
Lignans from Bursera fagaroides: Chemistry, Pharmacological Effects and Molecular Mechanism. A Current Review

 , ,
, ,
Abstract
:1. Introduction
2. Bursera fagaroides: Description, Distribution, and Uses in the Mexican Traditional Medicine
3. Anti-Cancer Studies of Extracts of B. Fagaroides
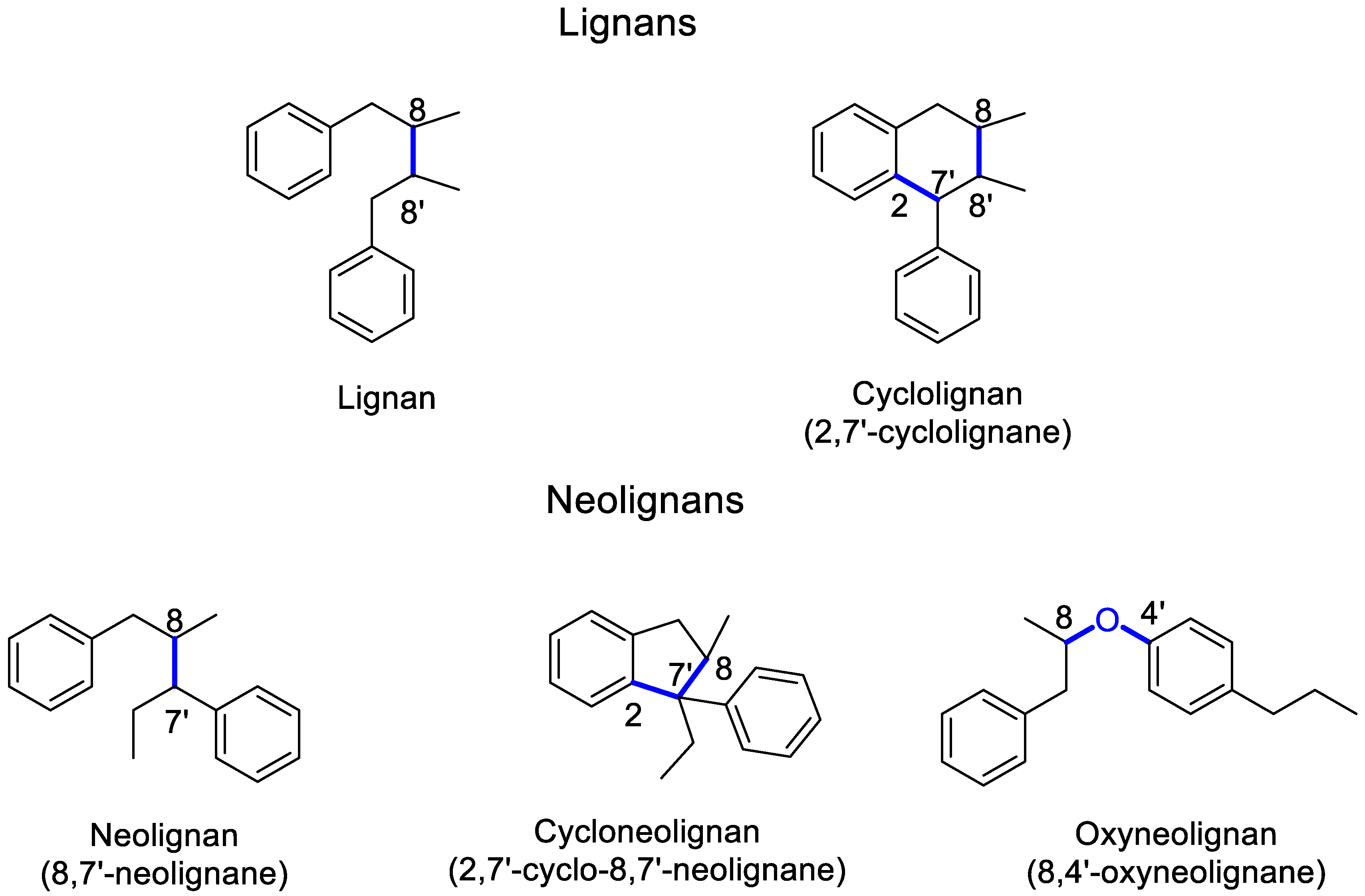
4. Lignans: Definition, Nomenclature, Biosynthesis and Pharmacological Relevance
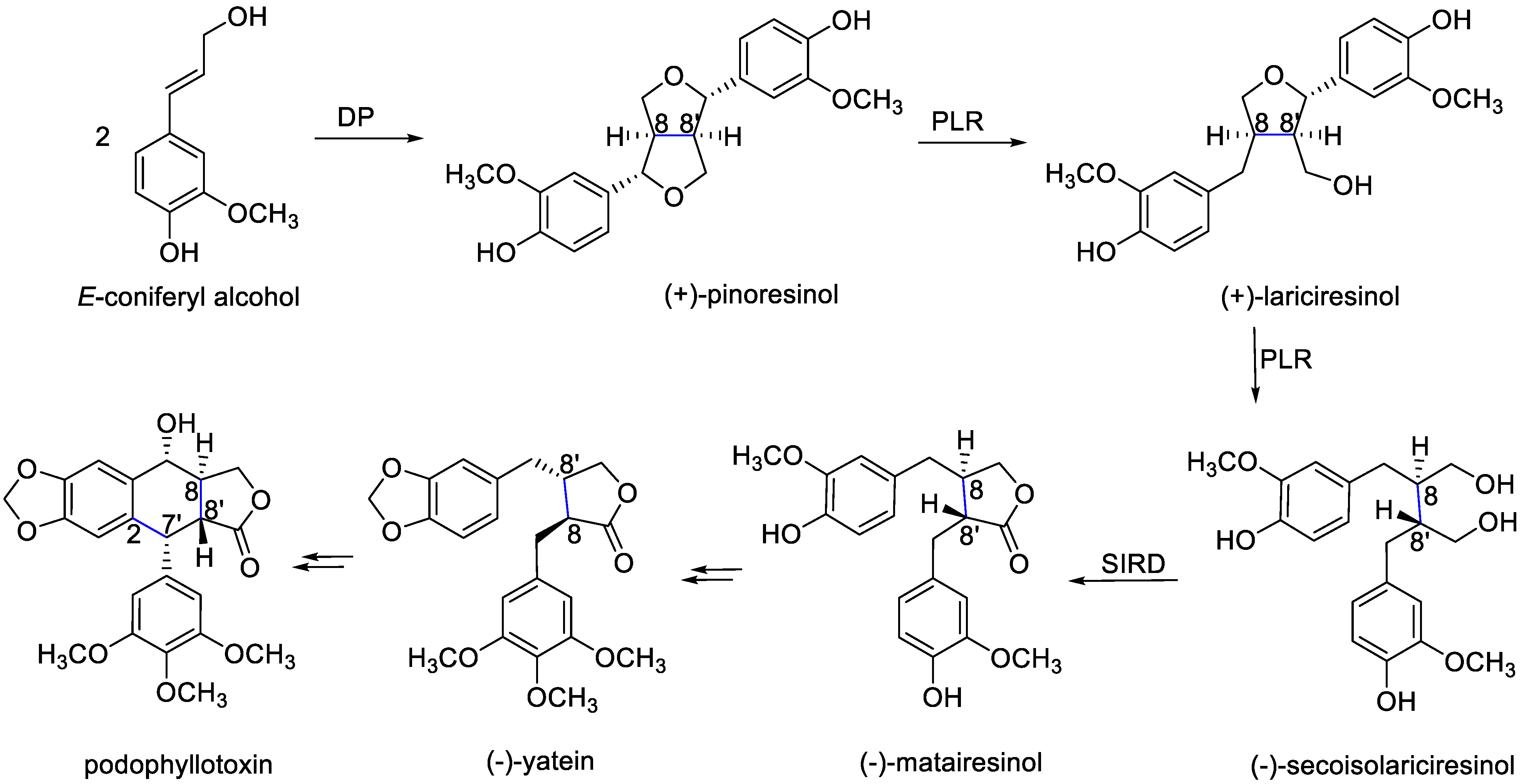
5. Lignans from B. Fagaroides
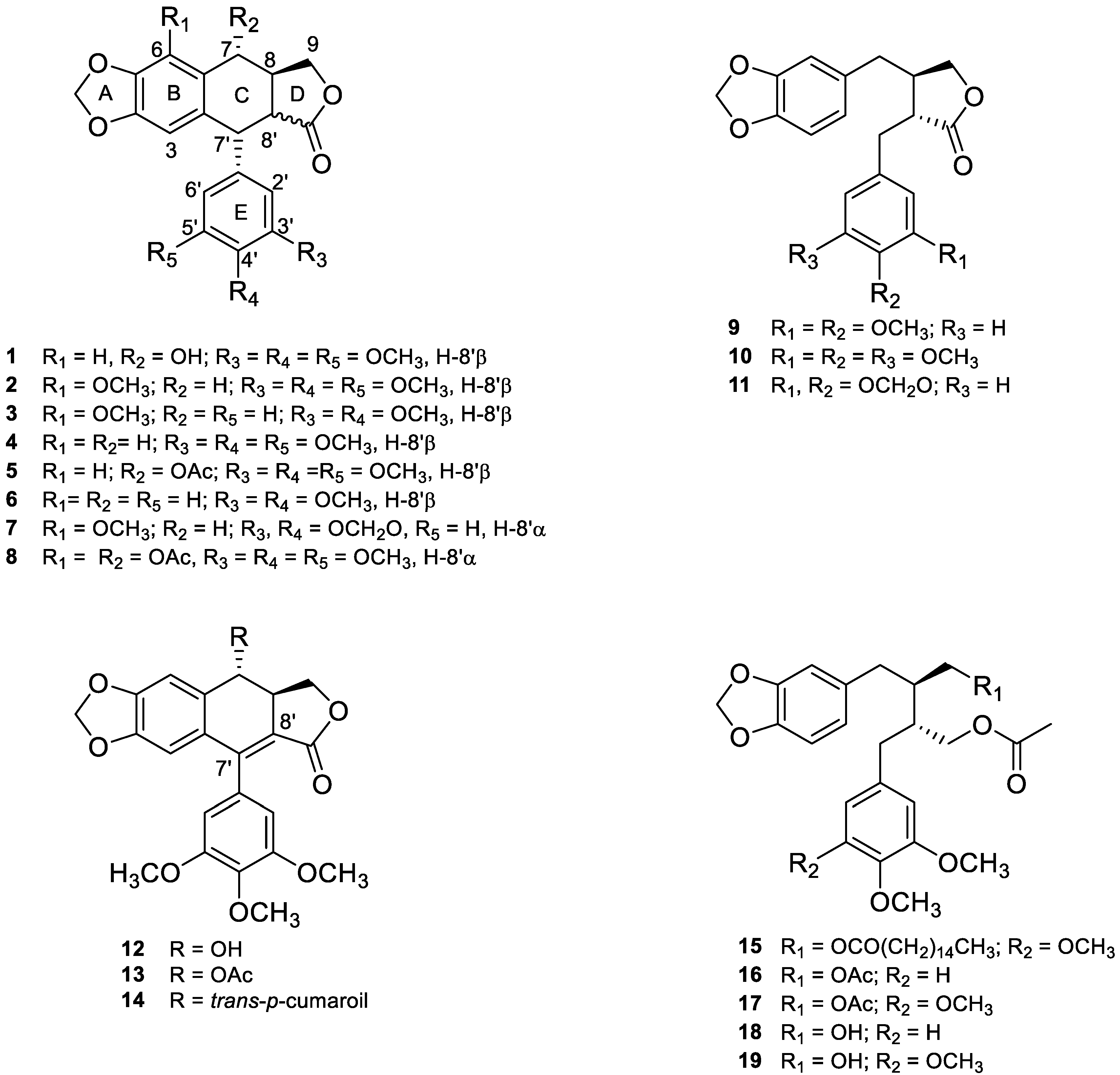
5.1. Chemical Structures
5.2. Isolation and Characterization
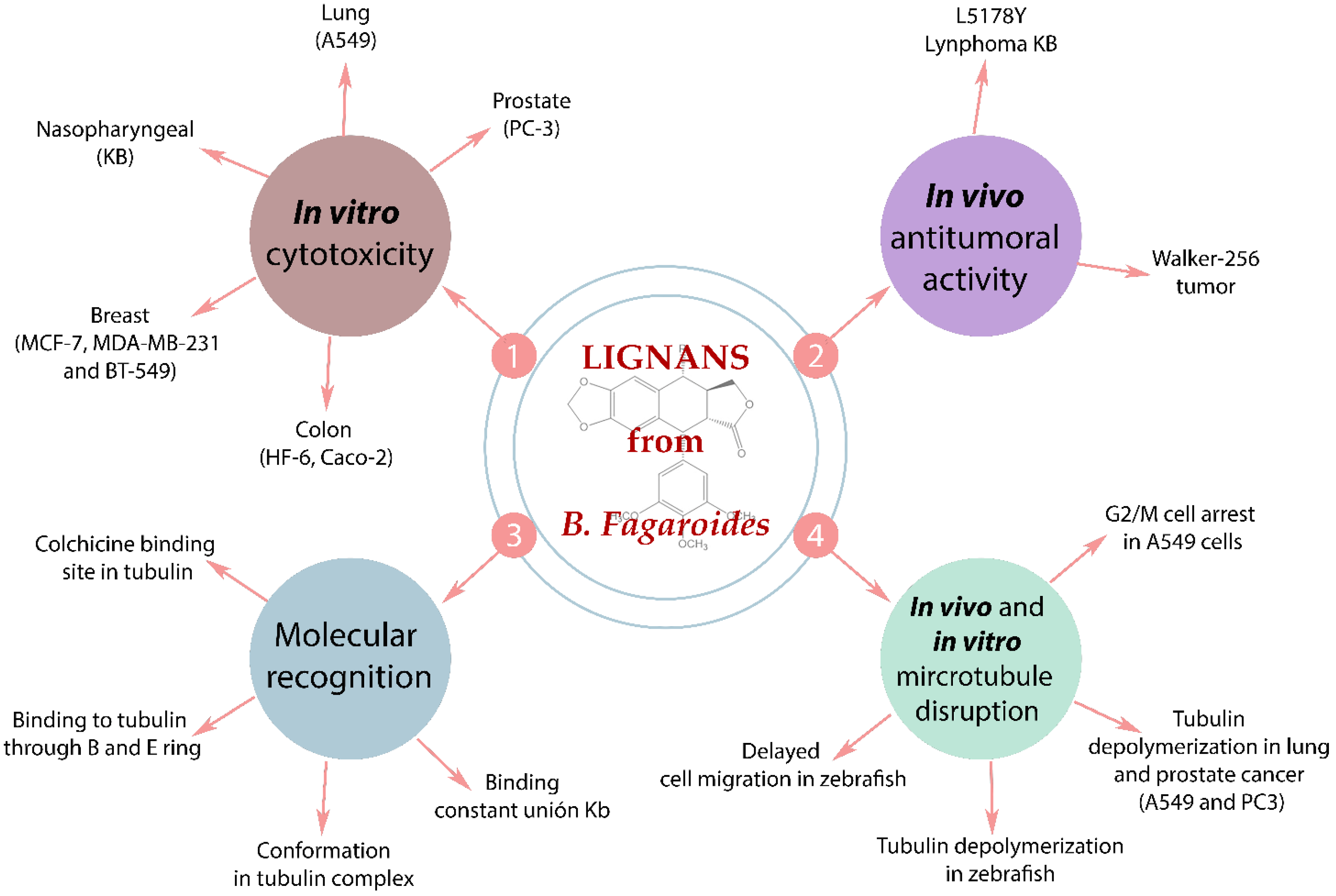
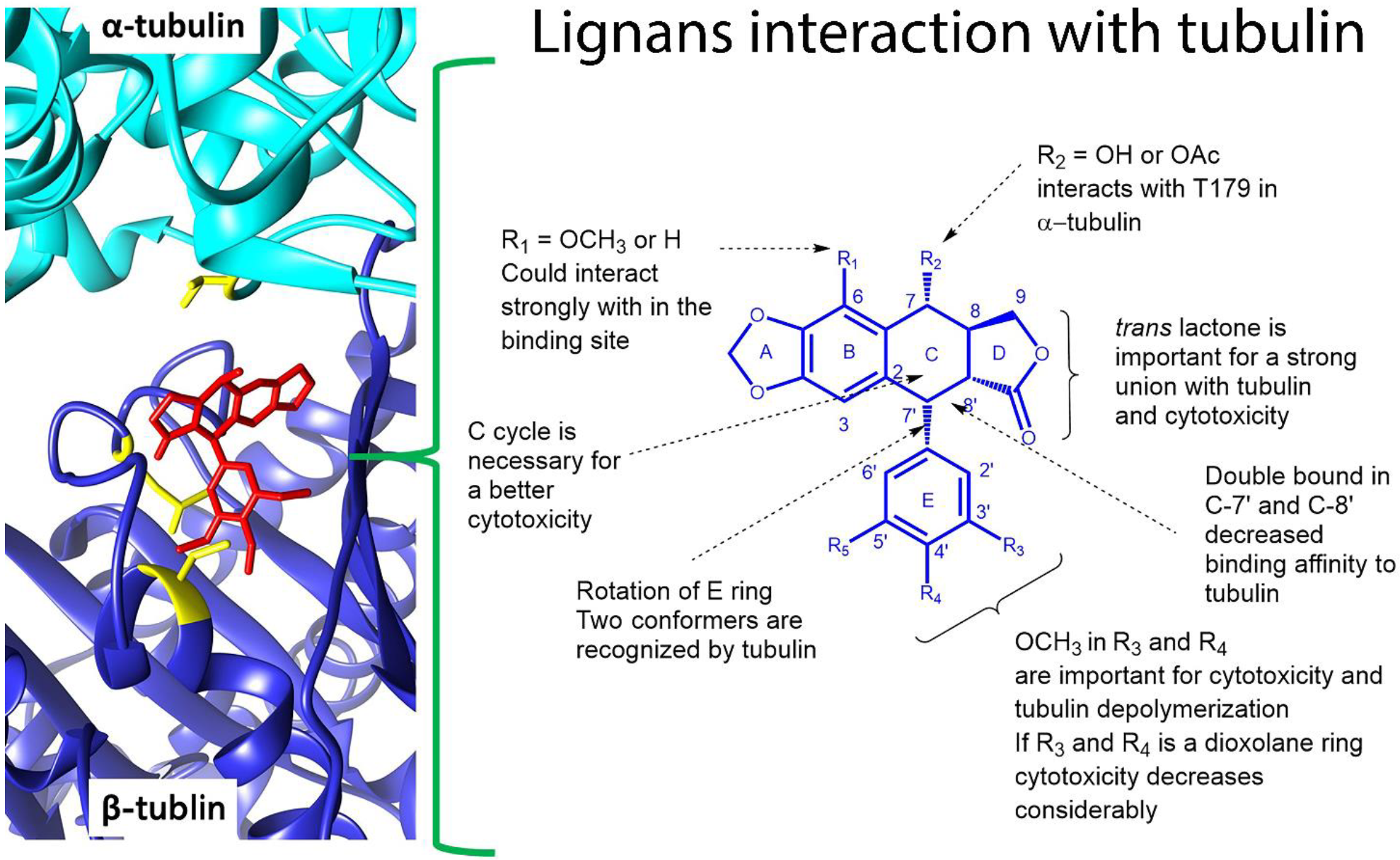
5.3. Anti-Cancer Molecular Mechanism of B. fagaroides Lignans
6. Other Effects
7. Conclusions and Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Mbaveng, A.T.; Manekeng, H.T.; Nguenang, G.S.; Dzotam, J.K.; Kuete, V.; Effertha, E. Cytotoxicity of 18 Cameroonian medicinal plants against drug sensitive and multi-factorial drug resistant cancer cells. J. Ethnopharmacol. 2018, 222, 21–33. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Roodhart, J.M.L.; Daenen, L.G.M.; Stigter, E.C.A.; Prins, H.J.; Gerrits, J.; Houthuijzen, J.M.; Gerritsen, M.G.; Schipper, H.S.; Backer, M.J.G.; van Amersfoort, M.; et al. Mesenchymal Stem Cells Induce Resistance to Chemotherapy through the Release of Platinum-Induced Fatty Acids. Cancer Cell 2011, 20, 370–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atmaca, H.; Çamli, Ç.; Sert, S. Ethanol Extract of Pinus nigra ssp. pallasiana var. şeneriana Inhibits Human Breast Cancer Cell Viability through Induction of Apoptosis. Celal Bayar Univ. J. Sci. 2018, 14, 35–40. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcotullio, M.C.; Curini, M.; Becerra, J.X. An Ethnopharmacological, Phytochemical and Pharmacological Review on Lignans from Mexican bursera spp. Molecules 2018, 23, 1976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becerra, J.X.; Noge, K.; Olivier, S.; Venable, L. The monophyly of Bursera and its impact for divergence times of Burseraceae. Taxon 2012, 61, 333–343. [Google Scholar] [CrossRef]
- Rzedowski, J.; Medina, R.; Calderón, G. Inventario del conocimiento taxonómico, así como de la diversidad y del endemismo regionales de las especies mexicanas de Bursera (Burseraceae). Acta Botánica Mexicana 2005, 70, 85–111. [Google Scholar] [CrossRef]
- Becerra, J.X. Evolution of Mexican Bursera (Burseraceae) inferred from ITS, ETS, and 5S nuclear ribosomal DNA sequences. Mol. Phylogenetics Evol. 2003, 26, 300–309. [Google Scholar] [CrossRef]
