
Slosh Simulation in a Computer Model of Canine Syringomyelia



Abstract
:1. Introduction
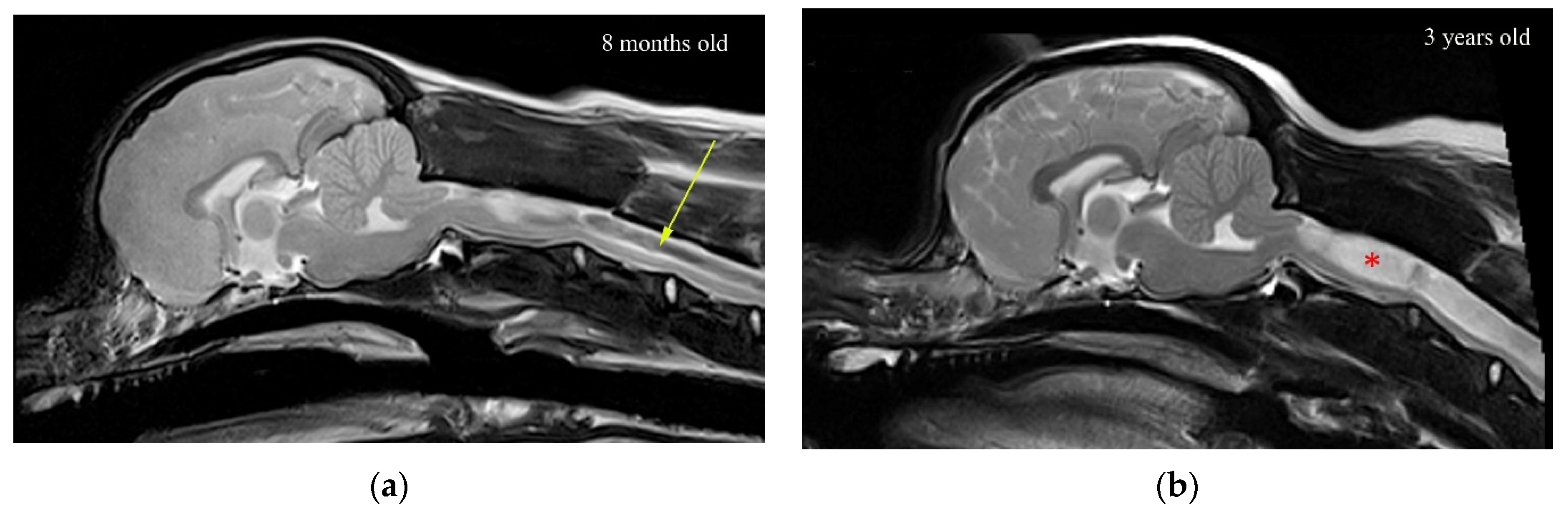
2. Materials and Methods
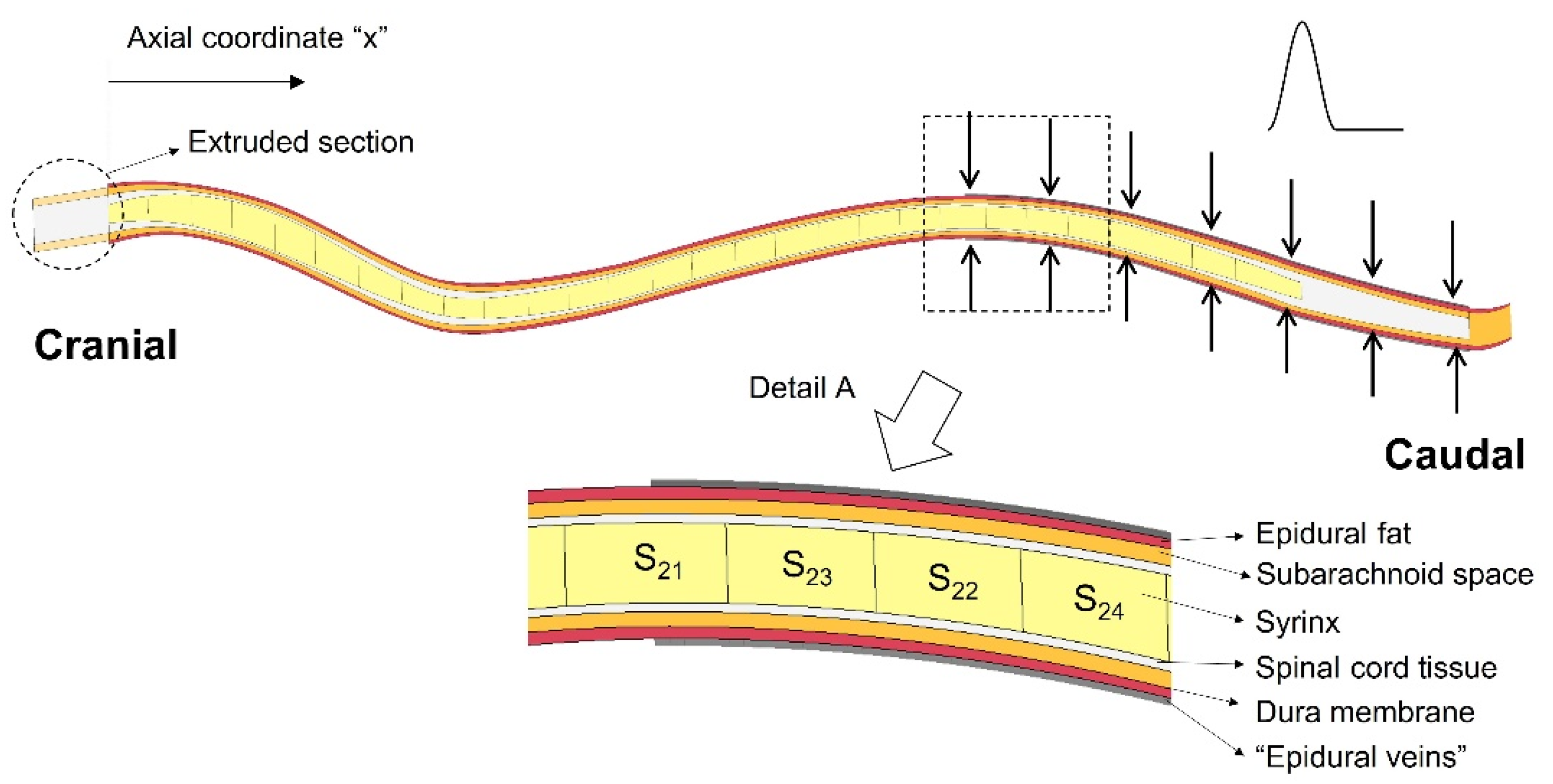
2.1. The Finite Element Model
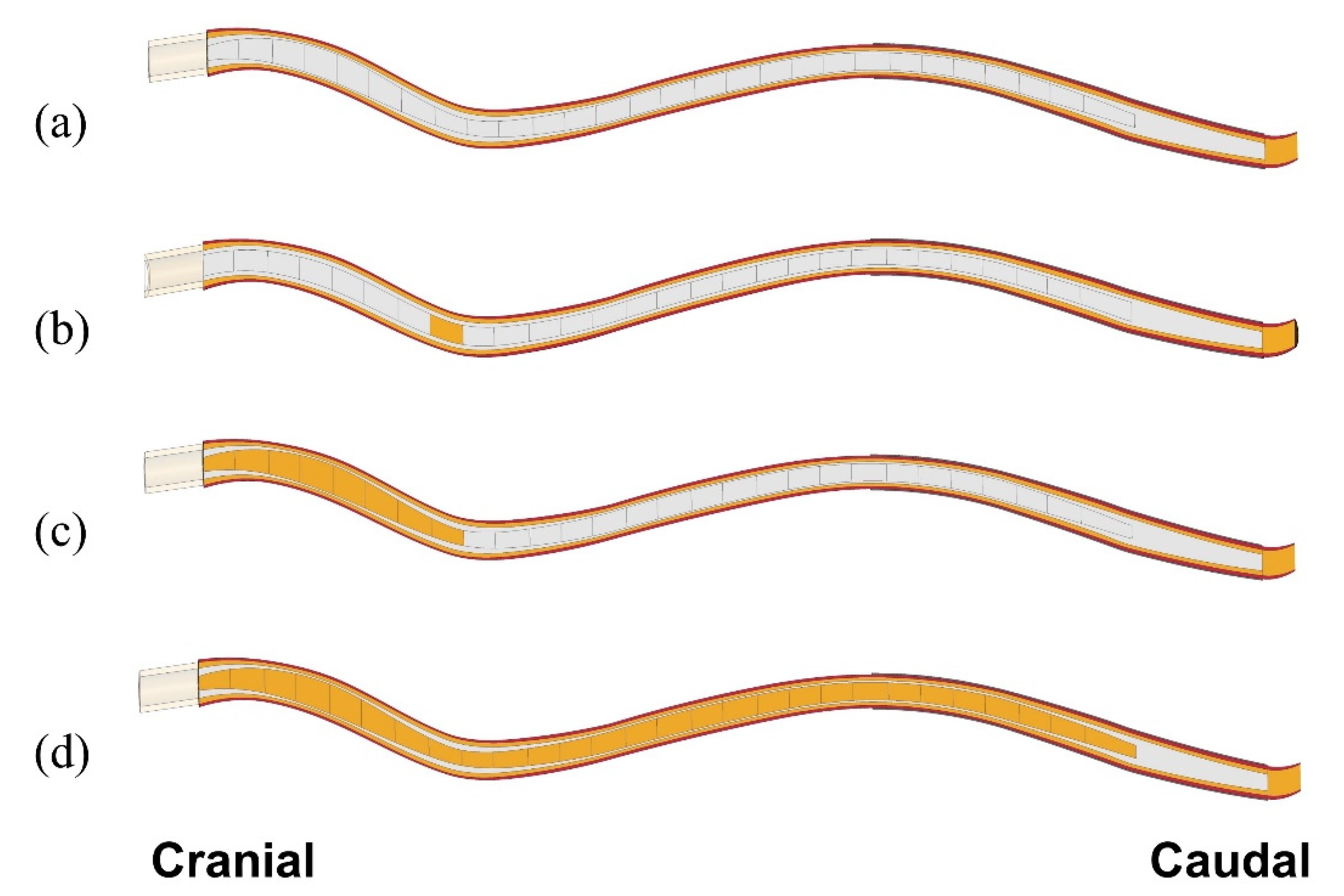
2.2. Simulations
2.3. Data Processing
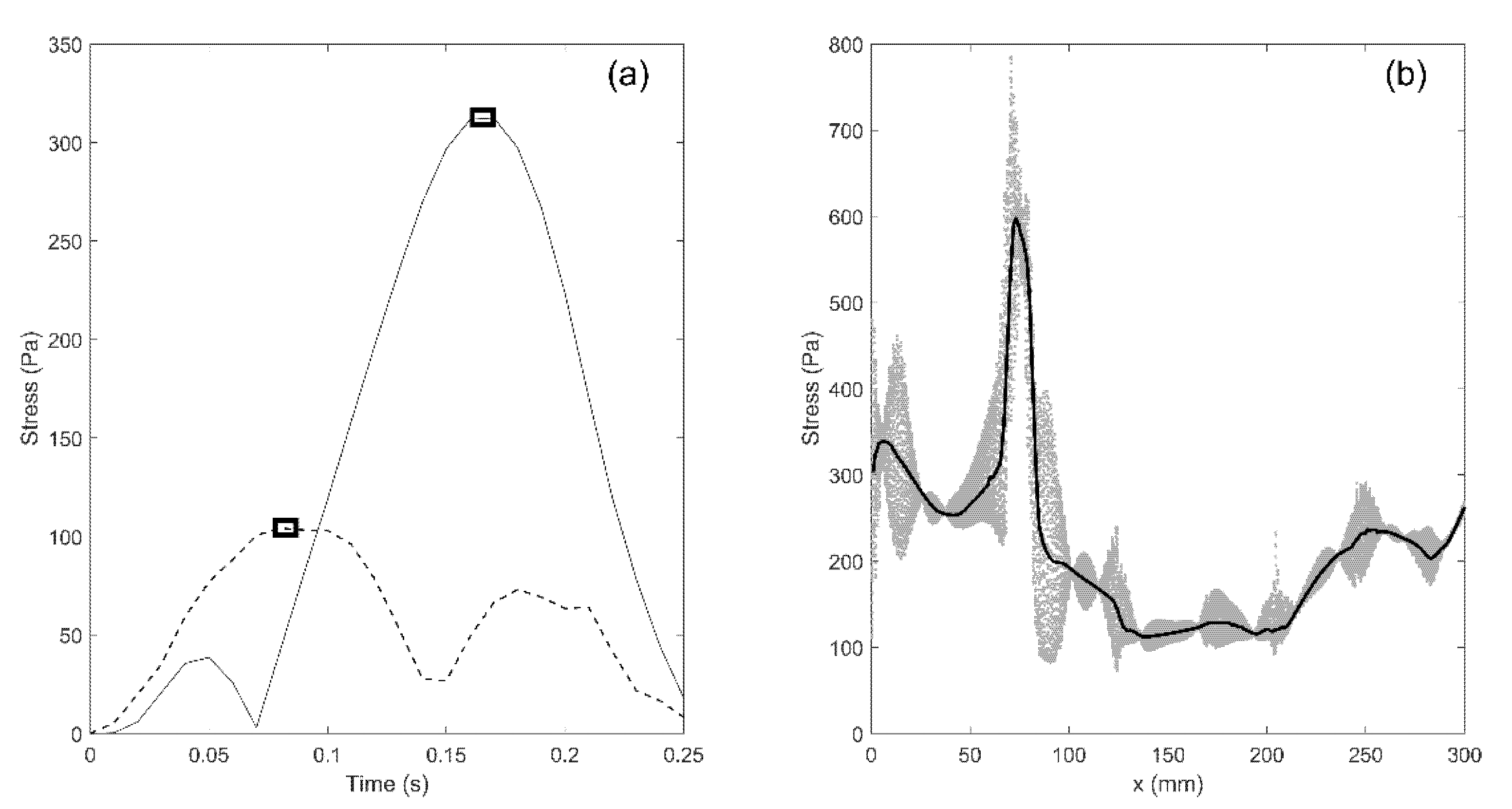
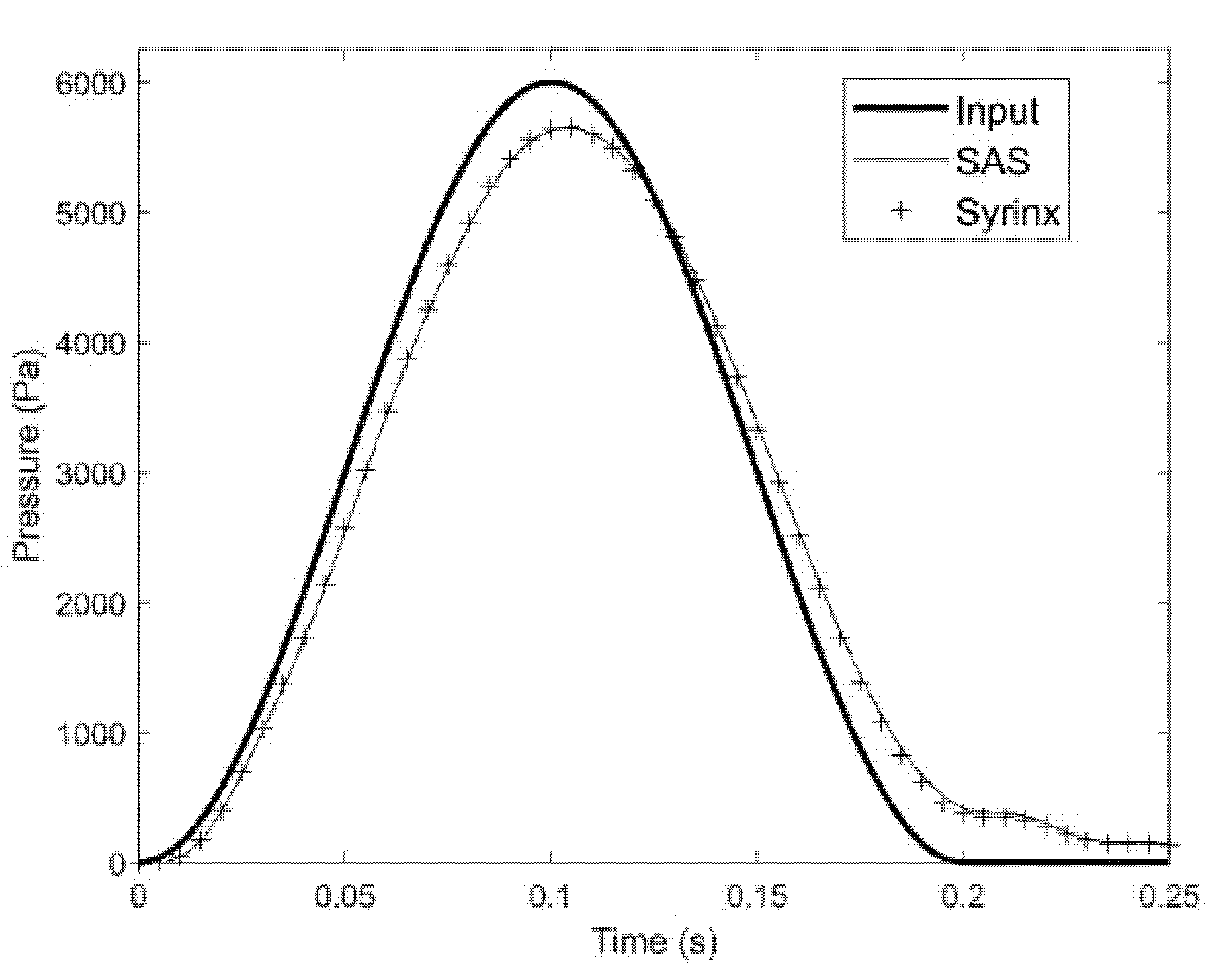
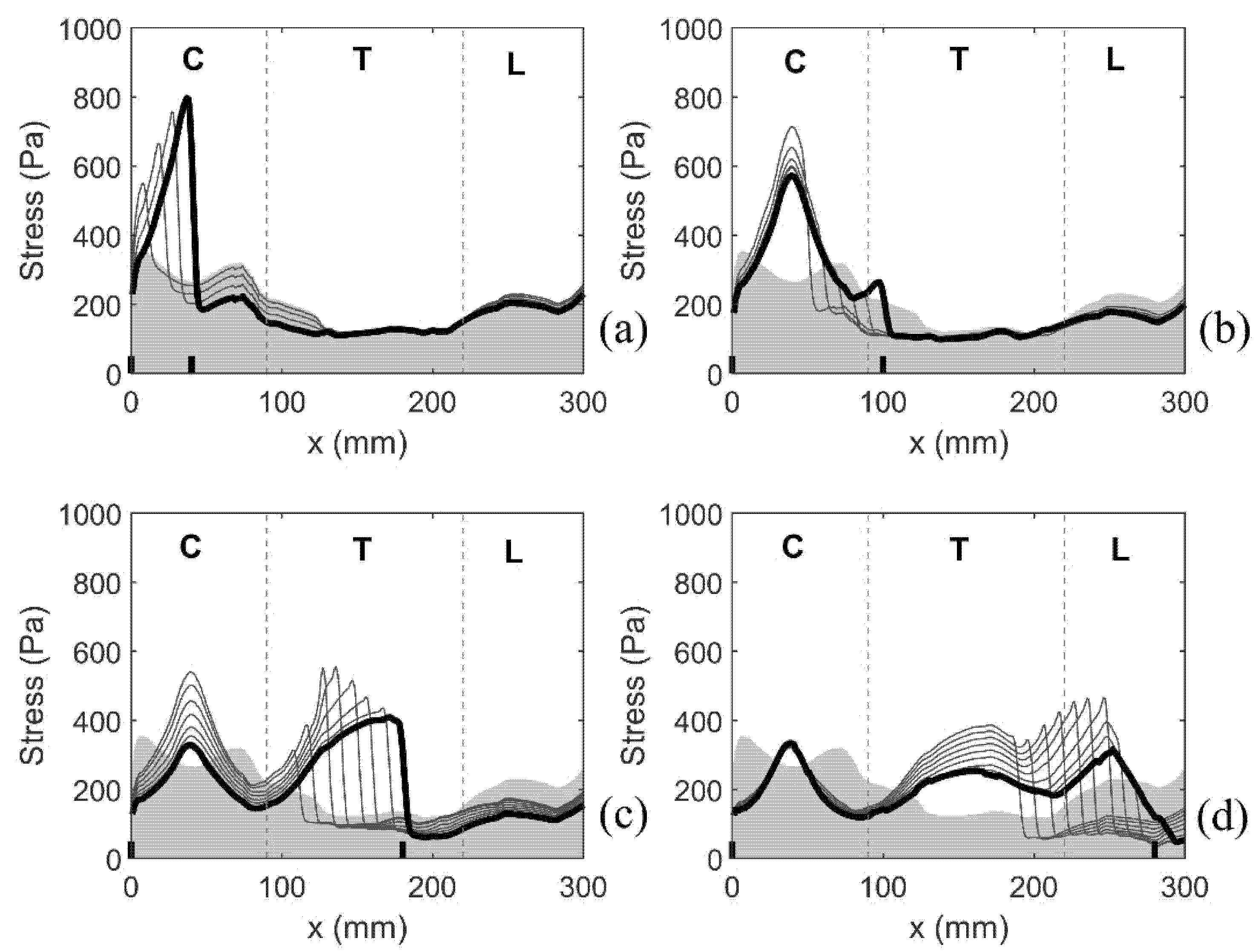
3. Results
3.1. General Observations
3.1.1. Pressure in the SAS and Syringes
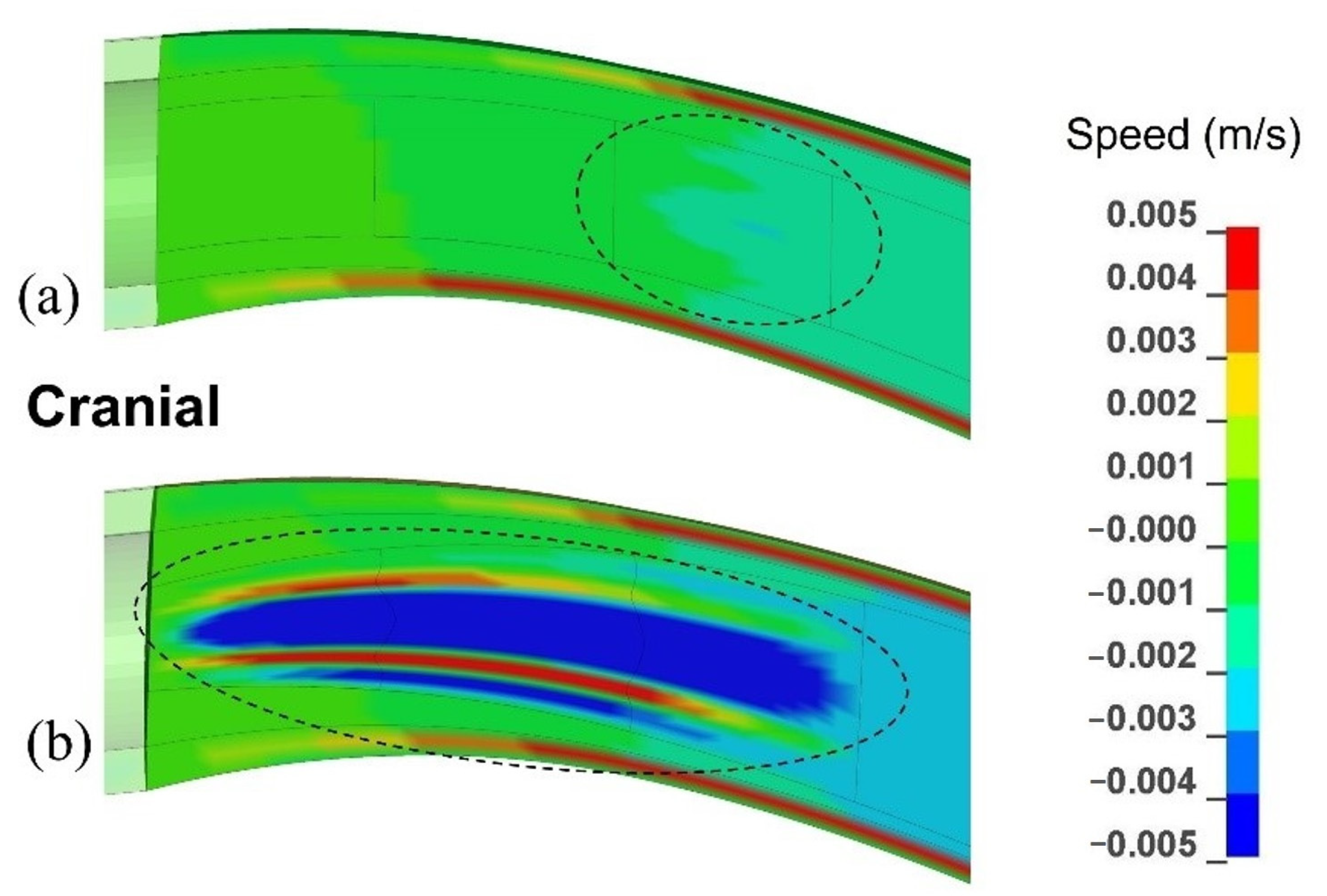
3.1.2. Movement in the Anatomical Layers
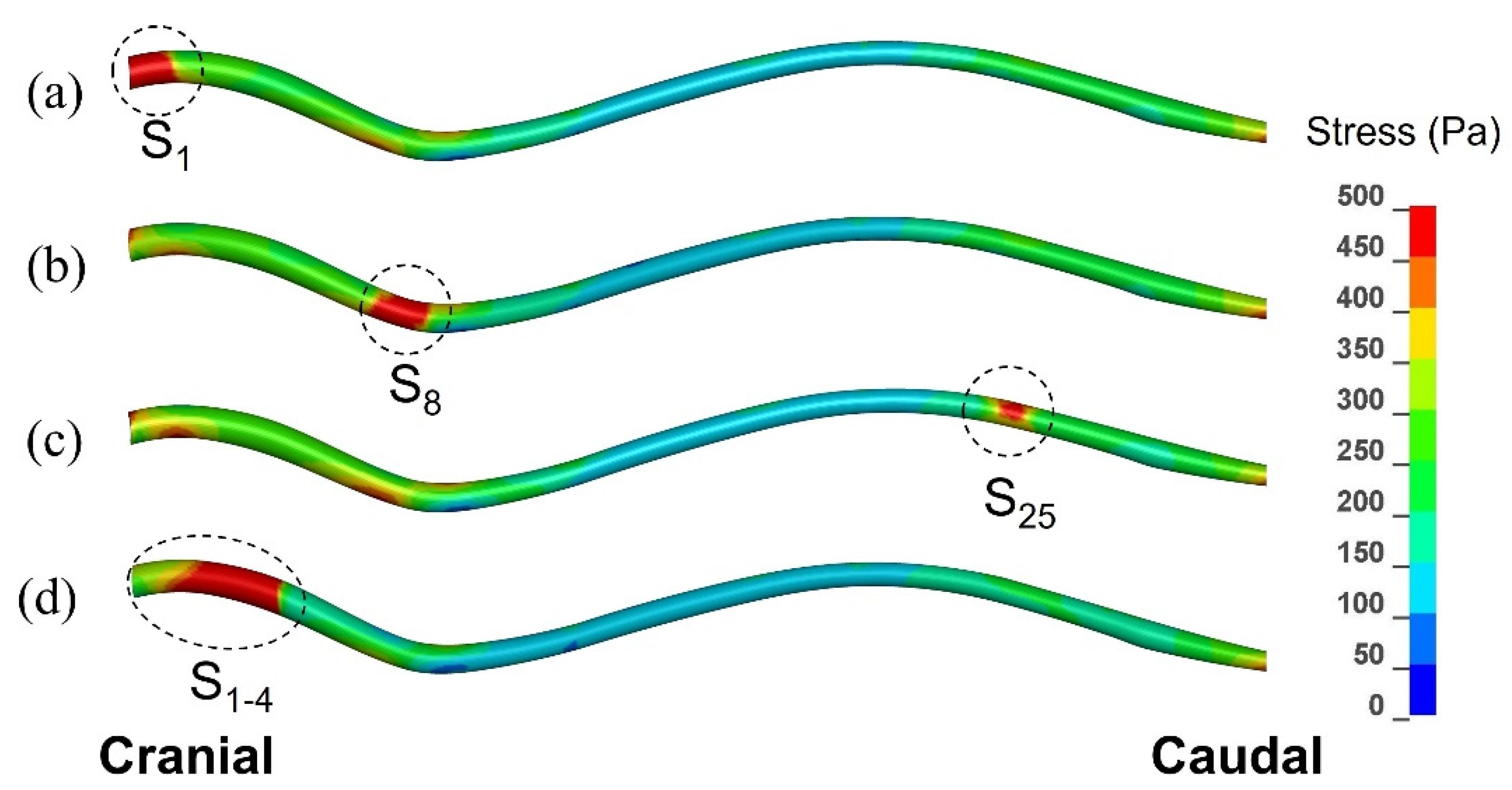
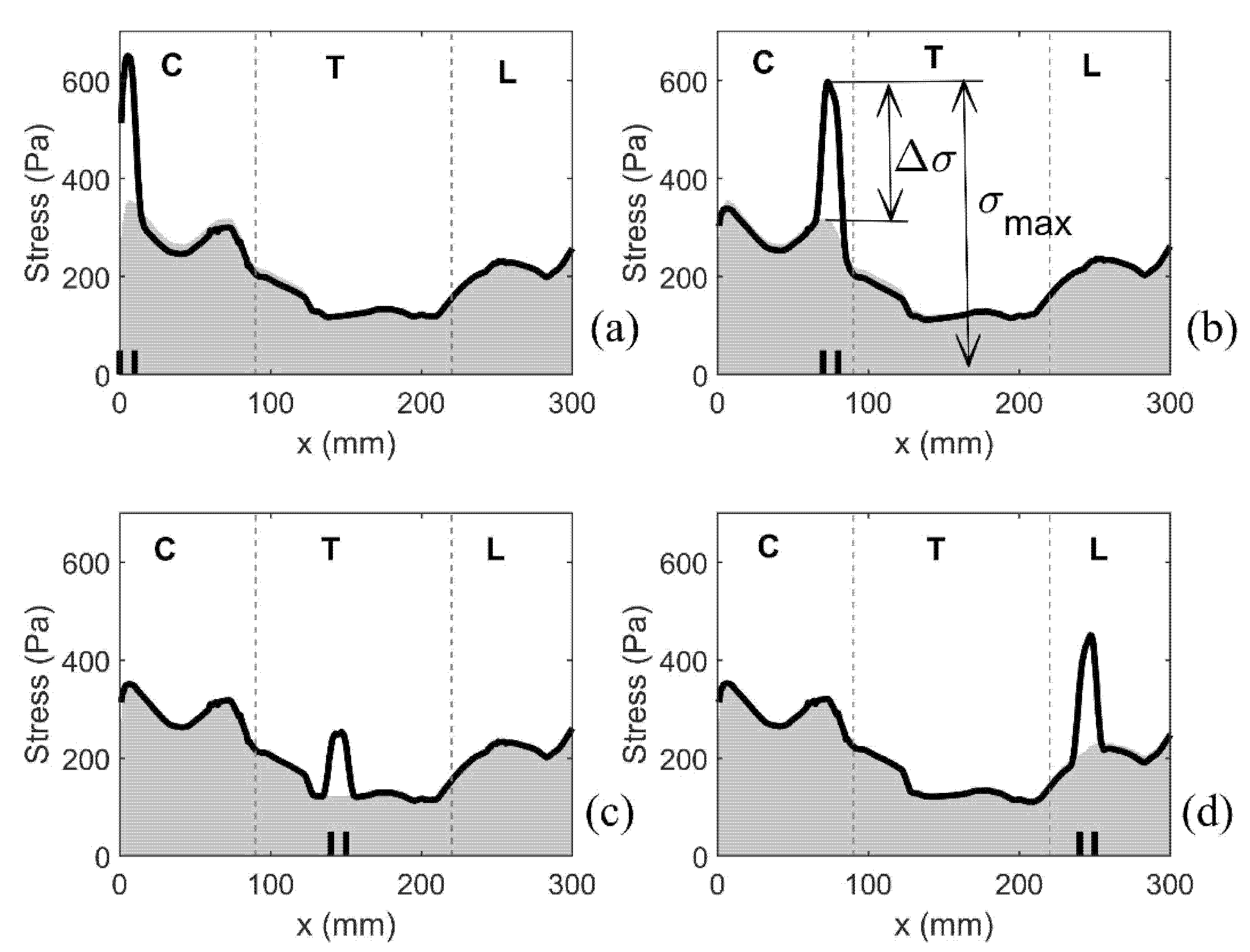
3.1.3. Patterns of the Mechanical Stress in the Spinal Cord
3.2. Simulation Results for Isolated Syringes and for the Expanding Syrinx
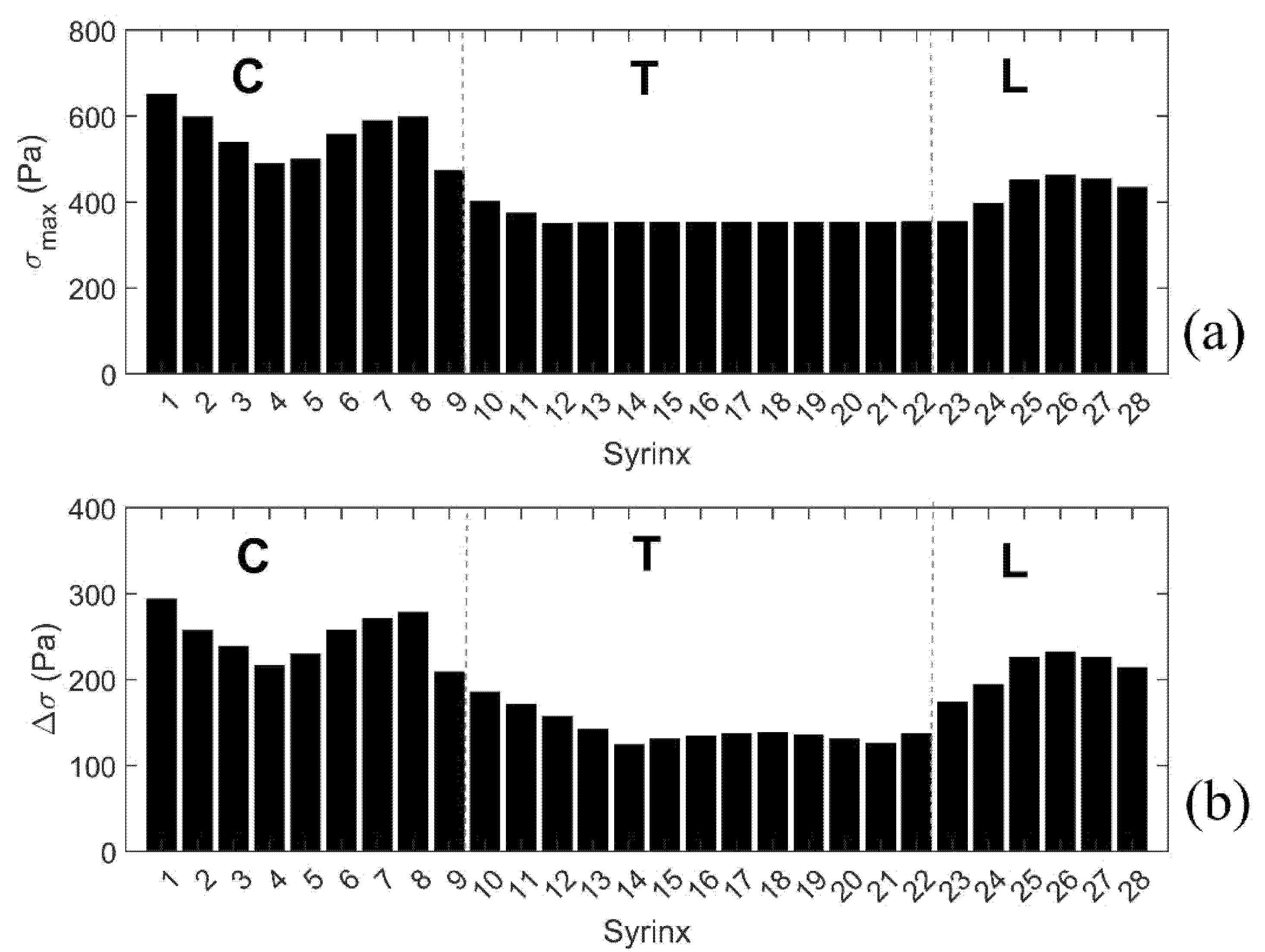
3.2.1. Small, Isolated Syringes
3.2.2. Expanding Syrinx
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, B. Post-traumatic syringomyelia, an update. Paraplegia 1990, 28, 296–313. [Google Scholar] [CrossRef] [Green Version]
