
Linear and Nonlinear Quantitative EEG Analysis during Neutral Hypnosis following an Opened/Closed Eye Paradigm

 ,
,  , ,
, ,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Protocol
2.3. Data Acquisition and Preprocessing
2.4. Power Analysis
2.5. Lempel-Ziv Complexity Analysis
2.6. Tsallis Entropy
2.7. Statistical Analysis
3. Results
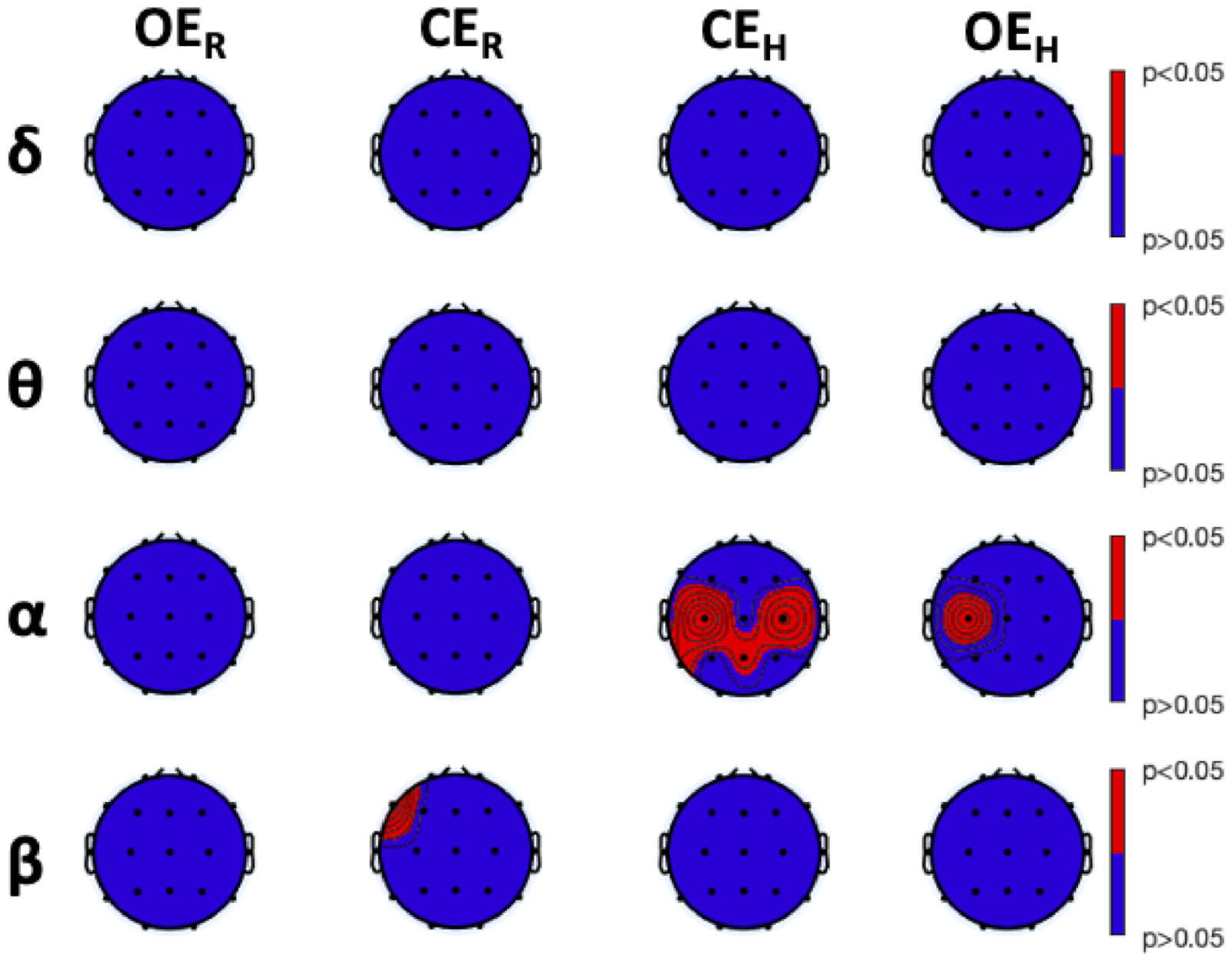
3.1. Power Analysis
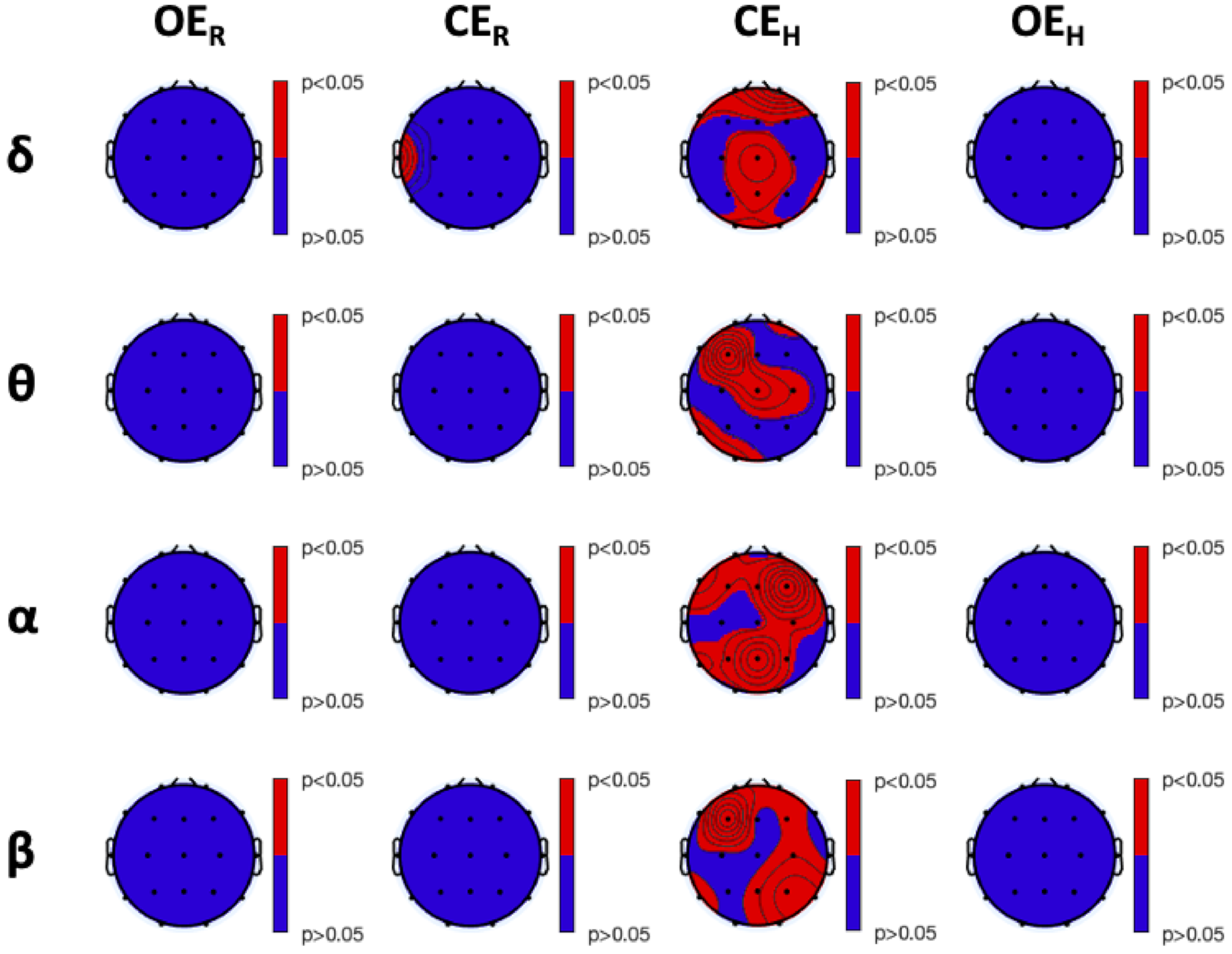
3.2. Lempel-Ziv Complexity Analysis
3.3. Tsallis Entropy Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goodman, M.S.; Kumar, S.; Zomorrodi, R.; Ghazala, Z.; Cheam, A.S.; Barr, M.S.; Daskalakis, Z.J.; Blumberger, D.M.; Fischer, C.; Flint, A.; et al. Theta-gamma coupling and working memory in Alzheimer’s dementia and mild cognitive impairment. Front. Aging Neurosci. 2018, 10, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oakley, D.A.; Halligan, P.W. Hypnotic suggestion: Opportunities for cognitive neuroscience. Nat. Rev. Neurosci. 2013, 14, 565–576. [Google Scholar] [CrossRef]
- Landry, M.; Lifshitz, M.; Raz, A. Brain correlates of hypnosis: A systematic review and meta-analytic exploration. Neurosci. Biobehav. Rev. 2017, 81, 75–98. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.P.; Adachi, T.; Tomé-Pires, C.; Lee, J.; Osman, Z.J.; Miró, J. Mechanisms of Hypnosis: Toward the Development of a Biopsychosocial Model. Int. J. Clin. Exp. Hypn. 2015, 63, 34–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynn, S.J.; Kirsch, I. Essentials of Clinical Hypnosis: An Evidence-Based Approach; American Psychological Association: Washington, DC, USA, 2006. [Google Scholar]
- Lipari, S.; Baglio, F.; Griffanti, L.; Mendozzi, L.; Garegnani, M.; Motta, A.; Cecconi, P.; Pugnetti, L. Altered and asymmetric default mode network activity in a “hypnotic virtuoso”: An fMRI and EEG study. Conscious. Cogn. 2012, 21, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Cardeña, E.; Lehmann, D.; Faber, P.L.; Jönsson, P.; Milz, P.; Pascual-Marqui, R.D.; Kochi, K. EEG sLORETA functional imaging during hypnotic arm levitation and voluntary arm lifting. Int. J. Clin. Exp. Hypn. 2012, 60, 31–53. [Google Scholar] [CrossRef]
- Gandhi, B.; Oakley, D.A. Does ‘hypnosis’ by any other name smell as sweet? The efficacy of ‘hypnotic’inductions depends on the label ‘hypnosis’. Conscious. Cogn. 2005, 14, 304–315. [Google Scholar] [CrossRef]
- Cardeña, E.; Jönsson, P.; Terhune, D.B.; Marcusson-Clavertz, D. The neurophenomenology of neutral hypnosis. Cortex 2013, 49, 375–385. [Google Scholar] [CrossRef]
