

Post-Tin-Mining Agricultural Soil Regeneration Using Local Organic Amendments Improve Nitrogen Fixation and Uptake in a Legume–Cassava Intercropping System
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Treatment
2.2. Plant Sampling and Analyses
2.3. Soil Sampling and Analyses
2.4. Calculation
2.4.1. Estimation of Nitrogen Fixation in Centrosema
2.4.2. Estimation of Fixed Amount of Nitrogen by Centrosema
2.4.3. Crop Nitrogen Uptake
2.4.4. Partial Balance of N
2.5. Statistical Analyses
3. Results
3.1. Nitrogen Isotope Discrimination (δ15N), Nitrogen Derived from N2–Fixation (%NDFA), and Nitrogen Fixation (N2–Fixation) of Centrosema
3.2. Nitrogen Content (%N) and Nitrogen Uptake (N Uptake) in Centrosema
3.3. Nitrogen Content (%N) and Nitrogen Uptake (N Uptake) in Plant Parts of Cassava
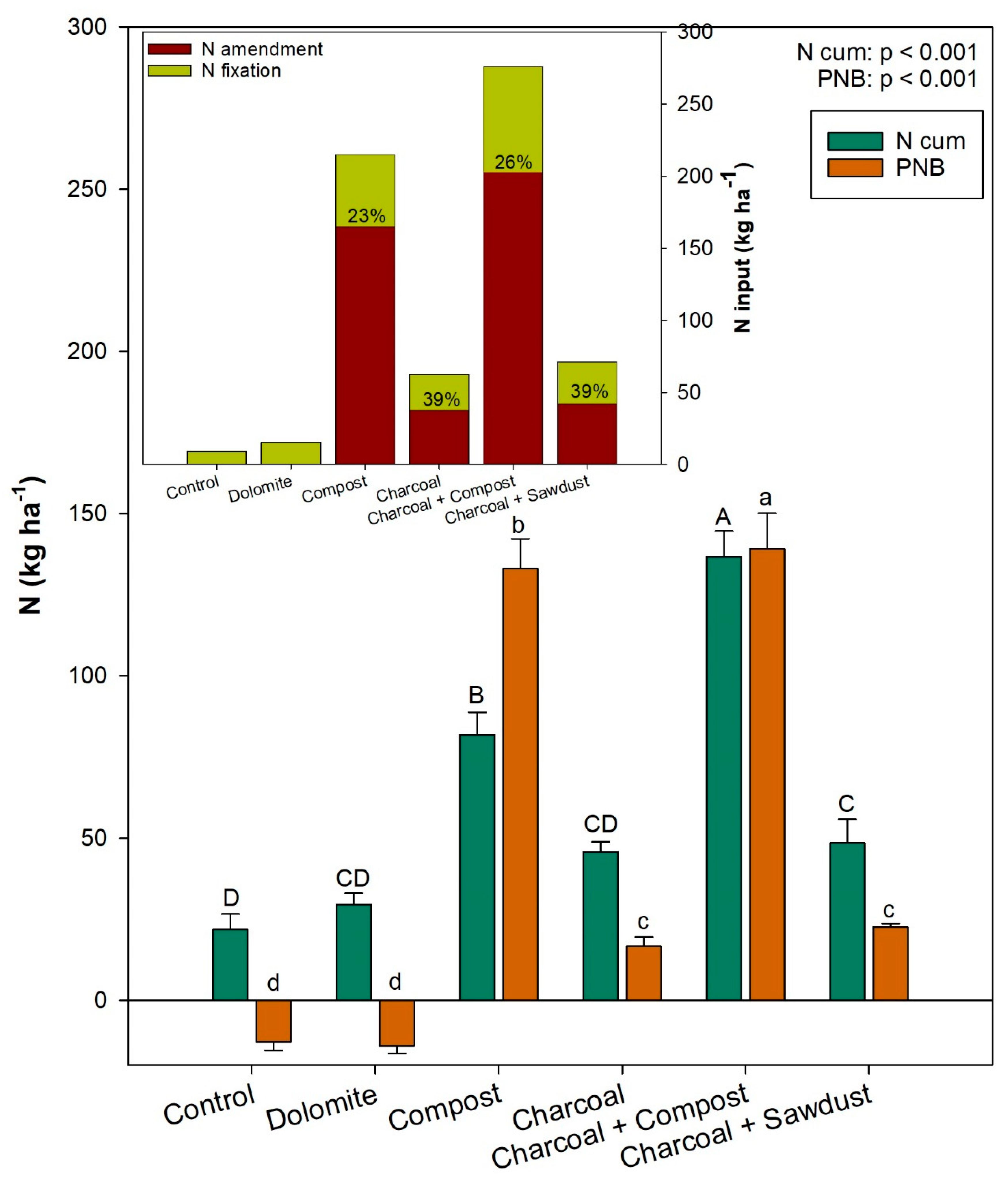
3.4. Total Nitrogen Uptake (TNuptake) in the System and Partial N Balance (PNB)
3.5. Soil N Dynamics
4. Discussion
4.1. Nitrogen Fixation by Centrosema
4.2. Crop Nitrogen Uptake
4.3. N Contribution to Soil
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anda, M.; Purwantari, N.D.; Yulistiani, D.; Sajimin; Suryani, E.; Husnain; Agus, F. Reclamation of post-tin mining areas using forages: A strategy based on soil mineralogy, chemical properties and particle size of the refused materials. CATENA 2022, 213, 106140. [Google Scholar] [CrossRef]
- Kumari, S.; Maiti, S.K. Reclamation of coalmine spoils with topsoil, grass, and legume: A case study from India. Environ. Earth Sci. 2019, 78, 429. [Google Scholar] [CrossRef]
- Maftukhah, R.; Kral, R.M.; Mentler, A.; Ngadisih, N.; Murtiningrum, M.; Keiblinger, K.M.; Gartner, M.; Hood-Nowotny, R. Post-Tin-Mining Agricultural Soil Regeneration Using Local Resources, Reduces Drought Stress and Increases Crop Production on Bangka Island, Indonesia. Agronomy 2023, 13, 50. [Google Scholar] [CrossRef]
- Kral, R.M.; Maftukhah, R.; Mentler, A.; Murtiningrum, M.; Ngadisih, N.; Keiblinger, K.M. Citizen science in the field: Co-experimentation at pilot scale for sustainable use of natural resources. Sustainability 2020, 12, 7700. [Google Scholar] [CrossRef]
- Kumari, S.; Ahirwal, J.; Maiti, S.K. Reclamation of industrial waste dump using grass-legume mixture: An experimental approach to combat land degradation. Ecol. Eng. 2022, 174, 106443. [Google Scholar] [CrossRef]
- Medoro, V.; Ferretti, G.; Galamini, G.; Rotondi, A.; Morrone, L.; Faccini, B.; Coltorti, M. Reducing Nitrogen Fertilization in Olive Growing by the Use of Natural Chabazite-Zeolitite as Soil Improver. Land 2022, 11, 1471. [Google Scholar] [CrossRef]
