
Multigene Phylogenetics and Morphology Reveal Five Novel Lasiodiplodia Species Associated with Blueberries


Abstract
:1. Introduction
2. Methods and Materials
2.1. Sample Collections and Fungal Isolation
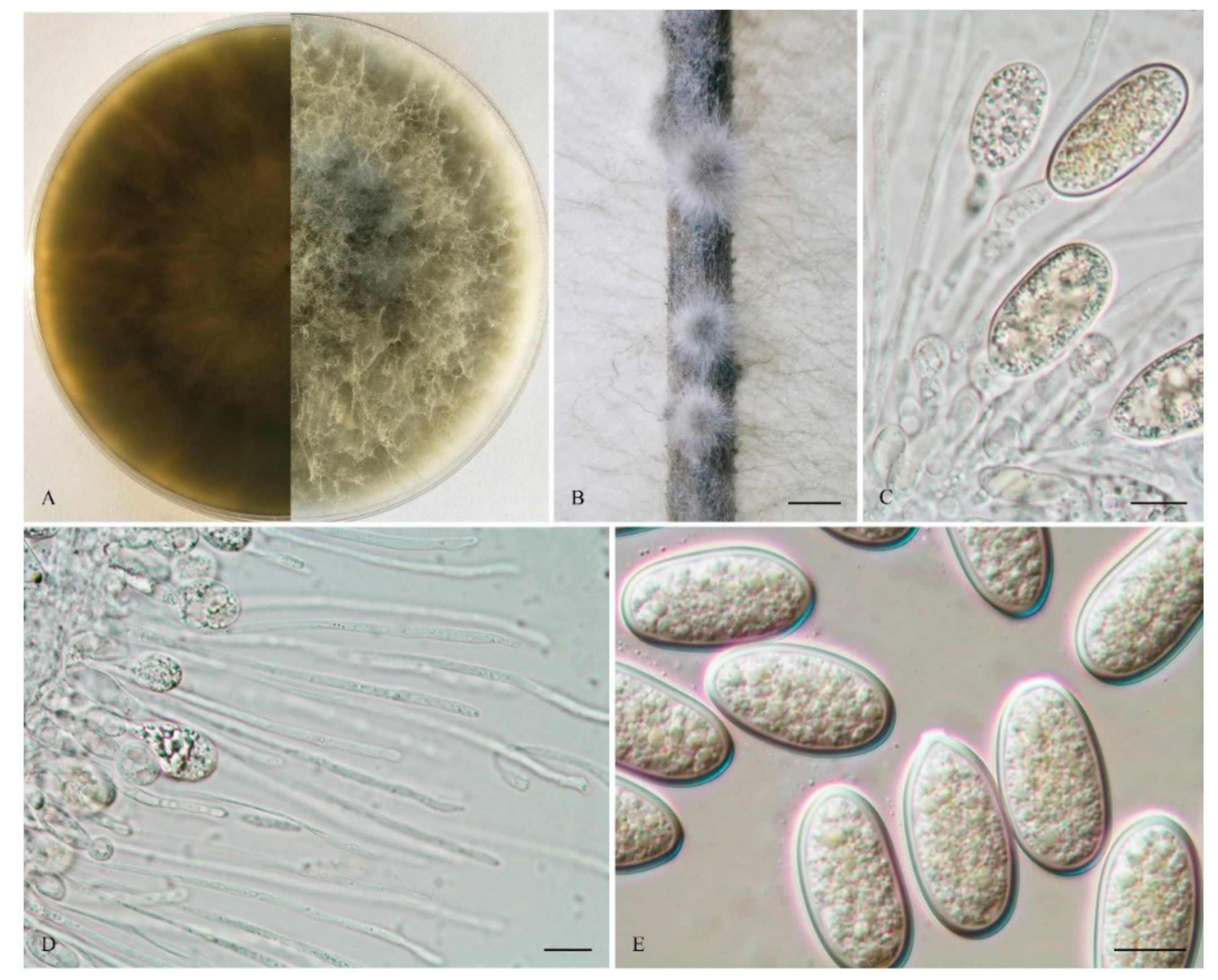
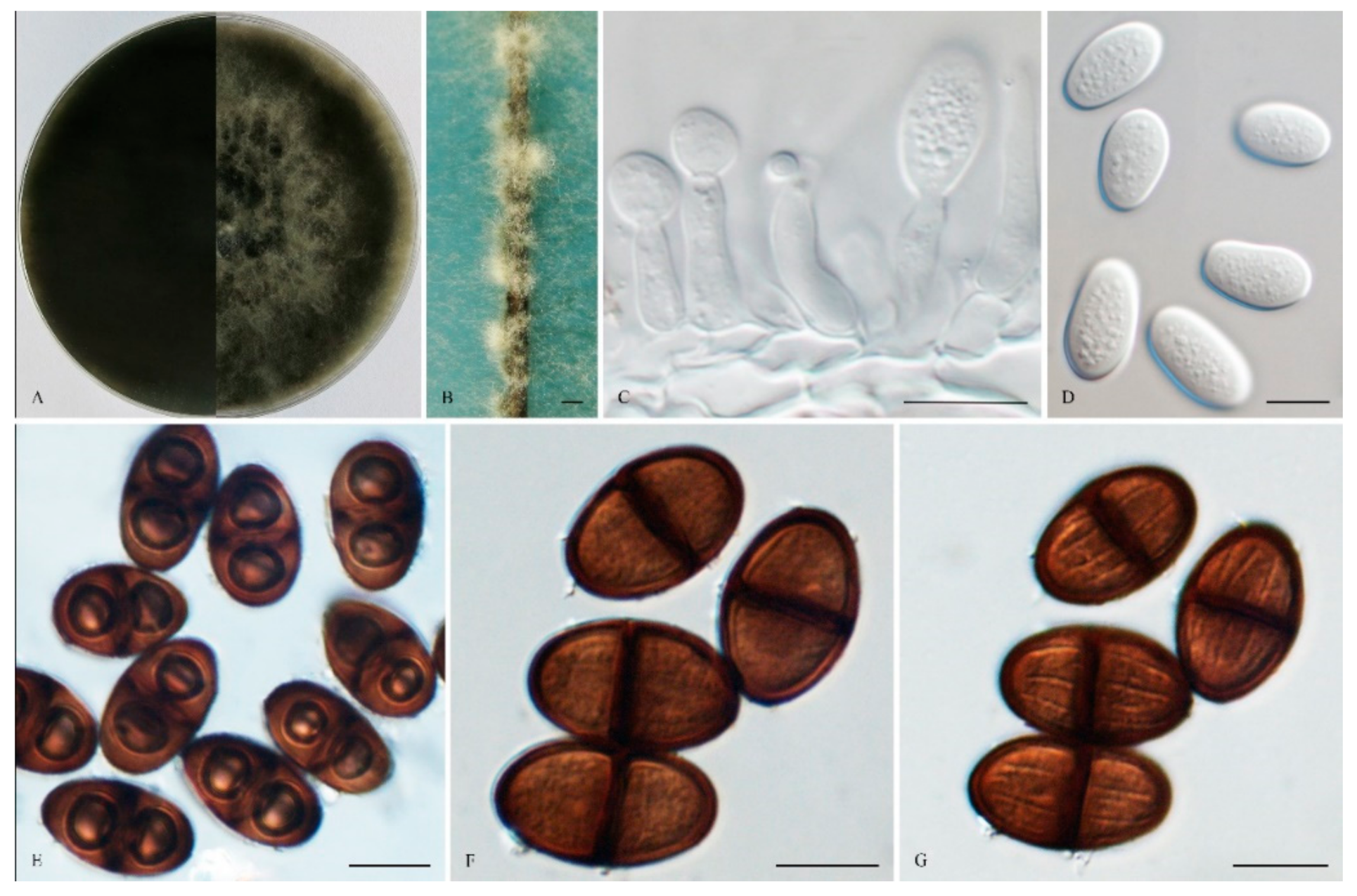
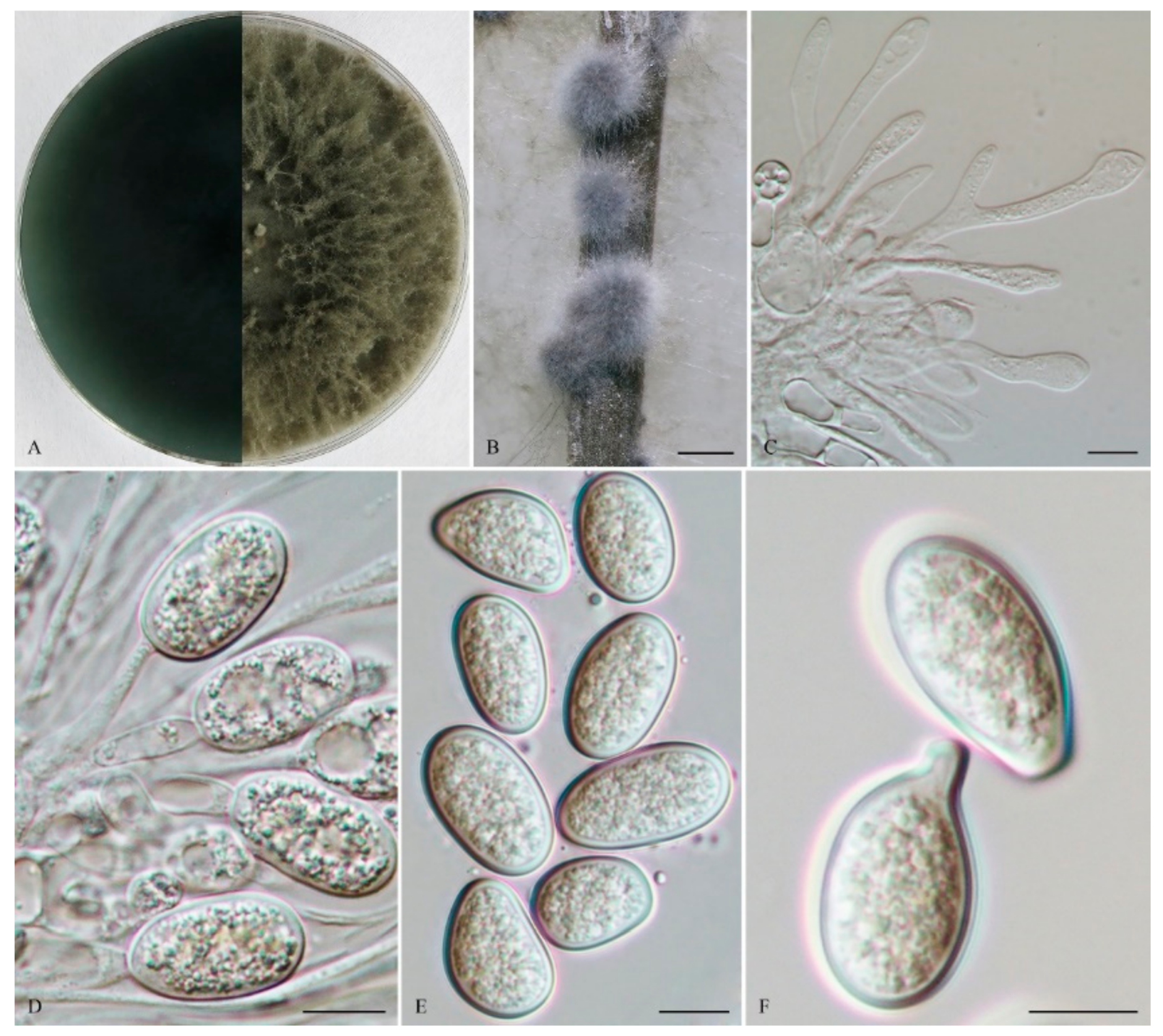
2.2. Morphological Characterization
2.3. DNA Extraction, PCR Amplification
2.4. Sequence Alignment and Phylogenetic Analysis
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shi, H.Z.; Liu, H.M. The current research situation of Blueberry. J. Jiangsu For. Sci. Technol. 2009, 36, 48–51. [Google Scholar]
- Boyzo, M.J.; Rebollar, A.A.; Silva, R.H.V.; Ramirez, M.G. First Report of Neofusicoccum parvum causing stem blight and dieback of blueberry in Mexico. Plant Dis. 2016, 100, 2524. [Google Scholar]
- Castillo, S.; Borrero, C.; Castaño, R.; Rodríguez, A.; Avilés, M. First report of canker disease caused by Neofusicoccum parvum and N australe on blueberry bushes in spain. Plant Dis. 2013, 97, 1112. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.T.; Rooneylatham, S.; Wright, A.F. First report of stem blight of blueberry in California caused by Neofusicoccum parvum. Plant Dis. 2014, 98, 1280. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.S.; Qiu, X.L.; Yi, K.S.; Yu, X.F.; Yu, L.; Kong, C.S.; Qiu, X.L.; Yi, K.S.; Yu, X.F.; Yu, L. First Report of Neofusicoccum vitifusiforme Causing Blueberry Blight of Blueberry in China. Plant Dis. 2010, 94, 1373. [Google Scholar] [CrossRef]
