
Altered Blood Levels of Anti-Gal Antibodies in Alzheimer’s Disease: A New Clue to Pathogenesis?

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Healthy Controls
2.2. Blood Collection and Processing
2.3. Determination of α-Gal Antibody Titers
2.4. Statistical Analysis
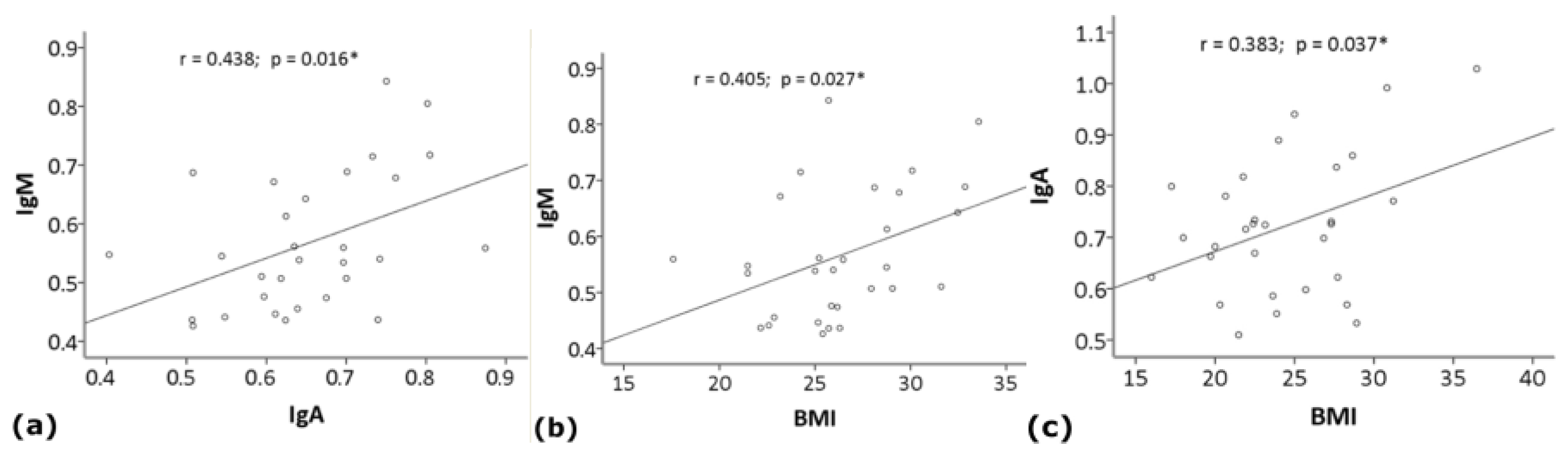
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alzheimer’s Association. 2020 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2020, 16, 391–460. [Google Scholar] [CrossRef]
- Fleming, R.; Zeisel, J.; Bennet, K. World Alzheimer Report 2020, Vol I and II; Alzheimer’s Disease International: London, UK, 2020. [Google Scholar]
- McGrowder, D.A.; Miller, F.; Vaz, K.; Nwokocha, C.; Wilson-Clarke, C.; Anderson-Cross, M.; Brown, J.; Anderson-Jackson, L.; Williams, L.; Latore, L.; et al. Cerebrospinal Fluid Biomarkers of Alzheimer’s Disease: Current Evidence and Future Perspectives. Brain Sci. 2021, 11, 215. [Google Scholar] [CrossRef]
- González-Sanmiguel, J.; Schuh, C.M.A.P.; Muñoz-Montesino, C.; Contreras-Kallens, P.; Aguayo, L.G.; Aguayo, S. Complex Interaction between Resident Microbiota and Misfolded Proteins: Role in Neuroinflammation and Neurodegeneration. Cells 2020, 9, 2476. [Google Scholar] [CrossRef]
- Almeida, I.C.; Ferguson, M.A.; Schenkman, S.; Travassos, L.R. Lytic anti-alpha-galactosyl antibodies from patients with chronic Chagas’ disease recognize novel O-linked oligosaccharides on mucin-like glycosyl-phosphatidylinositol-anchored glycoproteins of Trypanosoma cruzi. Biochem. J. 1994, 304, 793. [Google Scholar] [CrossRef] [PubMed]
- Welsh, R.M.; O’Donnell, C.L.; Reed, D.J.; Rother, R.P. Evaluation of the Galalpha1-3Gal epitope as a host modification factor eliciting natural humoral immunity to enveloped viruses. J. Virol. 1998, 72, 4650–4656. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Cai, L.; Wu, B.; Li, L.; Xiao, Z.; Cheng, J.; Wang, P.G. The wciN gene encodes an α-1,3-galactosyltransferase involved in the biosynthesis of the capsule repeating unit of Streptococcus pneumoniae serotype 6B. Biochemistry 2012, 51, 5804–5810. [Google Scholar] [CrossRef] [Green Version]
- Huai, G.; Qi, P.; Yang, H.; Wang, Y. Characteristics of α-Gal epitope, anti-Gal antibody, α1,3 galactosyltransferase and its clinical exploitation. Int. J. Mol. Med. 2016, 37, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyneveld, G.I.; Savelkoul, H.F.J.; Parmentier, H.K. Current Understanding of Natural Antibodies and Exploring the Possibilities of Modulation Using Veterinary Models. A Review. Front. Immunol. 2020, 11, 2139. [Google Scholar] [CrossRef]
- Galili, U. Discovery of the natural anti-Gal antibody and its past and future relevance to medicine. Xenotransplantation 2013, 20, 138–147. [Google Scholar] [CrossRef]
