
Ecological Connectivity of Vicuña (Vicugna vicugna) in a Remote Area of Chile and Conservation Implications

 , , , ,
, , , ,
Abstract
:1. Introduction
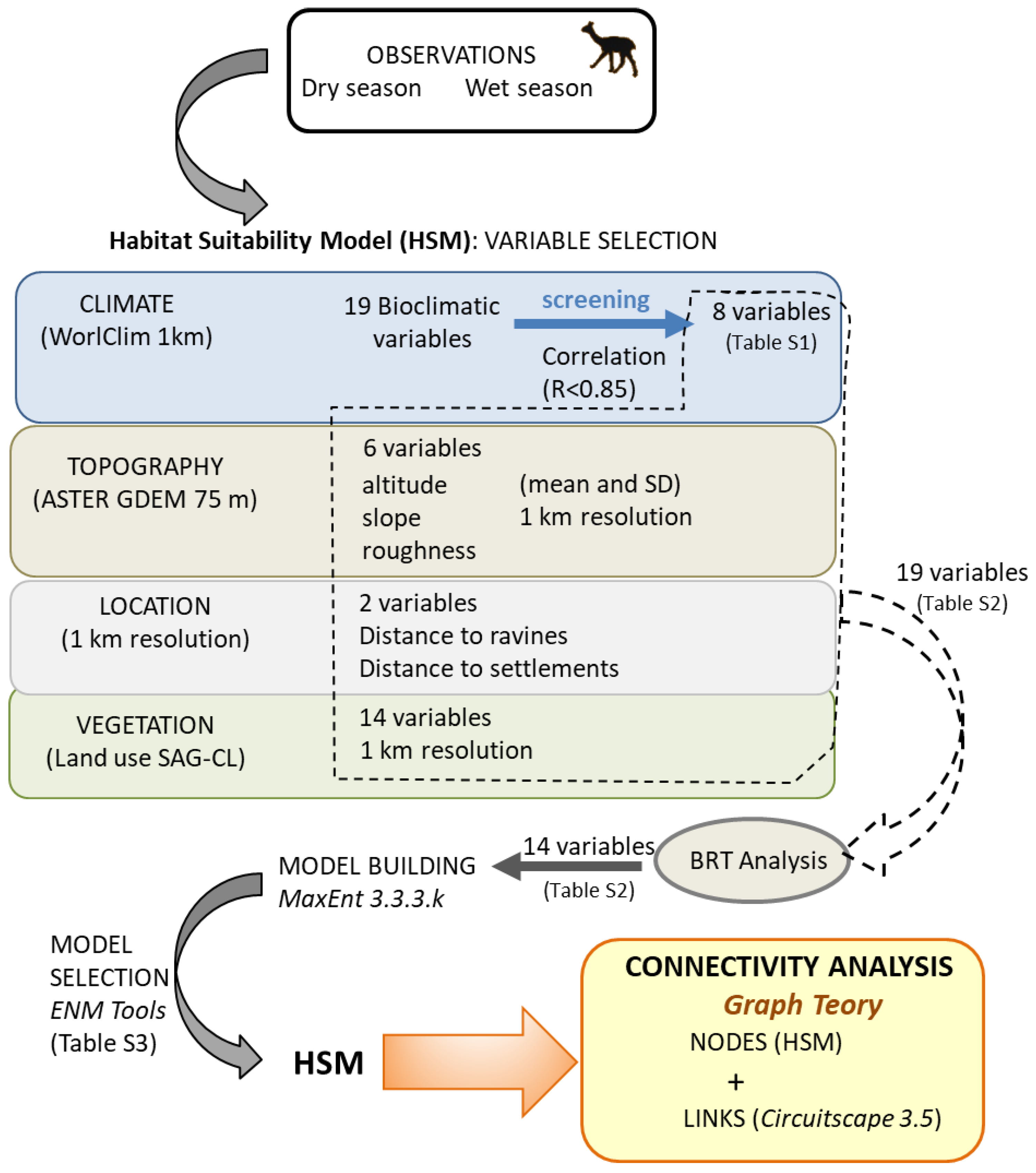
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. Connectivity Analysis
3. Results
3.1. Habitat Suitability Models
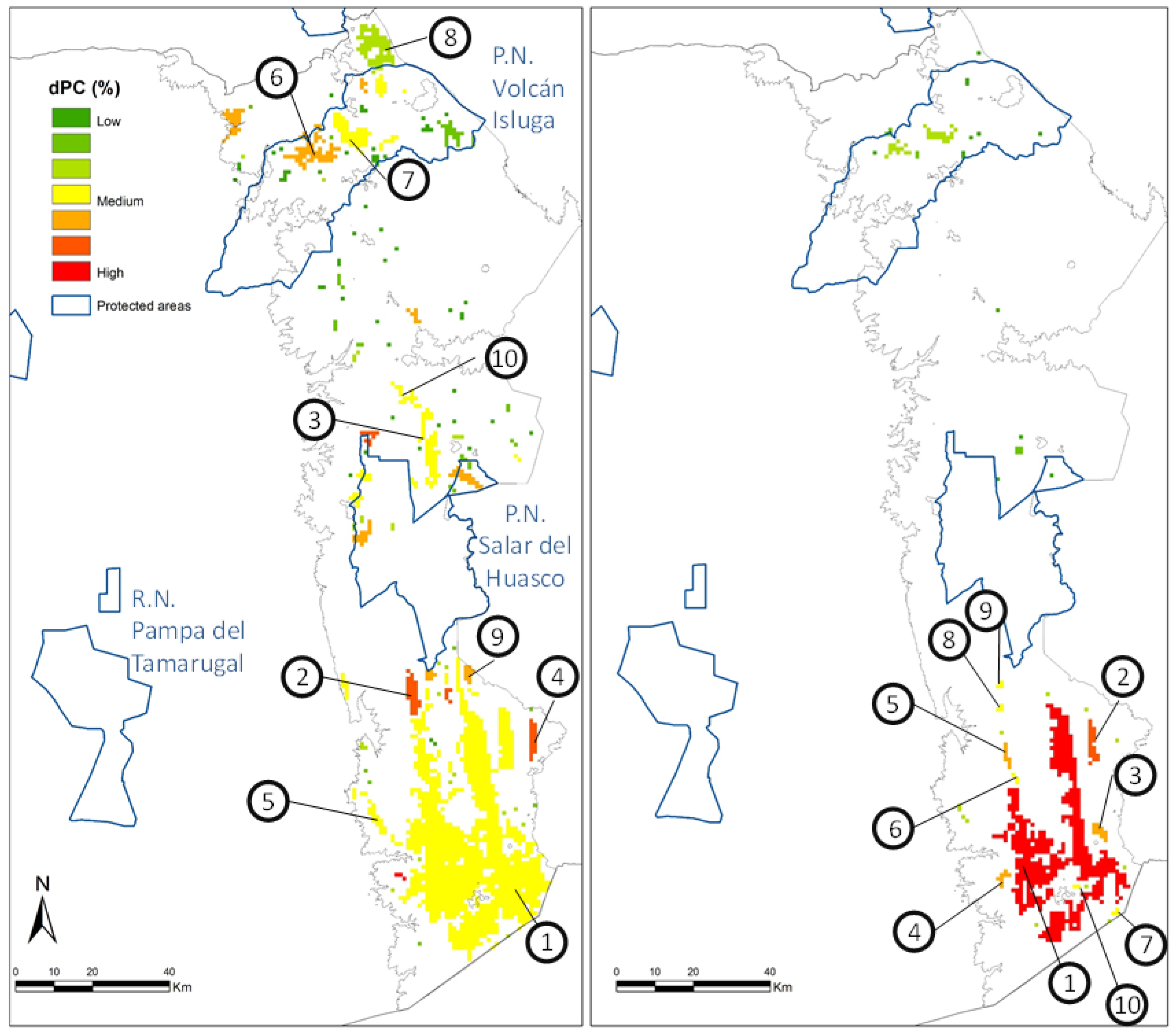
3.2. Connectivity Analysis

{kind=link}
{kind=link}
{kind=link}
| Threshold | ECAnorm | dPCintra | dPCflux | dPCconnector |
|---|---|---|---|---|
| MaxSS | 7.70 | 55.99 | 43.44 | 0.57 |
| AvPP | 8.98 | 66.5 | 33.32 | 0.18 |

| Threshold | Node | dPC | dPCintra | dPCflux | dPCconnector | Area (Ha) |
|---|---|---|---|---|---|---|
| MaxSS | 1 | 93.833 | 71.140 | 22.511 | 0.182 | 1146 |
| 2 | 4.019 | 0.046 | 3.943 | 0.031 | 29 | |
| 3 | 3.113 | 0.135 | 2.698 | 0.280 | 50 | |
| 4 | 2.331 | 0.020 | 2.312 | 0.000 | 19 | |
| 5 | 2.049 | 0.016 | 2.007 | 0.026 | 17 | |
| 6 | 1.627 | 0.176 | 1.431 | 0.019 | 57 | |
| 7 | 1.410 | 0.125 | 1.279 | 0.006 | 48 | |
| 8 | 1.405 | 0.236 | 1.169 | 0.000 | 66 | |
| 9 | 1.351 | 0.005 | 1.346 | 0.000 | 10 | |
| 10 | 1.034 | 0.014 | 0.845 | 0.174 | 16 | |
| AvPP | 1 | 98.589 | 79.569 | 18.811 | 0.208 | 519 |
| 2 | 6.165 | 0.130 | 6.026 | 0.009 | 21 | |
| 3 | 3.627 | 0.043 | 3.585 | 0.000 | 12 | |
| 4 | 2.464 | 0.019 | 2.445 | 0.000 | 8 | |
| 5 | 2.239 | 0.019 | 2.220 | 0.000 | 8 | |
| 6 | 0.878 | 0.003 | 0.876 | 0.000 | 3 | |
| 7 | 0.874 | 0.003 | 0.871 | 0.000 | 3 | |
| 8 | 0.698 | 0.003 | 0.696 | 0.000 | 3 | |
| 9 | 0.650 | 0.003 | 0.647 | 0.000 | 3 | |
| 10 | 0.608 | 0.001 | 0.607 | 0.000 | 2 |
4. Discussion
Conservation Implications
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CMS (Convention on the Conservation of Migratory Species). Improving Ways of Addressing Connectivity in the Conservation of Migratory Species, Resolution 12.26 (REV.COP13); Convention on Migratory Species: Gandhinagar, India, 2020; Available online: https://www.cms.int/sites/default/files/document/cms_cop13_doc.26.4.4_addressing-connectivity-in-conservation-of-migratory-species_e.pdf (accessed on 29 November 2023).
- Taylor, P.D.; Fahrig, L.; With, K.A. Landscape connectivity: A return to the basics. In Connectivity Conservation; Crooks, K.R., Sanjayan, M., Eds.; Cambridge University Press: Cambridge, UK, 2006; pp. 29–43. [Google Scholar]