- Becerra, J.X.; Venable, D.L.; Evans, P.H.; Bowers, W.S. Interactions between Chemical and Mechanical Defenses in the Plant Genus Bursera and Their Implications for Herbivores. Am. Zool. 2001, 41, 865–876. [Google Scholar]
- Case, R.J.; Tucker, A.O.; Maciarello, M.J.; Wheeler, K.A. Chemistry and Ethnobotany of Commercial Incense Copals, Copal Blanco, Copal Oro, and Copal Negro of North America. Econ. Bot. 2003, 57, 189–202. [Google Scholar] [CrossRef]
- Linares, E.; Bye, R. El copal en México. Biodiversitas 2008, 78, 8–11. [Google Scholar]
- Nakanishi, T.; Inatomi, Y.; Satomi, A.; Yamada, T.; Fukatsu, H.; Murata, H.; Inada, A.; Matsuura, N.; Ubukata, M.; Murata, J.; et al. New luteolin 3-O-acylated rhamnosides from leaves of Bursera graveolens. Heterocycles 2003, 60, 2077–2083. [Google Scholar]
- Souza, M.P.; Machado, M.I.L.; Braz-Filho, R. Six flavonoids from Bursera leptophloeos. Phytochemistry 1989, 28, 2467–2470. [Google Scholar] [CrossRef]
- Romero-Estrada, A.; Maldonado-Magaña, A.; González-Christen, J.; Marquina, S.; Garduño-Ramírez, M.L.; Rodríguez-López, V.; Alvarez, L. Anti-inflammatory and antioxidative effects of six pentacyclic triterpenes isolated from the Mexican copal resin of Bursera copallifera. BMC Complement. Altern. Med. 2016, 16, 422–431. [Google Scholar] [CrossRef] [Green Version]
- Columba-Palomares, M.C.; Villarreal, M.L.; Marquina, S.; Romero-Estrada, A.; Rodríguez-López, V.; Zamilpa, A.; Alvarez, L. Antiproliferative and Anti-inflammatory Acyl Glucosyl Flavones from the Leaves of Bursera copallifera. J. Mex. Chem. Soc. 2018, 62, 214–224. [Google Scholar] [CrossRef]
- Noge, K.; Becerra, J.X. Germacrene D, A Common Sesquiterpene in the Genus Bursera (Burseraceae). Molecules 2009, 14, 5289–5297. [Google Scholar] [CrossRef] [PubMed]
- García-Gutiérrez, H.A.; Cerda-García-Rojas, C.M.; Hernández-Hernández, J.D.; Román-Marín, L.U.; Joseph-Nathan, P. Oxygenated verticillene derivatives from Bursera suntui. Phytochemistry 2008, 69, 2844–2848. [Google Scholar] [CrossRef]
- Rojas-Sepulveda, A.M.; Mendieta-Serrano, M.; Mojica, M.Y.; Salas-Vidal, E.; Marquina, S.; Villarreal, M.L.; Puebla, A.M.; Delgado, J.I.; Alvarez, L. Cytotoxic Podophyllotoxin Type-Lignans from the Steam Bark of Bursera fagaroides var. fagaroides. Molecules 2012, 17, 9506–9519. [Google Scholar] [CrossRef] [Green Version]
- Antunez-Mojica, M.; León, A.; Rojas-Sepúlveda, A.M.; Marquina, S.; Mendieta-Serrano, M.; Salas-Vidal, E.; Villareal, M.L.; Alvarez, L. Aryldihydronaphthalene-type lignans from Bursera fagaroides var. Fagaroides and their antimitotic mechanism of action. RSC Adv. 2016, 6, 4950–4959. [Google Scholar] [CrossRef]