- Heiss, J.D.; Patronas, N.; DeVroom, H.L.; Shawker, T.; Ennis, R.; Kammerer, W.; Eidsath, A.; Talbot, T.; Morris, J.; Eskioglu, E.; et al. Elucidating the pathophysiology of syringomyelia. J. Neurosurg. 1999, 91, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Sixt, C.; Riether, F.; Will, B.E.; Tatagiba, M.S.; Roser, F. Evaluation of quality of life parameters in patients who have syringomyelia. J. Clin. Neurosci. 2009, 16, 1599–1603. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.E.; Knowler, S.P.; Rusbridge, C.; Noorman, E.; Jeffery, N.D. Prevalence of asymptomatic syringomyelia in Cavalier King Charles spaniels. Vet. Rec. 2011, 168, 667. [Google Scholar] [CrossRef]
- Stoodley, M.A.; Jones, N.R.; Yang, L.; Brown, C.J. Mechanisms underlying the formation and enlargement of noncommunicating syringomyelia: Experimental studies. Neurosurg. Focus 2000, 8, E2. [Google Scholar] [CrossRef] [PubMed]
- Brodbelt, A.R.; Stoodley, M.A.; Watling, A.M.; Tu, J.; Jones, N.R. Fluid flow in an animal model of post-traumatic syringomyelia. Eur. Spine J. 2003, 12, 300–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodbelt, A.R.; Stoodley, M.A.; Watling, A.; Rogan, C.; Tu, J.; Brown, C.J.; Burke, S.; Jones, N.R. The role of excitotoxic injury in post-traumatic syringomyelia. J. Neurotrauma 2003, 20, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Rusbridge, C. Chiari-like malformation with syringomyelia in the