- Crooks, K.R.; Sanjayan, M. Connectivity Conservation; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Baguette, M.; Blanchet, S.; Legrand, D.; Stevens, V.M.; Turlure, C. Individual dispersal, landscape connectivity and ecological networks. Biol. Rev. 2013, 88, 310–326. [Google Scholar] [CrossRef] [PubMed]
- Morelli, T.L.; Maher, S.P.; Lim, M.C.W.; Kastely, C.; Eastman, L.M.; Flint, L.E.; Flint, A.L.; Beissinger, S.R.; Moritz, C. Climate change refugia and habitat connectivity promote species persistence. Clim. Chang. Responses 2017, 4, 8. [Google Scholar] [CrossRef]
- Costanza, J.K.; Terando, A.J. Landscape Connectivity Planning for Adaptation to Future Climate and Land-Use Change. Curr. Landsc. Ecol. Rep. 2019, 4, 1–13. [Google Scholar] [CrossRef]
- Carroll, C.; Rohlf, D.J.; Li, Y.W.; Hartl, B.; Phillips, M.K.; Noss, R.F. Connectivity Conservation and Endangered Species Recovery: A Study in the Challenges of Defining Conservation-Reliant Species. Conserv. Lett. 2015, 8, 132–138. [Google Scholar] [CrossRef]
- Kauffman, M.J.; Cagnacci, F.; Chamaillé-Jammes, S.; Hebblewhite, M.; Hopcraft, J.G.C.; Merkle, J.A.; Mueller, T.; Mysterud, A.; Peters, W.; Roettger, C.; et al. Mapping out a future for ungulate migrations. Science 2021, 372, 566–569. [Google Scholar] [CrossRef]
- Rabinowitz, A.; Zeller, K. A rangewide model of landscape connectivity and conservation for the jaguar, Panthera onca. Biol. Conserv. 2010, 143, 939–945. [Google Scholar] [CrossRef]
- Ziółkowska, E.; Ostapowicz, K.; Kuemmerle, T.; Perzanowski, K.; Radeloff, V.C.; Kozak, J. Potential habitat connectivity of European bison (Bison bonasus) in the Carpathians. Biol. Conserv. 2012, 146, 188–196. [Google Scholar] [CrossRef]
- Kabir, M.; Hameed, S.; Ali, H.; Bosso, L.; Din, J.U.; Bischof, R.; Redpath, S.; Nawaz, M.A. Habitat suitability and movement corridors of grey wolf (Canis lupus) in Northern Pakistan. PLoS ONE 2017, 12, e0187027. [Google Scholar] [CrossRef]
- Batter, T.J.; Bush, J.P.; Sacks, B.N. Assessing genetic diversity and connectivity in a tule elk (Cervus canadensis nannodes) metapopulation in Northern California. Conserv. Genet. 2021, 22, 889–901. [Google Scholar] [CrossRef]
- González, B.A.; Donoso, D.S. Capítulo 4. Distribución geográfica actual de la vicuña austral. In La Vicuña Austral; González, B.A., Ed.; Facultad de Ciencias Forestales y de la Conservación de la Naturaleza, Corporación Nacional Forestal y Grupo Especialista en Camélidos Sudamericanos Silvestres: Santiago, Chile, 2020; ISBN 978-956-7669-74-5. [Google Scholar]
