
Effect of Microgravity Environment on Gut Microbiome and Angiogenesis

 ,
,  ,
,  and
and 
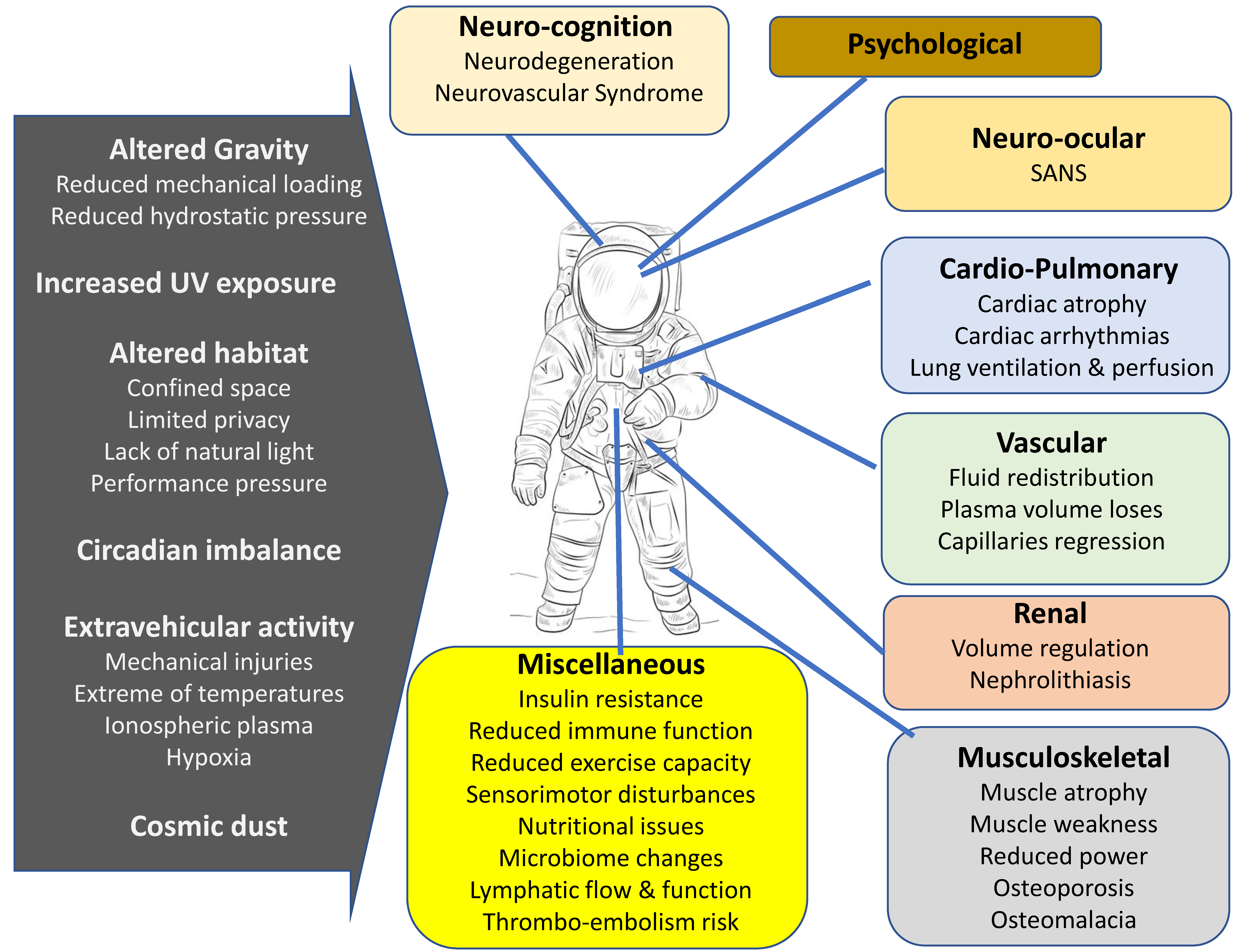{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Role of Gut Microbiota in Astronaut Health
3. Angiogenesis and the Influence of Microgravity
4. Gut Microbiome, Angiogenesis and Link with Visual Impairment
5. Angiogenesis and Skeletal Muscle
6. Metabolite Drivers of Angiogenesis and the Gut Microbiome
7. Utilization of Pro-, Pre- and Post-Biotics
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goswami, N.; White, O.; Blaber, A.; Evans, J.; van Loon, J.J.; Clement, G. Human physiology adaptation to altered gravity environments. Acta Astronaut. 2021, 189, 216–221. [Google Scholar] [CrossRef]
- Goswami, N.; Van Loon, J.J.W.A.; Roessler, A.; Blaber, A.P.; White, O. Editorial: Gravitational Physiology, Aging and Medicine. Front. Physiol. 2019, 10, 1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, R.; Akbar, N.; Khan, N.A. Gut microbiome and human health under the space environment. J. Appl. Microbiol. 2020, 130, 14–24. [Google Scholar] [CrossRef]
- Laurini, K.; Piedboeuf, J.C.; Schade, B.; Matsumoto, K.; Spiero, F.; Lorenzoni, A.; The Global Exploration Roadmap. IAC-11 B. 2018. Available online: https://www.globalspaceexploration.org/wordpress/wp-content/uploads/IAC62/ISECG%20Global%20Exploration%20Roadmap.pdf (accessed on 20 August 2021).