- Purata, S.E. (Ed.) Uso y Manejo de los Copales Aromáticos: Resinas y Aceites; CONABIO/RAISES: Mexico, 2008; pp. 1–60. Available online: https://bioteca.biodiversidad.gob.mx/janium-bin/detalle.pl?Id=20210712091900 (accessed on 9 June 2021).
- Puebla-Pérez, A.M.; Huacuja-Ruiz, L.; Rodríguez-Orozco, G.; Villaseñor-García, M.M.; Miranda-Beltrán, M.; Celis, A.; Sandoval-Ramírez, L. Cytotoxic and Antitumour Activity from Bursera fagaroides Ethanol Extract in Mice with L5178Y Lymphoma. Phytother. Res. 1998, 12, 545–548. [Google Scholar] [CrossRef]
- Rzedowski, J. Las especies de Bursera (Burseraceae) en la cuenca superior del río Papaloapan (México). Acta Botánica Mexicana 2004, 66, 23–151. [Google Scholar] [CrossRef] [Green Version]
- Castañeda-Miranda, A.G.; Chaparro, M.A.E.; Pacheco-Castro, A.; Chaparro, M.A.E.; Böhnel, H.N. Magnetic biomonitoring of atmospheric dust using tree leaves of Ficus benjamina in Querétaro (México). Environ. Monit. Assess. 2020, 192, 291–382. [Google Scholar] [CrossRef] [PubMed]
- Morales-Serna, J.A.; Cruz-Galicia, E.; García-Ríos, E.; Madrigal, D.; Gaviño, R.; Cárdenas, J.; Salmón, M. Three new diarylbutane lignans from the resin of Bursera fagaroides. Nat. Prod. Res. 2013, 27, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, E.; Sheth, K.; Cole, J.R. Antitumor agents from Bursera fagaroides (Burseraceae) (β-peltatin-A-methylether and 5′-desmethoxy-β-peltatin-A-methylether). Tetrahedron Lett. 1969, 32, 2759–2762. [Google Scholar] [CrossRef]
- Velázquez-Jiménez, R.; Torres-Valencia, J.M.; Cerda-García-Rojas, C.M.; Hernández-Hernández, J.D.; Román-Marín, L.U.; Manríquez-Torres, J.J.; Gómez-Hurtado, M.A.; Valdez-Calderón, A.; Motilva, V.; García-Mauriño, S.; et al. Absolute configuration of podophyllotoxin related lignans from Bursera fagaroides using vibrational circular dichroism. Phytochemistry 2011, 72, 2237–2243. [Google Scholar] [CrossRef]
- Acevedo, M.; Nuñez, P.; Gónzalez-Maya, L.; Cardoso-Taketa, A.; Villarreal, M.L. Cytotoxic and Anti-inflammatory Activities of Bursera species from Mexico. J. Clin. Toxicol. 2015, 5, 2–8. [Google Scholar]
- Antúnez-Mojica, M.; Rojas-Sepúlveda, A.M.; Mendieta-Serrano, M.A.; Leticia Gonzalez-Maya, L.; Marquina, S.; Salas-Vidal, E.; Alvarez, L. Lignans from Bursera fagaroides Affect in vivo Cell Behavior by Disturbing the Tubulin Cytoskeleton in Zebrafish Embryos. Molecules 2019, 24, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, G.P. Nomenclature of Lignans and Neolignans (IUPAC Recommendations). Pure Appl. Chem. 2000, 72, 1493–1523. [Google Scholar] [CrossRef]