- Bossolani, J.W.; Crusciol, C.A.C.; Merloti, L.F.; Moretti, L.G.; Costa, N.R.; Tsai, S.M.; Kuramae, E.E. Long-term lime and gypsum amendment increase nitrogen fixation and decrease nitrification and denitrification gene abundances in the rhizosphere and soil in a tropical no-till intercropping system. Geoderma 2020, 375, 114476. [Google Scholar] [CrossRef]
- Reilly, E.C.; Gutknecht, J.L.; Tautges, N.E.; Sheaffer, C.C.; Jungers, J.M. Nitrogen transfer and yield effects of legumes intercropped with the perennial grain crop intermediate wheatgrass. Field Crops Res. 2022, 286, 108627. [Google Scholar] [CrossRef]
- Mugi-Ngenga, E.; Bastiaans, L.; Zingore, S.; Anten, N.P.R.; Giller, K.E. The role of nitrogen fixation and crop N dynamics on performance and legacy effects of maize-grain legumes intercrops on smallholder farms in Tanzania. Eur. J. Agron. 2022, 141, 126617. [Google Scholar] [CrossRef]
- Islami, T.; Guritno, B.; Basuki, N.; Suryanto, A. Biochar for sustaining productivity of cassava based cropping systems in the degraded lands of East Java, Indonesia. J. Trop. Agric. 2011, 49, 40–46. [Google Scholar]
- Budianta, D.; Gofar, N.; Andika, G.A. Improvement of Sand Tailing Fertility Derived from Post Tin Mining Using Leguminous Crop Applied by Compost and Mineral Soil. J. Tanah Trop. 2013, 18, 217–223. [Google Scholar]
- Jensen, E.S.; Carlsson, G.; Hauggaard-Nielsen, H. Intercropping of grain legumes and cereals improves the use of soil N resources and reduces the requirement for synthetic fertilizer N: A global-scale analysis. Agron. Sustain. Dev. 2020, 40, 5. [Google Scholar] [CrossRef]
- Ashworth, A.J.; West, C.P.; Allen, F.L.; Keyser, P.D.; Weiss, S.A.; Tyler, D.D.; Taylor, A.M.; Warwick, K.L.; Beamer, K.P. Biologically Fixed Nitrogen in Legume Intercropped Systems: Comparison of Nitrogen-Difference and Nitrogen-15 Enrichment Techniques. Agron. J. 2015, 107, 2419–2430. [Google Scholar] [CrossRef]
- De Notaris, C.; Mortensen, E.Ø.; Sørensen, P.; Olesen, J.E.; Rasmussen, J. Cover crop mixtures including legumes can self-regulate to optimize N2 fixation while reducing nitrate leaching. Agric. Ecosyst. Environ. 2021, 309, 107287. [Google Scholar] [CrossRef]
- Constantin, J.; Mary, B.; Laurent, F.; Aubrion, G.; Fontaine, A.; Kerveillant, P.; Beaudoin, N. Effects of catch crops, no till and reduced nitrogen fertilization on nitrogen leaching and balance in three long-term experiments. Agric. Ecosyst. Environ. 2010, 135, 268–278. [Google Scholar] [CrossRef]
- Hama, J.R.; Strobel, B.W. Natural alkaloids from narrow-leaf and yellow lupins transfer to soil and soil solution in agricultural fields. Environ. Sci. Eur. 2020, 32, 126. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C. Ameliorating soil acidity of tropical Oxisols by liming for sustainable crop production. Adv. Agron. 2008, 99, 345–399. [Google Scholar]