- Sammonds, J.; Billones, R.; Ridgway, H.; Walter, M.; Jaspers, M. Survey of blueberry farms for Botryosphaeria dieback and crown rot pathogens. New Zealand Plant Prot. 2009, 62, 238–242. [Google Scholar] [CrossRef] [Green Version]
- Scarlett, K.A.; Shuttleworth, L.A.; Collins, D.; Rothwell, C.; Guest, D.I.; Daniel, R. Botryosphaeriales associated with stem blight and dieback of blueberry (Vaccinium spp.) in New South Wales and Western Australia. Australas. Plant Pathol. 2019, 48, 45–57. [Google Scholar] [CrossRef]
- Wright, A.F.; Harmon, P.F. First Report of Lasiodiplodia theobromae Associated with Stem Blight of Southern Highbush Blueberries in Florida. Plant Dis. 2009, 93, 962. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.F.; Harmon, P.F. Identification of Species in the Botryosphaeriaceae Family Causing Stem Blight on Southern Highbush Blueberry in Florida. Plant Dis. 2010, 94, 966–971. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.N. Etiology, Molecular Detection and Control of Blueberry Stem Blight in China. Ph.D. Thesis, Hebei Agricultural University, Baoding, China, 2016. [Google Scholar]
- Xu, C.N.; Zhang, H.J.; Zhou, Z.S.; Hu, T.; Wang, S.T.; Wang, Y.N.; Cao, K.Q. Identification and distribution of Botryosphaeriaceae species associated with blueberry stem blight in China. Eur. J. Plant Pathol. 2015, 143, 737–752. [Google Scholar] [CrossRef]
- Yu, L.; Impaprasert, R.; Zhao, J.R.; Xu, S.G.; Wu, X. Stem dieback of highbush blueberries caused by Neofusicoccum parvum in China. New Dis. Rep. 2016, 27, 3. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Wang, Y.; He, W.; Zhang, Y. Stem blight of blueberry caused by Lasiodiplodia vaccinii sp. nov. in China. Plant Dis. 2013, 13, 2041–2050. [Google Scholar] [CrossRef]
- Evans, E.A.; Ballen, F.H. An overview of US blueberry production, trade, and consumption, with special reference to Florida. Food Resour. Econ. 2017. Available online: https://edis.ifas.ufl.edu/fe952 (accessed on 15 September 2020).
- Popović, T.; Blagojević, J.; Aleksić, G.; Jelušić, A.; Krnjajić, S.; Milovanović, P. A blight disease on highbush blueberry associated with Macrophomina phaseolina in Serbia. Can. J. Plant Pathol. 2018, 40, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.D.; Sun, H.Y.; Chen, L. China Blueberry Industry Development Report. China Fruits 2016, 5, 1–10. Available online: http://caod.oriprobe.com/articles/49349906/wo_guo_lan_zuo_chan_ye_fa_zhan_bao_gao_.htm (accessed on 15 October 2020).