Cavalier King Charles spaniel: Long-term outcome after surgical management. Vet. Surg. 2007, 36, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Heiss, J.D.; Suffredini, G.; Smith, R.; DeVroom, H.L.; Patronas, N.J.; Butman, J.A.; Thomas, F.; Oldfield, E.H. Pathophysiology of persistent syringomyelia after decompressive craniocervical surgery. Clinical article. J. Neurosurg. Spine 2010, 13, 729–742. [Google Scholar] [CrossRef] [PubMed]
- Levine, D.N. The pathogenesis of syringomyelia associated with lesions at the foramen magnum: A critical review of existing theories and proposal of a new hypothesis. J. Neurol. Sci. 2004, 220, 3–21. [Google Scholar] [CrossRef]
- Brodbelt, A.R.; Stoodley, M.A. Post-traumatic syringomyelia: A review. J. Clin. Neurosci. 2003, 10, 401–408. [Google Scholar] [CrossRef]
- Martin, B.A.; Reymond, P.; Novy, J.; Baledent, O.; Stergiopulos, N. A coupled hydrodynamic model of the cardiovascular and cerebrospinal fluid system. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H1492–H1509. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Cirovic, S. A computational model of the cerebrospinal fluid system incorporating lumped-parameter cranial compartment and one-dimensional distributed spinal compartment. J. Biorheol. 2011, 25, 78–87. [Google Scholar] [CrossRef]