- Razafintsalama, H.; Trap, J.; Rabary, B.; Razakatiana, A.T.E.; Ramanankierana, H.; Rabeharisoa, L.; Becquer, T. Effect of Rhizobium Inoculation on Growth of Common Bean in Low-Fertility Tropical Soil Amended with Phosphorus and Lime. Sustainability 2022, 14, 4907. [Google Scholar] [CrossRef]
- Lusiba, S.G.; Maseko, S.T.; Odhiambo, J.J.O.; Adeleke, R. Biological N2 fixation, C accumulation and water-use efficiency (δ13C) of chickpea grown in three different soil types: Response to the addition of biochar from poultry litter and acacia. Acta Agric. Scand. Sect. B Soil Plant Sci. 2022, 72, 931–944. [Google Scholar] [CrossRef]
- Mia, S.; Van Groenigen, J.W.; Van de Voorde, T.F.J.; Oram, N.J.; Bezemer, T.M.; Mommer, L.; Jeffery, S. Biochar application rate affects biological nitrogen fixation in red clover conditional on potassium availability. Agric. Ecosyst. Environ. 2014, 191, 83–91. [Google Scholar] [CrossRef]
- Van Zwieten, L.; Rose, T.; Herridge, D.; Kimber, S.; Rust, J.; Cowie, A.; Morris, S. Enhanced biological N2 fixation and yield of faba bean (Vicia faba L.) in an acid soil following biochar addition: Dissection of causal mechanisms. Plant Soil 2015, 395, 7–20. [Google Scholar] [CrossRef]
- Rondon, M.A.; Lehmann, J.; Ramírez, J.; Hurtado, M. Biological nitrogen fixation by common beans (Phaseolus vulgaris L.) increases with bio-char additions. Biol. Fertil. Soils 2007, 43, 699–708. [Google Scholar] [CrossRef]
- Scheifele, M.; Hobi, A.; Buegger, F.; Gattinger, A.; Schulin, R.; Boller, T.; Mäder, P. Impact of pyrochar and hydrochar on soybean (Glycine max L.) root nodulation and biological nitrogen fixation. J. Plant Nutr. Soil Sci. 2017, 180, 199–211. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, C.; Wang, X.; Hai, L.; Hu, J. Increased abundance of nitrogen fixing bacteria by higher C/N ratio reduces the total losses of N and C in cattle manure and corn stover mix composting. Waste Manag. 2020, 103, 416–425. [Google Scholar] [CrossRef]
- Harindintwali, J.D.; Zhou, J.; Muhoza, B.; Wang, F.; Herzberger, A.; Yu, X. Integrated eco-strategies towards sustainable carbon and nitrogen cycling in agriculture. J. Environ. Manag. 2021, 293, 112856. [Google Scholar] [CrossRef]
- Shi, W.; Zhao, H.-Y.; Chen, Y.; Wang, J.-S.; Han, B.; Li, C.-P.; Lu, J.-Y.; Zhang, L.-M. Organic manure rather than phosphorus fertilization primarily determined asymbiotic nitrogen fixation rate and the stability of diazotrophic community in an upland red soil. Agric. Ecosyst. Environ. 2021, 319, 107535. [Google Scholar] [CrossRef]
- Agegnehu, G.; Bass, A.M.; Nelson, P.N.; Muirhead, B.; Wright, G.; Bird, M.I. Biochar and biochar-compost as soil amendments: Effects on peanut yield, soil properties and greenhouse gas emissions in tropical North Queensland, Australia. Agric. Ecosyst. Environ. 2015, 213, 72–85. [Google Scholar] [CrossRef]