- Phillips, A.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.; Groenewald, J.; Crous, P. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A. Botryosphaeriaceae: Current status of genera and species. Mycosphere 2016, 7, 1001–1073. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; McKenzie, E.H.C.; Jeewon, R.; Phillips, A.J.L.; Perera, R.H.; De Silva, N.I.; Maharachchikumburua, S.S.N.; Samarakoon, M.C.; Ekanayake, A.H.; et al. One stop shop III: Taxonomic update with molecular phylogeny for important phytopathogenic genera: 51–75 (2019). Fungal Divers. 2019, 98, 77–160. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; Jeewon, R.; Ghobad-Nejhad, M.; Wanasinghe, D.N.; Liu, N.; Phillips, A.J.L.; Oliveira-Filho, J.R.C.; Da Silva, G.A.; Gibertoni, T.B.; et al. One stop shop II: Taxonomic update with molecular phylogeny for important phytopathogenic genera: 26–50 (2019). Fungal Divers. 2019, 94, 41–129. [Google Scholar] [CrossRef]
- Poll, J.T.K.; Wood, F.H. Blueberries in New Zealand. Acta Hortic. 1985, 165, 35–46. [Google Scholar] [CrossRef]
- Wang, J.N.; Zhao, H.H.; Yu, Y.Y.; Li, X.D.; Liang, C.; Li, B.D. The pathogen causing Lasiodiplodia twig blight of blueberry. Mycosystema 2016, 35, 657–665. [Google Scholar]
- Milholland, R.D. Factors affecting sporulation and infection by the blueberry stem canker fungus, Botryosphaeria corticis. Phytopathology 1972, 62, 137–139. [Google Scholar] [CrossRef]
- Milholland, R.D. Histopathology and pathogenicity of Botryosphaeria dothidea on blueberry stems. Phytopathology 1972, 62, 654–660. [Google Scholar] [CrossRef]
- Phillips, A.J.; Oudemans, P.V.; Correia, A.; Alves, A. Characterisation and epitypification of Botryosphaeria corticis, the cause of blueberry cane canker. Fungal Divers. 2006, 21, 141–155. [Google Scholar]
- Creswell, T.C.; Milholland, R.D. Spore release and infection periods of Botryosphaeria dothidea on blueberry in North Carolina. Plant Dis. 1988, 72, 342. [Google Scholar] [CrossRef]
- Witcher, W.; Clayton, C.N. Blueberry stem blight caused by Botryosphaeria dothidea. Phytopathology 1962, 53, 705–712. [Google Scholar]
- Tennakoon, K.M.S.; Ridgway, H.J.; Jaspers, M.V.; Jones, E.E. Factors affecting Neofusicoccum ribis, infection and disease progression in blueberry. European journal of plant pathology. Eur. J. Plant Pathol. 2018, 151, 87–99. [Google Scholar]
- Espinoza, J.G.; Briceño, E.X.; Chávez, E.R.; Úrbez-Torres, J.R.; Latorre, B.A. Neofusicoccum spp. associated with stem canker and dieback of blueberry in Chile. Plant Dis. 2009, 93, 1187–1194. [Google Scholar] [CrossRef] [Green Version]
- Borrero, C.; Castaño, R.; Avilés, M. First report of Pestalotiopsis clavispora (Neopestalotiopsis clavispora) causing canker and twig dieback on blueberry bushes in Spain. Plant Dis. 2009, 102, 1178. [Google Scholar] [CrossRef]
- Choi, I.Y. First report of bark dieback on blueberry caused by Botryosphaeria dothidea in Korea. Plant Dis. 2011, 95, 227. [Google Scholar] [CrossRef]
- Daniel, R.; Scarlett, K.; Rothwell, C.T.; Shuttleworth, L.; Guest, D.I. Botryosphaeriaceae associated with stem blight and dieback of blueberry (Vaccinium corymbosum) in Australia. In Proceedings of the International Congress of Plant Pathology (ICPP) 2018: Plant Health in A Global Economy, Boston, MA, USA, 29 July–3 August 2018; Available online: https://apsnet.confex.com/apsnet/ICPP2018/meetingapp.cgi/Paper/10147 (accessed on 15 August 2020).
- Wiseman, M.S.; Serdani, M.; Putnam, M.L. A New Cane Dieback Disease of Northern Highbush Blueberry in the United States Caused by Lasiodiplodia mediterranea. Plant Dis. 2017, 101, 1317. [Google Scholar] [CrossRef]
- Lopes, A.; Barradas, C.; Phillips, A.J.; Alves, A. Diversity and phylogeny of Neofusicoccum species occurring in forest and urban environments in Portugal. Mycosphere 2016, 7, 906–920. [Google Scholar] [CrossRef]
- Dou, Z.P.; He, W.; Zhang, Y. Does morphology matter in taxonomy of Lasiodiplodia? An answer from Lasiodiplodia hyalina sp nov. Mycosphere 2017, 8, 1014–1027. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Phillips, A.J.; Hyde, K.D.; Chethana, K.W.; Zhang, W.; Zhao, W.S.; Yan, J.Y.; Li, X.H. Mycosphere Essays 14: Assessing the aggressiveness of plant pathogenic Botryosphaeriaceae. Mycosphere 2016, 7, 883–892. [Google Scholar] [CrossRef]
- Pavlic, D.; Slippers, B.; Coutinho, T.A.; Gryzenhout, M.; Wingfield, M.J. Lasiodiplodia gonubiensis sp. nov., a new Botryosphaeria anamorph from native Syzygium cordatum in South Africa. Stud Mycol. 2004, 50, 313–322. [Google Scholar]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jeewon, R.; Yeung, Q.S.Y.; Hyde, K.D. Novel phylogenetic group within Thozetella (Chaetosphaeriaceae): A new taxon based on morphology and DNA sequence analyses. Can. J. Microbiol. 2009, 55, 680–687. [Google Scholar] [CrossRef]