- Kihlstrom, J.F. Neuro-hypnotism: Prospects for hypnosis and neuroscience. Cortex 2013, 49, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Mazzoni, G.; Venneri, A.; McGeown, W.J.; Kirsch, I. Neuroimaging resolution of the altered state hypothesis. Cortex 2013, 49, 400–410. [Google Scholar] [CrossRef]
- Sabourin, M.E.; Cutcomb, S.D.; Crawford, H.J.; Pribram, K. EEG correlates of hypnotic susceptibility and hypnotic trance: Spectral analysis and coherence. Int. J. Psychophysiol. 1990, 10, 125–142. [Google Scholar] [CrossRef]
- Terhune, D.B.; Cardeña, E.; Lindgren, M. Differential frontal-parietal phase synchrony during hypnosis as a function of hypnotic suggestibility. Psychophysiology 2011, 48, 1444–1447. [Google Scholar] [CrossRef] [PubMed]
- De Pascalis, V.; Palumbo, G. EEG alpha asymmetry: Task difficulty and hypnotizability. Percept. Mot. Skills 1986, 62, 139–150. [Google Scholar] [CrossRef]
- Graffin, N.F.; Ray, W.J.; Lundy, R. EEG concomitants of hypnosis and hypnotic susceptibility. J. Abnorm. Psychol. 1995, 104, 123–131. [Google Scholar] [CrossRef]
- Crawford, H.J. Cognitive and psychophysiological correlates of hypnotic responsiveness and hypnosis. In Creative Mastery in Hypnosis and Hypnoanalysis: A Festschrift for Erika Fromm; Lawrence Erlbaum Associates, Inc.: Hillsdale, NJ, USA, 1990; pp. 47–54. [Google Scholar]
- MacLeod-Morgan, C.; Lack, L. Hemispheric specificity: A physiological concomitant of hypnotizability. Psychophysiology 1982, 19, 687–690. [Google Scholar] [CrossRef] [PubMed]
- Freeman, R.; Barabasz, A.; Barabasz, M.; Warner, D. Hypnosis and distraction differ in their effects on cold pressor pain. Am. J. Clin. Hypn. 2000, 43, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, D.D.; Dwyer, K.V.; Kelly, S.M. Relationship between QEEG relative power and hypnotic susceptibility. Am. J. Clin. Hypn. 2000, 43, 71–75. [Google Scholar] [CrossRef]
- Schacter, D.L. EEG theta waves and psychological phenomena: A review and analysis. Biol. Psychol. 1977, 5, 47–82. [Google Scholar] [CrossRef]
- Madeo, D.; Castellani, E.; Santarcangelo, E.L.; Mocenni, C. Hypnotic assessment based on the Recurrence Quantification Analysis of EEG recorded in the ordinary state of consciousness. Brain Cogn. 2013, 83, 227–233. [Google Scholar] [CrossRef]
- Chiarucci, R.; Madeo, D.; Loffredo, M.I.; Castellani, E.; Santarcangelo, E.L.; Mocenni, C. Cross-evidence for hypnotic susceptibility through nonlinear measures on EEGs of non-hypnotized subjects. Sci. Rep. 2014, 4, 5610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baghdadi, G.; Nasrabadi, A.M. Comparison of different EEG features in estimation of hypnosis susceptibility level. Comput. Biol. Med. 2012, 42, 590–597. [Google Scholar] [CrossRef]