- Haubensak, K.A.; D’Antonio, C.M.; Alexander, J. Effects of Nitrogen-Fixing Shrubs in Washington and Coastal California. Weed Technol. 2004, 18, 1475–1479. [Google Scholar] [CrossRef]
- Hood-Nowotny, R.; Watzinger, A.; Wawra, A.; Soja, G. The Impact of Biochar Incorporation on Inorganic Nitrogen Fertilizer Plant Uptake; An Opportunity for Carbon Sequestration in Temperate Agriculture. Geosciences 2018, 8, 420. [Google Scholar] [CrossRef]
- Hood-Nowotny, R.; Schwarzinger, B.; Schwarzinger, C.; Soliban, S.; Madakacherry, O.; Aigner, M.; Watzka, M.; Gilles, J. An Analysis of Diet Quality, How It Controls Fatty Acid Profiles, Isotope Signatures and Stoichiometry in the Malaria Mosquito Anopheles arabiensis. PLoS ONE 2012, 7, e45222. [Google Scholar] [CrossRef]
- Unkovich, M.J.; Pate, J.S.; Sanford, P.; Armstrong, E.L. Potential precision of the δN natural abundance method in field estimates of nitrogen fixation by crop and pasture legumes in south-west Australia. Aust. J. Agric. Res. 1994, 45, 119–132. [Google Scholar] [CrossRef]
- Yu, B.; Liu, G.; Liu, Q.; Huang, C.; Li, H.; Zhao, Z. Seasonal variation of deep soil moisture under different land uses on the semi-arid Loess Plateau of China. J. Soils Sediments 2019, 19, 1179–1189. [Google Scholar] [CrossRef]
- Hood-Nowotny, R.; Umana, N.H.-N.; Inselbacher, E.; Oswald-Lachouani, P.; Wanek, W. Alternative Methods for Measuring Inorganic, Organic, and Total Dissolved Nitrogen in Soil. Soil Sci. Soc. Am. J. 2010, 74, 1018. [Google Scholar] [CrossRef]
- Sørensen, P.; Jensen, E.S. Sequential diffusion of ammonium and nitrate from soil extracts to a polytetrafluoroethylene trap for 15N determination. Anal. Chim. Acta 1991, 252, 201–203. [Google Scholar] [CrossRef]
- Shearer, G.; Kohl, D.H. N2-Fixation in Field Settings: Estimations Based on Natural 15N Abundance. Funct. Plant Biol. 1986, 13, 699–756. [Google Scholar]
- Tsialtas, I.T.; Baxevanos, D.; Vlachostergios, D.N.; Dordas, C.; Lithourgidis, A. Cultivar complementarity for symbiotic nitrogen fixation and water use efficiency in pea-oat intercrops and its effect on forage yield and quality. Field Crops Res. 2018, 226, 28–37. [Google Scholar] [CrossRef]
- Rousk, K.; Sorensen, P.L.; Michelsen, A. Nitrogen Transfer from Four Nitrogen-Fixer Associations to Plants and Soils. Ecosystems 2016, 19, 1491–1504. [Google Scholar] [CrossRef]
- Peoples, M.B.; Brockwell, J.; Herridge, D.F.; Rochester, I.J.; Alves, B.J.R.; Urquiaga, S.; Boddey, R.M.; Dakora, F.D.; Bhattarai, S.; Maskey, S.L.; et al. The contributions of nitrogen-fixing crop legumes to the productivity of agricultural systems. Symbiosis 2009, 48, 1–17. [Google Scholar] [CrossRef]