- Yacobaccio, H. The historical relationship between people and the vicuña. In The Vicuña; Gordon, I.J., Ed.; Springer: Boston, MA, USA, 2009; pp. 7–20. [Google Scholar] [CrossRef]
- Wheeler, J.C.; Laker, J. The vicuña in the Andean altiplano. In The Vicuña: The Theory and Practice of Community-Based Wildlife Management; Gordon, I., Ed.; Springer: Boston, MA, USA, 2009; pp. 21–33. [Google Scholar] [CrossRef]
- Jungius, H. Bolivia and the vicuna. Oryx 1972, 11, 335–346. [Google Scholar] [CrossRef]
- Boswall, J. Vicuna in Argentina. Oryx 1972, 11, 449–456. [Google Scholar] [CrossRef]
- Grimwood, I. Notes on the Distribution and Status of Some Peruvian Mammals; Zoological Society: New York, NY, USA, 1969; Volume 21. [Google Scholar]
- Ceballos, G.; Ehrlich, P.R. Mammal Population Losses and the Extinction Crisis. Science 2002, 296, 904–907. [Google Scholar] [CrossRef] [PubMed]
- Acebes, P.; Wheeler, J.; Baldo, J.; Tuppia, P.; Lichtenstein, G.; Hoces, D.; Franklin, W.L. Vicugna vicugna: The IUCN Red List of Threatened Species; e.T22956A18540534; International Union for Conservation of Nature: Gland, Switzerland, 2018. [Google Scholar]
- Arzamendia, Y.; Carbajo, A.E.; Vilá, B. Social group dynamics and composition of managed wild vicuñas (Vicugna vicugna vicugna) in Jujuy, Argentina. J. Ethol. 2018, 36, 125–134. [Google Scholar] [CrossRef]
- González, B.A.; Donoso, D.S.; Villalobos, R.; Lagos, N.; Iriarte, A. Box 6.1. Ámbito de hogar y preferencia de hábitat de individuos de vicuña austral en el altiplano de Chile. In La Vicuña Austral; González, B.A., Ed.; Facultad de Ciencias Forestales y de la Conservación de la Naturaleza, Corporación Nacional Forestal y Grupo Especialista en Camélidos Sudamericanos: Santiago, Chile, 2020; ISBN 978-956-7669-74-5. [Google Scholar]
- Karandikar, H.; Donadio, E.; Smith, J.A.; Bidder, O.R.; Middleton, A.D. Spatial ecology of the Vicuña (Lama vicugna) in a high Andean protected area. J. Mammal. 2023, 104, 509–518. [Google Scholar] [CrossRef]
- Malo, J.E.; González, B.A.; Mata, C.; Vielma, A.; Donoso, D.S.; Fuentes, N.; Estades, C.F. Low habitat overlap at landscape scale between wild camelids and feral donkeys in the Chilean desert. Acta Oecol. 2016, 70, 1–9. [Google Scholar] [CrossRef]