- Hemley, S.J.; Biotech, B.; Tu, J.; Stoodley, M.A. Role of the blood-spinal cord-barrier in posttraumatic syringomyelia Laboratory investigation. J. Neurosurg.-Spine 2009, 11, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Heil, M.; Bertram, C.D. A poroelastic fluid-structure interaction model of syringomyelia. J. Fluid Mech. 2016, 809, 360–389. [Google Scholar] [CrossRef] [Green Version]
- Elliott, N.S.J.; Lucey, A.D.; Lockerby, D.A.; Brodbelt, A.R. Fluid-structure interactions in a cylindrical layered wave guide with application in the spinal column to syringomyelia. J. Fluid Struct. 2017, 70, 464–499. [Google Scholar] [CrossRef]
- Elliott, N.S.; Lockerby, D.A.; Brodbelt, A.R. A lumped-parameter model of the cerebrospinal system for investigating arterial-driven flow in posttraumatic syringomyelia. Med. Eng. Phys. 2011, 33, 874–882. [Google Scholar] [CrossRef]
- Elliott, N.S. Syrinx fluid transport: Modeling pressure-wave-induced flux across the spinal pial membrane. J. Biomech. Eng. 2012, 134, 031006. [Google Scholar] [CrossRef]
- Cirovic, S.; Kim, M. A one-dimensional model of the spinal cerebrospinal-fluid compartment. J. Biomech. Eng. 2012, 134, 021005. [Google Scholar] [CrossRef]