- Talapatra, S.K.; Talapatra, B. Shikimic Acid Pathway. In Chemistry of Plant Natural Products: Stereochemistry, Conformation, Synthesis, Biology and Medicine; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Davin, L.B.; Wang, H.W.; Crowell, A.L.; Bedgar, D.L.; Martin, D.M.; Sarkanen, S.; Lewis, N.G. Stereoselective bimolecular phenoxy radical coupling by an auxiliary (dirigent) protein without an active center. Science 1997, 275, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Corbin, C.; Drouet, S.; Markulin, L.; Auguin, D.; Lainé, É.; Davin, L.B.; Cort, J.R.; Lewis, N.G.; Hano, C. A genome-wide analysis of the flax (Linum usitatissimum L.) dirigent protein family: From gene identification and evolution to differential regulation. Plant Mol. Biol. 2018, 97, 73–101. [Google Scholar] [CrossRef]
- Umezawa, T. Diversity in lignan biosynthesis. Phytochem. Rev. 2003, 2, 371–390. [Google Scholar] [CrossRef]
- Hirano, T.; Fukuoka, K.; Oka, K.; Naito, T.; Hosaka, K.; Mitsuhashi, H.; Matsumoto, Y. Antiproliferative activity of mammalian lignan derivatives against the human breast carcinoma cell line, ZR-75-1. Cancer Investig. 1990, 8, 595–602. [Google Scholar] [CrossRef]
- Kardono, L.B.; Tsauri, S.; Padmawinata, K.; Pezzuto, J.M.; Kinghorn, A.D. Cytotoxic constituents of the bark of Plumeria rubra collected in Indonesia. J. Nat. Prod. 1990, 53, 1447–1455. [Google Scholar] [CrossRef]
- Drugbank. Podophyllotoxin. Available online: https://go.drugbank.com/drugs/DB01179 (accessed on 28 March 2021).
- Gutierrez-Gutierrez, F.; Puebla-Perez, A.M.; Gonzalez-Pozos, S.; Hernandez-Hernandez, J.M.; Perez-Rangel, A.; Alvarez, L.P.; Tapia-Pastrana, G.; Castillo-Romero, A. Antigiardial Activity of Podophyllotoxin-Type Lignans from Bursera fagaroides var. fagaroides. Molecules 2017, 22, 799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peña-Morán, O.; Villareal, M.L.; Álvarez, L.; Meneses-Acosta, A.; Rodríguez-López, V. Cytotoxicity, Post-Treatment Recovery, and Selectivity Analysis of Naturally Occurring Podophyllotoxins from Bursera fagaroides var. fagaroides on Breast Cancer Cell Lines. Molecules 2016, 21, 1013. [Google Scholar] [CrossRef] [PubMed]
- Antunez-Mojica, M.; Rodríguez-Salarichs, J.; Redondo-Horcajo, M.; León, A.; Barasoain, I.; Canales, A.; Cañada, F.J.; Jiménez-Barbero, J.; Alvarez, L.; Díaz, J.F. Structural and Biochemical Characterization of the Interaction of Tubulin with Potent Natural Analogues of Podophyllotoxin. J. Nat. Prod. 2016, 79, 2113–2121. [Google Scholar] [CrossRef]