- González, B.A.; Marín, J.C.; Toledo, V.; Espinoza, E. Wildlife forensic science in the investigation of poaching of vicuña. Oryx 2016, 50, 14–15. [Google Scholar] [CrossRef]
- Mata, C.; Malo, J.E.; Galaz, J.L.; Cadorzo, C.; Lagunas, H. A three-step approach to minimise the impact of a mining site on vicuña (Vicugna vicugna) and to restore landscape connectivity. Environ. Sci. Pollut. Res. 2016, 23, 13626–13636. [Google Scholar] [CrossRef]
- Franklin, W.L. Contrasting socioecologies of South American’s wild camelids: The vicuña and guanaco. In Advances in the Study of Mammalian Behavior; Eisenberg, J.F., Kleiman, D.G., Eds.; American Society of Mammalogists Special Publication: Topeka, KS, USA, 1983; Volume 7, pp. 573–629. [Google Scholar]
- Tirado, C.; Cortés, A.; Miranda-Urbina, E.; Carretero, M.A. Trophic preferences in an assemblage of mammal herbivores from Andean Puna (Northern Chile). J. Arid Environ. 2012, 79, 8–12. [Google Scholar] [CrossRef]
- Arzamendia, Y.; Cassini, M.H.; Vilá, B. Habitat use by vicuña Vicugna vicugna in Laguna Pozuelos Reserve, Jujuy, Argentina. Oryx 2006, 40, 198–203. [Google Scholar] [CrossRef]
- Moraga, C.; Donoso, D.S.; González, B.A. Capítulo 5. Metodologías de monitoreo en vicuñas y estimaciones de abundancia y densidad de vicuña austral en Chile. In La Vicuña Austral; González, B.A., Ed.; Facultad de Ciencias Forestales y de la Conservación de la Naturaleza, Corporación Nacional Forestal y Grupo Especialista en Camélidos Sudamericanos: Santiago, Chile, 2020. [Google Scholar]
- Miller, S.D.; Rottmann, J.; Raedeke, K.J.; Taber, R.D. Endangered mammals of Chile: Status and conservation. Biol. Conserv. 1983, 25, 335–352. [Google Scholar] [CrossRef]
- Luebert, F.; Pliscoff, P. Sinopsis Bioclimática y Vegetacional de Chile; Editorial Universitaria: Santiago de Chile, Chile, 2018. [Google Scholar]
- IUCN (International Union for Conservation of Nature). Vicugna vicugna: The IUCN Red List of Threatened Species; Version 2022-2; International Union for Conservation of Nature: Gland, Switzerland, 2022. [Google Scholar]
- Hughes, A.C.; Orr, M.C.; Ma, K.; Costello, M.J.; Waller, J.; Provoost, P.; Yang, Q.; Zhu, C.; Huijie Qiao, H. Sampling biases shape our view of the natural world. Ecography 2021, 44, 1259–1269. [Google Scholar] [CrossRef]
- Bunn, A.G.; Urban, D.L.; Keitt, T.H. Landscape connectivity: A conservation application of graph theory. J. Environ. Manag. 2000, 59, 265–278. [Google Scholar] [CrossRef]
