
Use of NanoSIMS to Identify the Lower Limits of Metabolic Activity and Growth by Serratia liquefaciens Exposed to Sub-Zero Temperatures


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Enrichment Protocols
2.2. Cell Enumeration via MPN
2.3. Isotope Labeling Experiments
2.4. Scanning Electron Microscopy (SEM)
2.5. NanoSIMS Protocols
2.6. Statistics
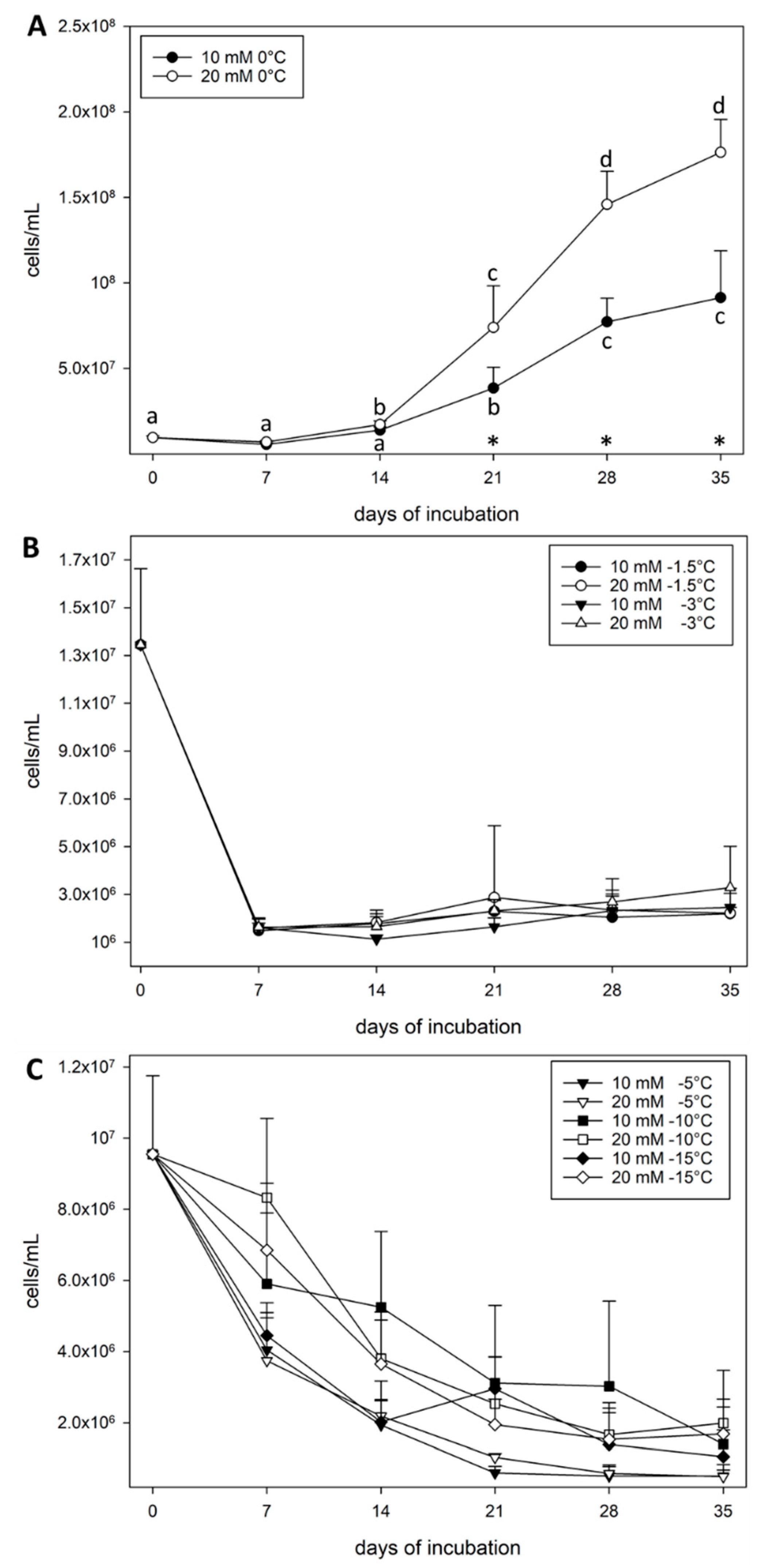
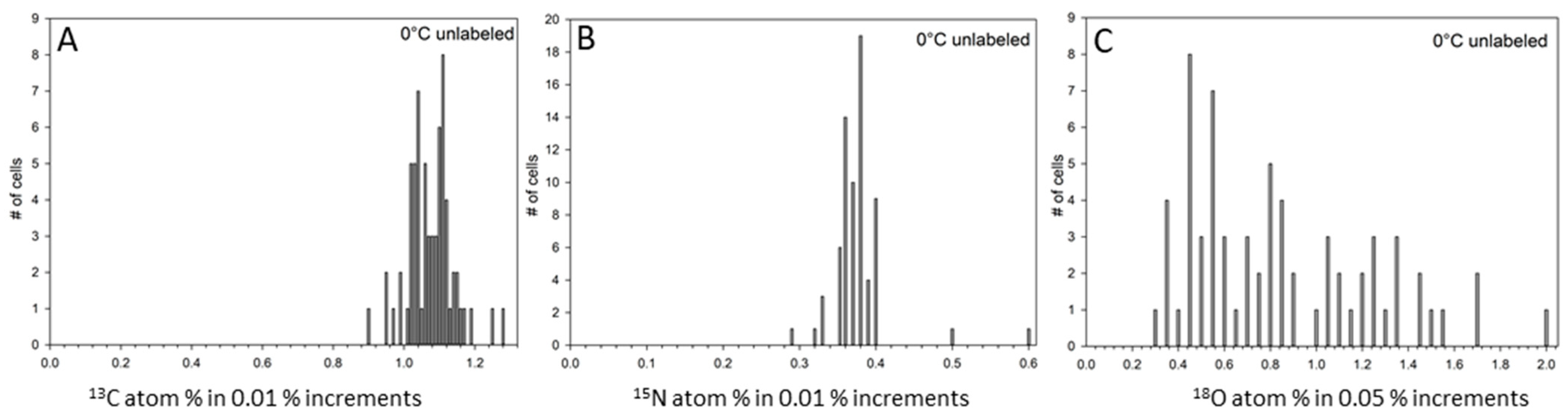
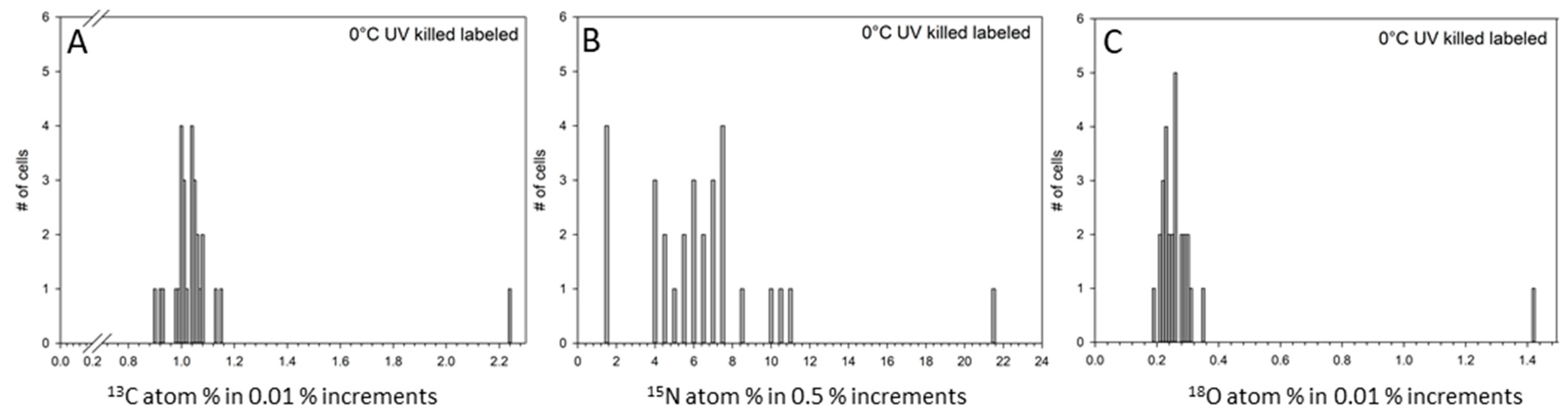
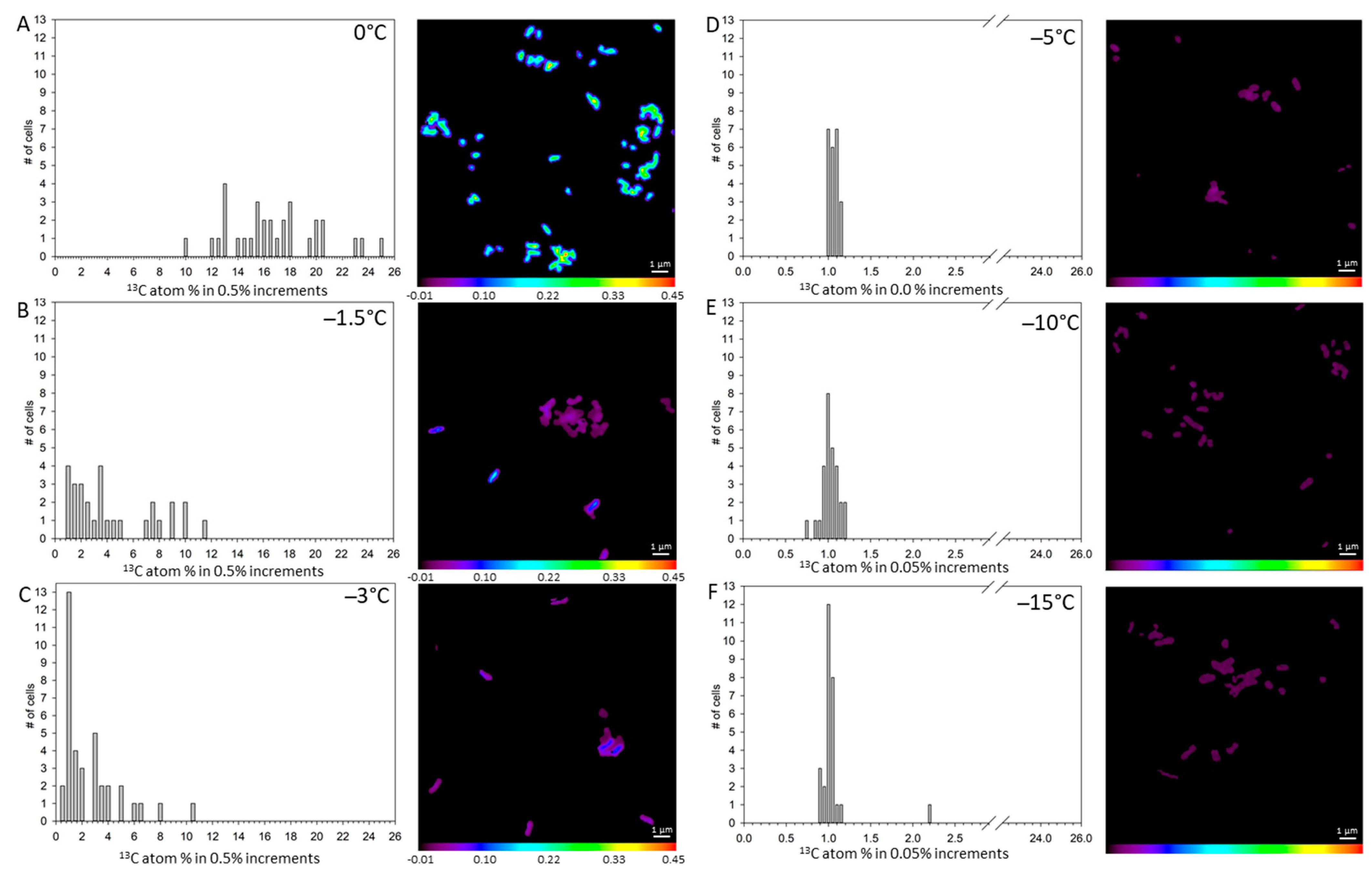
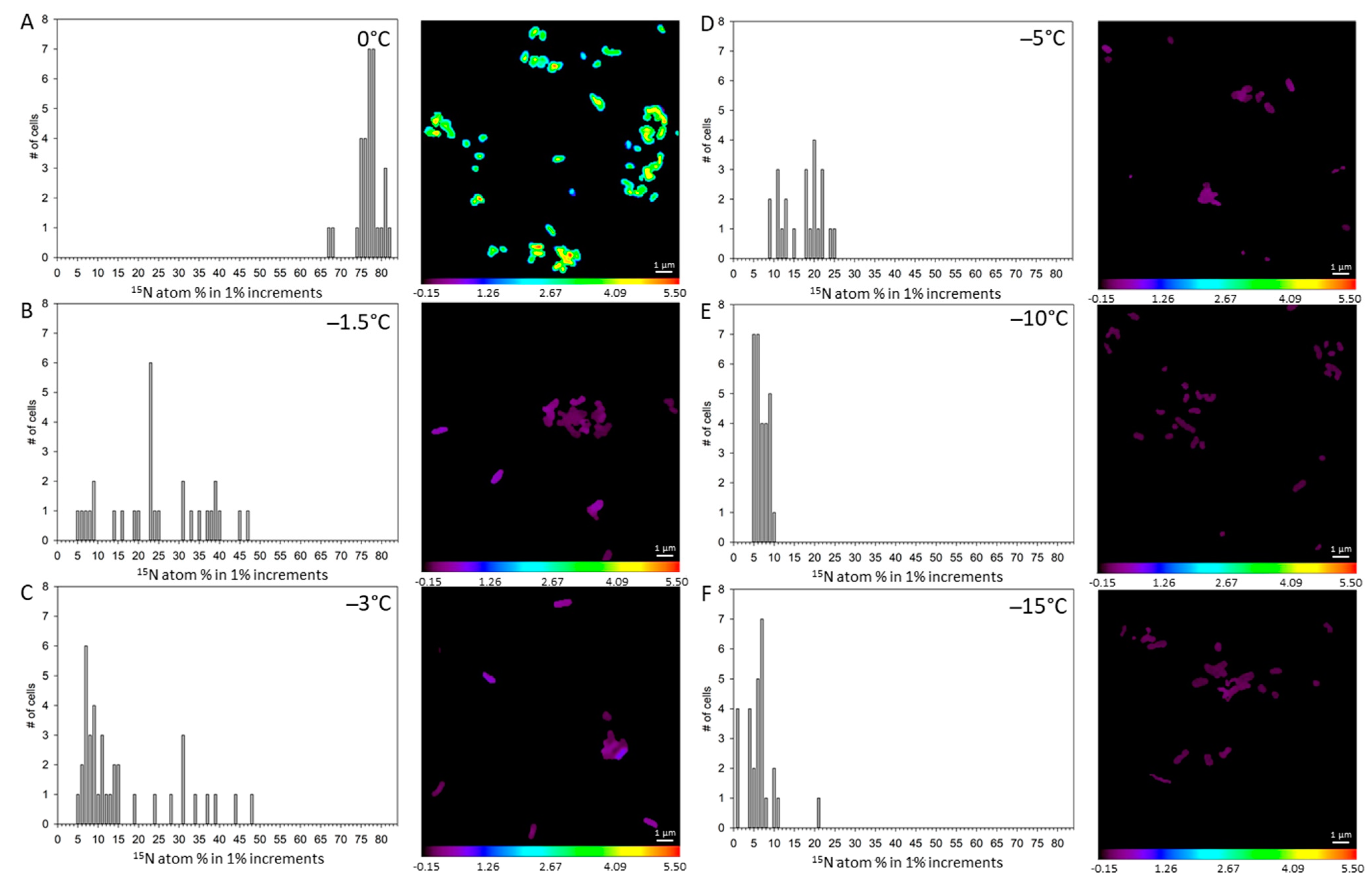
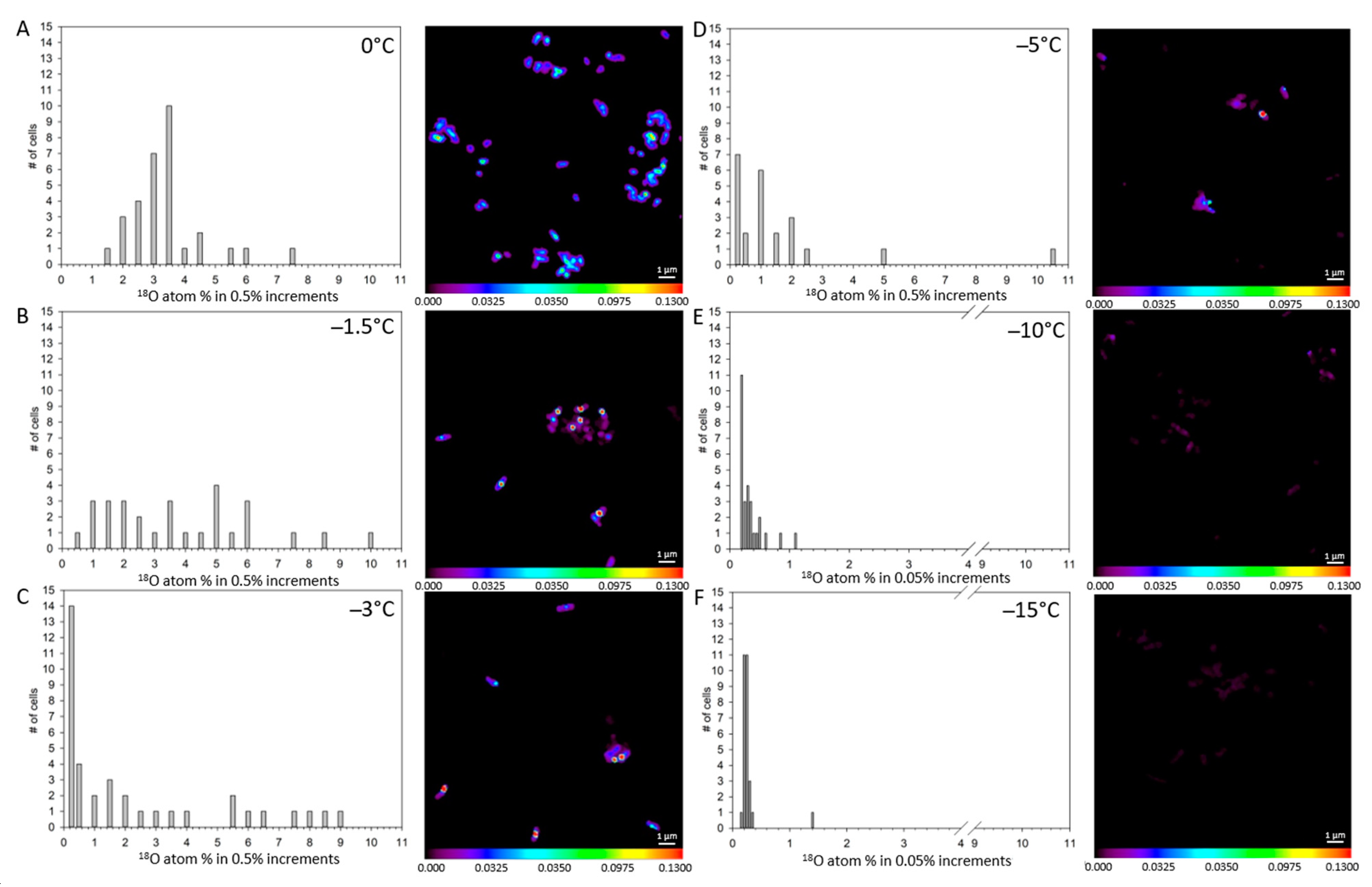
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Price, P.B.; Showers, T. Temperature dependence of metabolic rates for microbial growth, maintenance, and survival. Proc. Natl. Acad. Sci. USA 2004, 101, 4631–4636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, A.E.; Fritsen, C.H. Microbiota within the perennial ice cover of Lake Vida, Antarctica. FEMS Microbiol. Ecol. 2007, 59, 274–288. [Google Scholar] [CrossRef] [Green Version]
- Cockell, C.S.; Bush, T.; Bryce, C.; Direito, S.; Fox-Powell, M.; Harrison, J.P.; Lammer, H.; Landenmark, H.; Martin-Torres, J.; Nicholson, N.; et al. Habitability: A Review. Astrobiology 2016, 16, 89–117. [Google Scholar] [CrossRef]
- Rivkina, E.M.; Friedmann, E.I.; McKay, C.P.; Gilichinsky, D.A. Metabolic Activity of Permafrost Bacteria below the Freezing Point. Appl. Environ. Microbiol. 2000, 66, 3230–3233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christner, B.C.; Priscu, J.C.; Achberger, A.M.; Barbante, C.; Carter, S.P.; Christianson, K.; Michaud, A.B.; Mikucki, J.A.; Mitchell, A.C.; Skidmore, M.L.; et al. A microbial ecosystem beneath the West Antarctic ice sheet. Nature 2014, 512, 310–313. [Google Scholar] [CrossRef]
- Mikucki, J.A.; Auken, E.; Tulaczyk, D.; Virginia, R.A.; Schamper, C.; Sørensen, K.I.; Doran, P.T.; Dugan, H.; Foley, N. Deep groundwater and potential subsurface habitats beneath an Antarctic dry valley. Nat. Commun. 2015, 6, 6831. [Google Scholar] [CrossRef] [Green Version]
- Nersesova, Z.A.; Tsytovich, N.A. Unfrozen water in frozen soils, In Permafrost: Proceedings of International Conference; National Academy of Sciences: Washington, DC, USA, 1966; pp. 230–234. [Google Scholar]