- Gutiérrez-Gutiérrez, F.; Romo-Mancillas, A.; Puebla-Pérez, A.; Hernández-Hernández, J.M.; Castillo-Romero, A. Identification and molecular characterization of the tubulin-podophyllotoxin-type lignans binding site on Giardia lamblia. Chem. Biol. Drug. Des. 2019, 94, 2031–2040. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Arreguín, P.; Arteaga-Nieto, P.; Reynoso-Orozco, R.; Villagómez-Castro, J.C.; Sabanero-López, M.; Puebla-Pérez, A.M.; Calvo-Méndez, C. Bursera fagaroides, effect of an ethanolic extract on ornithine decarboxylase (ODC) activity in vitro and on the growth of Entamoeba histolytica. Exp. Parasitol. 2008, 119, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Zálešák, F.; Jean-Yves, D.; Bon, D.; Pospíšil, J. Lignans and Neolignans: Plant secondary metabolites as a reservoir of biologically active substances. Pharmacol. Res. 2019, 146, 1–27. [Google Scholar] [CrossRef]
- Schrecker, A.W.; Hartwell, J.L. Components of podophyllin XX. The absolute configuration of podophyllotoxin and related lignans. J. Org. Chem. 1956, 21, 381–382. [Google Scholar] [CrossRef]
- Petcher, T.J.; Weber, H.P.; Kuhn, M.; Von Wartburg, A. Crystal structure and absolute configuration of 2′-bromopodophyllotoxin—0.5 ethyl acetate. J. Chem. Soc. Perkin Trans. 1973, 2, 288–292. [Google Scholar] [CrossRef]
- Arora, S.K.; Bates, R.B.; Grady, R.A. Crystal and molecular structure of beta-peltatin a methyl ether. J. Org. Chem. 1975, 40, 28–31. [Google Scholar] [CrossRef]
- Bates, R.B.; Wood, J.B., III. Crystal and molecular structure of 5′-demothxy-.beta.-peltatin A methyl ether. J. Org. Chem. 1972, 37, 562–565. [Google Scholar] [CrossRef]
- Harmatha, J.; Buděšínský, M.; Trka, A. The structure of yatein. Determination of the positions, and configurations of benzyl groups in lignans of the 2,3-dibenzylbutyrolactone type. Collect. Czech. Chem. Commun. 1982, 47, 644–663. [Google Scholar] [CrossRef]
- Brewer, C.F.; Loike, J.D.; Horwitz, S.B.; Sternlicht, H.; Gensler, W.J. Conformational analysis of podophyllotoxin and its congeners. Structure–activity relationship in microtubule assembly. J. Med. Chem. 1979, 22, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Desai, D.C.; Jacob, J.; Almeida, A.; Kshirsagar, R.; Manju, S.L. Isolation, structural elucidation and antiinflammatory activity of astragalin, (−)hinokinin, aristolactam I and aristolochic acids (I & II) from Aristolochia indica. Nat. Prod. Res. 2014, 28, 1413–1417. [Google Scholar] [PubMed]
- Hendrawati, O.; Woerdenbag, H.J.; Michiels, P.J.A.; Aantjes, H.G.; Dam, A.v.; Kayser, O. Identification of lignans and related compounds in Anthriscus sylvestris by LC-ESI-MS/MS and LC-SPE-NMR. Phytochemistry 2011, 72, 2172–2179. [Google Scholar] [CrossRef]