- Dou, Z.P.; He, W.; Zhang, Y. Lasiodiplodia chinensis sp. nov., a new holomorphic species from China. Mycosphere 2017, 8, 521–532. [Google Scholar] [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Shinsky, J.J., White, T.J., Eds.; Elsevier: Berlin/Heidelberg, Germany, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Alves, A.; Crous, P.W.; Correia, A.; Phillips, A.J. Morphological and molecular data reveal cryptic speciation in Lasiodiplodia theobromae. Fungal Divers. 2008, 28, 1–3. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruywagen, E.M.; Slippers, B.; Roux, J.; Wingfield, M.J. Phylogenetic species recognition and hybridisation in Lasiodiplodia: A case study on species from baobabs. Fungal Biol. 2017, 121, 420–436. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, H.K.; Fournier, J.; Crous, P.W.; Jeewon, R.; Pointing, S.B.; Hyde, K.D. Towards a phylogenetic clarification of Lophiostoma/Massarina and morphologically similar genera in the Pleosporales. Fungal Divers. 2009, 38, 225–251. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swofford, D.L. Paup*: Phylogenetic analysis using parsimony (*and other methods) 4.0. B5. Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Jeewon, R.; Fournier, J.; Hyde, K.D. Multi-gene phylogeny and morphotaxonomy of Amniculicola lignicola: A novel freshwater fungus from France and its relationships to the Pleosporales. Mycol. Res. 2008, 112, 1186–1194. [Google Scholar] [CrossRef]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of Akaike information criterion and Bayesian approaches over likelihood ratio tests. Syst Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef]
- Page, R.D. Tree View: An application to display phylogenetic trees on personal computers. Bioinformatics 1996, 12, 357–358. [Google Scholar] [CrossRef] [Green Version]
- Trakunyingcharoen, T.; Lombard, L.; Groenewald, J.Z.; Cheewangkoon, R.; To-Anun, C.; Crous, P.W. Caulicolous Botryosphaeriales from Thailand. Persoonia 2015, 34–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdollahzadeh, J.; Javadi, A.; Goltapeh, E.M.; Zare, R.; Phillips, A.J. Phylogeny and morphology of four new species of Lasiodiplodia from Iran. Persoonia 2010, 25, 1. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Gálvez, E.; Guerrero, P.; Barradas, C.; Crous, P.W.; Alves, A. Phylogeny and pathogenicity of Lasiodiplodia species associated with dieback of mango in Peru. Fungal Biol. 2017, 121, 452–465. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, S.; Zhao, L.; Sun, X.; He, W.; Zhang, Y.; Dai, Y.C. Lasiodiplodia spp. associated with Aquilaria crassna in Laos. Mycol. Prog. 2019, 18, 683–701. [Google Scholar] [CrossRef]
- Begoude, B.D.; Slippers, B.; Wingfield, M.J.; Roux, J. Botryosphaeriaceae associated with Terminalia catappa in Cameroon, South Africa and Madagascar. Mycol Prog. 2010, 9, 101–123. [Google Scholar] [CrossRef] [Green Version]
- Linaldeddu, B.T.; Deidda, A.; Scanu, B.; Franceschini, A.; Serra, S.; Berraf-Tebbal, A.; Boutiti, M.Z.; Jamâa, M.B.; Phillips, A.J. Diversity of Botryosphaeriaceae species associated with grapevine and other woody hosts in Italy, Algeria and Tunisia, with descriptions of Lasiodiplodia exigua and Lasiodiplodia mediterranea sp. nov. Fungal Divers. 2015, 71, 201–214. [Google Scholar] [CrossRef]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- de Silva, N.I.; Phillips, A.J.; Liu, J.K.; Lumyong, S.; Hyde, K.D. Phylogeny and morphology of Lasiodiplodia species associated with Magnolia forest plants. Sci. Rep. 2019, 9, 1. Available online: https://www.nature.com/articles/s41598-019-50804-x (accessed on 15 October 2020). [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Cultures | Host | Locality | Longitude and Latitude | GenBank | |||
|---|---|---|---|---|---|---|---|---|
| ITS | tef1-α | TUB | RPB2 | |||||
| L. aquilariae | CGMCC 3.18471 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783442 | KY848600 | N/A | KY848562 |
| L. avicenniae | CMW41467 | Avicennia marina | South Africa | 25°44′ S, 28°15′ E * | KP860835 | KP860680 | KP860758 | KU587878 |
| L. avicenniae | LAS199 | Avicennia marina | South Africa | 25°44′ S, 28°15′ E * | KU587957 | KU587947 | KU587868 | KU587880 |
| L. brasiliensis | CMM 2321 | Carica papaya | Brazil * | 15°47′ S, 47°55′ W | KY783475 | KY848612 | KY848556 | KY848595 |
| L. brasiliensis | CMM 4015 | Mangifera indica | Brazil * | 15°47′S, 47°55′W | JX464063 | JX464049 | N/A | N/A |
| L. brasiliensis | CMW 35884 | Adansonia madagascariensis | Madagascar * | 18°52′ S, 47°29′ E | KU887094 | KU886972 | KU887466 | KU696345 |