- Bore, E.K.; Apostel, C.; Halicki, S.; Kuzyakov, Y.; Dippold, M.A. Microbial metabolism in soil at subzero temperatures: Adaptation mechanisms revealed by position-specific 13C labeling. Front. Microbiol. 2017, 8, 946. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, A.; Burkhardt, J.; Cockell, C.S.; Cray, J.A.; Dijksterhuis, J.; Fox-Powell, M.; Kee, T.P.; Kminek, G.; McGenity, T.J.; Timmis, K.N.; et al. Multiplication of microbes below 0.690 water activity: Implications for terrestrial and extraterrestrial life. Environ. Microbiol. 2015, 17, 257–277. [Google Scholar] [CrossRef]
- Hoehler, T.M.; Jorgensen, B.B. Microbial life under extreme energy limitation. Nat. Rev. Microbiol. 2013, 11, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Lorv, J.S.H.; Rose, D.R.; Glick, B.R. Bacterial ice crystal controlling proteins. Scientifica 2014, 20. [Google Scholar] [CrossRef] [Green Version]
- Buffington, B. Trajectory design for the Europa Clipper mission concept. Aiaa/Aas Astrodyn. Spec. Conf. 2014. [Google Scholar] [CrossRef]
- Clark, K.; Boldt, J.; Greeley, R.; Hand, K.; Jun, I.; Lock, R.; Pappalardo, R.; van Houten, T.; Yan, T. Return to Europa: Overview of the Jupiter Europa orbiter mission. Adv. Space Res. 2011, 48, 629–650. [Google Scholar] [CrossRef]
- Rummel, J.D.; Beaty, D.W.; Jones, M.A.; Bakermans, C.; Barlow, N.G.; Boston, P.J.; Chevrier, V.F.; Clark, B.C.; de Vera, J.P.; Gough, R.V.; et al. A new analysis of Mars “Special Regions”: Findings of the second MEPAG Special Regions Science Analysis Group (SR-SAG2). Astrobiology 2014, 14, 887–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piqueux, S.; Buz, J.; Edwards, C.S.; Bandfield, J.L.; Kleinböhl, A.; Kass, D.M.; Hayne, P.O.; The MCS THEMIS Teams. Widespread shallow water ice on Mars at high latitudes and mid latitudes. Geophys. Res. Lett. 2019, 46, 14290–14298. [Google Scholar] [CrossRef]
- Zent, A.P. A historical search for habitable ice at the Phoenix landing site. Icarus 2008, 196, 385–408. [Google Scholar] [CrossRef]
- Stoker, C.R.; Zent, A.; Catling, D.C.; Douglas, S.; Marshall, J.R.; Archer Jr., D.; Clark, B.; Kounaves, S.P.; Lemmon, M.T.; Quinn, R.; et al. Habitability of the Phoenix landing site. Geophys. Res. 2010, 115, E00E20. [Google Scholar] [CrossRef]