- Sakar, M.K.; Er, N.; Dilek, E.; Del Olmo, E.; San Feliciano, A. (−)-Desoxypodophyllotoxin and diterpenoids from uniperus nana Willd. berries. Acta Pharm. Turcica 2002, 44, 213–219. [Google Scholar]
- Da Silva, R.; Heleno, V.C.G.; de Albuquerque, S.; Bastos, J.K.; e Silva, M.L.A.; Donate, P.M.; da Silva, G.V.J. Complete assignment of 1H and 13C NMR data for three aryltetralin lignan lactones. Magn. Reson. Chem. 2004, 42, 985–989. [Google Scholar] [CrossRef]
- Nakanishi, T.; Inatomi, Y.; Murata, H.; Shigeta, K.; Iida, N.; Inada, A.; Murata, J.; Perez Farrera, M.A.; Iinuma, M.; Tanaka, T.; et al. A new and known cytotoxic aryltetralin-type lignans from stems of Bursera graveolens. Chem. Pharm. Bull. 2005, 53, 229–231. [Google Scholar] [CrossRef] [Green Version]
- Jolad, S.D.; Wiedhopf, R.M.; Cole, J.R. Cytotoxic agents from Bursera morelensis (Burseraceae): Deoxypodophyllotoxin and a new lignan, 5′-desmethoxydeoxypodophyllotoxin. J. Pharm. Sci. 1977, 66, 892–893. [Google Scholar] [CrossRef] [PubMed]
- Jutiviboonsuk, A.; Zhang, H.; Tan, G.T.; Ma, C.; Hung, N.V.; Cuong, N.M.; Bunyapraphatsara, N.; Soejarto, D.D.; Fong, H.H.S. Bioactive constituents from roots of Bursera tonkinensis. Phytochemistry 2005, 66, 2745–2751. [Google Scholar] [CrossRef] [PubMed]
- Cancer Facts & Figures 2020. 2020. Available online: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2020.html (accessed on 26 April 2021).
- Gordaliza, M.; Castro, M.D.; Miguel del Corral, J.M.; Feliciano, A.S. Feliciano, Antitumor properties of podophyllotoxin and related compounds. Curr. Pharm. Des. 2000, 6, 1811–1839. [Google Scholar] [CrossRef] [PubMed]
- Mounina, G.; Zhou, J.Z.; Lu-Yong, Z. Podophyllotoxin, a medicinal agent of plant origin: Past, present and future. Chin. J. Nat. Med. 2012, 10, 161–169. [Google Scholar]
- Ardalani, H.; Avan, A.; Ghayour-Mobarhan, M. Podophyllotoxin: A novel potential natural anti-cancer agent. Avicenna J. Phytomed. 2017, 7, 285–294. [Google Scholar]
- Longstaff, E.; Von Krogh, G. Condyloma eradication: Self-therapy with 0.15–0.5% podophyllotoxin versus 20–25% podophyllin preparations—An integrated safety assessment. Regul. Toxicol. Pharmacol. 2001, 33, 117–137. [Google Scholar] [CrossRef]
- Xu, X.Y.; Wang, D.Y.; Li, Y.P.; Deyrup, S.T.; Zhang, H.J. Plant-derived lignans as potential antiviral agents: A systematic review. Phytochem. Rev. 2021. [Google Scholar] [CrossRef]