| L. bruguierae | CMW41470 | Bruguiera gymnorrhiza | South Africa * | 25°44′ S, 28°15′ E | KP860833 | KP860678 | KP860756 | KU587875 |
| L. bruguierae | CMW42480 | Bruguiera gymnorrhiza | South Africa * | 25°44′ S, 28°15′ E | KP860832 | KP860677 | KP860755 | KU587876 |
| L. caatinguensis | CMM1325 | Citrus sinensis | Brazil * | 15°47′ S, 47°55′ W | KT154760 | KT008006 | KT154767 | N/A |
| L. caatinguensis | IBL381 | Spondias purpurea | Brazil * | 15°47′ S, 47°55′ W | KT154757 | KT154751 | KT154764 | N/A |
| L. chinensis | CGMCC3.18044 | Vaccinium uliginosum | Shandong, China | 36°03′ N, 120°22′ E | KX499875 | KX499913 | KX499988 | KX499951 |
| L. chinensis | CGMCC3.18061 | unknown | Hainan, China | 20°0′ N, 110°12′ E | KX499889 | KX499927 | KX500002 | KX499965 |
| L. chinensis | CGMCC3.18066 | Hevea brasiliensis | Hainan, China | 20°0′ N, 110°12′ E | KX499899 | KX499937 | KX500012 | KX499974 |
| L. chinensis | CGMCC3.18067 | Sterculia lychnophora | Hainan, China | 20°0′ N, 110°12′ E | KX499901 | KX499939 | KX500014 | KX499976 |
| L. citricola | IRAN1521C | Citrus sp. | Iran * | 33°05′ N, 43°06′ E | GU945353 | GU945339 | KU887504 | KU696350 |
| L. citricola | IRAN1522C | Citrus sp. | Iran * | 33°05′ N, 43°06′ E | GU945354 | GU945340 | KU887505 | KU696351 |
| L. clavispora | CGMCC 3.19594 | Vaccinium uliginosum | Fujian, China | 26°06′ N, 119°17′ E | MK802166 | N/A | MK816339 | MK809507 |
| L. clavispora | CGMCC 3.19595 | Vaccinium uliginosum | Fujian, China | 26°06′ N, 119°17′ E | MK802165 | N/A | MK816338 | MK809506 |
| L. crassispora | CMW 13488 | Eucalyptus urophylla | Venezuela * | 10°28′ N, 66°53′ W | DQ103552 | DQ103559 | KU887507 | KU696352 |
| L. crassispora | WAC12533 | Santalum album | Australia * | 32° S, 150° E | DQ103550 | DQ103557 | KU887506 | KU696353 |
| L. curvata | CGMCC 3.18456 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783437 | KY848596 | KY848529 | KY848557 |
| L. curvata | CGMCC 3.18476 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783443 | KY848601 | KY848532 | KY848563 |
| L. euphorbicola | CMM 3609 | Jatropha curcas | Brazil * | 15°47′ S, 47°55′ W | KU887149 | KU887026 | KU887455 | KU696346 |
| L. euphorbicola | CMW 33350 | Adansonia digitata | Botswana * | 24°36′ S, 25°40′ E | KU887187 | KU887063 | KU887494 | KU696347 |
| L. euphorbicola | CMW 36231 | Adansonia digitata | Zimbabwe * | 17°49′ S, 31°03′ E | KF234543 | KF226689 | KF254926 | N/A |
| L. exigua | BL184 | Retama raetam | Tunisia * | 34°44′ N, 10°44′ E | KJ638318 | KJ638337 | N/A | N/A |
| L. exigua | CBS 137785 | Retama raetam | Tunisia * | 34°44′ N, 10°44′ E | KJ638317 | KJ638336 | KU887509 | KU696355 |
| L. fujianensis | CGMCC 3.19593 | Vaccinium uliginosum | Fujian, China | 26°06′ N, 119°17′ E | MK802164 | MK887178 | MK816337 | MK809505 |
| L. gilanensis | IRAN 1501C | Unknown | Iran * | 33°05′ N, 43°06′ E | GU945352 | GU945341 | KU887510 | KU696356 |
| L. gilanensis | IRAN 1523C | Unknown | Iran * | 33°05′ N, 43°06′ E | GU945351 | GU945342 | KU887511 | KU696357 |
| L. gonubiensis | CMW 14077 | Syzygium cordatum | South Africa * | 25°44′ S, 28°15′ E | AY639595 | DQ103566 | DQ458860 | KU696359 |
| L. gonubiensis | CMW 14078 | Syzygium cordatum | South Africa * | 25°44′ S, 28°15′ E | AY639594 | DQ103567 | EU673126 | KU696358 |
| L. gravistriata | CMM 4564 | Anacardium humile | Brazil * | 15°47′ S, 47°55′ W | KT250949 | KT250950 | N/A | N/A |
| L. gravistriata | CMM 4565 | Anacardium humile | Brazil * | 15°47′ S, 47°55′ W | KT250947 | KT266812 | N/A | N/A |
| L. henanica | CGMCC 3.19176 | Vaccinium uliginosum | Shandong, China | 36°03′ N, 120°22′ E | MH729351 | MH729357 | MH729360 | MH729354 |
| L. hormozganensis | IRAN 1498C | Mangifera indica | Iran * | 33°05′ N, 43°06′ E | GU945356 | GU945344 | KU887514 | KU696360 |
| L. hormozganensis | IRAN 1500C | Olea sp. | Iran * | 33°05′ N, 43°06′ E | GU945355 | GU945343 | KU887515 | KU696361 |
| L. iraniensis | CMM 3610 | Jatropha curcas | Brazil * | 15°47′ S, 47°55′ W | KF234544 | KF226690 | KF254927 | N/A |
| L. iraniensis | CMW 36237 | Adansonia digitata | Mozambique * | 25°56′ S, 32°35′ E | KU887121 | KU886998 | KU887499 | KU696348 |
| L. iraniensis | CMW 36239 | Adansonia digitata | Mozambique * | 25°56′ S, 32°35′ E | KU887123 | KU887000 | KU887501 | KU696349 |
| L. iraniensis | IRAN 1502C | Juglans sp. | Iran * | 33°05′ N, 43°06′ E | GU945347 | GU945335 | KU887517 | KU696362 |
| L. iraniensis | IRAN 1520C | Salvadora persica | Iran * | 33°05′ N, 43°06′ E | GU945348 | GU945336 | KU887516 | KU696363 |
| L. irregularis | CGMCC 3.18468 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783472 | KY848610 | KY848553 | KY848592 |