- Lee, J.S.; Spiegel, D.; Kim, S.B.; Lee, J.H.; Kim, S.I.; Yang, B.H.; Choi, J.H.; Kho, Y.C.; Nam, J.H. Fractal analysis of EEG in hypnosis and its relationship with hypnotizability. Int. J. Clin. Exp. Hypn. 2007, 55, 14–31. [Google Scholar] [CrossRef] [PubMed]
- Yargholi, E.; Nasrabadi, A.M. The impacts of hypnotic susceptibility on chaotic dynamics of EEG signals during standard tasks of Waterloo-Stanford Group Scale. J. Med. Eng. Technol. 2013, 37, 273–281. [Google Scholar] [CrossRef]
- Yargholi, E.; Nasrabadi, A.M. Chaos–chaos transition of left hemisphere EEGs during standard tasks of Waterloo-Stanford Group Scale of hypnotic susceptibility. J. Med. Eng. Technol. 2015, 39, 281–285. [Google Scholar] [CrossRef]
- Stam, C.J. Nonlinear dynamical analysis of EEG and MEG: Review of an emerging field. Clin. Neurophysiol. 2005, 116, 2266–2301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.S.; Roy, R.; Jensen, E. EEG complexity as a measure of depth of anesthesia for patients. IEEE Trans. Biomed. Eng. 2001, 48, 1424–1433. [Google Scholar] [CrossRef]
- Ibáñez-Molina, A.J.; Iglesias-Parro, S.; Soriano, M.F.; Aznarte, J.I. Multiscale Lempel-Ziv complexity for EEG measures. Clin. Neurophysiol. 2015, 126, 541–548. [Google Scholar] [CrossRef]
- Tuominen, J.; Kallio, S.; Kaasinen, V.; Railo, H. Segregated brain state during hypnosis. Neurosci. Conscious. 2021, 2021, niab002. [Google Scholar] [CrossRef]
- Lipping, T.; Ferenets, R.; Mortier, E.P.; Struys, M.M.R.F. A new method for evaluating the performance of depth-of-hypnosis indices-the D-value. In Proceedings of the 2007 29th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Lyon, France, 22–26 August 2007; pp. 6487–6490. [Google Scholar] [CrossRef]
- Bai, Y.; Liang, Z.; Li, X.; Voss, L.J.; Sleigh, J.W. Permutation Lempel–Ziv complexity measure of electroencephalogram in GABAergic anaesthetics. Physiol. Meas. 2015, 36, 2483. [Google Scholar] [CrossRef] [PubMed]
- Hudetz, A.G.; Liu, X.; Pillay, S.; Boly, M.; Tononi, G. Propofol anesthesia reduces Lempel-Ziv complexity of spontaneous brain activity in rats. Neurosci. Lett. 2016, 628, 132–135. [Google Scholar] [CrossRef] [Green Version]
- Hinterberger, T.; Schoner, J.; Halsband, U. Analysis of electrophysiological state patterns and changes during hypnosis induction. Int. J. Clin. Exp. Hypn. 2011, 59, 165–179. [Google Scholar] [CrossRef] [PubMed]