- Madeleine, J.B.; Forget, F.; Head, J.W.; Levrard, B.; Montmessin, F.; Millour, E. Amazonian northern mid-latitude glaciation on Mars: A proposed climate scenario. Icarus 2009, 203, 390–405. [Google Scholar] [CrossRef] [Green Version]
- Schuerger, A.C.; Ulrich, R.; Berry, B.J.; Nicholson, W.L. Growth of Serratia liquefaciens under 7 mbar, 0 °C, and CO2-enriched anoxic atmospheres. Astrobiology 2013, 13, 115–131. [Google Scholar] [CrossRef] [Green Version]
- Schuerger, A.C.; Mancinelli, R.L.; Kern, R.G.; Rothschild, L.J.; McKay, C.P. Survival of endospores of Bacillus subtilis on spacecraft surfaces under simulated Martian environments: Implications for the forward contamination of Mars. Icarus 2003, 165, 253–276. [Google Scholar] [CrossRef]
- Schuerger, A.C.; Nicholson, W.L. Interactive effects of hypobaria, low temperature, and CO2 atmospheres inhibit the growth of mesophilic Bacillus spp. Under simulated martian conditions. Icarus 2006, 185, 143–152. [Google Scholar] [CrossRef]
- Schwendner, P.; Schuerger, A.C. Exploring microbial activity in low-pressure environments. Curr. Issues Mol. Biol. 2020, 38, 163–196. [Google Scholar] [CrossRef] [PubMed]
- Youle, M.; Rohwer, F.; Stacy, A.; Whiteley, M.; Steel, B.C.; Delalez, N.J.; Nord, A.L.; Berry, R.M.; Armitage, J.P.; Kamoun, S.; et al. The microbial Olympics. Nat. Rev. Microbiol. 2012, 10, 583–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakermans, C.; Skidmore, M. Microbial respiration in ice at subzero temperatures (−4 °C to −33 °C). Environ. Micobiol. Rep. 2011, 3, 774–782. [Google Scholar] [CrossRef]
- Panikov, N.S.; Flanagan, P.W.; Oechel, W.C.; Mastepanov, M.A.; Christensen, T.R. Microbial activity in soils frozen to below −39 °C. Soil Biol. Biochem. 2006, 38, 785–794. [Google Scholar] [CrossRef]
- Schaefer, K.; Jafarov, E. A parameterization of respiration in frozen soils based on substrate availability. Biogeosciences 2016, 13, 1991–2001. [Google Scholar] [CrossRef] [Green Version]
- La Duc, M.T.; Dekas, A.; Osman, S.; Moissl, C.; Newcombe, D.; Venkateswaran, K. Isolation and characterization of bacteria capable of tolerating the extreme conditions of clean room environments. Appl. Environ. Microbiol. 2007, 73, 2600–2611. [Google Scholar] [CrossRef] [Green Version]