- Garrett-Bakelman, F.E.; Darshi, M.; Green, S.J.; Gur, R.C.; Lin, L.; Macias, B.R.; McKenna, M.J.; Meydan, C.; Mishra, T.; Nasrini, J.; et al. The NASA Twins Study: A multidimensional analysis of a year-long human spaceflight. Science 2019, 364, eaau8650. [Google Scholar] [CrossRef] [PubMed]
- Kinross, J.M.; Darzi, A.W.; Nicholson, J.K. Gut microbiome-host interactions in health and disease. Genome Med. 2011, 3, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mohajeri, M.H.; Brummer, R.J.M.; Rastall, R.A.; Weersma, R.K.; Harmsen, H.J.M.; Faas, M.; Eggersdorfer, M. The role of the microbiome for human health: From basic science to clinical applications. Eur. J. Nutr. 2018, 57, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Valdes, A.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef] [Green Version]
- Sampson, T.R.; Mazmanian, S.K. Control of Brain Development, Function, and Behavior by the Microbiome. Cell Host Microbe 2015, 17, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Zeevi, D.; Korem, T.; Zmora, N.; Israeli, D.; Rothschild, D.; Weinberger, A.; Ben-Yacov, O.; Lador, D.; Avnit-Sagi, T.; Lotan-Pompan, M.; et al. Personalized Nutrition by Prediction of Glycemic Responses. Cell 2015, 163, 1079–1094. [Google Scholar] [CrossRef] [Green Version]
- Veronese, N.; Solmi, M.; Caruso, M.G.; Giannelli, G.; Osella, A.R.; Evangelou, E.; Maggi, S.; Fontana, L.; Stubbs, B.; Tzoulaki, I. Dietary fiber and health outcomes: An umbrella review of systematic reviews and meta-analyses. Am. J. Clin. Nutr. 2018, 107, 436–444. [Google Scholar] [CrossRef] [Green Version]
- Voorhies, A.A.; Ott, C.M.; Mehta, S.; Pierson, D.L.; Crucian, B.E.; Feiveson, A.; Oubre, C.M.; Torralba, M.; Moncera, K.; Zhang, Y.; et al. Study of the impact of long-duration space missions at the International Space Station on the astronaut microbiome. Sci. Rep. 2019, 9, 9911. [Google Scholar] [CrossRef] [PubMed]
- Andriessen, E.M.; Wilson, A.M.; Mawambo, G.; Dejda, A.; Miloudi, K.; Sennlaub, F.; Sapieha, P. Gut microbiota influences pathological angiogenesis in obesity-driven choroidal neovascularization. EMBO Mol. Med. 2016, 8, 1366–1379. [Google Scholar] [CrossRef] [PubMed]
- Mader, T.H.; Gibson, C.R.; Otto, C.A.; Sargsyan, A.E.; Miller, N.R.; Subramanian, P.S.; Hart, S.F.; Lipsky, W.; Patel, N.B.; Lee, A.G. Persistent Asymmetric Optic Disc Swelling After Long-Duration Space Flight: Implications for Pathogenesis. J. Neuro-Ophthalmol. 2017, 37, 133–139. [Google Scholar] [CrossRef]
- Lee, A.G.; Mader, T.H.; Gibson, C.R.; Brunstetter, T.J.; Tarver, W.J. Space flight-associated neuro-ocular syndrome (SANS). Eye 2018, 32, 1164–1167. [Google Scholar] [CrossRef] [Green Version]