- Fan, F.; Zhang, F.; Song, Y.; Sun, J.; Bao, X.; Guo, T.; Li, L. Nitrogen fixation of faba bean (Vicia faba L.) interacting with a non-legume in two contrasting intercropping systems. Plant Soil 2006, 283, 275–286. [Google Scholar] [CrossRef]
- Unkovich, M.; Herridge, D.; Peoples, M.; Cadisch, G.; Boddey, B.; Giller, K.; Alves, B.; Chalk, P. Measuring Plant-Associated Nitrogen Fixation in Agricultural Systems; Australian Centre for International Agricultural Research (ACIAR): Canberra, ACT, Australia, 2008. [Google Scholar]
- Landriscini, M.R.; Galantini, J.A.; Duval, M.E.; Capurro, J.E. Nitrogen balance in a plant-soil system under different cover crop-soybean cropping in Argentina. Appl. Soil Ecol. 2019, 133, 124–131. [Google Scholar] [CrossRef]
- Mia, S.; Dijkstra, F.A.; Singh, B. Enhanced biological nitrogen fixation and competitive advantage of legumes in mixed pastures diminish with biochar aging. Plant Soil 2018, 424, 639–651. [Google Scholar] [CrossRef]
- Romanyà, J.; Casals, P. Biological Nitrogen Fixation Response to Soil Fertility Is Species-Dependent in Annual Legumes. J. Soil Sci. Plant Nutr. 2020, 20, 546–556. [Google Scholar] [CrossRef]
- Guinet, M.; Nicolardot, B.; Revellin, C.; Durey, V.; Carlsson, G.; Voisin, A.-S. Comparative effect of inorganic N on plant growth and N 2 fixation of ten legume crops: Towards a better understanding of the differential response among species. Plant Soil 2018, 432, 207–227. [Google Scholar] [CrossRef]
- Kermah, M.; Franke, A.C.; Adjei-Nsiah, S.; Ahiabor, B.D.K.; Abaidoo, R.C.; Giller, K.E. N2-fixation and N contribution by grain legumes under different soil fertility status and cropping systems in the Guinea savanna of northern Ghana. Agric. Ecosyst. Environ. 2018, 261, 201–210. [Google Scholar] [CrossRef]
- Mathenge, C.; Thuita, M.; Masso, C.; Gweyi-Onyango, J.; Vanlauwe, B. Variability of soybean response to rhizobia inoculant, vermicompost, and a legume-specific fertilizer blend in Siaya County of Kenya. Soil Tillage Res. 2019, 194, 104290. [Google Scholar] [CrossRef]
- Xu, C.-Y.; Hosseini-Bai, S.; Hao, Y.; Rachaputi, R.C.N.; Wang, H.; Xu, Z.; Wallace, H. Effect of biochar amendment on yield and photosynthesis of peanut on two types of soils. Environ. Sci. Pollut. Res. 2015, 22, 6112–6125. [Google Scholar] [CrossRef]
- Quilliam, R.S.; DeLuca, T.H.; Jones, D.L. Biochar application reduces nodulation but increases nitrogenase activity in clover. Plant Soil 2013, 366, 83–92. [Google Scholar] [CrossRef]
- Haddad, S.A.; Mowrer, J.; Thapa, B. Biochar and compost from cotton residues inconsistently affect water use efficiency, nodulation, and growth of legumes under arid conditions. J. Environ. Manag. 2022, 307, 114558. [Google Scholar] [CrossRef]