- Cirovic, S. A coaxial tube model of the cerebrospinal fluid pulse propagation in the spinal column. J. Biomech. Eng. 2009, 131, 021008. [Google Scholar] [CrossRef]
- Chang, H.S.; Nakagawa, H. Theoretical analysis of the pathophysiology of syringomyelia associated with adhesive arachnoiditis. J. Neurol. Neurosurg. Psychiatry 2004, 75, 754–757. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.S.; Nakagawa, H. Hypothesis on the pathophysiology of syringomyelia based on simulation of cerebrospinal fluid dynamics. J. Neurol. Neurosurg. Psychiatry 2003, 74, 344–347. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, P.W.; Berkouk, K.; Lucey, A.D. Pressure wave propagation in fluid-filled co-axial elastic tubes. Part 2: Mechanisms for the pathogenesis of syringomyelia. J. Biomech. Eng. 2003, 125, 857–863. [Google Scholar] [CrossRef]
- Cardillo, G.; Camporeale, C. Modeling fluid–structure interactions between cerebro-spinal fluid and the spinal cord. J. Fluid Struct. 2021, 102, 103251. [Google Scholar] [CrossRef]
- Bilston, L.E.; Stoodley, M.A.; Fletcher, D.F. The influence of the relative timing of arterial and subarachnoid space pulse waves on spinal perivascular cerebrospinal fluid flow as a possible factor in syrinx development. J. Neurosurg. 2010, 112, 808–813. [Google Scholar] [CrossRef]