- Nieto-Yañez, O.J.; Resendiz-Albor, A.A.; Ruiz-Hurtado, P.A.; Rivera-Yañez, N.; Rodriguez-Canales, M.; Rodriguez-Sosa, M.; Juarez-Avelar, I.; Rodriguez-Lopez, M.G.; Canales-Martinez, M.M.; Rodriguez-Monroy, M.A. In vivo and in vitro antileishmanial effects of methanolic extract from bark of Bursera aptera. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.Y.; Seo, K.H.; Jeong, R.H.; Lee, S.M.; Kim, G.S.; Noh, H.J.; Kim, S.Y.; Kim, G.W.; Kim, J.Y.; Baek, N.I. Anti-inflammatory lignans from the fruits of Acanthopanax sessiliflorus. Molecules 2013, 18, 41–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, T.S.C.; Lucarini, R.; Volpe, A.C.; de Andrade, C.Q.J.; Souza, A.M.P.; Pauletti, P.M.; Januário, A.H.; Símaro, G.V.; Bastos, J.K.; Cunha, W.R.; et al. In vivo and in silico anti-inflammatory mechanism of action of the semisynthetic ()-cubebin derivatives ()-hinokinin and ()-O-benzylcubebin. Bioorg. Med. Chem. Lett. 2017, 27, 176–179. [Google Scholar] [CrossRef] [Green Version]
- De Souza Pereira, J.J.; Pereira, A.D.P.; Jandú, J.J.; da Paz, J.A.; Crovella, S.; dos Santos Correia, M.T.; de Azevêdo Silva, J. Commiphora leptophloeos phytochemical and antimicrobial characterization. Front. Microbiol. 2017, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.L.A.; Coimbra, H.S.; Pereira, A.C.; Almeida, V.A.; Lima, T.C.; Costa, E.S.; Vinholis, A.H.C.; Royo, V.A.; Silva, R.; Filho, A.A.S.; et al. Evaluation of Piper cubeba extract, ()-cubebin and its semi-synthetic derivatives against oral pathogens. Phytoth. Res. 2007, 21, 420–422. [Google Scholar] [CrossRef]
- Huang, R.L.; Huang, Y.L.; Ou, J.C.; Chen, C.C.; Hsu, F.L.; Chang, C. Screening of 25 compounds isolated from Phyllanthus species for anti-human Hepatitis B virus in vitro. Phytother. Res. 2003, 17, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.S.; Koo, K.A.; Ma, C.J.; Sung, S.H.; Kim, Y.C. Neuroprotective lignans from Biota orientalis leaves. Nat. Prod. Sci. 2008, 14, 167–170. [Google Scholar]
- Haribabu, K.; Ajitha, M.; Mallavadhani, U.V. Quantitative estimation of (-)-hinokinin, a trypanosomicidal marker in Piper cubeba, and some of its commercial formulations using HPLC-PDA. J. Pharm. Anal. 2015, 5, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Esperandim, V.R.; da Silva Ferreira, D.; Rezende, K.C.; Cunha, W.R.; Saraiva, J.; Bastos, J.K.; e Silva, M.L.; de Albuquerque, S. Evaluation of the in vivo therapeutic properties of (-)-cubebin and (-)-hinokinin against Trypanosoma cruzi. Experim. Parasitol. 2013, 133, 442–446. [Google Scholar] [CrossRef]
- Usia, T.; Watabe, T.; Kadota, S.; Tezuka, Y. Metabolite-cytochrome P450 complex formation by methylenedioxyphenyl lignans of Piper cubeba: Mechanism-based inhibition. Life Sci. 2005, 76, 2381–2391. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Chang, Y.L.; Teng, C.M.; Chen, I.S. Anti-platelet aggregation alkaloids and lignans from Hernandia nymphaeifolia. Planta Med. 2000, 66, 251–256. [Google Scholar] [CrossRef]