- Galili, U. Human Natural Antibodies to Mammalian Carbohydrate Antigens as Unsung Heroes Protecting against Past, Present, and Future Viral Infections. Antibodies 2020, 9, 25. [Google Scholar] [CrossRef]
- Galili, U. Significance of the evolutionary α1,3-galactosyltransferase (GGTA1) gene inactivation in preventing extinction of apes and old world monkeys. J. Mol. Evol. 2015, 80, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.G.; McKelvey, K.J.; March, L.M.; Hunter, D.J.; Xue, M.; Jackson, C.J.; Morris, J.M. Aberrant levels of natural IgM antibodies in osteoarthritis and rheumatoid arthritis patients in comparison to healthy controls. Immunol. Lett. 2016, 170, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Palma, J.; Tokarz-Deptuła, B.; Deptuła, J.; Deptuła, W. Natural antibodies–facts known and unknown. Cent. Eur. J. Immunol. 2018, 43, 466–475. [Google Scholar] [CrossRef]
- Le Berre, L.; Rousse, J.; Gourraud, P.A.; Imbert-Marcille, B.M.; Salama, A.; Evanno, G.; Semana, G.; Nicot, A.; Dugast, E.; Guérif, P.; et al. Decrease of blood anti-α1,3 Galactose Abs levels in multiple sclerosis (MS) and clinically isolated syndrome (CIS) patients. Clin. Immunol. 2017, 180, 128–135. [Google Scholar] [CrossRef]
- Montassier, E.; Berthelot, L.; Soulillou, J.P. Are the decrease in circulating anti-α1,3-Gal IgG and the lower content of galactosyl transferase A1 in the microbiota of patients with multiple sclerosis a novel environmental risk factor for the disease? Mol. Immunol. 2018, 93, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Galili, U.; Anaraki, F.; Thall, A.; Hill-Black, C.; Radic, M. One percent of human circulating B lymphocytes are capable of producing the natural anti-Gal antibody. Blood 1993, 82, 2485–2493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaison, P.L.; Kannan, V.M.; Geetha, M.; Appukuttan, P. Epitopes recognized by serum anti-α-galactoside antibody are present on brain glycoproteins in man. J. Niosci. 1993, 18, 187–193. [Google Scholar] [CrossRef]
- Nishi, K.; Tanegashima, A.; Yamamoto, Y.; Ushiyama, I.; Ikemoto, K.; Yamasaki, S.; Nishimura, A.; Rand, S.; Brinkmann, B. Utilization of lectin-histochemistry in forensic neuropathology: Lectin staining provides useful information for postmortem diagnosis in forensic neuropathology. Leg. Med. 2003, 5, 117–131. [Google Scholar] [CrossRef]
- Montassier, E.; Al-Ghalith, G.A.; Mathé, C.; Le Bastard, Q.; Douillard, V.; Garnier, A.; Guimon, R.; Raimondeau, B.; Touchefeu, Y.; Duchalais, E.; et al. Distribution of Bacterial α1,3-Galactosyltransferase Genes in the Human Gut Microbiome. Front. Immunol 2020, 10, 3000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westfall, S.; Dinh, D.M.; Pasinetti, G.M. Investigation of Potential Brain Microbiome in Alzheimer’s Disease: Implications of Study Bias. J. Alzheimer’s Dis. 2020, 75, 559–570. [Google Scholar] [CrossRef] [PubMed]