- Bilston, L.E.; Fletcher, D.F.; Brodbelt, A.R.; Stoodley, M.A. Arterial pulsation-driven cerebrospinal fluid flow in the perivascular space: A computational model. Comput. Methods Biomech. Biomed. Eng. 2003, 6, 235–241. [Google Scholar] [CrossRef]
- Bertram, C.D.; Heil, M. A Poroelastic Fluid/Structure-Interaction Model of Cerebrospinal Fluid Dynamics in the Cord With Syringomyelia and Adjacent Subarachnoid-Space Stenosis. J. Biomech. Eng. 2017, 139. [Google Scholar] [CrossRef] [Green Version]
- Bertram, C.D.; Brodbelt, A.R.; Stoodley, M.A. The origins of syringomyelia: Numerical models of fluid/structure interactions in the spinal cord. J. Biomech. Eng.-T Asme. 2005, 127, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Bertram, C.D.; Bilston, L.E.; Stoodley, M.A. Tensile radial stress in the spinal cord related to arachnoiditis or tethering: A numerical model. Med. Biol. Eng. Comput. 2008, 46, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Bertram, C.D. Evaluation by fluid/structure-interaction spinal-cord simulation of the effects of subarachnoid-space stenosis on an adjacent syrinx. J. Biomech. Eng. 2010, 132, 061009. [Google Scholar] [CrossRef] [PubMed]