- Pascual-Hortal, L.; Saura, S. Comparison and development of new graph-based landscape connectivity indices: Towards the priorization of habitat patches and corridors for conservation. Landsc. Ecol. 2006, 21, 959–967. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Lobo, J.M. Threshold criteria for conversion of probability of species presence to either- or presence-absence. Acta Oecol. 2007, 31, 361–369. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G.; Pearson, R. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- McRae, B.H.; Dickson, B.G.; Keitt, T.H.; Shah, V.B. Using circuit theory to model connectivity in ecology, evolution, and conservation. Ecology 2008, 89, 2712–2724. [Google Scholar] [CrossRef]
- Saura, S.; Estreguil, C.; Mouton, C.; Rodríguez-Freire, M. Network analysis to assess landscape connectivity trends: Application to European forests (1990–2000). Ecol. Indic. 2011, 11, 407–416. [Google Scholar] [CrossRef]
- Sutherland, G.D.; Harestad, A.S.; Price, K.; Lertzman, K.P. Scaling of natal dispersal distances in terrestrial birds and mammals. Conserv. Ecol. 2000, 4, 16. [Google Scholar] [CrossRef]
- Bowman, J.; Jaeger, J.A.; Fahrig, L. Dispersal distance of mammals is proportional to home range size. Ecology 2002, 83, 2049–2055. [Google Scholar] [CrossRef]
- Bonacic, C. Sustainable Use of the Vicuña in Chile. Master’s Thesis, University of Reading, Reading, UK, 1996. [Google Scholar]
- Saura, S.; Rubio, L. A common currency for the different ways in which patches and links can contribute to habitat availability and connectivity in the landscape. Ecography 2010, 33, 523–537. [Google Scholar] [CrossRef]
- Saura, S.; Torné, J. Conefor Sensinode 2.2: A software package for quantify the importance of habitat patches for landscape connectivity. Environ. Model. Softw. 2009, 24, 135–139. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef] [PubMed]
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Beger, M.; Metaxas, A.; Balbar, A.C.; McGowan, J.A.; Daigle, R.; Kuempel, C.D.; Treml, E.A.; Possingham, H.P. Demystifying ecological connectivity for actionable spatial conservation planning. Trends Ecol. Evol. 2022, 37, 1079–1091. [Google Scholar] [CrossRef] [PubMed]