- McKhann, G.M.; Knopman, D.S.; Chertkow, H.; Hyman, B.T.; Jack, C.R., Jr.; Kawas, C.H.; Klunk, W.E.; Koroshetz, W.J.; Manly, J.J.; Mayeux, R.; et al. The diagnosis of dementia due to Alzheimer’s disease: Recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimer’s Dement. 2011, 7, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Burk, C.M.; Beitia, R.; Lund, P.K.; Dellon, E.S. High rate of galactose-alpha-1,3-galactose sensitization in both eosinophilic esophagitis and patients undergoing upper endoscopy. Dis. Esophagus 2016, 29, 558–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safaie, P.; Ham, M.; Kuang, P.; Mehta, A.S.; Wang, M.; Cheifetz, A.S.; Robson, S.; Lau, D.; Block, T.M.; Moss, A.C. Lectin-reactive anti-α-gal in patients with Crohn’s disease: Correlation with clinical phenotypes. Inflamm. Bowel Dis. 2013, 19, 2796–2800. [Google Scholar] [CrossRef] [Green Version]
- Chinuki, Y.; Morita, E. Alpha-Gal-containing biologics and anaphylaxis. Allergol. Int. 2019, 68, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Naso, F.; Stefanelli, U.; Buratto, E.; Lazzari, G.; Perota, A.; Galli, C.; Gandaglia, A. Alpha-Gal Inactivated Heart Valve Bioprostheses Exhibit an Anti-Calcification Propensity Similar to Knockout Tissues. Tissue Eng. Part A 2017, 23, 1181–1195. [Google Scholar] [CrossRef] [PubMed]
- Celarain, N.; Tomas-Roig, J. Aberrant DNA methylation profile exacerbates inflammation and neurodegeneration in multiple sclerosis patients. J. Neuroinflamm. 2020, 17, 21. [Google Scholar] [CrossRef]
- Mangold, A.; Lebherz, D.; Papay, P.; Liepert, J.; Hlavin, G.; Lichtenberger, C.; Adami, A.; Zimmermann, M.; Klaus, D.; Reinisch, W.; et al. Anti-Gal titers in healthy adults and inflammatory bowel disease patients. Transp. Proc. 2011, 43, 3964–3968. [Google Scholar] [CrossRef]
- Weksler, M.E.; Relkin, N.; Turkenich, R.; La Russe, S.; Zhou, L.; Szabo, P. Patients with Alzheimer disease have lower levels of serum anti-amyloid peptide antibodies than healthy elderly individuals. Exp. Gerontol. 2002, 37, 943–948. [Google Scholar] [CrossRef]
- Britschgi, M.; Olin, C.E.; Johns, H.T.; Takeda-Uchimura, Y.; LeMieux, M.C.; Rufibach, K.; Rajadas, J.; Zhang, H.; Tomooka, B.; Robinson, W.H.; et al. Neuroprotective natural antibodies to assemblies of amyloidogenic peptides decrease with normal aging and advancing Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2009, 106, 12145–12150. [Google Scholar] [CrossRef] [Green Version]
- Heineke, M.H.; van Egmond, M. Immunoglobulin A: Magic bullet or Trojan horse? Eur. J. Clin. Investig. 2017, 47, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Leblhuber, F.; Walli, J.; Tilz, G.P.; Wachter, H.; Fuchs, D. Systemische Veränderungen des Immunsystems bei Patienten mit Alzheimer-Demenz [Systemic changes of the immune system in patients with Alzheimer’s dementia]. Dtsch. Med. Wochenschr. 1998, 123, 787–791. [Google Scholar] [CrossRef]