- Bertram, C.D. A numerical investigation of waves propagating in the spinal cord and subarachnoid space in the presence of a syrinx. J. Fluid. Struct. 2009, 25, 1189–1205. [Google Scholar] [CrossRef]
- Williams, B. On the pathogenesis of syringomyelia: A review. J. R. Soc. Med. 1980, 73, 798–806. [Google Scholar] [CrossRef]
- Williams, B. Simultaneous cerebral and spinal fluid pressure recordings. I. Technique, physiology, and normal results. Acta Neurochir. 1981, 58, 167–185. [Google Scholar] [CrossRef] [PubMed]
- Williams, B. Cerebrospinal fluid pressure changes in response to coughing. Brain 1976, 99, 331–346. [Google Scholar] [CrossRef]
- Williams, B. Pathogenesis of syringomyelia. Acta Neurochir. 1993, 123, 159–165. [Google Scholar] [CrossRef]
- Cirovic, S.; Lloyd, R.; Jovanovik, J.; Volk, H.A.; Rusbridge, C. Computer simulation of syringomyelia in dogs. BMC Vet. Res. 2018, 14, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusbridge, C.; Stringer, F.; Knowler, S.P. Clinical application of diagnostic imaging of Chiari-like malformation and syringomyelia. Front. Vet. Sci. 2018, 5, 280. [Google Scholar] [CrossRef] [PubMed]