| L. laeliocattleyae | BOT 29 | Mangifera indica | Egypt * | 30°03′ N, 31°14′ E | JN814401 | JN814428 | N/A | N/A |
| L. laeliocattleyae | CBS 130992 | Mangifera indica | Egypt * | 30°03′ N, 31°14′ E | JN814397 | JN814424 | KU887508 | KU696354 |
| L. laosensis | CGMCC 3.18464 | Aquilaria crassna | Laos | 17°59′N, 102°34′E | KY783471 | KY848609 | KY848552 | KY848591 |
| L. laosensis | CGMCC 3.18473 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783450 | KY848603 | KY848536 | KY848570 |
| L. lignicola | CBS 134112 | dead wood | Thailand * | 13°43′ N, 100°28′ E | JX646797 | KU887003 | JX646845 | KU696364 |
| L. lignicola | CGMCC 3.18460 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783462 | N/A | N/A | KY848582 |
| L. lignicola | CGMCC 3.18483 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783449 | N/A | N/A | KY848569 |
| L. lignicola | MFLUCC 11-0656 | dead wood | Thailand * | 13°43′ N, 100°28′ E | JX646798 | N/A | JX646846 | N/A |
| L. macroconidica | CGMCC 3.18479 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783438 | KY848597 | KY848530 | KY848558 |
| L. macrospora | CMM3833 | Jatropha curcas | Brazil * | 15°47′ S, 47°55′ W | KF234557 | KF226718 | KF254941 | N/A |
| L. mahajangana | CMW 27801 | Terminalia catappa | Madagascar * | 18°52′ S, 47°29′ E | FJ900595 | FJ900641 | FJ900630 | KU696365 |
| L. mahajangana | CMW 27818 | Terminalia catappa | Madagascar * | 18°52′ S, 47°29′ E | FJ900596 | FJ900642 | FJ900631 | KU696366 |
| L. margaritacea | CBS 122065 | Adansonia gibbosa | Western Australia * | 31°56′ S, 115°55′ E | EU144051 | EU144066 | N/A | N/A |
| L. margaritacea | CBS 122519 | Adansonia gibbosa | Western Australia * | 31°56′ S, 115°55′ E | EU144050 | EU144065 | KU887520 | KU696367 |
| L. mediterranea | CBS 137783 | Quercus ilex | Italy * | 41°54′ N, 12°18′ E | KJ638312 | KJ638331 | KU887521 | KU696368 |
| L. mediterranea | CBS 137784 | Vitis vinifera | Italy * | 41°54′ N, 12°18′ E | KJ638311 | KJ638330 | KU887522 | KU696369 |
| L. microcondia | CGMCC 3.18485 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783441 | KY848614 | N/A | KY848561 |
| L. missouriana | UCD 2193MO | Vitis sp. | USA * | 38° N, 97° W | HQ288225 | HQ288267 | HQ288304 | KU696370 |
| L. missouriana | UCD 2199MO | Vitis sp. | USA * | 38° N, 97° W | HQ288226 | HQ288268 | HQ288305 | KU696371 |
| L. nanpingensis | CGMCC 3.19596 | Vaccinium uliginosum | Fujian, China | 26°06′ N, 119°17′ E | MK802167 | N/A | MK816340 | MK809508 |
| L. nanpingensis | CGMCC 3.19597 | Vaccinium uliginosum | Fujian, China | 26°06′ N, 119°17′ E | MK802168 | N/A | MK816341 | MK809509 |
| L. paraphysoides | CGMCC 3.19174 | Vaccinium uliginosum | Shandong, China | 36°03′ N, 120°22′ E | MH729349 | MH729355 | MH729358 | MH729352 |
| L. paraphysoides | CGMCC 3.19175 | Vaccinium uliginosum | Shandong, China | 36°03′ N, 120°22′ E | MH729350 | MH729356 | MH729359 | MH729353 |
| L. parva | CBS 456.78 | Cassava field-soil | Colombia, USA | 34°0′ N, 81°1′ W | EF622083 | EF622063 | KU887523 | KU696372 |
| L. parva | CBS 494.78 | Cassava field-soil | Colombia, USA | 34°0′ N, 81°1′ W | EF622084 | EF622064 | EU673114 | KU696373 |
| L. plurivora | STE-U 4583 | Vitis vinifera | South Africa * | 25°44′ S, 28°15′ E | AY343482 | EF445396 | KU887525 | KU696375 |
| L. plurivora | STE-U 5803 | Prunus salicina | South Africa * | 25°44′ S, 28°15′ E | EF445362 | EF445362 | EF445362 | EF445362 |
| L. pontae | CMM1277 | Spondias purpurea | Brazil * | 15°47′ S, 47°55′ W | KT151794 | KT151791 | KT151797 | N/A |
| L. pseudotheobromae | CBS 116459 | Gmelina arborea | Costa Rica * | 9°55′ N, 84°3′ W | EF622077 | EF622057 | EU673111 | KU696376 |
| L. pseudotheobromae | CGMCC 3.18047 | Pteridium aquilinum | China * | 39°54′ N, 116°23′ E | KX499876 | KX499914 | KX499989 | KX499952 |
| L. pseudotheobromae | CGMCC 3.18451 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783468 | KY848621 | N/A | KY848588 |
| L. pseudotheobromae | CGMCC 3.18452 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783467 | KY848620 | KY848549 | KY848587 |
| L. pseudotheobromae | CGMCC 3.18453 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783460 | KY848618 | KY848545 | KY848580 |
| L. pseudotheobromae | CGMCC 3.18457 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783436 | KY848613 | N/A | N/A |
| L. pseudotheobromae | CGMCC 3.18461 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783446 | N/A | N/A | KY848566 |
| L. pseudotheobromae | CGMCC 3.18465 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783445 | N/A | N/A | KY848565 |
| L. pseudotheobromae | CGMCC 3.18466 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783444 | KY848615 | KY848533 | KY848564 |