- De Pascalis, V.; Ray, W.J.; Tranquillo, I.; D’Amico, D. EEG activity and heart rate during recall of emotional events in hypnosis: Relationships with hypnotizability and suggestibility. Int. J. Psychophysiol. 1998, 29, 255–275. [Google Scholar] [CrossRef]
- De Pascalis, V. EEG spectral analysis during hypnotic induction, hypnotic dream and age regression. Int. J. Psychophysiol. Off. J. Int. Organ. Psychophysiol. 1993, 15, 153–166. [Google Scholar] [CrossRef]
- Barry, R.J.; Clarke, A.R.; Johnstone, S.J.; Magee, C.A.; Rushby, J.A. EEG differences between eyes-closed and eyes-open resting conditions. Clin. Neurophysiol. 2007, 118, 2765–2773. [Google Scholar] [CrossRef]
- De Pascalis, V.; Scacchia, P. Hypnotizability and placebo analgesia in waking and hypnosis as modulators of auditory startle responses in healthy women: An ERP study. PLoS ONE 2016, 11, e0159135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumas, R.A. EEG alpha-hypnotizability correlations: A review. Psychophysiology 1977, 14, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Barabasz, A.F. EEG alpha-hypnotizability correlations are not simple covariates of subject self-selection. Biol. Psychol. 1983, 17, 169–172. [Google Scholar] [CrossRef]
- Perlini, A.H.; Spanos, N.P. EEG alpha methodologies and hypnotizability: A critical review. Psychophysiology 1991, 28, 511–530. [Google Scholar] [CrossRef]
- Depascalis, V.; Silveri, A.; Palumbo, G. EEG asymmetry during covert mental activity and its relationship with hypnotizability. Int. J. Clin. Exp. Hypn. 1988, 36, 38–52. [Google Scholar] [CrossRef]
- Cardeña, E.; Kallio, S.; Terhune, D.B.; Buratti, S.; Lööf, A. The effects of translation and sex on hypnotizability testing. Contemp. Hypn. 2007, 24, 154–160. [Google Scholar] [CrossRef]
- Page, R.A.; Green, J.P. An update on age, hypnotic suggestibility, and gender: A brief report. Am. J. Clin. Hypn. 2007, 49, 283–287. [Google Scholar] [CrossRef]
- Költo, A.; Gosi-Greguss, A.C.; Varga, K.; Bányai, É.I. The Influence of Time and Gender on Hungarian Hypnotizability Scores1. Int. J. Clin. Exp. Hypn. 2014, 62, 84–110. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Salmaso, D.; Longoni, A.M. Problems in the assessment of hand preference. Cortex 1985, 21, 533–549. [Google Scholar] [CrossRef]
- Cacioppo, S.; Bianchi-Demicheli, F.; Bischof, P.; DeZiegler, D.; Michel, C.M.; Landis, T. Hemispheric specialization varies with EEG brain resting states and phase of menstrual cycle. PLoS ONE 2013, 8, e63196. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhou, R.; Cui, H.; Wu, M.; Wang, Q.; Zhao, Y.; Liu, Y. Variations in resting frontal alpha asymmetry between high-and low-neuroticism females across the menstrual cycle. Psychophysiology 2015, 52, 182–191. [Google Scholar] [CrossRef] [PubMed]