- Lehmann, J.; Pereira da Silva, J.; Steiner, C.; Nehls, T.; Zech, W.; Glaser, B. Nutrient availability and leaching in an archaeological Anthrosol and a Ferralsol of the Central Amazon basin: Fertilizer, manure and charcoal amendments. Plant Soil 2003, 249, 343–357. [Google Scholar] [CrossRef]
- Lambers, J.H.R.; Harpole, W.S.; Tilman, D.; Knops, J.; Reich, P.B. Mechanisms responsible for the positive diversity–productivity relationship in Minnesota grasslands. Ecol. Lett. 2004, 7, 661–668. [Google Scholar] [CrossRef]
- Feng, L.; Yang, W.; Zhou, Q.; Tang, H.; Ma, Q.; Huang, G.; Wang, S. Effects of interspecific competition on crop yield and nitrogen utilisation in maize-soybean intercropping system. Plant Soil Environ. 2021, 67, 460–467. [Google Scholar] [CrossRef]
- Hale, S.E.; Nurida, N.L.; Mulder, J.; Sørmo, E.; Silvani, L.; Abiven, S.; Joseph, S.; Taherymoosavi, S.; Cornelissen, G. The effect of biochar, lime and ash on maize yield in a long-term fi eld trial in a Ultisol in the humid tropics. Sci. Total Environ. 2020, 719, 137455. [Google Scholar] [CrossRef]
- Jien, S.-H.; Chen, W.-C.; Ok, Y.S.; Awad, Y.M.; Liao, C.-S. Short-term biochar application induced variations in C and N mineralization in a compost-amended tropical soil. Environ. Sci. Pollut. Res. 2018, 25, 25715–25725. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef]
- Goergen, E.; Chambers, J.C.; Blank, R. Effects of water and nitrogen availability on nitrogen contribution by the legume, Lupinus argenteus Pursh. Appl. Soil Ecol. 2009, 42, 200–208. [Google Scholar] [CrossRef]
- De Notaris, C.; Rasmussen, J.; Sørensen, P.; Olesen, J.E. Nitrogen leaching: A crop rotation perspective on the effect of N surplus, field management and use of catch crops. Agric. Ecosyst. Environ. 2018, 255, 1–11. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Y.; Yan, X.; Li, J.; Jiao, N.; Hu, S. Biochar amendments increase the yield advantage of legume-based intercropping systems over monoculture. Agric. Ecosyst. Environ. 2017, 237, 16–23. [Google Scholar] [CrossRef]
- Walley, F.L.; Kyei-Boahen, S.; Hnatowich, G.; Stevenson, C. Nitrogen and phosphorus fertility management for desi and kabuli chickpea. Can. J. Plant Sci. 2005, 85, 73–79. [Google Scholar] [CrossRef]
- Xiu, L.; Zhang, W.; Wu, D.; Sun, Y.; Zhang, H.; Gu, W.; Wang, Y.; Meng, J.; Chen, W. Biochar can improve biological nitrogen fixation by altering the root growth strategy of soybean in Albic soil. Sci. Total Environ. 2021, 773, 144564. [Google Scholar] [CrossRef]
- Sánchez-Navarro, A.; Salas-Sanjuan, M.d.C.; Blanco-Bernardeau, M.A.; Sánchez-Romero, J.A.; Delgado-Iniesta, M.J. Medium-Term Effect of Organic Amendments on the Chemical Properties of a Soil Used for Vegetable Cultivation with Cereal and Legume Rotation in a Semiarid Climate. Land 2023, 12, 897. [Google Scholar] [CrossRef]