- Adair, T.H.; Montani, J.P. Angiogenesis. In Colloquium Series on Integrated Systems Physiology: From Molecule to Function; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2010. [Google Scholar]
- Cabral, T.; Mello, L.G.M.; Lima, L.H.; Polido, J.; Regatieri, C.V.; Belfort, R., Jr.; Mahajan, V.B. Retinal and choroidal angiogenesis: A review of new targets. Int. J. Retin. Vitr. 2017, 3, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vico, L.; Hargens, A. Skeletal changes during and after spaceflight. Nat. Rev. Rheumatol. 2018, 14, 229–245. [Google Scholar] [CrossRef]
- Baum, O.; Olfert, I.M.; Egginton, S.; Hellsten, Y. Advances and challenges in skeletal muscle angiogenesis. Am. J. Physiol.-Heart Circ. Physiol. 2016, 310, H326–H336. [Google Scholar] [CrossRef]
- Turroni, S.; Rampelli, S.; Biagi, E.; Consolandi, C.; Severgnini, M.; Peano, C.; Quercia, S.; Soverini, M.; Carbonero, F.G.; Bianconi, G.; et al. Temporal dynamics of the gut microbiota in people sharing a confined environment, a 520-day ground-based space simulation, MARS500. Microbiome 2017, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bull, M.J.; Plummer, N.T. Part 1: The Human Gut Microbiome in Health and Disease. Integr. Med. (Encinitas Calif.) 2014, 13, 17–22. [Google Scholar]
- Kang, D.-W.; Adams, J.B.; Gregory, A.C.; Borody, T.; Chittick, L.; Fasano, A.; Khoruts, A.; Geis, E.; Maldonado, J.; McDonough-Means, S.; et al. Microbiota Transfer Therapy alters gut ecosystem and improves gastrointestinal and autism symptoms: An open-label study. Microbiome 2017, 5, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.; Minuto, C.; Cryan, J.F.; Clarke, G.; Dinan, T.G. Cross Talk: The Microbiota and Neurodevelopmental Disorders. Front. Neurosci. 2017, 11, 490. [Google Scholar] [CrossRef] [Green Version]
- Ghoshal, U.C.; Shukla, R.; Ghoshal, U.; Gwee, K.-A.; Ng, S.C.; Quigley, E.M.M. The Gut Microbiota and Irritable Bowel Syndrome: Friend or Foe? Int. J. Inflamm. 2012, 2012, 151085. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- Amedei, A.; Morbidelli, L. Circulating Metabolites Originating from Gut Microbiota Control Endothelial Cell Function. Molecules 2019, 24, 3992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voorhies, A.A.; Lorenzi, H.A. The Challenge of Maintaining a Healthy Microbiome during Long-Duration Space Missions. Front. Astron. Space Sci. 2016, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Dietlein, L.F. Skylab: A Beginning. 1977. Available online: https://ntrs.nasa.gov/api/citations/19860011667/downloads/19860011667.pdf?attachment=true (accessed on 20 August 2021).