- De Pascalis, V.; Bellusci, A.; Russo, P.M. Italian norms for the Stanford hypnotic susceptibility scale, form C. Int. J. Clin. Exp. Hypn. 2000, 48, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Weitzenhoffer, A.M.; Hilgard, E.R. Stanford Hypnotic Susceptibility Scale, Form C; Consulting Psychologists Press: Palo Alto, CA, USA, 1962; Volume 27. [Google Scholar]
- Spielberger, C.D. State-Trait anxiety inventory. Corsini Encycl. Psychol. 2010, 1-1. [Google Scholar]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullen, T.R.; Kothe, C.A.E.; Chi, Y.M.; Ojeda, A.; Kerth, T.; Makeig, S.; Jung, T.P.; Cauwenberghs, G. Real-Time Neuroimaging and Cognitive Monitoring Using Wearable Dry EEG. IEEE Trans. Bio-Med. Eng. 2015, 62, 2553–2567. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.Y.; Hsu, S.H.; Pion-Tonachini, L.; Jung, T.P. Evaluation of Artifact Subspace Reconstruction for Automatic Artifact Components Removal in Multi-Channel EEG Recordings. IEEE Trans. Biomed. Eng. 2020, 67, 1114–1121. [Google Scholar] [CrossRef]
- Palmer, J.A.; Kreutz-Delgado, K.; Makeig, S. AMICA: An Adaptive Mixture of Independent Component Analyzers with Shared Components; Technical Report; Swartz Center for Computatonal Neursoscience, University of California San Diego: San Diego, CA, USA, 2012. [Google Scholar]
- Lempel, A.; Ziv, J. On the Complexity of Finite Sequences. IEEE Trans. Inf. Theory 1976, 22, 75–81. [Google Scholar] [CrossRef]
- Kaspar, F.; Schuster, H. Easily calculable measure for the complexity of spatiotemporal patterns. Phys. Rev. A 1987, 36, 842. [Google Scholar] [CrossRef] [PubMed]
- Abásolo, D.; Hornero, R.; Gómez, C.; García, M.; López, M. Analysis of EEG background activity in Alzheimer’s disease patients with Lempel–Ziv complexity and central tendency measure. Med. Eng. Phys. 2006, 28, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Al-Nuaimi, A.H.H.; Jammeh, E.; Sun, L.; Ifeachor, E. Complexity measures for quantifying changes in electroencephalogram in Alzheimer’s disease. Complexity 2018, 2018, 8915079. [Google Scholar] [CrossRef]
- Tsallis, C. Possible generalization of Boltzmann-Gibbs statistics. J. Stat. Phys. 1988, 52, 479–487. [Google Scholar] [CrossRef]
- Tsallis, C. Nonextensive statistics: Theoretical, experimental and computational evidences and connections. Braz. J. Phys. 1999, 29, 1–35. [Google Scholar] [CrossRef]