- Santhosh, K.S.; Akhila, D.S.; Dechamma, M.M.; Rajeshwari, V. An integrative approach to understand the role of the nitrogen fixing microbial consortia in the environment. J. Pharmacogn. Phytochem. 2019, 8, 909–915. [Google Scholar]
- Clocchiatti, A.; Hannula, S.E.; Hundscheid, M.P.J.; Klein Gunnewiek, P.J.A.; de Boer, W. Utilizing woody materials for fungal-based management of soil nitrogen pools. Appl. Soil Ecol. 2023, 181, 104663. [Google Scholar] [CrossRef]
- Huang, R.; Li, B.; Chen, Y.; Tao, Q.; Xu, Q.; Wen, D.; Gao, X.; Li, Q.; Tang, X.; Wang, C. Biochar Application Increases Labile Carbon and Inorganic Nitrogen Supply in a Continuous Monocropping Soil. Land 2022, 11, 473. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | N Applied (kg ha−1) |
|---|---|
| Control | 0 |
| Dolomite | 0 |
| Compost | 165 |
| Charcoal | 38 |
| Charcoal and compost | 203 |
| Charcoal and sawdust | 42 |
| Treatments | TNuptake (kg ha−1) | TN2–fixation (kg ha−1) | TNDFS (kg ha−1) |
|---|---|---|---|
| Control | 12 ± 3c | 9 ± 3c | 3 ± 0.8 |
| Dolomite | 17 ± 1c | 16 ± 2bc | 2 ± 0.6 |
| Compost | 61 ± 5b | 50 ± 3ab | 11 ± 6 |
| Charcoal | 29 ± 3c | 25 ± 3b | 5 ± 1 |
| Charcoal + compost | 111 ± 7a | 73 ± 10a | 38 ± 12 |
| Charcoal + sawdust | 34 ± 6bc | 29 ± 7b | 5 ± 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maftukhah, R.; Keiblinger, K.M.; Ngadisih, N.; Murtiningrum, M.; Kral, R.M.; Mentler, A.; Hood-Nowotny, R. Post-Tin-Mining Agricultural Soil Regeneration Using Local Organic Amendments Improve Nitrogen Fixation and Uptake in a Legume–Cassava Intercropping System. Land 2023, 12, 1107. https://doi.org/10.3390/land12051107
Maftukhah R, Keiblinger KM, Ngadisih N, Murtiningrum M, Kral RM, Mentler A, Hood-Nowotny R. Post-Tin-Mining Agricultural Soil Regeneration Using Local Organic Amendments Improve Nitrogen Fixation and Uptake in a Legume–Cassava Intercropping System. Land. 2023; 12(5):1107. https://doi.org/10.3390/land12051107
Chicago/Turabian StyleMaftukhah, Rizki, Katharina M. Keiblinger, Ngadisih Ngadisih, Murtiningrum Murtiningrum, Rosana M. Kral, Axel Mentler, and Rebecca Hood-Nowotny. 2023. "Post-Tin-Mining Agricultural Soil Regeneration Using Local Organic Amendments Improve Nitrogen Fixation and Uptake in a Legume–Cassava Intercropping System" Land 12, no. 5: 1107. https://doi.org/10.3390/land12051107
APA StyleMaftukhah, R., Keiblinger, K. M., Ngadisih, N., Murtiningrum, M., Kral, R. M., Mentler, A., & Hood-Nowotny, R. (2023). Post-Tin-Mining Agricultural Soil Regeneration Using Local Organic Amendments Improve Nitrogen Fixation and Uptake in a Legume–Cassava Intercropping System. Land, 12(5), 1107. https://doi.org/10.3390/land12051107