- Moissl-Eichinger, C.; Pukall, R.; Probst, A.J.; Stieglmeier, M.; Schwendner, P.; Mora, M.; Barczyk, S.; Bohmeier, M.; Rettberg, P. Lessons learned from the microbial analysis of the Herschel spacecraft during assembly, integration, and test operations. Astrobiology 2013, 13, 1125–1139. [Google Scholar] [CrossRef] [PubMed]
- Vaishampayan, P.; Probst, A.J.; La Duc, M.T.; Bargoma, E.; Benardini, J.N.; Andersen, G.L.; Venkateswaran, K. New perspectives on viable microbial communities in low-biomass cleanroom environments. ISME J. 2013, 7, 312–324. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, M.K. Mechanism of bacterial adaptation to low temperature. J. Biosci. 2006, 31, 157–165. [Google Scholar] [CrossRef]
- D’Amico, S.; Collins, T.; Marx, J.C.; Feller, G.; Gerday, C. Psychrophilic microorganisms: Challenges for life. Embo Rep. 2006, 7, 385–389. [Google Scholar] [CrossRef]
- Gilichinsky, D.; Rivkina, E.; Shcherbakova, V.; Laurinavichuis, K.; Tiedje, J. Supercooled water brines within permafrost—an unknown ecological niche for microorganisms: A model for astrobiology. Astrobiology 2003, 3, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Musat, N.; Foster, R.; Vagner, T.; Adam, B.; Kuypers, M.M.M. Detecting metabolic activities in single cells, with emphasis on nanoSIMS. FEMS Microbiol. Rev. 2012, 36, 486–511. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, A.M.; Clode, P.L.; Fletcher, I.A.; Nunan, N.; Stockdale, E.A.; O’Donnell, A.G.; Murphy, D.V. A novel method for the study of the biophysical interface in soils using nano-scale secondary ion mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Eichorst, S.; Strasser, F.; Woyke, T.; Schintlmeister, A.; Wagner, M.; Woebken, D. Advancements in the application of NanoSIMS and Raman microspectroscopy to investigate the activity of microbial cells in soils. FEMS Microbiol. Ecol. 2015, 91, fiv106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spizizen, J. Transformation of biochemically deficient strains of Bacillus subtilis by deoxyribonucleate. Proc. Natl. Acad. Sci. USA 1958, 44, 1072–1078. [Google Scholar] [CrossRef] [Green Version]