- González, B.A.; Vásquez, J.P.; Gómez-Uchida, D.; Cortés, J.; Rivera, R.; Aravena, N.; Chero, A.M.; Agapito, A.M.; Varas, V.; Wheleer, J.C.; et al. Phylogeography and Population Genetics of Vicugna vicugna: Evolution in the Arid Andean High Plateau. Front. Genet. 2019, 10, 445. [Google Scholar] [CrossRef] [PubMed]
- Unnithan Kumar, S.; Turnbull, J.; Hartman Davies, O.; Hodgetts, T.; Cushman, S.A. Moving beyond landscape resistance: Considerations for the future of connectivity modelling and conservation science. Lands Ecol. 2022, 37, 2465–2480. [Google Scholar] [CrossRef]
- d’Arc, N.R.; Cassini, M.H.; Vilá, B.L. Habitat use by vicuñas Vicugna vicugna in the Laguna Blanca Reserve (Catamarca, Argentina). J. Arid Environ. 2000, 46, 107–115. [Google Scholar]
- Fuentes-Allende, N.; Quilodrán, C.S.; Jofré, A.; González, B.A. Behavioral responses of vicuñas to human activities at priority feeding sites associated with roads in the highland desert of northern Chile. Int. J. Agric. Nat. Resour. 2023, 50, 75–83. [Google Scholar] [CrossRef]
- Mosca-Torres, M.E.; Puig, S. Seasonal diet of vicuñas in the Los Andes protected area (Salta, Argentina): Are they optimal foragers? J. Arid Environ. 2010, 74, 450–457. [Google Scholar] [CrossRef]
- MMA (Ministerio del Medio Ambiente). Plan Nacional de Protección de Humedales 2018–2022; Ministerio del Medio Ambiente, Gobierno de Chile: Santiago, Chile, 2018.
- IUCN. Red List Categories and Criteria, Version 3.1, 2nd ed.; IUCN Library System: Gland, Switzerland; Cambridge, UK, 2012; 32p, ISBN 978-2-8317-1435-6. [Google Scholar]
- RCE (Reunión Comité Clasificación de Especies Silvestres). Ministerio del Medio Ambiente, Gobierno de Chile. 2019. Available online: https://clasificacionespecies.mma.gob.cl/wp-content/uploads/2019/12/Acta_RCE_4_Decimosexto_Proc_9_octubre_2019.pdf (accessed on 26 October 2023).
- Cushman, S.A.; Elliot, N.B.; Macdonald, D.W.; Loveridge, A.J. A multi-scale assessment of population connectivity in African lions (Panthera leo) in response to landscape change. Landsc. Ecol. 2016, 31, 1337–1353. [Google Scholar] [CrossRef]
- Quilodrán, C.S.; Montoya-Burgos, J.I.; Currat, M. Harmonizing hybridization dissonance in conservation. Commun. Biol. 2020, 3, 391. [Google Scholar] [CrossRef]
- SNASPE (Sistema Nacional de Áreas Silvestres del Estado). Biblioteca del Congreso Nacional de Chile. Información Territorial. Available online: https://www.bcn.cl/siit/mapas_vectoriales/index_html (accessed on 22 December 2022).