- Loderstedt, S.; Benigni, L.; Chandler, K.; Cardwell, J.M.; Rusbridge, C.; Lamb, C.R.; Volk, H.A. Distribution of syringomyelia along the entire spinal cord in clinically affected Cavalier King Charles Spaniels. Vet. J. 2011, 190, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Hallquist, J.O. LS-DYNA Theory Manual; Livermote Software Technology Corporation: Livermore, CA, USA, 2006. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spinal Cord | E = 62.5 kPa, ν = 0.49, ρ = 1000 kg/m3 |
| Dura | E = 1.25 MPa, ν = 0.4, ρ = 1000 kg/m3 |
| Epidural fat | E = 1 kPa, ν = 0.4999, ρ = 900 kg/m3 |
| CSF and fluid in the syrinx | μ = 0.001 Pa s, ρ = 1000 kg/m3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cirovic, S.; Rusbridge, C. Slosh Simulation in a Computer Model of Canine Syringomyelia. Life 2021, 11, 1083. https://doi.org/10.3390/life11101083
Cirovic S, Rusbridge C. Slosh Simulation in a Computer Model of Canine Syringomyelia. Life. 2021; 11(10):1083. https://doi.org/10.3390/life11101083
Chicago/Turabian StyleCirovic, Srdjan, and Clare Rusbridge. 2021. "Slosh Simulation in a Computer Model of Canine Syringomyelia" Life 11, no. 10: 1083. https://doi.org/10.3390/life11101083
APA StyleCirovic, S., & Rusbridge, C. (2021). Slosh Simulation in a Computer Model of Canine Syringomyelia. Life, 11(10), 1083. https://doi.org/10.3390/life11101083