- Rettedal, E.A.; Brözel, V.S. Characterizing the diversity of active bacteria in soil by comprehensive stable isotope probing of DNA and RNA with H218O. Microbiol. Open 2015, 4, 208–219. [Google Scholar] [CrossRef]
- Jakosky, B.M.; Nealson, K.H.; Bakermans, C.; Ley, R.E.; Mellon, M.T. Subfreezing activity of microorganisms and the potential habitability of Mars’ Polar Regions. Astrobiology 2003, 3, 343–350. [Google Scholar] [CrossRef]
- Popa, R.; Weber, P.K.; Pett-Ridge, J.; Finzi, J.A.; Fallon, S.J.; Hutcheon, I.D.; Nealson, K.H.; Capone, D.G. Carbon and nitrogen fixation and metabolite exchange in and between individual cells of Anabaena oscillarioides. ISME J. 2007, 1, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Lechene, C.P.; Luyten, Y.; McMahon, G.; Distel, D.L. Quantitative imaging of nitrogen fixation by individual bacteria within animal cells. Science 2007, 317, 1563–1566. [Google Scholar] [CrossRef] [Green Version]
- Musat, N.; Halm, H.; Winterholler, B.; Hoppe, P.; Peduzzi, S.; Hillion, F.; Horreard, F.; Amann, R.; Jørgensen, B.B.; Kuypers, M.M.M. A single-cell view on the ecophysiology of anaerobic phototrophic bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 17861–17866. [Google Scholar] [CrossRef] [Green Version]
- Behrens, S.; Lösekann, T.; Pett-Ridge, J.; Weber, P.K.; Ng, W.-O.; Stevenson, B.S.; Hutcheon, I.D.; Relmann, D.A.; Spormann, A.M. Linking microbial phylogeny to metabolic activity at the single-cell level by using enhanced element labeling-catalyzed reporter deposition fluorescence in situ hybridization (EL-FISH) and NanoSIMS. Appl. Environ. Microbiol. 2008, 74, 3143–3150. [Google Scholar] [CrossRef] [Green Version]
- Finzi-Hart, J.A.; Pett-Ridge, J.; Weber, P.K.; Popa, R.; Fallon, S.J.; Gunderson, T.; Hutcheon, I.D.; Nealson, K.H.; Capone, D.G. Fixation and fate of C and N in the cyanobacterium Trichodesmium using nanometer-scale secondary ion mass spectrometry. Proc. Natl. Acad. Sci. USA 2009, 106, 6345–6350. [Google Scholar] [CrossRef] [Green Version]
- Summons, R.E.; Franzmann, P.D.; Nichols, P.D. Carbon Isotopic fractionation associated with methylotrophic methanogenesis. Org. Geochem. 1998, 28, 465–475. [Google Scholar] [CrossRef]