- Sonter, L.J.; Ali, S.H.; Watson, J.E.M. Mining and biodiversity: Key issues and research needs in conservation science. Proc. R. Soc. B 2018, 285, 20181926. [Google Scholar] [CrossRef] [PubMed]
- Krosby, M.; Tewksbury, J.; Haddad, N.M.; Hoekstra, J. Ecological connectivity for a changing climate. Conserv. Biol. 2010, 24, 1686–1689. [Google Scholar] [CrossRef] [PubMed]
- Oxford Analytica. Climate change will hit the Andean region unevenly. Expert Briefs 2022. [Google Scholar] [CrossRef]
- Sarricolea Espinoza, P.; Romero Aravena, H. Variabilidad y cambios climáticos observados y esperados en el Altiplano del norte de Chile. Rev. Geogr. Norte Gd. 2015, 62, 169–183. (In Spanish) [Google Scholar] [CrossRef]
- Chavez, R.O.; Meseguer-Ruiz, O.; Olea, M.; Calderon-Seguel, M.; Yager, K.; Meneses, R.I.; Lastra, J.A.; Nuñez-Hidalgo, I.; Sarricolea, P.; Serrano-Notivoli, R.; et al. Andean peatlands at risk? Spatiotemporal patterns of extreme NDVI anomalies, water extraction and drought severity in a large-scale mining area of Atacama, northern Chile. Int. J. Appl. Earth Obs. Geoinf. 2023, 116, 103138. [Google Scholar] [CrossRef]
- SERNAGEOMIN (Servicio Nacional de Geología y Minería). Anuario de la Minería de Chile 2022; Servicio Nacional de Geología y Minería: Santiago, Chile, 2023; 235p.
- Hamann, R. Mining companies’ role in sustainable development: The ‘why’ and ‘how’ of corporate social responsibility from a business perspective. Dev. South. Afr. 2003, 20, 237–254. [Google Scholar] [CrossRef]
- Hartman, B.D.; Bookhagen, B.; Chadwick, O.A. The effects of check dams and other erosion control structures on the restoration of Andean bofedal. Restor. Ecol. 2016, 24, 761–772. [Google Scholar] [CrossRef]


| Variables | Contribution (%) | Jackknife Test of Training Gain | ||
|---|---|---|---|---|
| Only the Variable | Without the Variable | |||
| Topographic | Mean altitude | 0.57 | 0.014 | 1.048 |
| SD altitude | 10.34 | 0.168 | 0.938 | |
| Mean gradient | 0.62 | 0.010 | 1.030 | |
| Location | Distance to ravines | 11.14 | 0.222 | 0.891 |
| Distance to settlements | 4.90 | 0.025 | 1.020 | |
| Climatic | BIO 1 | 5.10 | 0.013 | 0.974 |
| BIO2 | 0.20 | 0.062 | 1.051 | |
| BIO 3 | 1.02 | 0.097 | 1.038 | |
| BIO 4 | 5.10 | 0.227 | 1.047 | |
| BIO 7 | 3.02 | 0.064 | 1.052 | |
| BIO 12 | 40.34 | 0.463 | 0.926 | |
| Vegetation | Bofedal | 1.16 | 0.002 | 1.039 |
| Steppe | 9.76 | 0.026 | 0.891 | |
| Very open shrub | 6.72 | 0.129 | 1.010 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mata, C.; González, B.A.; Donoso, D.S.; Fuentes-Allende, N.; Estades, C.F.; Malo, J.E. Ecological Connectivity of Vicuña (Vicugna vicugna) in a Remote Area of Chile and Conservation Implications. Land 2024, 13, 472. https://doi.org/10.3390/land13040472
Mata C, González BA, Donoso DS, Fuentes-Allende N, Estades CF, Malo JE. Ecological Connectivity of Vicuña (Vicugna vicugna) in a Remote Area of Chile and Conservation Implications. Land. 2024; 13(4):472. https://doi.org/10.3390/land13040472
Chicago/Turabian StyleMata, Cristina, Benito A. González, Denise S. Donoso, Nicolás Fuentes-Allende, Cristián F. Estades, and Juan E. Malo. 2024. "Ecological Connectivity of Vicuña (Vicugna vicugna) in a Remote Area of Chile and Conservation Implications" Land 13, no. 4: 472. https://doi.org/10.3390/land13040472
APA StyleMata, C., González, B. A., Donoso, D. S., Fuentes-Allende, N., Estades, C. F., & Malo, J. E. (2024). Ecological Connectivity of Vicuña (Vicugna vicugna) in a Remote Area of Chile and Conservation Implications. Land, 13(4), 472. https://doi.org/10.3390/land13040472