- Henney, M.R.; Raylor, G.R.; Molina, T.C. Mycological profile of crew during 56-day simulated orbital flight. Mycopathologia 1978, 63, 131–144. [Google Scholar] [CrossRef]
- Nicogossian, A.E.; Doarn, C.R.; Hu, Y. Evolution of Human Capabilities and Space Medicine. In Space Physiology and Medicine; Springer: New York, NY, USA, 2016; pp. 3–57. [Google Scholar]
- Brereton, N.; Pitre, F.; Gonzalez, E. Reanalysis of the Mars500 experiment reveals common gut microbiome alterations in astronauts induced by long-duration confinement. Comput. Struct. Biotechnol. J. 2021, 19, 2223–2235. [Google Scholar] [CrossRef] [PubMed]
- Avila-Herrera, A.; Thissen, J.; Urbaniak, C.; Be, N.A.; Smith, D.J.; Karouia, F.; Mehta, S.; Venkateswaran, K.; Jaing, C. Crewmember microbiome may influence microbial composition of ISS habitable surfaces. PLoS ONE 2020, 15, e0231838. [Google Scholar] [CrossRef] [PubMed]
- Morrison, M.D.; Thissen, J.B.; Karouia, F.; Mehta, S.; Urbaniak, C.; Venkateswaran, K.; Smith, D.J.; Jaing, C. Investigation of Spaceflight Induced Changes to Astronaut Microbiomes. Front. Microbiol. 2021, 12, 1368. [Google Scholar] [CrossRef]
- Tirumalai, M.R.; Karouia, F.; Tran, Q.; Stepanov, V.G.; Bruce, R.J.; Ott, C.M.; Pierson, D.L.; Fox, G.E. The adaptation of Escherichia coli cells grown in simulated microgravity for an extended period is both phenotypic and genomic. npj Microgravity 2017, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tirumalai, M.R.; Karouia, F.; Tran, Q.; Stepanov, V.G.; Bruce, R.J.; Ott, C.M.; Pierson, D.L.; Fox, G.E. Evaluation of Acquired Antibiotic Resistance in Escherichia coli Exposed to Long-Term Low-Shear Modeled Microgravity and Background Antibiotic Exposure. mBio 2019, 10, e02637-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Han, C.; Lu, Z.; Ge, P.; Cui, Y.; Zhao, D.; Yang, X.; Wu, B.; Qiang, L.; Zhang, Y.; et al. Simulated microgravity suppresses MAPK pathway-mediated innate immune response to bacterial infection and induces gut microbiota dysbiosis. FASEB J. 2020, 34, 14631–14644. [Google Scholar] [CrossRef] [PubMed]
- Vroom, M.M.; Rodriguez-Ocasio, Y.; Lynch, J.B.; Ruby, E.G.; Foster, J.S. Modeled microgravity alters lipopolysaccharide and outer membrane vesicle production of the beneficial symbiont Vibrio fischeri. npj Microgravity 2021, 7, 1–10. [Google Scholar] [CrossRef]
- Goswami, N.; Roma, P.G.; De Boever, P.; Clément, G.; Hargens, A.R.; Loeppky, J.A.; Evans, J.M.; Peter Stein, T.; Blaber, A.P.; Van Loon, J.J.W.A.; et al. Using the Moon as a high-fidelity analogue environment to study biological and behavioral effects of long-duration space exploration. Planet Space Sci. 2012, 74, 111–120. [Google Scholar] [CrossRef]
- Maier, J.A.M.; Cialdai, F.; Monici, M.; Morbidelli, L. The Impact of Microgravity and Hypergravity on Endothelial Cells. BioMed Res. Int. 2015, 2015, 434803. [Google Scholar] [CrossRef]
- Cotrupi, S.; Ranzani, D.; Maier, J.A. Impact of modeled microgravity on microvascular endothelial cells. Biochim. Biophys. Acta (BBA)-Bioenerg. 2005, 1746, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Versari, S.; Villa, A.; Bradamante, S.; Maier, J.A. Alterations of the actin cytoskeleton and increased nitric oxide synthesis are common features in human primary endothelial cell response to changes in gravity. Biochim. Biophys. Acta (BBA)-Bioenerg. 2007, 1773, 1645–1652. [Google Scholar] [CrossRef] [Green Version]