- Dekas, A.E.; Poretsky, R.S.; Orphan, V.J. Deep-Sea Archaea Fix and Share Nitrogen in Methane-consuming Microbial Consortia. Science 2009, 326, 422–426. [Google Scholar] [CrossRef] [Green Version]
- Orphan, V.J.; Turk, K.A.; Green, A.M.; House, C.H. Patterns of 15N assimilation and growth of methanotrophic ANME-2 archaea and sulfate-reducing bacteria within structured syntrophic consortia revealed by FISH-SIMS. Environ. Microbiol. 2009, 11, 1777–1791. [Google Scholar] [CrossRef] [PubMed]
- Cliff, J.B.; Gaspar, D.J.; Bottomley, P.J.; Myrold, D.D. Exploration of inorganic C and N assimilation by soil microbes with time-of-flight secondary ion mass spectrometry. Appl. Environ. Microbiol. 2002, 68, 4067–4073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cliff, J.B.; Bottomley, P.J.; Gaspar, D.J.; Myrold, D.D. Nitrogen mineralization and assimilation at millimeter scales. Soil Biol. Biochem. 2007, 39, 823–826. [Google Scholar] [CrossRef]
- Schwartz, E. Analyzing microorganisms in environmental samples using stable isotope probing with H218O. Cold Spring Harb. Protoc. 2009, 77. [Google Scholar] [CrossRef] [PubMed]
- Papp, K.; Mau, R.L.; Hayer, M.; Koch, B.J.; Hungate, B.A.; Schwartz, E. Quantitative stable isotope probing with H218O reveals that most bacterial taxa in soil synthesize new ribosomal RNA. ISME J. 2018, 12, 3043–3045. [Google Scholar] [CrossRef]
- Woods, A.; Watwood, M.; Schwartz, E. Identification of a toluene-degrading bacterium from a soil sample through H218O DNA stable Isotope probing. Appl. Environ. Microbiol. 2011, 77, 5995–5999. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, E. Characterization of growing microorganisms in soil by stable isotope probing with H218O. Appl. Environ. Microbiol. 2007, 73, 2541–2546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langer, S.; Vogts, A.; Schulz-Vogt, N. Simultaneous Visualization of Enzymatic Activity in the Cytoplasm and at Polyphosphate Inclusions in Beggiatoa sp. Strain 35Flor Incubated with 18O-Labeled Water. mSphere 2018, 3, e00489–e18. [Google Scholar] [CrossRef] [Green Version]