- Gell-Mann, M.; Tsallis, C. Nonextensive Entropy: Interdisciplinary Applications; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Scott, D.W. Sturges’ rule. Wiley Interdiscip. Rev. Comput. Stat. 2009, 1, 303–306. [Google Scholar] [CrossRef]
- Zhang, D.; Jia, X.; Ding, H.; Ye, D.; Thakor, N.V. Application of Tsallis entropy to EEG: Quantifying the presence of burst suppression after asphyxial cardiac arrest in rats. IEEE Trans. Biomed. Eng. 2009, 57, 867–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- Peritz, E. Book Reviews: Multiple Comparison Procedures Y. Hochberg and A. C. Tamhane New York: Wiley, 1987. xxii + 450 pp. J. Educ. Stat. 1989, 14, 103–106. [Google Scholar] [CrossRef]
- Klinger, E.; Gregoire, K.C.; Barta, S.G. Physiological correlates of mental activity: Eye movements, alpha, and heart rate during imagining, suppression, concentration, search, and choice. Psychophysiology 1973, 10, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Cooper, N.R.; Burgess, A.P.; Croft, R.J.; Gruzelier, J.H. Investigating evoked and induced electroencephalogram activity in task-related alpha power increases during an internally directed attention task. Neuroreport 2006, 17, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Crawford, H. Cognitive and physiological flexibility: Multiple pathways to hypnotic responsiveness. In Suggestion and Suggestibility; Springer: Berlin/Heidelberg, Germany, 1989; pp. 155–167. [Google Scholar]
- Crawford, H.J.; Gruzelier, J.H. A midstream view of the neuropsychophysiology of hypnosis: Recent research and future directions. In Contemporary Hypnosis Research; Guilford Press: New York, NY, USA, 1992. [Google Scholar]
- Gruzelier, J. The neuropsychology of hypnosis. In Hypnosis: Current Clinical, Experimental and Forensic Practices; Croom Helm: Kent, UK, 1988. [Google Scholar]
- Gruzelier, J. Neuropsychological investigations of hypnosis: Cerebral laterality and beyond. In Hypnosis: Theory, Research and Clinical Practice; Free University Press: Amsterdam, The Netherlands, 1990. [Google Scholar]
- Gruzelier, J. A working model of the neurophysiology of hypnosis: A review of evidence. Contemp. Hypn. 1998, 15, 3–21. [Google Scholar] [CrossRef]
- Sheer, D.E. Sensory and cognitive 40-Hz event-related potentials: Behavioral correlates, brain function, and clinical application. In Brain Dynamics; Springer: Berlin/Heidelberg, Germany, 1989; pp. 339–374. [Google Scholar]
- Pascalis, V.d. Psychophysiological correlates of hypnosis and hypnotic susceptibility. Int. J. Clin. Exp. Hypn. 1999, 47, 117–143. [Google Scholar] [CrossRef]