- de la Rubia Ortí, J.E.; Sancho Castillo, S.; Benlloch, M.; Julián Rochina, M.; Corchón Arreche, S.; García-Pardo, M.P. Impact of the Relationship of Stress and the Immune System in the Appearance of Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 55, 899–903. [Google Scholar] [CrossRef]
- de la Rubia Ortí, J.E.; Prado-Gascó, V.; Sancho Castillo, S.; Julián-Rochina, M.; Romero Gómez, F.J.; García-Pardo, M.P. Cortisol and IgA are Involved in the Progression of Alzheimer’s Disease. A Pilot Study. Cell. Mol. Neurobiol. 2019, 39, 1061–1065. [Google Scholar] [CrossRef]
- Doss, S.; Wandinger, K.P.; Hyman, B.T.; Panzer, J.A.; Synofzik, M.; Dickerson, B.; Mollenhauer, B.; Scherzer, C.R.; Ivinson, A.J.; Finke, C.; et al. High prevalence of NMDA receptor IgA/IgM antibodies in different dementia types. Ann. Clin. Transl. Neurol. 2014, 1, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Smalla, K.H.; Angenstein, F.; Richter, K.; Gundelfinger, E.D.; Staak, A. Identification of fucose alpha(1-2) galactose epitope-containing glycoproteins from rat hippocampus. Neuroreport 1998, 9, 813–817. [Google Scholar] [CrossRef]
- Galili, U.; Buehler, J.; Shohet, S.B.; Macher, B.A. The human natural anti-Gal IgG. III. The subtlety of immune tolerance in man as demonstrated by crossreactivity between natural anti-Gal and anti-B antibodies. J. Exp. Med. 1987, 165, 693–704. [Google Scholar] [CrossRef]
- Pul, R.; Nguyen, D.; Schmitz, U.; Marx, P.; Stangel, M. Comparison of intravenous immunoglobulin preparations on microglial function in vitro: More potent immunomodulatory capacity of an IgM/IgA-enriched preparation. Clin. Neuropharmacol. 2002, 25, 254–259. [Google Scholar] [CrossRef]
- Bredesen, D.E.; Amos, E.C.; Canick, J.; Ackerley, M.; Raji, C.; Fiala, M.; Ahdidan, J. Reversal of cognitive decline in Alzheimer’s disease. Aging 2016, 8, 1250–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vossenkämper, A.; Blair, P.A.; Safinia, N.; Fraser, L.D.; Das, L.; Sanders, T.J.; Stagg, A.J.; Sanderson, J.D.; Taylor, K.; Chang, F.; et al. A role for gut-associated lymphoid tissue in shaping the human B cell repertoire. J. Exp. Med. 2013, 210, 1665–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerhardt, S.; Mohajeri, M.H. Changes of Colonic Bacterial Composition in Parkinson’s Disease and Other Neurodegenerative Diseases. Nutrients 2018, 10, 708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szablewski, L. Human Gut Microbiota in Health and Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 62, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, F.; Cechova, K.; Amlerova, J.; Hort, J. Antibiotics, gut microbiota, and Alzheimer’s disease. J. Neuroinflamm. 2019, 16, 108. [Google Scholar] [CrossRef]
- Liu, S.; Gao, J.; Zhu, M.; Liu, K.; Zhang, H.L. Gut Microbiota and Dysbiosis in Alzheimer’s Disease: Implications for Pathogenesis and Treatment. Mol. Neurobiol. 2020, 57, 5026–5043. [Google Scholar] [CrossRef]