- Perfumo, A.; Elsaesser, A.; Littmann, S.; Foster, R.A.; Kyupers, M.M.M.; Cockell, C.S.; Kminek, G. Epifluorescence, SEM, TEM and nanoSIMS image analysis of the cold phenotype of Clostridium psychrophilum at subzero temperatures. FEMS Microbiol. Ecol. 2014, 90, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Raymond-Bouchard, I.; Chourey, K.; Altshuler, I.; Iyer, R.; Hettich, R.L.; Whyte, L.G. Mechanisms of subzero growth in the cryophile Planococcus halocryophilus determined through proteomic analysis. Environ. Microbiol. 2017, 19, 4460–4479. [Google Scholar] [CrossRef] [PubMed]
- Bakermans, C.; Tsapin, A.I.; Souza-Egipsy, V.; Gilichinsky, D.A.; Nealson, K.H. Reproduction and metabolism at −10 °C of bacteria isolated from Siberian permafrost. Environ. Microbiol. 2003, 5, 321–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peteranderl, R.; Lechene, C. Measure of carbon and nitrogen stable isotope ratios in cultures cells. J. Am. Soc. Mass Spectrom. 2004, 15, 478–485. [Google Scholar] [CrossRef] [Green Version]
- Musat, N.; Stryhanyuk, H.; Bombach, P.; Adrian, L.; Audinot, J.-N.; Richnow, H.H. The effect of FISH and CARD-FISH on the isotopic composition of 13C- and 15N-labeled Pseudomonas putida cells measured by nanoSIMS. Syst. Appl. Microbiol. 2014, 37, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Richards, O.C.; Boyer, P.D. 18O labeling of DNA during synthesis and stability of label during replication. J. Mol. Biol. 1966, 19, 109–119. [Google Scholar] [CrossRef]
- Kempes, C.P.; van Bodegom, P.M.; Wolpert, D.; Libby, D.; Amend, J.; Hoehler, T. Drivers of bacterial maintenance and minimal energy requirements. Front. Microbiol. 2017, 8, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, P.B. A habitat for psychrophiles in deep Antarctic ice. Proc. Natl. Acad. Sci. USA 2000, 97, 1247–1251. [Google Scholar] [CrossRef] [Green Version]
- Tung, H.C.; Price, P.B.; Bramall, N.E.; Vrdoljak, G. Microorganisms metabolizing on clay grains in 3-km-deep Greenland basal ice. Astrobiology 2006, 6, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Rohde, R.A.; Price, P.B. Diffusion-controlled metabolism for long-term survival of single isolated microorganisms trapped within ice crystals. Proc. Natl. Acad. Sci. USA 2007, 104, 16592–16597. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viable Cells per mL | T0 | T70 | ||||