- Picking, D.; Chambers, B.; Barker, J.; Shah, I.; Porter, R.; Naughton, D.P.; Delgoda, R. Inhibition of cytochrome P450 activities by extracts of Hyptis verticillata Jacq.: Assessment for potential HERB-drug interactions. Molecules 2018, 23, 430. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | KB | PC-3 | MCF-7 | HF-6 | A549 | A2780 | MDA-MB-231 | BT-549 | Ref |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 4.61 × 10−6 | 2.29 | 2.51 × 10−5 | 4.34 × 10−4 | 0.015 | 0.018 | NR | NR | [19,40] |
| 2 | 4.43 × 10−6 | 2.21 | 2.24 × 10−5 | 4.2 × 10−4 | NR | NR | NR | NR | [19] |
| 3 | 2.19 × 10−5 | 2.51 × 10−5 | 2.56 × 10−4 and 7.22 | 1.00 | 0.033 | 0.084 | 2.44 | 1.26 | [19,39,40] |
| 4 | 3.76 | 5.02 × 10−3 | 3.76 | 3.09 | NR | NR | NR | NR | [19] |
| 5 | 2.25 | 0.0109 | >8 and 0.132 | 5.28 | 0.025 | 0.034 | 0.18 | 0.011 | [19,39,40] |
| 6 | NR | NR | 0.040 | NR | NR | NR | 0.14 | 0.021 | [39] |
| 7 | 7.54 | 5.22 × 10−3 | 9.60 | 7.54 | 8.67 | 12.94 | NR | NR | [19,40] |
| 9 | 1.07 | 4.58 × 10−3 | 1.07 | 1.83 | NR | NR | NR | NR | [19] |
| 12 | 0.25 | 2.42 × 10−5 | >9.7 | 0.012 | NR | NR | NR | NR | [20] |
| 13 | 0.29 | 0.06 | >8.8 and 0.35 | 0.066 | NR | NR | 0.16 | 0.06 | [20,40] |
| 14 | 3.61 | 2.42 × 10−5 | >7.2 | 1.27 | NR | NR | NR | NR | [20] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antúnez-Mojica, M.; Romero-Estrada, A.; Hurtado-Díaz, I.; Miranda-Molina, A.; Alvarez, L. Lignans from Bursera fagaroides: Chemistry, Pharmacological Effects and Molecular Mechanism. A Current Review. Life 2021, 11, 685. https://doi.org/10.3390/life11070685
Antúnez-Mojica M, Romero-Estrada A, Hurtado-Díaz I, Miranda-Molina A, Alvarez L. Lignans from Bursera fagaroides: Chemistry, Pharmacological Effects and Molecular Mechanism. A Current Review. Life. 2021; 11(7):685. https://doi.org/10.3390/life11070685
Chicago/Turabian StyleAntúnez-Mojica, Mayra, Antonio Romero-Estrada, Israel Hurtado-Díaz, Alfonso Miranda-Molina, and Laura Alvarez. 2021. "Lignans from Bursera fagaroides: Chemistry, Pharmacological Effects and Molecular Mechanism. A Current Review" Life 11, no. 7: 685. https://doi.org/10.3390/life11070685
APA StyleAntúnez-Mojica, M., Romero-Estrada, A., Hurtado-Díaz, I., Miranda-Molina, A., & Alvarez, L. (2021). Lignans from Bursera fagaroides: Chemistry, Pharmacological Effects and Molecular Mechanism. A Current Review. Life, 11(7), 685. https://doi.org/10.3390/life11070685