Quezel, P.; Medail, F. Ecologie et Biogéographie des Forêts du Bassin Méditerranéen; Elsevier: Paris, France, 2003; 571p. [Google Scholar]
- Benabadji, N.; Aboura, R.; Benchouk, F. La régression des steppes méditerranéennes: Le cas d’un faciès à Lygeum spartum L. d’Oranie (Algérie). Rev. Ecol. Mediterr. 2009, 35, 75–90. [Google Scholar] [CrossRef]
- Barbero, M.; Quezel, P.; Loisel, R. Les apports de la phytoécologie dans l’interprétation des changements et perturbations induites par l’homme sur les écosystèmes forestiers méditerranéens. Forêt Médit. 1990, 12, 194–215. [Google Scholar]
- Whitaker, R.H. Communities and Ecosystems; Macmillan Co. Coiinc: New York, NY, USA, 1975. [Google Scholar]
- Belhacini, F. Contribution à une Étude Floristique et Biogéographique des Matorrals du Versant sud de la Région de Tlemcen. Master’s Thesis, Université Aboubakr Belkaïd Tlemcen, Chetouane, Algeria, 2011. [Google Scholar]
- Larbi, R. Analyse de la Diversité Floristique et de la Phytodynamique de la Série de Végétation à Cedrus Atlantica au Djurdjura Centro- Méridional (Secteur de Tikjda). Master’s Thesis, Université Mouloud Mameri Tizi Ouzou, Tizi Ouzou, Algeria, 2015. [Google Scholar]
- Chebli, Y.; EL Otmani, S.; Elame, F.; Moula, N.; Chentouf, M.; Hornick, J.L.; Cabaraux, J.F. Silvopastoral system in Morocco: Focus on their importance, strategic functions, and recent changes in the Mediterranean side. Sustainability 2021, 131, 10744. [Google Scholar] [CrossRef]
- Mele, C.; Medagli, P.; Acogli, R.; Beccarisi, L.; Albano, A.; Marchiori, S. Flora of Salento (Apulia, Southeastern Italy): An annotated checklist. Fl. Medit. 2006, 16, 193–245. [Google Scholar]
- Simonson, W.D.; Allen, H.D. Cork oak (Quercus suber L.) forests of western Mediterranean mountains: A plant community comparison. Ecol. Mediterr. 2014, 40, 35–48. [Google Scholar] [CrossRef]
- Olff, H.; Ritchie, M.E. Effects of herbivores on grassland plant diversity. Trend. Ecol. Evol. 1998, 13, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Curtin, C.G. Livestock grazing, rest, and restoration in arid landscapes. Conserv. Biol. 2002, 16, 840–842. Available online: https://www.jstor.org/stable/3061232 (accessed on 15 April 2022). [CrossRef]
- Pacala, S.W.; Crawley, M.J. Herbivores and plant diversity. Am. Nat. 1992, 140, 243–260. Available online: https://www.jstor.org/stable/2462608 (accessed on 15 April 2022). [CrossRef]
- Cory, V.L. Methods of determining forage preferences of stock. Ecology 1930, 11, 760–763. [Google Scholar] [CrossRef]
- Casey, N.H.; Van Niekerk, W.A. The Boer Goat I. Origin, adaptability, performance testing, reproduction, and milk production. Small Rumin. Res. 1988, 1, 291–302. [Google Scholar] [CrossRef]
- Molina-Alcaide, E.; Yáñez-Ruiz, D. Potential use of olive by-products in ruminant feeding: A review. Anim. Feed Sci. Technol. 2008, 147, 247–264. [Google Scholar] [CrossRef]
- Archer, S.; Smeins, F.E. Ecosystem-level processes. In Grazing Management: An Ecological Perspective; Heitschmidt, R.K., Stuth, J.W., Eds.; Timber Press: Portland, OR, USA, 1991; pp. 109–139. [Google Scholar]
- El Otmani, S.; Chebli, Y.; Chentouf, M.; Hornick, J.L.; Cabaraux, J.F. Effects of Olive Cake and Cactus Cladodes as Alternative Feed Resources on Goat Milk Production and Quality. Agriculture 2021, 11, 3. [Google Scholar] [CrossRef]
- Chebli, Y.; El Otmani, S.; Cabaraux, J.-F.; Keli, A.; Chentouf, M. Using GPS Tracking Collars and Sensors to Monitor the Grazing Activity of Browsing Goats in Forest Rangeland. Eng. Proc. 2022, 27, 37. [Google Scholar] [CrossRef]
- Chebli, Y.; El Otmani, S.; Hornick, J.-L.; Keli, A.; Bindelle, J.; Chentouf, M.; Cabaraux, J.-F. Using GPS Collars and Sensors to Investigate the Grazing Behavior and Energy Balance of Goats Browsing in a Mediterranean Forest Rangeland. Sensors 2022, 22, 781. [Google Scholar] [CrossRef]
- Chebli, Y.; Chentouf, M.; Hornick, J.L.; Cabaraux, J.F. Biomass production and use of silvopastoral areas in Rif Mountains of Morocco. Options Mediterr. 2016, 116, 309–312. [Google Scholar]
- Aharon, H.; Henkin, Z.; Ungar, E.D.; Kababya, D.; Baram, H.; Perevolotsky, A. Foraging behaviour of the newly introduced Boer goat breed in a Mediterranean woodland: A research observation. Small Rumin. Res. 2007, 69, 144–153. [Google Scholar] [CrossRef]
- Ventura-Cordero, J.; González-Pech, P.; Sandoval-Castro, C.A.; Torres-Acosta, J.F.J.; Tun-Garrido, J. Feed resource selection by Criollo goats browsing a tropical deciduous forest. Anim. Prod. Sci. 2017, 58, 2314–2320. [Google Scholar] [CrossRef]
- Chebli, Y.; Boulaich, H.; Chentouf, M.; El Otmani, S. Seasonal feeding behavior of goat kids browsing in Mediterranean forest rangeland of Bouhachem Natural Park, Northern Morocco. Livest. Res. Rural. Dev. 2022, 34, 107. [Google Scholar]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chebli, Y.; Chentouf, M.; Cabaraux, J.-F.; El Otmani, S. Floristic Composition, Diversity, Palatability, and Forage Availability of Forest Rangelands in the Southern Mediterranean Region of Northern Morocco. Land 2023, 12, 215. https://doi.org/10.3390/land12010215
Chebli Y, Chentouf M, Cabaraux J-F, El Otmani S. Floristic Composition, Diversity, Palatability, and Forage Availability of Forest Rangelands in the Southern Mediterranean Region of Northern Morocco. Land. 2023; 12(1):215. https://doi.org/10.3390/land12010215
Chicago/Turabian StyleChebli, Youssef, Mouad Chentouf, Jean-François Cabaraux, and Samira El Otmani. 2023. "Floristic Composition, Diversity, Palatability, and Forage Availability of Forest Rangelands in the Southern Mediterranean Region of Northern Morocco" Land 12, no. 1: 215. https://doi.org/10.3390/land12010215
APA StyleChebli, Y., Chentouf, M., Cabaraux, J. -F., & El Otmani, S. (2023). Floristic Composition, Diversity, Palatability, and Forage Availability of Forest Rangelands in the Southern Mediterranean Region of Northern Morocco. Land, 12(1), 215. https://doi.org/10.3390/land12010215