|---|---|---|---|---|---|---|
| −1.5 °C | −3 °C | −5 °C | −10 °C | −15 °C | ||
| 10 mM | 7.2 × 107 | 6.5 × 107 | 3.2 × 107 | 5.8 × 104 | 1.2 × 105 | 1.1 × 105 |
| ±1.7 × 107 | ±3.0 × 107 | ±1.7 × 107 | ±1.7 × 104 | ±1.8 × 104 | ±4.2 × 104 | |
| 20 mM | 7.2 × 107 | 9.4 × 107 | 4.3 × 107 | 7.2 × 104 | 2.5 × 105 | 1.5 × 105 |
| ±1.7 × 107 | ±4.0 × 107 | ±1.7 × 107 | ±4.4 × 102 | ±1.7 × 103 | ±1.7 × 103 | |
| Treatment | 13C atom-% | Std Error | 15N atom-% | Std Error | 18O atom-% | Std Error |
|---|---|---|---|---|---|---|
| 0 °C unlabeled (n = 67) | 1.08 d | 0.008 | 0.38 e | 0.005 | 0.87 c | 0.050 |
| 0 °C UV-killed (n = 33) | 1.04 e | 0.0034 | 17.49 c | 0.303 | 0.63 d | 0.013 |
| 0 °C labeled (n = 31) | 16.93 a | 0.635 | 77.28 a | 0.674 | 3.63 a | 0.217 |
| −1.5 °C labeled (n = 29) | 4.70 b | 0.601 | 25.08 b | 2.429 | 4.06 a | 0.424 |
| −3 °C labeled (n = 37) | 2.83 c | 0.373 | 16.97 c | 1.904 | 2.47 b | 0.472 |
| −5 °C labeled (n = 23) | 1.08 d | 0.010 | 17.66 c | 0.908 | 1.75 bc | 0.535 |
| −10 °C labeled (n = 28) | 1.05 de | 0.018 | 7.51 d | 0.358 | 0.37 e | 0.041 |
| −15 °C labeled (n = 28) | 1.07 de | 0.045 | 6.75 d | 0.746 | 0.30 e | 0.041 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwendner, P.; Nguyen, A.N.; Schuerger, A.C. Use of NanoSIMS to Identify the Lower Limits of Metabolic Activity and Growth by Serratia liquefaciens Exposed to Sub-Zero Temperatures. Life 2021, 11, 459. https://doi.org/10.3390/life11050459
Schwendner P, Nguyen AN, Schuerger AC. Use of NanoSIMS to Identify the Lower Limits of Metabolic Activity and Growth by Serratia liquefaciens Exposed to Sub-Zero Temperatures. Life. 2021; 11(5):459. https://doi.org/10.3390/life11050459
Chicago/Turabian StyleSchwendner, Petra, Ann N. Nguyen, and Andrew C. Schuerger. 2021. "Use of NanoSIMS to Identify the Lower Limits of Metabolic Activity and Growth by Serratia liquefaciens Exposed to Sub-Zero Temperatures" Life 11, no. 5: 459. https://doi.org/10.3390/life11050459
APA StyleSchwendner, P., Nguyen, A. N., & Schuerger, A. C. (2021). Use of NanoSIMS to Identify the Lower Limits of Metabolic Activity and Growth by Serratia liquefaciens Exposed to Sub-Zero Temperatures. Life, 11(5), 459. https://doi.org/10.3390/life11050459