- Morbidelli, L.; Monici, M.; Marziliano, N.; Cogoli, A.; Fusi, F.; Waltenberger, J.; Ziche, M. Simulated hypogravity impairs the angiogenic response of endothelium by up-regulating apoptotic signals. Biochem. Biophys. Res. Commun. 2005, 334, 491–499. [Google Scholar] [CrossRef]
- Krüger, M.; Pietsch, J.; Bauer, J.; Kopp, S.; Carvalho, D.T.O.; Baatout, S.; Moreels, M.; Melnik, D.; Wehland, M.; Egli, M.; et al. Growth of Endothelial Cells in Space and in Simulated Microgravity—A Comparison on the Secretory Level. Cell. Physiol. Biochem. 2019, 52, 1039–1060. [Google Scholar]
- Kirchen, M.E.; O’Connor, K.M.; Gruber, H.E.; Sweeney, J.R.; Fras, I.A.; Stover, S.J.; Sarmiento, A.; Marshall, G.J. Effects of microgravity on bone healing in a rat fibular osteotomy model. Clin. Orthop. Relat. Res. 1995, 318, 231–242. [Google Scholar]
- Henry, M.K.; Unsworth, B.R.; Sychev, V.; Guryeva, T.S.; Dadasheva, O.A.; Piert, S.J.; Lagel, K.E.; Dubrovin, L.C.; Jahns, G.C.; Boda, K.; et al. Launch conditions might affect the formation of blood vessels in the quail chorioallantoic membrane. Folia Vet. 1998, 42, S25–S31. [Google Scholar] [PubMed]
- Davidson, J.M.; Aquino, A.M.; Woodward, S.C.; Wilfinger, W.W. Sustained microgravity reduces intrinsic wound healing and growth factor responses in the rat. FASEB J. 1999, 13, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Goswami, N. Falls and Fall-Prevention in Older Persons: Geriatrics Meets Spaceflight! Front Physiol. 2017, 8, 603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswami, N.; Batzel, J.J.; Clément, G.; Stein, T.P.; Hargens, A.; Sharp, M.K.; Blaber, A.P.; Roma, P.G.; Hinghofer-Szalkay, H.G. Maximizing information from space data resources: A case for expanding integration across research disciplines. Graefe’s Arch. Clin. Exp. Ophthalmol. 2012, 113, 1645–1654. [Google Scholar] [CrossRef]
- Delp, M.D.; Charvat, J.M.; Limoli, C.L.; Globus, R.K.; Ghosh, P. Apollo Lunar Astronauts Show Higher Cardiovascular Disease Mortality: Possible Deep Space Radiation Effects on the Vascular Endothelium. Sci. Rep. 2016, 6, 29901. [Google Scholar] [CrossRef] [Green Version]
- Infanger, M.; Kossmehl, P.; Shakibaei, M.; Baatout, S.; Witzing, A.; Grosse, J.; Bauer, J.; Cogoli, A.; Faramarzi, S.; Derradji, H.; et al. Induction of three-dimensional assembly and increase in apoptosis of human endothelial cells by simulated microgravity: Impact of vascular endothelial growth factor. Apoptosis 2006, 11, 749–764. [Google Scholar] [CrossRef]
- Mader, T.H.; Gibson, C.R.; Miller, N.R.; Subramanian, P.S.; Patel, N.B.; Lee, A.G. An overview of spaceflight-associated neuro-ocular syndrome (SANS). Neurol. India 2019, 67, 206–S211. [Google Scholar] [CrossRef] [PubMed]
- Cepeda, C.; Vergnes, L.; Carpo, N.; Schibler, M.J.; Bentolila, L.A.; Karouia, F.; Espinosa-Jeffrey, A. Human Neural Stem Cells Flown into Space Proliferate and Generate Young Neurons. Appl. Sci. 2019, 9, 4042. [Google Scholar] [CrossRef] [Green Version]
- Shaka, S.; Carpo, N.; Tran, V.; Ma, Y.Y.; Karouia, F. Human Neural Stem Cells in Space Proliferate more than Ground Control Cells: Implications for Long-Term Space Travel. J. Stem Cell Res. Dev. Ther. 2021, 7, 69. [Google Scholar]
- Corydon, T.J.; Mann, V.; Slumstrup, L.; Kopp, S.; Sahana, J.; Askou, A.L.; Magnusson, N.E.; Echegoyen, D.; Bek, T.; Sundaresan, A.; et al. Reduced Expression of Cytoskeletal and Extracellular Matrix Genes in Human Adult Retinal Pigment Epithelium Cells Exposed to Simulated Microgravity. Cell. Physiol. Biochem. 2016, 40, 1–17. [Google Scholar] [CrossRef]