- Pluta, R.; Ułamek-Kozioł, M.; Januszewski, S.; Czuczwar, S.J. Gut microbiota and pro/prebiotics in Alzheimer’s disease. Aging 2020, 12, 5539–5550. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, D.; Zhou, G.; Li, C. Dietary Pattern, Gut Microbiota, and Alzheimer’s Disease. J. Agric. Food Chem. 2020, 68, 12800–12809. [Google Scholar] [CrossRef]
- Megur, A.; Baltriukienė, D.; Bukelskienė, V.; Burokas, A. The Microbiota-Gut-Brain Axis and Alzheimer’s Disease: Neuroinflammation Is to Blame? Nutrients 2020, 13, 37. [Google Scholar] [CrossRef]
- Miyake, S.; Kim, S.; Suda, W.; Oshima, K.; Nakamura, M.; Matsuoka, T.; Chihara, N.; Tomita, A.; Sato, W.; Kim, S.W.; et al. Dysbiosis in the Gut Microbiota of Patients with Multiple Sclerosis, with a Striking Depletion of Species Belonging to Clostridia XIVa and IV Clusters. PLoS ONE 2015, 10, e0137429. [Google Scholar] [CrossRef] [Green Version]
- Brenner, S.R. Blue-green algae or cyanobacteria in the intestinal micro-flora may produce neurotoxins such as Beta-N-Methylamino-L-Alanine (BMAA) which may be related to development of amyotrophic lateral sclerosis, Alzheimer’s disease and Parkinson-Dementia-Complex in humans and Equine Motor Neuron Disease in horses. Med. Hypotheses 2013, 80, 103. [Google Scholar] [CrossRef]
- Schwartz, K.; Boles, B.R. Microbial amyloids--functions and interactions within the host. Curr. Opin. Microbiol. 2013, 16, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Kesika, P.; Suganthy, N.; Sivamaruthi, B.S.; Chaiyasut, C. Role of gut-brain axis, gut microbial composition, and probiotic intervention in Alzheimer’s disease. Life Sci. 2021, 264, 118627. [Google Scholar] [CrossRef]
- Guevara, J.; Espinosa, B.; Zenteno, E.; Vázguez, L.; Luna, J.; Perry, G.; Mena, R. Altered glycosylation pattern of proteins in Alzheimer disease. J. Neuropathol. Exp. Neurol. 1998, 57, 905–914. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Variables | AD (N. 30) | HS (N. 30) | F (1,59)/X2 # | p |
|---|---|---|---|---|
| Females/males (N.) | 19/11 | 15/15 | 1.086 | 0.297 |
| Age (mean ± SD, y) (range, y) | 83.77 ± 5.89 (70–96) | 80.83 ± 6.04 (70–93) | 3.631 | 0.062 |
| Education level (mean + SD, y) | 9.10 ± 5.27 | 11.77 ± 4.19 | 4.708 | 0.034 |
| BMI (mean ± SD, kg/m2) | 24.37 ± 4.56 | 26.37 ± 3.70 | 3.475 | 0.067 |
| MMSE (score) | 17.88 ± 6.85 | 30.07 ± 1.31 | 91.632 | <0.001 |
| Blood group (N; %) | ||||
| 0 | 13; 43.3% | 14; 46.6% | 0.067 | 0.795 |
| A | 13; 43.3% | 8; 26.6% | 1.832 | 0.176 |
| B | 3; 10% | 6; 20% | 1.176 | 0.278 |
| AB | 1; 3.3% | 2; 6.6% | 0.351 | 0.554 |
| Medical history (N; %) | ||||
| Smoke * | 3; 10% | 3; 10% | 0.000 | 1.000 |
| Dyslipidemia | 11; 36.6% | 12; 40% | 0.071 | 0.791 |
| Diabetes | 8; 26.6% | 6; 20% | 0.373 | 0.542 |
| Hypertension | 17; 56.6% | 19; 63.3 % | 0.278 | 0.598 |