- Ray, W.J. EEG concomitants of hypnotic susceptibility. Int. J. Clin. Exp. Hypn. 1997, 45, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Lutzenberger, W.; Elbert, T.; Birbaumer, N.; Ray, W.J.; Schupp, H. The scalp distribution of the fractal dimension of the EEG and its variation with mental tasks. Brain Topogr. 1992, 5, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Gómez, C.; Hornero, R. Entropy and complexity analyses in Alzheimer’s disease: An MEG study. Open Biomed. Eng. J. 2010, 4, 223. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Xu, M.; Zhou, P.; He, F.; Ming, D. EEG complexity and functional connectivity during precise timing prediction. In Proceedings of the 2019 41st Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Berlin, Germany, 23–27 July 2019; IEEE: Piscataway, NJ, USA, 2019; pp. 2909–2912. [Google Scholar]
- Mohammadi, Y.; Moradi, M.H. Prediction of Depression Severity Scores Based on Functional Connectivity and Complexity of the EEG Signal. Clin. EEG Neurosci. 2021, 52, 52–60. [Google Scholar] [CrossRef]
- Friston, K.J. Theoretical neurobiology and schizophrenia. Br. Med. Bull. 1996, 52, 644–655. [Google Scholar] [CrossRef]
- Friston, K.J. Dysfunctional connectivity in schizophrenia. World Psychiatry 2002, 1, 66. [Google Scholar] [PubMed]
- Wang, D.J.; Jann, K.; Fan, C.; Qiao, Y.; Zang, Y.F.; Lu, H.; Yang, Y. Neurophysiological basis of multi-scale entropy of brain complexity and its relationship with functional connectivity. Front. Neurosci. 2018, 12, 352. [Google Scholar] [CrossRef] [PubMed]
- Baccalá, L.A.; Sameshima, K. Partial directed coherence: A new concept in neural structure determination. Biol. Cybern. 2001, 84, 463–474. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HS vs. MS | |||
| median ± mad | median ± mad | p-value | |
| Fp1 | 0.4146 ± 0.0424 | 0.4861 ± 0.0491 | 0.0317 |
| F7 | 0.4225 ± 0.0374 | 0.4943 ± 0.0381 | 0.0232 |
| F3 | 0.4126 ± 0.0262 | 0.4784 ± 0.0391 | 0.0324 |
| Pz | 0.4106 ± 0.0278 | 0.4458 ± 0.0359 | 0.1233 |
| HS vs. LS | |||
| median ± mad | median ± mad | p-value | |
| Fp1 | 0.4146 ± 0.0424 | 0.4650 ± 0.0285 | 0.2317 |
| F7 | 0.4225 ± 0.0374 | 0.4837 ± 0.0310 | 0.0615 |
| F3 | 0.4126 ± 0.0262 | 0.4664 ± 0.0248 | 0.0597 |
| Pz | 0.4106 ± 0.0278 | 0.4465 ± 0.0258 | 0.0404 |
| MS vs. LS | |||
| median ± mad | median ± mad | p-value | |
| Fp1 | 0.4146 ± 0.0424 | 0.4650 ± 0.0285 | 0.7812 |
| F7 | 0.4225 ± 0.0374 | 0.4837 ± 0.0310 | 0.9971 |
| F3 | 0.4126 ± 0.0262 | 0.4664 ± 0.0248 | 0.9979 |
| Pz | 0.4106 ± 0.0278 | 0.4465 ± 0.0258 | 0.7097 |
| HS vs. MS | ||||
| median ± mad | median ± mad | p-value | ||
| F7 | 0.4531 ± 0.0079 | 0.4601 ± 0.0108 | 0.4469 | |
| HS vs. LS | ||||