- Horai, R.; Zárate-Bladés, C.; Dillenburg-Pilla, P.; Chen, J.; Kielczewski, J.L.; Silver, P.B.; Jittayasothorn, Y.; Chan, C.-C.; Yamane, H.; Honda, K.; et al. Microbiota-Dependent Activation of an Autoreactive T Cell Receptor Provokes Autoimmunity in an Immunologically Privileged Site. Immunity 2015, 43, 343–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, R.; Langmann, T. Gut flora connects obesity with pathological angiogenesis in the eye. EMBO Mol. Med. 2016, 8, 1361–1363. [Google Scholar] [CrossRef]
- Smith, S.M.; McCoy, T.; Gazda, D.; Morgan, J.; Heer, M.; Zwart, S.R. Space Flight Calcium: Implications for Astronaut Health, Spacecraft Operations, and Earth. Nutrients 2012, 4, 2047–2068. [Google Scholar] [CrossRef] [Green Version]
- Honka, M.-J.; Latva-Rasku, A.; Bucci, M.; A Virtanen, K.; Hannukainen, J.; Kalliokoski, K.; Nuutila, P. Insulin-stimulated glucose uptake in skeletal muscle, adipose tissue and liver: A positron emission tomography study. Eur. J. Endocrinol. 2018, 178, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Huenchullan, S.F.; Tam, C.S.; Ban, L.A.; Ehrenfeld-Slater, P.; Mclennan, S.V.; Twigg, S.M. Skeletal muscle adiponectin induction in obesity and exercise. Metabolism 2019, 102, 154008. [Google Scholar] [CrossRef] [Green Version]
- Judge, A.R.; Powers, S.K.; Ferreira, L.F.; Bamman, M.M. Meeting Synopsis: Advances in Skeletal Muscle Biology in Health and Disease (Gainesville, Florida, February 22nd to 24th 2012)—Day 1: “Cell Signaling Mechanisms Mediating Muscle Atrophy and Hypertrophy” and “muscle Force, Calcium Handling, and Stress Response”. Front. Physiol. 2012, 3, 200. [Google Scholar] [PubMed] [Green Version]
- Hendrickse, P.; Degens, H. The role of the microcirculation in muscle function and plasticity. J. Muscle Res. Cell Motil. 2019, 40, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Wagenmakers, A.J.M.; Strauss, J.; Shepherd, S.O.; Keske, M.; Cocks, M. Increased muscle blood supply and transendothelial nutrient and insulin transport induced by food intake and exercise: Effect of obesity and ageing. J. Physiol. 2015, 594, 2207–2222. [Google Scholar] [CrossRef] [Green Version]
- Poole, D.C.; Copp, S.W.; Hirai, D.M.; Musch, T.I. Dynamics of muscle microcirculatory and blood-myocyte O2 flux during contractions. Acta Physiol. 2010, 202, 293–310. [Google Scholar] [CrossRef]
- Laughlin, M.; Roseguini, B. Mechanisms for exercise training-induced increases in skeletal muscle blood flow capacity: Differences with interval sprint training versus aerobic endurance training. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2008, 59, 71–88. [Google Scholar]
- Vincent, M.A.; Clerk, L.H.; Lindner, J.R.; Price, W.J.; Jahn, L.A.; Leong-Poi, H.; Barrett, E.J. Mixed meal and light exercise each recruit muscle capillaries in healthy humans. Am. J. Physiol. Metab. 2006, 290, E1191–E1197. [Google Scholar] [CrossRef] [PubMed]
- Groen, B.B.L.; Hamer, H.M.; Snijders, T.; Van Kranenburg, J.; Frijns, D.; Vink, H.; van Loon, L.J. Skeletal muscle capillary density and microvascular function are compromised with aging and type 2 diabetes. J. Appl. Physiol. 2014, 116, 998–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turroni, S.; Magnani, M.; Kc, P.; Lesnik, P.; Vidal, H.; Heer, M. Gut Microbiome and Space Travelers’ Health: State of the Art and Possible Pro/Prebiotic Strategies for Long-Term Space Missions. Front. Physiol. 2020, 11, 1135. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-M.; Wei, L.; Chiu, Y.-S.; Hsu, Y.-J.; Tsai, T.-Y.; Wang, M.-F.; Huang, C.-C. Lactobacillus plantarum TWK10 Supplementation Improves Exercise Performance and Increases Muscle Mass in Mice. Nutrients 2016, 8, 205. [Google Scholar] [CrossRef] [PubMed]