| L. pseudotheobromae | CGMCC 3.18470 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783458 | N/A | N/A | KY848578 |
| L. pseudotheobromae | CGMCC 3.18474 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783452 | N/A | KY848538 | KY848572 |
| L. pseudotheobromae | CGMCC 3.18475 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783459 | KY848617 | KY848544 | KY848579 |
| L. pyriformis | CBS 121770 | Acacia mellifera | Namibia * | 22°33′S, 17°04′E | EU101307 | EU101352 | KU887527 | KU696378 |
| L. pyriformis | CBS 121771 | Acacia mellifera | Namibia * | 22°33′ S, 17°04′ E | EU101308 | EU101353 | KU887528 | KU887528 |
| L. rubropurpurea | WAC 12535 | Eucalyptus grandis | Australia * | 32° S, 151° E | DQ103553 | DQ103571 | EU673136 | KU696380 |
| L. rubropurpurea | WAC 12536 | Eucalyptus grandis | Australia * | 32° S, 152° E | DQ103554 | DQ103572 | KU887530 | KU696381 |
| L. sterculiae | CBS342.78 | Sterculia oblonga | Germany * | 52°31′ N, 13°26 E | KX464140 | KX464634 | KX464908 | KX463989 |
| L. subglobosa | CMM3872 | Jatropha curcas | Brazil * | 15°47′S, 47°55′W | KF234558 | KF226721 | KF254942 | N/A |
| L. subglobosa | CMM4046 | Jatropha curcas | Brazil * | 15°47′ S, 47°55′ W | KF234560 | KF226723 | KF254944 | N/A |
| L. tenuiconidia | CGMCC 3.18449 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783466 | KY848619 | N/A | KY848586 |
| L. thailandica | CBS 138653 | Phyllanthus acidus | Thailand * | 13°43′ N, 100°28′ E | KM006433 | KM006464 | N/A | N/A |
| L. thailandica | CBS 138760 | Mangifera indica | Thailand * | 13°43′ N, 100°28′ E | KJ193637 | KJ193681 | N/A | N/A |
| L. theobromae | CBS 111530 | Fruit along coral reef coast | Papua New Guinea * | 9°25′ S, 147°22′ E | EF622074 | EF622054 | KU887531 | KU696382 |
| L. theobromae | CBS 164.96 | Unknown | Unknown | – | AY640255 | AY640258 | KU887532 | KU696383 |
| L. tropica | CGMCC 3.18477 | Aquilaria crassna | Laos | 17°59′ N, 102°34′ E | KY783454 | KY848616 | KY848540 | KY848574 |
| L. venezuelensis | WAC 12539 | Acacia mangium | Venezuela * | 10°28′ N, 66°53′ W | DQ103547 | DQ103568 | KU887533 | KU696384 |
| L. venezuelensis | WAC 12540 | Acacia mangium | Venezuela * | 10°28′ N, 66°53′ W | DQ103548 | DQ103569 | KU887534 | KU887534 |
| L. viticola | UCD 2553AR | Vitis sp. | USA * | 38° N, 97° W | HQ288227 | HQ288269 | HQ288306 | KU696385 |
| L. viticola | UCD 2604MO | Vitis sp. | USA * | 38° N, 97° W | HQ288228 | HQ288270 | HQ288307 | KU696386 |
| L. vitis | CBS 124060 | Vitis vinifera | Italy * | 41°54′ N, 12°18′ E | KX464148 | KX464642 | KX464917 | KX463994 |
| Diplodia mutila | CMW 7060 | Fraxinus excelsior | Netherlands * | 52°22′ N, 4°51′ E | AY236955 | AY236904 | AY236933 | EU339574 |
| D. seriata | CBS 112555 | Vitis vinifera | Portugal | 38°43′ N, 9°7′ W | AY259094 | AY573220 | DQ458856 | N/A |
| Species | Base Pair Difference | Nucleotides Difference (tef1-α) |
|---|---|---|
| L. clavispora and L. gonubiensis | A instead of G | 30 |
| T instead of G | 33 | |
| T instead of gap | 35, 36, 37 | |
| G instead of gap | 38, 42 | |
| C instead of gap | 39, 40, 41 | |
| C instead of T | 44, 48, 105 | |
| G instead of A | 121 | |
| L. fujianensis and L. thailandica | T instead of C | 4 |
| A instead of G | 7 | |
| C instead of A | 27 | |
| gap instead of C | 71, 74, 76 | |
| gap instead of A | 72 | |
| gap instead of G | 73, 75, 78 | |
| gap instead of T | 77 | |
| C instead of T | 92, 153, 296 | |
| C instead of G | 185 | |
| G instead of C | 495 | |
| L. paraphysoides and L. citricola | gap instead of A | 9 |
| T instead of C | 111 | |
| gap instead of G | 197 | |
| A instead of G | 248 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, Y.; Bhoyroo, V.; Rampadarath, S.; Jeewon, R. Multigene Phylogenetics and Morphology Reveal Five Novel Lasiodiplodia Species Associated with Blueberries. Life 2021, 11, 657. https://doi.org/10.3390/life11070657
Wang Y, Zhang Y, Bhoyroo V, Rampadarath S, Jeewon R. Multigene Phylogenetics and Morphology Reveal Five Novel Lasiodiplodia Species Associated with Blueberries. Life. 2021; 11(7):657. https://doi.org/10.3390/life11070657
Chicago/Turabian StyleWang, Yu, Ying Zhang, Vishwakalyan Bhoyroo, Sillma Rampadarath, and Rajesh Jeewon. 2021. "Multigene Phylogenetics and Morphology Reveal Five Novel Lasiodiplodia Species Associated with Blueberries" Life 11, no. 7: 657. https://doi.org/10.3390/life11070657
APA StyleWang, Y., Zhang, Y., Bhoyroo, V., Rampadarath, S., & Jeewon, R. (2021). Multigene Phylogenetics and Morphology Reveal Five Novel Lasiodiplodia Species Associated with Blueberries. Life, 11(7), 657. https://doi.org/10.3390/life11070657