| median ± mad | median ± mad | p-value | ||
| F7 | 0.4531 ± 0.0079 | 0.4704 ± 0.0088 | 0.0144 | |
| MS vs. LS | ||||
| median ± mad | median ± mad | p-value | ||
| F7 | 0.4601 ± 0.0108 | 0.4704 ± 0.0088 | 0.1247 | |
| HS vs. MS | ||||
| median ± mad | median ± mad | p-value | ||
| C3 | 0.2352 ± 0.0206 | 0.2478 ± 0.0159 | 0.6142 | |
| C4 | 0.2299 ± 0.0162 | 0.2508 ± 0.0161 | 0.1663 | |
| T5 | 0.2339 ± 0.0094 | 0.2472 ± 0.0122 | 0.1472 | |
| Pz | 0.2299 ± 0.0136 | 0.2455 ± 0.0139 | 0.1423 | |
| HS vs. LS | ||||
| median ± mad | median ± mad | p-value | ||
| C3 | 0.2352 ± 0.0206 | 0.2599 ± 0.0098 | 0.0154 | |
| C4 | 0.2299 ± 0.0162 | 0.2579 ± 0.0109 | 0.0166 | |
| T5 | 0.2339 ± 0.0094 | 0.2579 ± 0.0120 | 0.0404 | |
| Pz | 0.2299 ± 0.0136 | 0.2532 ± 0.0158 | 0.0485 | |
| MS vs. LS | ||||
| median ± mad | median ± mad | p-value | ||
| C3 | 0.2478 ± 0.0159 | 0.2599 ± 0.0098 | 0.0722 | |
| C4 | 0.2508 ± 0.0161 | 0.2579 ± 0.0109 | 0.3966 | |
| T5 | 0.2472 ± 0.0122 | 0.2579 ± 0.0120 | 0.6563 | |
| Pz | 0.2455 ± 0.0139 | 0.2532 ± 0.0158 | 0.7168 | |
| HS vs. MS | ||||
| median ± mad | median ± mad | p-value | ||
| C3 | 0.2425 ± 0.0271 | 0.2372 ± 0.0152 | 0.8871 | |
| HS vs. LS | ||||
| median ± mad | median ± mad | p-value | ||
| C3 | 0.2425 ± 0.0271 | 0.2578 ± 0.0135 | 0.0547 | |
| MS vs. LS | ||||
| median ± mad | median ± mad | p-value | ||
| C3 | 0.2372 ± 0.0152 | 0.2578 ± 0.0135 | 0.0809 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rho, G.; Callara, A.L.; Petri, G.; Nardelli, M.; Scilingo, E.P.; Greco, A.; Pascalis, V.D. Linear and Nonlinear Quantitative EEG Analysis during Neutral Hypnosis following an Opened/Closed Eye Paradigm. Symmetry 2021, 13, 1423. https://doi.org/10.3390/sym13081423
Rho G, Callara AL, Petri G, Nardelli M, Scilingo EP, Greco A, Pascalis VD. Linear and Nonlinear Quantitative EEG Analysis during Neutral Hypnosis following an Opened/Closed Eye Paradigm. Symmetry. 2021; 13(8):1423. https://doi.org/10.3390/sym13081423
Chicago/Turabian StyleRho, Gianluca, Alejandro Luis Callara, Giovanni Petri, Mimma Nardelli, Enzo Pasquale Scilingo, Alberto Greco, and Vilfredo De Pascalis. 2021. "Linear and Nonlinear Quantitative EEG Analysis during Neutral Hypnosis following an Opened/Closed Eye Paradigm" Symmetry 13, no. 8: 1423. https://doi.org/10.3390/sym13081423
APA StyleRho, G., Callara, A. L., Petri, G., Nardelli, M., Scilingo, E. P., Greco, A., & Pascalis, V. D. (2021). Linear and Nonlinear Quantitative EEG Analysis during Neutral Hypnosis following an Opened/Closed Eye Paradigm. Symmetry, 13(8), 1423. https://doi.org/10.3390/sym13081423