- Cruys, B.; Wong, B.; Kuchnio, A.; Verdegem, D.; Cantelmo, A.R.; Conradi, L.-C.; Vandekeere, S.; Bouché, A.; Cornelissen, I.; Vinckier, S.; et al. Glycolytic regulation of cell rearrangement in angiogenesis. Nat. Commun. 2016, 7, 12240. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sun, X.; Carmeliet, P. Hallmarks of Endothelial Cell Metabolism in Health and Disease. Cell Metab. 2019, 30, 414–433. [Google Scholar] [CrossRef]
- Marcu, R.; Choi, Y.J.; Xue, J.; Fortin, C.L.; Wang, Y.; Nagao, R.J.; Xu, J.; MacDonald, J.W.; Bammler, T.K.; Murry, C.E.; et al. Human Organ-Specific Endothelial Cell Heterogeneity. iScience 2018, 4, 20–35. [Google Scholar] [CrossRef] [Green Version]
- Burns, P.; Wilson, D. Angiogenesis mediated by metabolites is dependent on vascular endothelial growth factor (VEGF). Angiogenesis 2003, 6, 73–77. [Google Scholar] [CrossRef]
- Scupham, A.J.; Presley, L.L.; Wei, B.; Bent, E.; Griffith, N.; McPherson, M.; Zhu, F.; Oluwadara, O.; Rao, N.; Braun, J.; et al. Abundant and Diverse Fungal Microbiota in the Murine Intestine. Appl. Environ. Microbiol. 2006, 72, 793–801. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, G.R.; Chang, P.V. Deciphering the Chemical Lexicon of Host–Gut Microbiota Interactions. Trends Pharmacol. Sci. 2019, 40, 430–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Humaran, L.G.B.; Gratadoux, J.-J.; Blugeon, S.; Bridonneau, C.; Furet, J.-P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, S.A.; Fu, J.; Chang, P.V. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2020, 117, 19376–19387. [Google Scholar] [CrossRef]
- Zhang, Z.; Tang, H.; Chen, P.; Xie, H.; Tao, Y. Demystifying the manipulation of host immunity, metabolism, and extraintestinal tumors by the gut microbiome. Signal Transduct. Target. Ther. 2019, 4, 1–34. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Castro-Wallace, S.; Stahl, S.; Voorhies, A.; Lorenzi, H.; Douglas, G.L. Response of Lactobacillus acidophilus ATCC 4356 to low-shear modeled microgravity. Acta Astronaut. 2017, 139, 463–468. [Google Scholar] [CrossRef]
- Sakai, T.; Moteki, Y.; Takahashi, T.; Shida, K.; Kiwaki, M.; Shimakawa, Y.; Matsui, A.; Chonan, O.; Morikawa, K.; Ohta, T.; et al. Probiotics into outer space: Feasibility assessments of encapsulated freeze-dried probiotics during 1 month’s storage on the International Space Station. Sci. Rep. 2018, 8, 10687. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Varela, R.G.; Garcia, H.; Mata-Haro, V.; González-Córdova, A.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddiqui, R.; Qaisar, R.; Goswami, N.; Khan, N.A.; Elmoselhi, A. Effect of Microgravity Environment on Gut Microbiome and Angiogenesis. Life 2021, 11, 1008. https://doi.org/10.3390/life11101008
Siddiqui R, Qaisar R, Goswami N, Khan NA, Elmoselhi A. Effect of Microgravity Environment on Gut Microbiome and Angiogenesis. Life. 2021; 11(10):1008. https://doi.org/10.3390/life11101008
Chicago/Turabian StyleSiddiqui, Ruqaiyyah, Rizwan Qaisar, Nandu Goswami, Naveed Ahmed Khan, and Adel Elmoselhi. 2021. "Effect of Microgravity Environment on Gut Microbiome and Angiogenesis" Life 11, no. 10: 1008. https://doi.org/10.3390/life11101008
APA StyleSiddiqui, R., Qaisar, R., Goswami, N., Khan, N. A., & Elmoselhi, A. (2021). Effect of Microgravity Environment on Gut Microbiome and Angiogenesis. Life, 11(10), 1008. https://doi.org/10.3390/life11101008