| Myocardial infarction | 3; 10% | 3; 10% | 0.000 | 1.000 |
| TIA/Stroke | 3; 10% | 1; 3.33% | 1.071 | 0.301 |
| Drugs (N; %) | ||||
| Antihypertensive | 17; 56.6% | 18; 60% | 0.069 | 0.793 |
| Lipid-lowering | 10; 30% | 11; 36.6% | 0.073 | 0.787 |
| Hypoglycemic | 8; 26.6% | 6; 20% | 0.373 | 0.542 |
| Antiacid | 12; 40% | 11; 36.6% | 0.071 | 0.791 |
| Antiplatelet | 13; 43.3% | 12; 40% | 0.069 | 0.793 |
| Anti-inflammatory | 3; 10% | 3; 10% | 0.000 | 1.000 |
| Ig (O.D. 450 nm) | AD | HS | Partial η2 | F | df | p |
|---|---|---|---|---|---|---|
| IgG | 0.696 ± 0.18 | 0.837 ± 0.16 | 0.121 | 5.784 | (1, 42) | 0.021 |
| IgA | 0.721 ± 0.13 | 0.652 ± 0.13 | 0.119 | 5.661 | (1, 42) | 0.022 |
| IgM | 0.413 ± 0.98 | 0.56 ± 0.11 | 0.419 | 30.313 | (1, 42) | <0.001 |
| Micro-Organisms | Increasing | Decreasing | References |
|---|---|---|---|
| Cyanobacteria | X | [42,50] | |
| Chlamydia pneumoniae | X | [42,43,45] | |
| Borrelia burgdorferi | X | [43,45] | |
| Escherichia coli | X | [43,44,45,46,47] | |
| Shigella | X | [43,44,45,46,47] | |
| Enterococcus | X | [41,43,44,46,47] | |
| Blautia glucerasea/producta | X | [41,48] | |
| Clostridium perfringens/saccharoliticum | X | [41,44,45,47] | |
| Gemellaceae (Gemella) | X | [41,48] | |
| Mogibacteriaceae | X | [41,48] | |
| Veillonellaceae (Dialister) | X | [41] | |
| Tissirellaceae | X | [41,48] | |
| Bifidobacteriaceae (Bifidobacterium) | X | [41] | |
| Bacterodaceae (Bacteroides) | X | [41,44,45,46,47] | |
| Akkermansia | X | [46] | |
| Bacillus subtilis | X | [46,47] | |
| Klebsiella pneumonia | X | [44,46] | |
| Mycobacterium spp. | X | [46] | |
| Staphylococcus aureus | X | [44,46] | |
| Streptococcus spp. | X | [46] | |
| Fusobacteriaceae | X | [47] | |
| Prevotellaceae | X | [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angiolillo, A.; Gandaglia, A.; Arcaro, A.; Carpi, A.; Gentile, F.; Naso, F.; Di Costanzo, A. Altered Blood Levels of Anti-Gal Antibodies in Alzheimer’s Disease: A New Clue to Pathogenesis? Life 2021, 11, 538. https://doi.org/10.3390/life11060538
Angiolillo A, Gandaglia A, Arcaro A, Carpi A, Gentile F, Naso F, Di Costanzo A. Altered Blood Levels of Anti-Gal Antibodies in Alzheimer’s Disease: A New Clue to Pathogenesis? Life. 2021; 11(6):538. https://doi.org/10.3390/life11060538
Chicago/Turabian StyleAngiolillo, Antonella, Alessandro Gandaglia, Alessia Arcaro, Andrea Carpi, Fabrizio Gentile, Filippo Naso, and Alfonso Di Costanzo. 2021. "Altered Blood Levels of Anti-Gal Antibodies in Alzheimer’s Disease: A New Clue to Pathogenesis?" Life 11, no. 6: 538. https://doi.org/10.3390/life11060538
APA StyleAngiolillo, A., Gandaglia, A., Arcaro, A., Carpi, A., Gentile, F., Naso, F., & Di Costanzo, A. (2021). Altered Blood Levels of Anti-Gal Antibodies in Alzheimer’s Disease: A New Clue to Pathogenesis? Life, 11(6), 538. https://doi.org/10.3390/life11060538